Embed Size (px)
Citation preview

Differential effect of dietary spermine on alkaline phosphatase activityin jejunum and ileum of unweaned rats
Olivier Peulen 1, Myriam Gharbi 1, Brigitte Powroznik, Guy Dandrifosse*
Department of Biochemistry and General Physiology, Institute of Chemistry B6C, Immunology Centre,University of Liège, B-4000 Liège (Sart-Tilman), Belgium
Received 30 March 2004; accepted 11 June 2004
Available online 06 July 2004
Abstract
Spermine is a low molecular weight polyamine involved in the postnatal maturation of the gut. When it is administered orally to sucklingrats it induces the maturation of their spleen, liver, pancreas, and small intestine. We showed that this polyamine modulates differently theactivity of alkaline phosphatase in jejunum and ileum in suckling rat. In 14-day-old rat which had received spermine orally for 3 days, oncedaily, an increase of alkaline phosphatase activity in the jejunum and a decrease of this activity in the ileum was observed. Alkalinephosphatase was located at the bottom of the villus in the control jejunum and in the whole length of the villus in spermine-treated rats. On thecontrary, in ileum of controls, this enzyme was present in the whole length of the villus but disappeared in the spermine-treated animals. Anenzyme mass shift was observed in the small intestine after spermine administration. Spermine administration did not change the expressionof genes coding for alkaline phosphatase, suggesting a post-transcriptional modification.© 2004 Elsevier SAS. All rights reserved.
Keywords: Alkaline phosphatase; Zymography; Intestine; Weaning; Maturation; Differentiation
1. Introduction
In mammals, at weaning, the gastrointestinal tract has toadapt to a drastic change from milk to solid food. The natureand activity of the enzymes located in the brush borders ofthe enterocytes were modified after solid food ingestion: forexample, lactase specific activity decreases, in contrast tomaltase and sucrase activities, which increase [1]. In parallelto this digestive adaptation, immunological changes occur toadapt the animals to the new microbial and nutritional anti-genic content of the small intestine [2].
The factors triggering postnatal maturation of the smallintestine are still poorly understood. Exogenous administra-tion of cortisone [1], thyroxine [3], insulin [4] and epidermalgrowth factor (EGF) [5] have been shown to induce preco-cious physiological and morphological maturation of theintestine in the rats. Both insulin [6] and EGF [7] increaseornithine decarboxylase (ODC) activity in the small intestine
and lead to an increase of polyamine synthesis. Difluorom-ethylornithine, an irreversible inhibitor of ODC activity, pre-vents the normal increase of polyamine concentration in theintestinal mucosa at weaning and delays postnatal intestinalmaturation [8]. Moreover, this maturation is also delayed inornithine transcarbamylase deficient mice [9]. These resultssuggest a relation between polyamines and maturation.
It appears that spermine, a low molecular weightpolyamine, induces the maturation of small intestine mucosawhen orally ingested by suckling rats [10]. Among others,the spermine-induced maturation is characterised by a de-crease in lactase specific activity and by an increase in mal-tase and sucrase specific activities. It is identical to thenatural one which occurs at weaning [11] and is dependenton the immune system [12,13].
In small intestine, alkaline phosphatase (IAP) is coded bytwo different genes (IAP-1 and IAP-2). After weaning,mRNA (2.7 kb) coding for IAP increases in jejunum whenIAP activity decreases twofold [14] suggesting a post-transcriptional regulation. Reducing western-blot shows a65 kDa IAP band in jejunum before and after weaning [14].
* Corresponding author. Tel.: +32-4-366-3577; fax: +32-4-366-2887.E-mail address: [email protected] (G. Dandrifosse).1 The first two authors contributed equally to this research.
Biochimie 86 (2004) 487–493
www.elsevier.com/locate/biochi
0300-9084/$ - see front matter © 2004 Elsevier SAS. All rights reserved.doi:10.1016/j.biochi.2004.06.002

In this report we analysed the effects of spermine ingestionby suckling rats on IAP activity and gene expression.
2. Materials and methods
2.1. Animals
Wistar rats (Rattus norvegicus) were obtained from theanimal house of the Medicine Faculty of the Liege Univer-sity. They were housed in an environmentally controlledroom at 23 °C with a 12-h light–dark cycle. They were fedwith standard laboratory diet (Carfill, Bruxelles, Belgium)and had water ad libitum. Day of birth was reported as day 0.In our experiments, there was no body weight differencebetween male or female pups. No distinction between genderwas made. Since it is well known that experimental valuesvary from one litter to another, the comparisons of resultswere always made between animals from the same litter. Theanimal welfare committees of the University of Liege and theFRSM (Fonds de la Recherche Scientifique Médicale) ap-proved the animal experiments.
2.2. Experimental design
Spermine (0.4 µmol/g body weight in 50 µl water) wasadministered orally as reported elsewhere [10]. Control ani-mals were treated in the same way, but received only thevehicle (50 µl of water). Spermine was given for 3 days (day11, 12 and 13) once daily at 9:00 h. After spermine adminis-tration, the suckling rats were immediately returned to theirmother. They were killed on day 14 by cervical dislocation.
2.3. Small intestine preparation
The small intestine was immediately removed after sacri-fice, then washed in cold saline (NaCl 9 g/l, 4 °C). Theduodenum was removed from the intestine until the Treitzligament. The small intestine was divided into two pieces ofequal length called jejunum and ileum. Each part was ho-mogenised in distilled water (1 g wet weight per 4.15 ml)using a homogeniser (ultra-turrax, Ika T8). The homogenatewas kept in portions maintained at –70 °C until analysis.
