Embed Size (px)
Citation preview

Differential effects of enalapril and losartan on bodycomposition and indices of muscle quality in aged maleFischer 344 × Brown Norway rats
Christy S. Carter & Silvia Giovaninni & Dong-Oh Seo & Jameson DuPree &
Drake Morgan & Hae Young Chung & Hazel Lees & Michael Daniels &
Gene B. Hubbard & Shuko Lee & Yuji Ikeno & Thomas C. Foster &
Thomas W. Buford & Emanuele Marzetti
Received: 14 July 2010 /Accepted: 14 November 2010 /Published online: 14 December 2010# American Aging Association 2010
Abstract The primary purpose of the present set ofstudies was to provide a direct comparison of theeffects of the angiotensin-converting enzyme inhibitorenalapril and the angiotensin receptor blocker losartanon body composition, physical performance, andmuscle quality when administered late in life to agedrats. Overall, enalapril treatment consistently attenu-ated age-related increases in adiposity relative to bothplacebo and losartan. The maximal effect was
achieved after 3 months of treatment (between 24and 27 months of age), at a dose of 40 mg/kg and wasobserved in the absence of any changes in physicalactivity, body temperature, or food intake. In addition,the reduction in fat mass was not due to changes inpathology given that enalapril attenuated age-relatedincreases in tumor development relative to placebo-and losartan-treated animals. Both enalapril andlosartan attenuated age-related decreases in grip
AGE (2011) 33:167–183DOI 10.1007/s11357-010-9196-y
C. S. Carter (*) : S. Giovaninni :D.-O. Seo : J. DuPree :H. Lees : T. W. Buford : E. MarzettiDepartment of Aging and Geriatric Research,Institute on Aging, University of Florida,Gainesville, FL 32610, USAe-mail: [email protected]
S. GiovaninniDepartment of Gerontology, Geriatrics and PhysicalSciences, Catholic University of the Sacred Heart,Rome, Italy
D. MorganDepartment of Psychiatry, University of Florida,Gainesville, FL 32610, USA
H. Y. ChungDepartment of Pharmacy, Longevity Scienceand Technology Institutes, Research Institute for DrugDevelopment, Pusan National University,Geumjeong-gu,Busan 609-735, South Korea
M. DanielsDepartment of Statistics, University of Florida,Gainesville, FL 32610, USA
G. B. Hubbard :Y. IkenoBarshop Institute and Department of Pathology,University of Texas Health Science Center,San Antonio, TX 78245, USA
S. Lee :Y. IkenoAudie Murphy VA Hospital (STVHCS),San Antonio, TX 78245, USA
T. C. FosterDepartment of Neuroscience, University of Florida,Gainesville, FL 32610, USA
E. MarzettiDepartment of Orthopedics and Traumatology,Catholic University of the Sacred Heart,Rome, Italy

strength, suggesting that changes in body compositionappear dissociated from improvements in physicalfunction and may reflect a differential impact ofenalapril and losartan on muscle quality. To linkchanges in adiposity to improvements in skeletalmuscle quality, we performed gene array analyses togenerate hypotheses regarding cell signaling path-ways altered with enalapril treatment. Based on theseresults, our primary follow-up pathway wasmitochondria-mediated apoptosis of myocytes. Rela-tive to losartan- and placebo-treated rats, onlyenalapril decreased DNA fragmentation and caspase-dependent apoptotic signaling. These data suggestthat attenuation of the severity of skeletal muscleapoptosis promoted by enalapril may represent adistinct mechanism through which this compoundimproves muscle strength/quality.
Keywords Age-related adiposity . Body composition .