Alternatively, 1 cm-length piece of jejunum and ileum wasimmediately snap-frozen in Tissue Tek medium (Sakura,Netherlands). Serial 6 µm thick sections were cut with aMicron HM 500 OM (Microm) cryostat, mounted on poly-L-lysine-coated slides, air-dried for 15 min, and fixed in coldacetone for 10 min.
2.4. Disaccharidase activity analysis
Sucrase (EC 3.2.1.48), maltase (EC 3.2.1.20) and lactase(EC 3.2.1.23) activities were assayed according to Dahlqvist[15]. They were expressed as µmol substrate hydrolysed permin. They were related to 1 g of protein (specific activ-
ity = SA). Protein content was estimated according to Brad-ford [16] using bovine serum albumin as protein standard.
2.5. Alkaline phosphatase activity analysis
Alkaline phosphatase (EC 3.1.3.1) activity was assayedon homogenates according to Millington and Tovell [17]using paranitrophenolphosphate as substrate. It was ex-pressed as µmol substrate hydrolysed per min and related to1 g of protein (specific activity = SA). Protein content wasestimated according to Bradford [16] using bovine serumalbumin as protein standard.
2.6. In situ alkaline phosphatase analysis
Aceton-fixed cryosections were rehydrated in phosphatebuffered solution (PBS). Alkaline phosphatase localisationwas analysed by histochemistry using ready-to-use tabletscontaining naphtol substrate for alkaline phosphatase andFast Red chromogen (Roche Diagnostics GmbH, Germany).Tablets were dissolved in 2 ml of Tris–HCl (0.1 M, pH 8.2).The solution was directly applied on the section.
2.7. Alkaline phosphatase zymography
Proteins for zymography were isolated from homogenatesby lysis solution [Tris–HCl (50 mM, pH 7.4), NaCl(150 mM), Igepal (1%, v/v), sodium deoxycholate (1%,w/v), iodoacetamide (5 mM), Triton X-100 (1%, v/v), PMSF(2 mM)] and subsequent clarification at 22 000 × g for40 min.
Proteins were separated by unreducing SDS-PAGE (8% or12%, w/v). Before loading, 30 µg proteins were mixed in 1:1with loading buffer [Tris–HCl (65 mM, pH 6.8), glycerol(26%, v/v), SDS (2%, w/v), bromophenol blue (1%, w/v)]and submitted to separation at 20 V/cm without previous heatdenaturation. After separation, the gel was incubated at roomtemperature in a substrate solution [Sigma Fast™BCIP/NBT tablets (5-bromo-4-chloro-3-indolyl phos-phate/nitroblue tetrazolium)] until the appearance of alkalinephosphatase bands on the gel (at least overnight). Banddensitometry was estimated using Quantity One software(Bio-Rad).
2.8. Alkaline phosphatase RT-PCR
Total RNA was extracted from jejunum and ileum byRNAgent kit (Promega). Single-stranded cDNA was synthe-sised from 1 µg total RNA with avian myeloblastosis virus(AMV) reverse transcriptase (15 U), dNTP (1 mM), recom-binant ribonuclease inhibitor (1 U/µl), oligo(dT)15 primers(0.025 µg/µl)] in 20 µl AMV buffer [Tris–HCl (10 mM, pH9), KCl (50 mM), Triton X-100 (0.1%, v/v), MgCl2 (5 mM)].Reverse transcription was performed at 42 °C for 15 min,then AMV was inactivated at 95 °C for 5 min. This handlingwas followed by a 4 °C step for 5 min to avoid AMV–cDNAcomplex formation.
488 O. Peulen et al. / Biochimie 86 (2004) 487–493
PCR were performed from 3 µl of first-strand cDNAreaction using an Amplitron II system (Thermolyne) with1 U Taq polymerase (Promega), dNTP (200 µM), MgCl2(1.5 mM) in 100 µl PCR buffer [Tris–HCl (10 mM, pH 9),KCl (50 mM), Triton X-100 (0.1%, v/v)]. Oligonucleotidesdescribed in Table 1 were used as PCR primers. They weredesigned using Primer3 web-based software [18]. Primerswere picked up in adjacent exons. PCR consisted of a firstdenaturation step at 95 °C for 5 min, followed by 20–40 am-plification cycles (95 °C for 1 min, 58 °C for 1 min, 72 °C for1 min). A terminal elongation was performed for 5 min at72 °C. Samples were taken after 20, 30 and 40 PCR cyclesand submitted to horizontal agarose gel (1%, w/v) electro-phoresis at 80 V in order to get quantification (band densito-metry using Quantity One analysis software, Bio-Rad) in thelinear amplification zone.
Abundance of tissue non-specific alkaline phosphatasemRNA was determined by cDNA array hybridisation (AtlasRat 1.2 array, Clontech) following manufacturer recommen-dations.
2.9. Statistical analysis
The results are reported as means with their standarddeviation (S.D.). Statistical analysis was performed usingStudent’s t-test for testing treatment effect. When heterosce-dasticity (Levene’s test) was suspected, non-parametric testwas used.
3. Results
The intestinal disaccharidase SA were analysed afterspermine treatment as a positive control of spermineeffect. Spermine ingested once a day for 3 days (postnataldays 11, 12 and 13), induced a modification in the disac-charidase SA level in small intestine. Lactase SA wassignificantly reduced (137.93 ± 19.54 vs.86.28 ± 24.77 µmol/min g protein; P < 0.05, N = 4),whereas maltase and sucrase SA were significantly increased(88.45 ± 20.14 vs. 316.35 ± 61.02 µmol/min g protein;P < 0.01, N = 4 and 3.20 ± 0.74 vs. 59.91 ± 14.11 µmol/min gprotein; P < 0.01, N = 4), indicating a successful maturationof the small intestine.
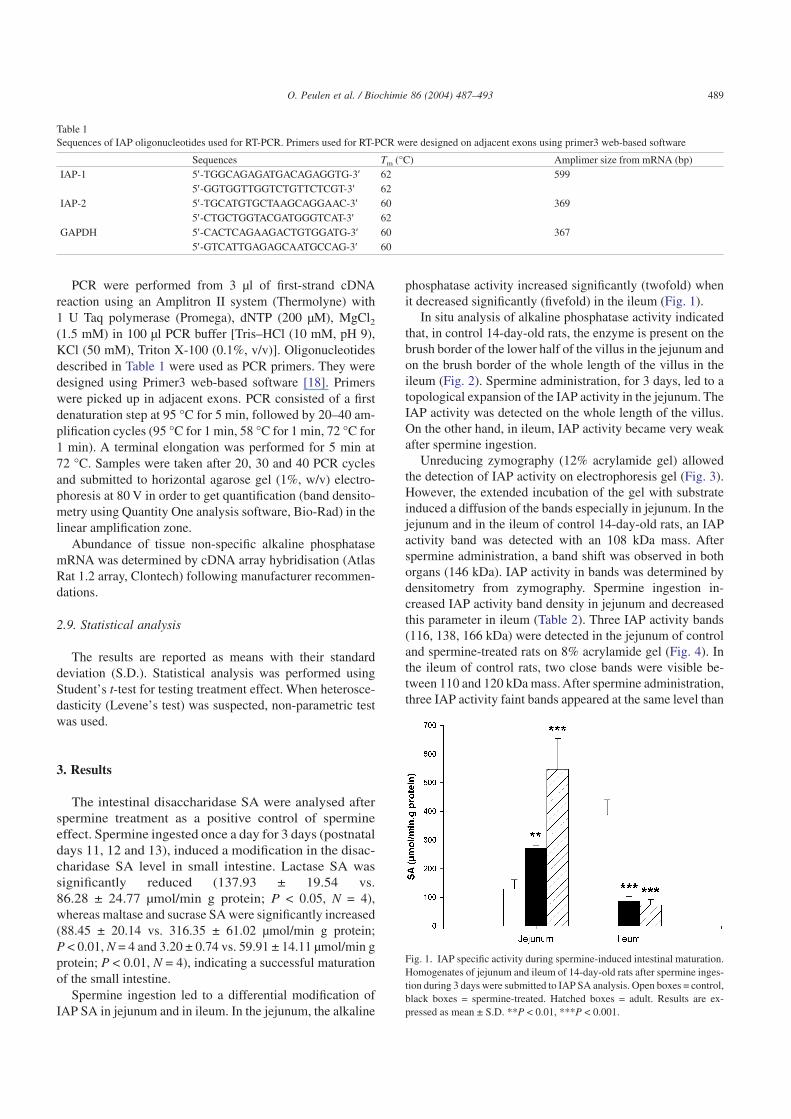
Spermine ingestion led to a differential modification ofIAP SA in jejunum and in ileum. In the jejunum, the alkaline
phosphatase activity increased significantly (twofold) whenit decreased significantly (fivefold) in the ileum (Fig. 1).
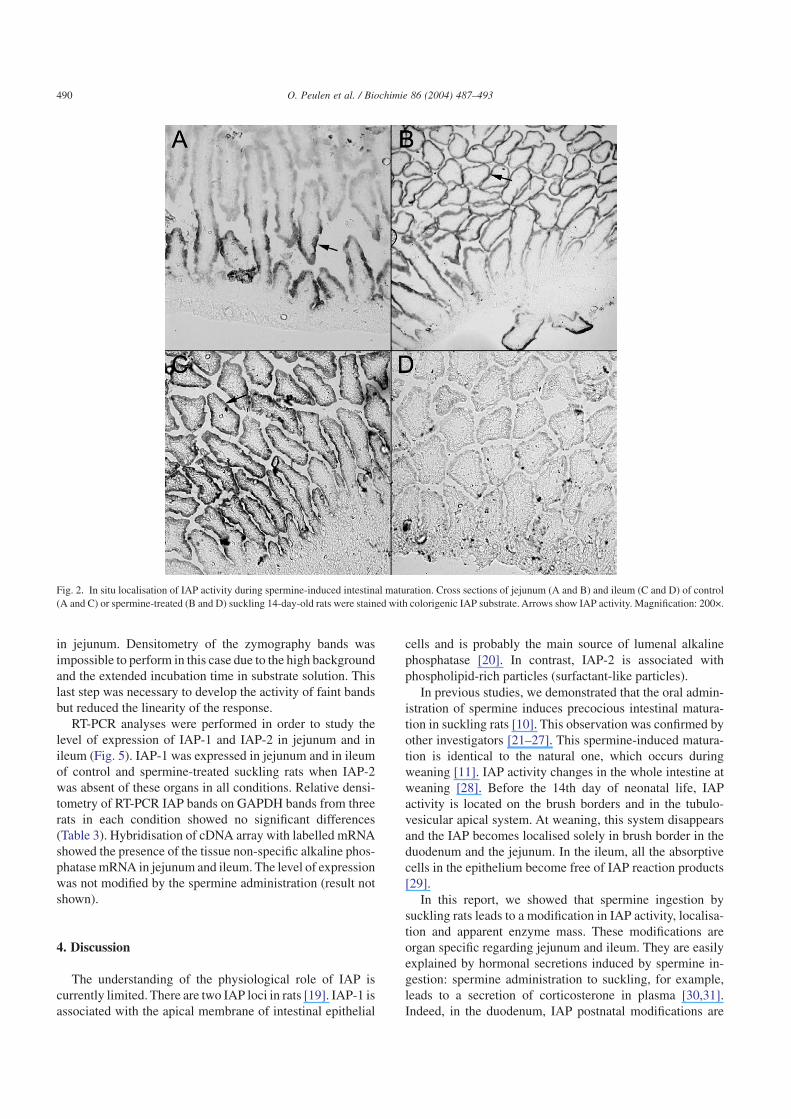
In situ analysis of alkaline phosphatase activity indicatedthat, in control 14-day-old rats, the enzyme is present on thebrush border of the lower half of the villus in the jejunum andon the brush border of the whole length of the villus in theileum (Fig. 2). Spermine administration, for 3 days, led to atopological expansion of the IAP activity in the jejunum. TheIAP activity was detected on the whole length of the villus.On the other hand, in ileum, IAP activity became very weakafter spermine ingestion.