Sarcopenia . Renin–angiotensin system . Physicalfunction .Muscle quality
Introduction
Age-related changes in body composition haveimportant clinical implications, given that loss ofmuscle and gain of fat mass are independentlyassociated with declining performance as well asincreased risk for disability and mortality in olderpersons (Goodpaster et al. 2001, 2006; Newman et al.2003; Visser et al. 1998). Behavioral interventions,such as moderate calorie restriction and physicalexercise, have proven beneficial against age-associatedchanges in body composition in experimental settings;however, pharmacological approaches may be particu-larly relevant late in life, since not all older individualsbenefit from or are capable of participating in traditionaldiet and/or exercise programs. Therefore, the present setof studies was designed to provide a direct comparisonof the effects of the angiotensin-converting enzymeinhibitor (ACEi) enalapril and the angiotensin receptorblocker (ARB) losartan on body composition, musclequality, and physical function, when administered late inlife to aged rats.
Developing preclinical models of late-life inter-vention strategies for combating declining physicalfunction has enormous significance (de Grey 2007;Rae et al. 2010). With the continued “graying” of the
worldwide population, the number of individuals atrisk of developing physical disability continues toincrease and the skyrocketing social, emotional, andeconomic cost (Olshansky et al. 2009) of caring forsuch individuals mandates the need for testing theeffectiveness of health-promoting interventions withinthis cohort. To address this need, we have used theFischer 344×Brown Norway (F344BN) rat as ourmodel, since several studies have shown that thisstrain proceeds from 80% to 50% mortality between24 and 30 months of age. In humans, this samepattern of survival mirrors an exponential increase indisability. In fact, in both rats and humans, assessmentof functional limitations in the 50% survival range ishighly predictive of future disability and ultimatelymortality (Carter et al. 2002; Guralnik et al. 1994). Inthe context of changing body composition, a similarassociation exists (Newman et al. 2001, 2003;Goodpaster et al. 2006). In the F344BN, there is agradual increase in both lean and fat mass through24 months of age, a decrease in muscle, a furtherincrease in fat through 27 months, and a decrease inboth compartments thereafter (Carter et al. 2004;Rice et al. 2005). The similarity between theF344BN rat strain and humans in terms of timingof age-related body composition changes and declin-ing performance makes this strain a reasonablemodel for studying the relationship between age,adiposity, muscle quality, and physical function.
Observational studies in humans and convergingevidence from our history of preclinical studiessuggest that the use of ACEis attenuates age-relatedincreases in whole-body adiposity and physicalperformance decline in the absence of any sizeablemuscle hypertrophic effect (Bahi et al. 2004; Foianiniet al. 2000; Gayagay et al. 1998; Carter et al. 2004,2005; Onder et al. 2002). These data lend credence tothe hypothesis that it is not necessary to optimize thequantity of muscle that is preserved with an interven-tion but that it is the quality of muscle that remainswhich determines functionality. Since then, severalother studies have demonstrated similar effects ofACEis, in particular enalapril, on body composition invarious strains of rats and mice, across different ages,and under normal and high-fat feeding scenarios (deCavanagh et al. 2007; Santos et al. 2009; Weisinger etal. 2009b). Taken together, these data suggest that thephysiological mechanism by which ACEis mayenhance muscle quality and subsequently physical
168 AGE (2011) 33:167–183

function is through their impact on whole-bodyadiposity.
However, the point is further complicated giventhat ACEis have dual physiological actions: (1) theyblock angiotensin II (ANGII) production by prevent-ing the conversion of angiotensin 1 (ANGI) to ANGIIand (2) they block of the proteolytic degradation ofbradykinin (BK) (Henriksen and Jacob 2003). There-fore, in the current study, we directly compared theadiposity reducing effectiveness of enalapril withlosartan, an ARB which only blocks the action ofANGII by antagonizing the ANGII type 1 (AT1)receptor. This allows for a pharmacological approachin dissociating the contribution of each pathway(ANGII vs BK) in mitigating age-related increasesin adiposity. We also measured a variety of physio-logical endpoints which may themselves cause areduction in body weight (food intake, temperature,glucose and insulin levels, pathology) and which maybe secondary to the effects of enalapril. Furthermore,in order to expand our understanding of how thisdecrease in adiposity may play a role in preservingskeletal muscle quality, we used gene array technol-ogy in order to identify potential cell signalingpathways that are altered with enalapril treatmentand which may contribute to declining function.Based on these data, we focused on mitochondria-mediated apoptosis in both enalapril- and losartan-treated animals to further assess whether thispathway may be differentially regulated by thesetwo treatments.