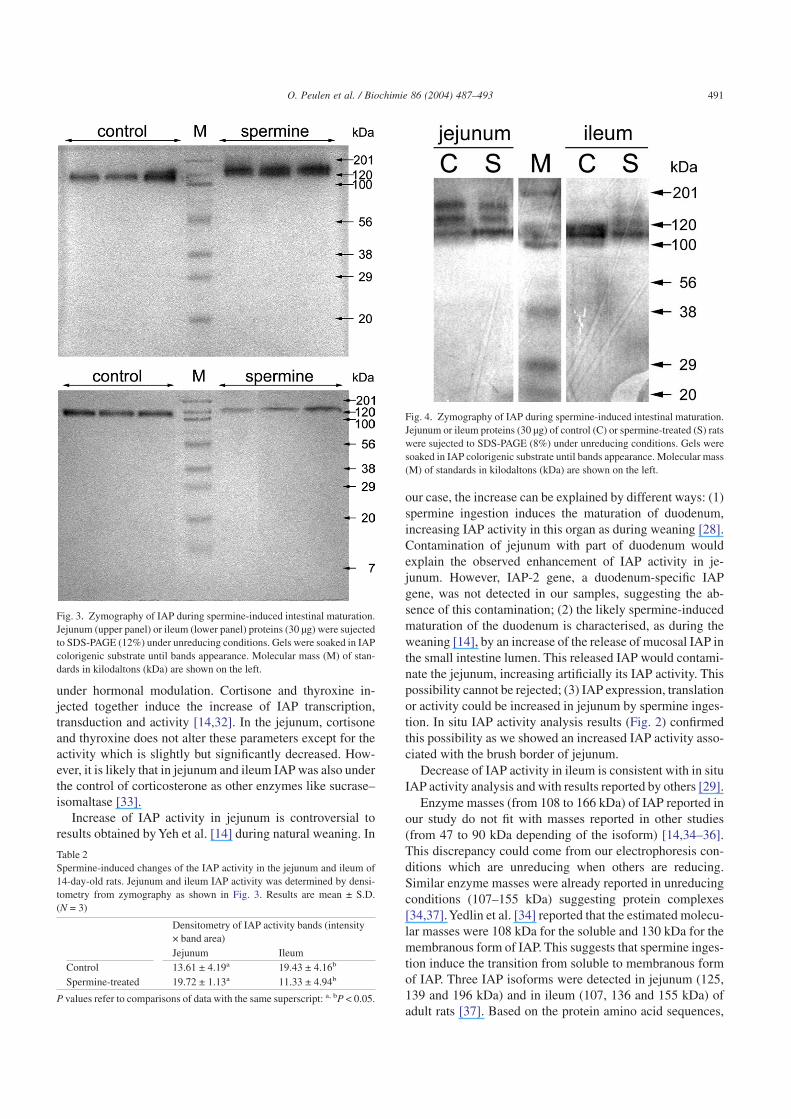
Unreducing zymography (12% acrylamide gel) allowedthe detection of IAP activity on electrophoresis gel (Fig. 3).However, the extended incubation of the gel with substrateinduced a diffusion of the bands especially in jejunum. In thejejunum and in the ileum of control 14-day-old rats, an IAPactivity band was detected with an 108 kDa mass. Afterspermine administration, a band shift was observed in bothorgans (146 kDa). IAP activity in bands was determined bydensitometry from zymography. Spermine ingestion in-creased IAP activity band density in jejunum and decreasedthis parameter in ileum (Table 2). Three IAP activity bands(116, 138, 166 kDa) were detected in the jejunum of controland spermine-treated rats on 8% acrylamide gel (Fig. 4). Inthe ileum of control rats, two close bands were visible be-tween 110 and 120 kDa mass. After spermine administration,three IAP activity faint bands appeared at the same level than
Table 1Sequences of IAP oligonucleotides used for RT-PCR. Primers used for RT-PCR were designed on adjacent exons using primer3 web-based software
Sequences Tm (°C) Amplimer size from mRNA (bp)IAP-1 5′-TGGCAGAGATGACAGAGGTG-3′ 62 599
5′-GGTGGTTGGTCTGTTCTCGT-3′ 62IAP-2 5′-TGCATGTGCTAAGCAGGAAC-3′ 60 369
5′-CTGCTGGTACGATGGGTCAT-3′ 62GAPDH 5′-CACTCAGAAGACTGTGGATG-3′ 60 367
5′-GTCATTGAGAGCAATGCCAG-3′ 60
Fig. 1. IAP specific activity during spermine-induced intestinal maturation.Homogenates of jejunum and ileum of 14-day-old rats after spermine inges-tion during 3 days were submitted to IAP SA analysis. Open boxes = control,black boxes = spermine-treated. Hatched boxes = adult. Results are ex-pressed as mean ± S.D. **P < 0.01, ***P < 0.001.
489O. Peulen et al. / Biochimie 86 (2004) 487–493
in jejunum. Densitometry of the zymography bands wasimpossible to perform in this case due to the high backgroundand the extended incubation time in substrate solution. Thislast step was necessary to develop the activity of faint bandsbut reduced the linearity of the response.
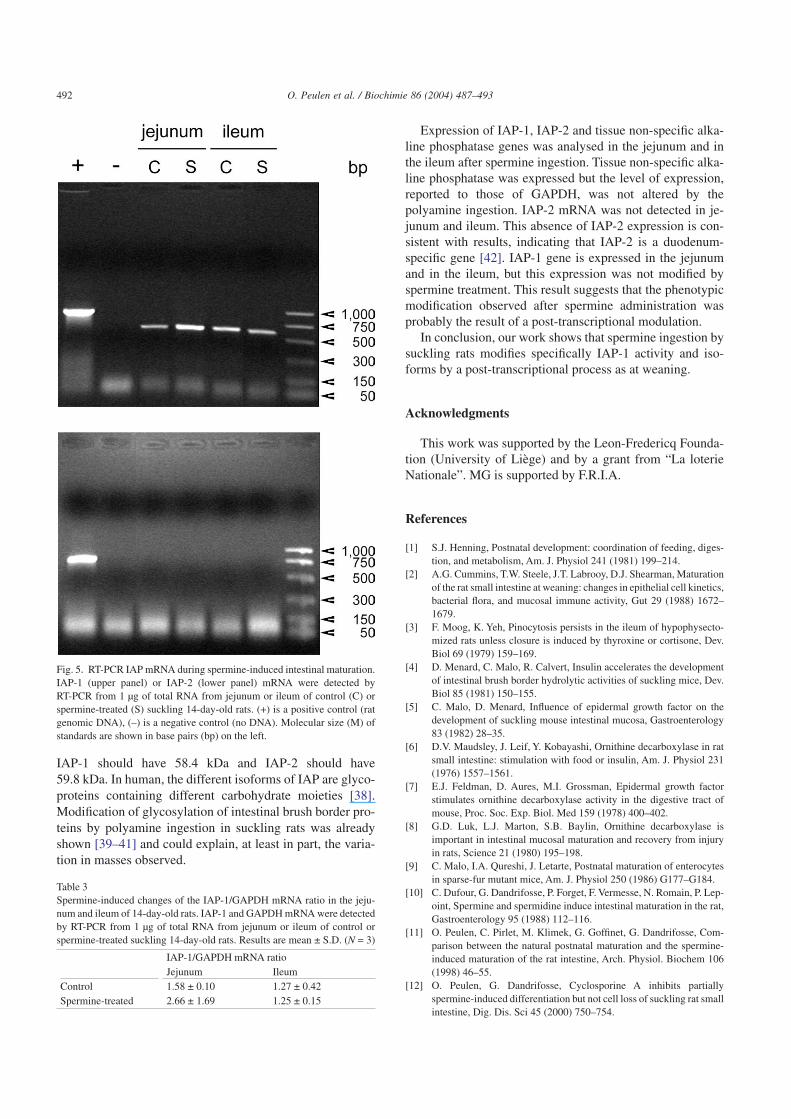
RT-PCR analyses were performed in order to study thelevel of expression of IAP-1 and IAP-2 in jejunum and inileum (Fig. 5). IAP-1 was expressed in jejunum and in ileumof control and spermine-treated suckling rats when IAP-2was absent of these organs in all conditions. Relative densi-tometry of RT-PCR IAP bands on GAPDH bands from threerats in each condition showed no significant differences(Table 3). Hybridisation of cDNA array with labelled mRNAshowed the presence of the tissue non-specific alkaline phos-phatase mRNA in jejunum and ileum. The level of expressionwas not modified by the spermine administration (result notshown).
4. Discussion
The understanding of the physiological role of IAP iscurrently limited. There are two IAP loci in rats [19]. IAP-1 isassociated with the apical membrane of intestinal epithelial
cells and is probably the main source of lumenal alkalinephosphatase [20]. In contrast, IAP-2 is associated withphospholipid-rich particles (surfactant-like particles).
In previous studies, we demonstrated that the oral admin-istration of spermine induces precocious intestinal matura-tion in suckling rats [10]. This observation was confirmed byother investigators [21–27]. This spermine-induced matura-tion is identical to the natural one, which occurs duringweaning [11]. IAP activity changes in the whole intestine atweaning [28]. Before the 14th day of neonatal life, IAPactivity is located on the brush borders and in the tubulo-vesicular apical system. At weaning, this system disappearsand the IAP becomes localised solely in brush border in theduodenum and the jejunum. In the ileum, all the absorptivecells in the epithelium become free of IAP reaction products[29].