Methods
Animals
For all studies, we used male F344BN rats purchasedfrom the National Institute on Aging (NIA) Colony atHarlan Industries (Indianapolis, IN). The F344BN ratstrain has been extensively used for aging studies andwas selected for these experiments given that theseanimals demonstrate age-related body compositionchanges (i.e., increase in adiposity and decrease inlean mass) resembling those occurring in humans(Schwartz 1998). Animals were received at 22 monthsof age and housed individually on a 12-h light/darkcycle in a specific pathogen-free facility accredited bythe American Association for Accreditation of Labo-
ratory Animal Care. From 22 to 24 months of age,animals were allowed to acclimate to their housingconditions and to establish baseline rates of foodintake and body weights. All experimental protocolswere approved by the University of Florida’s AnimalCare and Use Committee.
Experimental design
The first experiment consisted of a longitudinalstudy to directly compare the effects of enalapriland losartan and to replicate our previous publishedfindings that enalapril both reduces whole-bodyadiposity and mitigates declining physical function(Carter et al. 2004). Rats were randomly assigned toreceive 40 mg/kg enalapril (n=16) 30 mg/kg losartan(n=18) or placebo (n=19) from 24 to 30 months ofage. Drug delivery was accomplished by compound-ing the various treatments into bacon-flavored foodtablets (Bio-Serv, Frenchtown, NJ). Placebo-containing food tablets were identical to thosedelivering enalapril or losartan, except that the drugwas omitted. Determinations of body compositionand physical performance were performed at base-line (24 months), 27 and 30 months of age. Theresults of these experiments demonstrated that onlyenalapril induces a decrease in adiposity, with themaximal effect occurring between 24 and 27 monthsof age.
Based on the results of the first experiment, thesecond experiment was designed to characterize theeffects of enalapril (20 or 40 mg/kg; n=33 in eachgroup for a total of n=66), losartan (30 mg/kg; n=39), and placebo (n=34) on markers of skeletalmuscle aging over a narrower age range (24 to27 months of age). This timeframe represents thecritical window determined in the first experiment andreflects timing during the aging process, where malesof this strain continue to gain fat mass whilesimultaneously losing muscle mass (Rice et al.2005). We performed gene array analyses (enalapril40 mg/kg and placebo; n=4 each group) to generatehypotheses regarding cell signaling pathways thatmight be altered with enalapril treatment. Based onthese results, our primary follow-up pathway wasmitochondria-mediated apoptosis of myocytes. Therationale to focus on mitochondria-driven apoptosisrelies on the major role postulated for this apoptoticpathway in the pathogenesis of sarcopenia of aging
AGE (2011) 33:167–183 169

which is correlated with declining physical function(Marzetti et al. 2009b). The muscle selected for thisstudy was the gastrocnemius, which undergoessignificant age-related atrophy accompanied by ele-vations in apoptosis markers (Marzetti et al. 2008b,2009a). To ensure that any observed change in bodycomposition was not secondary to other physiologicalprocesses, which by themselves are known to impactbody weight and composition, we measured locomo-tor activity, body temperature, food intake, andglucose and insulin levels. In addition, we assessedoverall tissue pathology, in collaboration with theBarshop Longevity Institute, to ensure that thetreatment itself did not promote conditions, such asincreased tumorigenesis, that would also result inweight loss.