In this report, we showed that spermine ingestion bysuckling rats leads to a modification in IAP activity, localisa-tion and apparent enzyme mass. These modifications areorgan specific regarding jejunum and ileum. They are easilyexplained by hormonal secretions induced by spermine in-gestion: spermine administration to suckling, for example,leads to a secretion of corticosterone in plasma [30,31].Indeed, in the duodenum, IAP postnatal modifications are
Fig. 2. In situ localisation of IAP activity during spermine-induced intestinal maturation. Cross sections of jejunum (A and B) and ileum (C and D) of control(A and C) or spermine-treated (B and D) suckling 14-day-old rats were stained with colorigenic IAP substrate. Arrows show IAP activity. Magnification: 200×.
490 O. Peulen et al. / Biochimie 86 (2004) 487–493
under hormonal modulation. Cortisone and thyroxine in-jected together induce the increase of IAP transcription,transduction and activity [14,32]. In the jejunum, cortisoneand thyroxine does not alter these parameters except for theactivity which is slightly but significantly decreased. How-ever, it is likely that in jejunum and ileum IAP was also underthe control of corticosterone as other enzymes like sucrase–isomaltase [33].
Increase of IAP activity in jejunum is controversial toresults obtained by Yeh et al. [14] during natural weaning. In
our case, the increase can be explained by different ways: (1)spermine ingestion induces the maturation of duodenum,increasing IAP activity in this organ as during weaning [28].Contamination of jejunum with part of duodenum wouldexplain the observed enhancement of IAP activity in je-junum. However, IAP-2 gene, a duodenum-specific IAPgene, was not detected in our samples, suggesting the ab-sence of this contamination; (2) the likely spermine-inducedmaturation of the duodenum is characterised, as during theweaning [14], by an increase of the release of mucosal IAP inthe small intestine lumen. This released IAP would contami-nate the jejunum, increasing artificially its IAP activity. Thispossibility cannot be rejected; (3) IAP expression, translationor activity could be increased in jejunum by spermine inges-tion. In situ IAP activity analysis results (Fig. 2) confirmedthis possibility as we showed an increased IAP activity asso-ciated with the brush border of jejunum.
Decrease of IAP activity in ileum is consistent with in situIAP activity analysis and with results reported by others [29].
Enzyme masses (from 108 to 166 kDa) of IAP reported inour study do not fit with masses reported in other studies(from 47 to 90 kDa depending of the isoform) [14,34–36].This discrepancy could come from our electrophoresis con-ditions which are unreducing when others are reducing.Similar enzyme masses were already reported in unreducingconditions (107–155 kDa) suggesting protein complexes[34,37].Yedlin et al. [34] reported that the estimated molecu-lar masses were 108 kDa for the soluble and 130 kDa for themembranous form of IAP. This suggests that spermine inges-tion induce the transition from soluble to membranous formof IAP. Three IAP isoforms were detected in jejunum (125,139 and 196 kDa) and in ileum (107, 136 and 155 kDa) ofadult rats [37]. Based on the protein amino acid sequences,
Fig. 3. Zymography of IAP during spermine-induced intestinal maturation.Jejunum (upper panel) or ileum (lower panel) proteins (30 µg) were sujectedto SDS-PAGE (12%) under unreducing conditions. Gels were soaked in IAPcolorigenic substrate until bands appearance. Molecular mass (M) of stan-dards in kilodaltons (kDa) are shown on the left.
Table 2Spermine-induced changes of the IAP activity in the jejunum and ileum of14-day-old rats. Jejunum and ileum IAP activity was determined by densi-tometry from zymography as shown in Fig. 3. Results are mean ± S.D.(N = 3)
Densitometry of IAP activity bands (intensity× band area)Jejunum Ileum
Control 13.61 ± 4.19a 19.43 ± 4.16b
Spermine-treated 19.72 ± 1.13a 11.33 ± 4.94b
P values refer to comparisons of data with the same superscript: a, bP < 0.05.
Fig. 4. Zymography of IAP during spermine-induced intestinal maturation.Jejunum or ileum proteins (30 µg) of control (C) or spermine-treated (S) ratswere sujected to SDS-PAGE (8%) under unreducing conditions. Gels weresoaked in IAP colorigenic substrate until bands appearance. Molecular mass(M) of standards in kilodaltons (kDa) are shown on the left.
491O. Peulen et al. / Biochimie 86 (2004) 487–493
IAP-1 should have 58.4 kDa and IAP-2 should have59.8 kDa. In human, the different isoforms of IAP are glyco-proteins containing different carbohydrate moieties [38].Modification of glycosylation of intestinal brush border pro-teins by polyamine ingestion in suckling rats was alreadyshown [39–41] and could explain, at least in part, the varia-tion in masses observed.
Expression of IAP-1, IAP-2 and tissue non-specific alka-line phosphatase genes was analysed in the jejunum and inthe ileum after spermine ingestion. Tissue non-specific alka-line phosphatase was expressed but the level of expression,reported to those of GAPDH, was not altered by thepolyamine ingestion. IAP-2 mRNA was not detected in je-junum and ileum. This absence of IAP-2 expression is con-sistent with results, indicating that IAP-2 is a duodenum-specific gene [42]. IAP-1 gene is expressed in the jejunumand in the ileum, but this expression was not modified byspermine treatment. This result suggests that the phenotypicmodification observed after spermine administration wasprobably the result of a post-transcriptional modulation.
In conclusion, our work shows that spermine ingestion bysuckling rats modifies specifically IAP-1 activity and iso-forms by a post-transcriptional process as at weaning.
Acknowledgments
This work was supported by the Leon-Fredericq Founda-tion (University of Liège) and by a grant from “La loterieNationale”. MG is supported by F.R.I.A.
References
[1] S.J. Henning, Postnatal development: coordination of feeding, diges-tion, and metabolism, Am. J. Physiol 241 (1981) 199–214.
[2] A.G. Cummins, T.W. Steele, J.T. Labrooy, D.J. Shearman, Maturationof the rat small intestine at weaning: changes in epithelial cell kinetics,bacterial flora, and mucosal immune activity, Gut 29 (1988) 1672–1679.
[3] F. Moog, K. Yeh, Pinocytosis persists in the ileum of hypophysecto-mized rats unless closure is induced by thyroxine or cortisone, Dev.Biol 69 (1979) 159–169.
[4] D. Menard, C. Malo, R. Calvert, Insulin accelerates the developmentof intestinal brush border hydrolytic activities of suckling mice, Dev.Biol 85 (1981) 150–155.
[5] C. Malo, D. Menard, Influence of epidermal growth factor on thedevelopment of suckling mouse intestinal mucosa, Gastroenterology83 (1982) 28–35.
[6] D.V. Maudsley, J. Leif, Y. Kobayashi, Ornithine decarboxylase in ratsmall intestine: stimulation with food or insulin, Am. J. Physiol 231(1976) 1557–1561.
[7] E.J. Feldman, D. Aures, M.I. Grossman, Epidermal growth factorstimulates ornithine decarboxylase activity in the digestive tract ofmouse, Proc. Soc. Exp. Biol. Med 159 (1978) 400–402.
[8] G.D. Luk, L.J. Marton, S.B. Baylin, Ornithine decarboxylase isimportant in intestinal mucosal maturation and recovery from injuryin rats, Science 21 (1980) 195–198.
[9] C. Malo, I.A. Qureshi, J. Letarte, Postnatal maturation of enterocytesin sparse-fur mutant mice, Am. J. Physiol 250 (1986) G177–G184.
[10] C. Dufour, G. Dandrifosse, P. Forget, F. Vermesse, N. Romain, P. Lep-oint, Spermine and spermidine induce intestinal maturation in the rat,Gastroenterology 95 (1988) 112–116.
[11] O. Peulen, C. Pirlet, M. Klimek, G. Goffinet, G. Dandrifosse, Com-parison between the natural postnatal maturation and the spermine-induced maturation of the rat intestine, Arch. Physiol. Biochem 106(1998) 46–55.
[12] O. Peulen, G. Dandrifosse, Cyclosporine A inhibits partiallyspermine-induced differentiation but not cell loss of suckling rat smallintestine, Dig. Dis. Sci 45 (2000) 750–754.
Fig. 5. RT-PCR IAP mRNA during spermine-induced intestinal maturation.IAP-1 (upper panel) or IAP-2 (lower panel) mRNA were detected byRT-PCR from 1 µg of total RNA from jejunum or ileum of control (C) orspermine-treated (S) suckling 14-day-old rats. (+) is a positive control (ratgenomic DNA), (–) is a negative control (no DNA). Molecular size (M) ofstandards are shown in base pairs (bp) on the left.
Table 3Spermine-induced changes of the IAP-1/GAPDH mRNA ratio in the jeju-num and ileum of 14-day-old rats. IAP-1 and GAPDH mRNA were detectedby RT-PCR from 1 µg of total RNA from jejunum or ileum of control orspermine-treated suckling 14-day-old rats. Results are mean ± S.D. (N = 3)
IAP-1/GAPDH mRNA ratioJejunum Ileum
Control 1.58 ± 0.10 1.27 ± 0.42Spermine-treated 2.66 ± 1.69 1.25 ± 0.15
492 O. Peulen et al. / Biochimie 86 (2004) 487–493

[13] O. Peulen, G. Dandrifosse, Spermine-induced maturation in Wistar ratintestine: a cytokine dependent mechanism, J. Pediatr. Gastroenterol.Nutr. 38 (2004) 524–532.
[14] K. Yeh, M. Yeh, P.R. Holt, D.H. Alpers, Development and hormonalmodulation of postnatal expression of intestinal alkaline phosphatasemRNA species and their encoded isoenzymes, Biochem. J 301 (1994)893–899.
[15] A. Dahlqvist, Method for assay of intestinal disaccharidases, Anal.Biochem 7 (1964) 18–25.
[16] M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein–dyebinding, Anal. Biochem 72 (1976) 248–254.
[17] P.F. Millington, P.W. Tovell, Quantitative changes in alkaline phos-phatase in epithelial cells of rat small intestine from birth to weaning,Histochem. J. 1 (1969) 311–321.
[18] S. Rozen, H.J. Skaletsky, Primer3 on the WWW for general users andfor biologist programmers, in: S. Krawetz, S. Misener (Eds.), Bioin-formatics Methods and Protocols: Methods in Molecular Biology,Humana Press, Totoya, 2000, pp. 365–386.
[19] Q. Xie, D.H. Alpers, The two isozymes of rat intestinal alkalinephosphatase are products of two distinct genes, Physiol. Genom 3(2000) 1–8.
[20] S. Miura, M. Yoshioka, Y. Hamada, A. Morita, H. Asakura,M. Tsuchiya, Alkaline phosphatase of rat intestinal lymph: its charac-terization and the effect of fat administration, Clin. Chim. Acta 186(1990) 239–248.
[21] J.P. Buts, N. De Keyser, J. Kolanowski, E. Sokal, F. Van Hoof,Maturation of villus and crypt cell functions in rat small intestine.Role of dietary polyamines, Dig. Dis. Sci 38 (1993) 1091–1098.
[22] K. Shimizu, S. Mushiake, N. Yoshimura, T. Harada, S. Okada, Theeffect of spermine on the disaccharidase activities in suckling rats ofdifferent age, Cell Biol. Int 17 (1993) 543–546.
[23] G.E. Wild, A.S. Daly, N. Sauriol, G. Bennett, Effect of exogenouslyadministered polyamine on the structural maturation and enzymeontogeny of the postnatal rat intestine, Biol. Neonate 63 (1993)246–257.
[24] G. Capano, K.J. Bloch, E.J. Schiffrin, J.A. Dascoli, E.J. Israel,P.R. Harmatz, Influence of the polyamine, spermidine, on intestinalmaturation and dietary antigen uptake in the neonatal rat, J. Pediatr.Gastroenterol. Nutr 19 (1994) 34–42.
[25] E. Harada, Y. Hashimoto, B. Syuto, Orally administered spermineinduces precocious intestinal maturation of macromolecular transportand disaccharidase development in suckling rats, Comp. Biochem.Physiol. A Mol. Integr. Physiol 109 (1994) 667–673.
[26] B. Dorhout, A. van Faassen, C.M. van Beusekom, A.W. Kingma, E. deHoog, G.T. Nagel, A. Karrenbeld, E.R. Boersma, F.A. Muskiet, Oraladministration of deuterium-labelled polyamines to sucking rat pups:luminal uptake, metabolic fate and effects on gastrointestinal matura-tion, Br. J. Nutr 78 (1997) 639–654.
[27] J.C. ter Steege, W.A. Buurman, P.P. Forget, Spermine induces matu-ration of the immature intestinal immune system in neonatal mice, J.Pediatr. Gastroenterol. Nutr 25 (1997) 332–340.
[28] F. Moog, The functional differentiation of the small intestine. II. Thedifferentiation of alkaline phosphomonoesterase in the duodenum ofthe mouse, J. Exp. Zool 118 (1951) 187–208.
[29] K. Ono, The fine structure and localization of alkaline phosphataseactivity of the small intestinal epithelium in the postnatal developingrat, Acta Histochem 52 (1975) 117–133.
[30] M. Kaouass, J. Sulon, P. Deloyer, G. Dandrifosse, Spermine-inducedintestinal postnatal maturation : are glucocorticoids necessary? Arch.Int. Physiol. Biochim 101 (1992) B118.
[31] M. Kaouass, J. Sulon, P. Deloyer, G. Dandrifosse, Spermine-inducedprecocious intestinal maturation in suckling rats: possible involve-ment of glucocorticoids, J. Endocrinol 141 (1994) 279–283.
[32] T. Uezato, M. Fujita, Developmental transition of alkaline phos-phatase from suckling to adult type in rat small intestine: molecularspecies and effect of injected cortisone and thyroxine, J. Biochem 94(1983) 1483–1488.
[33] K.Y. Yeh, M. Yeh, P.R. Holt, Differential effects of thyroxine andcortisone on jejunal sucrase expression in suckling rats, Am. J.Physiol 256 (1989) G604–G612.
[34] S.T. Yedlin, G.P. Young, B. Seetharam, S. Seetharam, D.H. Alpers,Characterization and comparison of soluble and membranous formsof intestinal alkaline phosphatase from the suckling rat, J. Biol. Chem256 (1981) 5620–5626.
[35] N.L. Sussman, S. Seetharam, M.C. Blaufuss, D.H. Alpers, Translationof rat intestinal RNA yields two alkaline phosphatases, Biochem. J234 (1986) 563–568.
[36] M.J. Engle, A. Mahmood, D.H. Alpers, Two rat intestinal alkalinephosphatase isoforms with different carboxyl-terminal peptides areboth membrane-bound by a glycan phosphatidylinositol linkage, J.Biol. Chem 270 (1995) 11935–11940.
[37] H. Wada, I. Yagami, N. Niwa, T. Hayakawa, H. Tsuge, Distributionand properties of rat intestinal alkaline phosphatase isoenzymes, Exp.Anim 50 (2001) 153–158.
[38] T. Komoda, Y. Sakagishi, The function of carbohydrate moiety andalteration of carbohydrate composition in human alkaline phos-phatase isoenzymes, Biochim. Biophys. Acta 523 (1978) 395–406.
[39] S. Greco, I. Hugueny, P. George, P. Perrin, P. Louisot, M.C. Biol,Influence of spermine on intestinal maturation of the glycoproteinglycosylation process in neonatal rats, Biochem. J 345 (2000) 69–75.
[40] S. Greco, P. George, I. Hugueny, P. Louisot, M.C. Biol, Spermidine-induced glycoprotein fucosylation in immature rat intestine, C.R.Acad. Sci. III 322 (1999) 543–549.
[41] S. Greco, E. Niepceron, I. Hugueny, P. George, P. Louisot, M.C. Biol,Dietary spermidine and spermine participate in the maturation ofgalactosyltransferase activity and glycoprotein galactosylation in ratsmall intestine, J. Nutr 131 (2001) 1890–1897.
[42] C. Calhau, F. Martel, C. Hipolito-Reis, I. Azevedo, Differencesbetween duodenal and jejunal rat alkaline phosphatase, Clin. Biochem33 (2000) 571–577.
493O. Peulen et al. / Biochimie 86 (2004) 487–493





