Methods for experiment 1
Determination of body composition via time-domainnuclear magnetic resonance Body composition wasdetermined by time-domain nuclear magnetic reso-nance (TD-NMR) in restrained but awake and alertrats (TD-NMR Minispec, Bruker Optics, The Wood-lands, TX, USA). The MiniSpec identifies threecomponents of body composition (fat, free body fluid,and lean tissue in grams) by acquiring and analyzingTD-NMR signals from all protons in the sample area.Scans were acquired by putting live conscious rats(i.e., without anesthetics) into a sample holder(90-mm diameter and ~250-mm length) with a screwtop that tightens to the length of the rat. The sampleholder was then inserted into the analyzer. The totalscan time for each rat was approximately 2 min.Based on the finding that the greatest change in bodyweight occurred between approximately 24 and27 months of age, for each individual animal, wecalculated the percent change from baseline, for bothlean and fat grams, from 24 to 27 months of age. Atwo-way repeated analysis of variance (ANOVA) wasused for testing the effects of time (24 vs 27 months)and treatment (enalapril 40 mg/kg vs losartan 30 mg/kg vs placebo). This same methodology was appliedin experiment 2 at 24 and 27 months of age andanalyzed using the same statistical test describedabove.
Assessment of physical performance Forelimb gripstrength was determined using an automated grip
strength meter (Columbus Instruments, Columbus,OH). The rat was grasped by the tail and suspendedabove a grip ring. After about 3 s, the rat was gentlylowered toward the grip ring and allowed to grasp thering with its forepaws. The remainder of the rat’sbody was quickly lowered to a horizontal position andthe animal's tail pulled until its grasp of the ring wasbroken. The force in grams was determined with acomputerized electronic pull strain gauge that wasfitted directly to the grasping ring, for three consec-utive pulls, and the maximum was determined. Atwo-way repeated ANOVA was used for testing theeffects of time (24 vs 27 vs 30 months) and treatment(enalapril 40 mg/kg vs losartan 30 mg/kg vs placebo).Data are reported as % change of kilogram of forceper kilogram of body weight.
Methods for experiment 2
Measurement of locomotor activity One day prior tothe TD-NMR experiments at 24 and 27 months ofage, locomotor activity was assessed. At the onsetof the dark cycle, rats were brought into theprocedure room in their home cages and allowedto acclimate to the environment for 1 h. Rats werethen placed into activity monitors (Med Associates,St. Albans VT) and allowed to acclimate for 15 minand then were monitored for 1 h. These devices arecubicles (43.5×43.5×23 cm), with a series ofinfrared emitters and receivers arrayed along thesides. These allow the automatic recording of ananimal’s position and distance moved (cm) withinthe chamber. Total distance traveled (cm) across the1-h session was the final unit of measure. A two-way repeated ANOVA was used for testing theeffects of time (24 vs 27 months) and treatment(enalapril 20 mg/kg vs enalapril 40 mg/kg vslosartan 30 mg/kg vs placebo).
Determination of body temperature via implantedmicrochip A glass-encapsulated IPTT-300 tempera-ture transponder was implanted subcutaneously ineach rat (Biomedic Data Systems, Inc., Seaford DE),while under momentary light anesthesia (approxi-mately 2% isoflurane at 2 ml/min O2). The device isdesigned for harmless nonsurgical subcutaneousimplantation and is approximately 14 mm in lengthby 2 mm in diameter. Temperature data were transmit-
170 AGE (2011) 33:167–183

ted to a dedicated handheld reader. Three consecutivetemperatures were obtained and averaged, at 24 and27 months of age. A two-way repeated ANOVA wasused for testing the effects of time (24 vs 27months) andtreatment (enalapril 20 mg/kg vs enalapril 40 mg/kg vslosartan 30 mg/kg vs placebo).
Serum measurements Rats were sacrificed by rapiddecapitation, using a dedicated guillotine. Wholeblood was collected and processed for subsequentdetermination of serum glucose and insulin. Theabdomen was quickly opened, and tissues wereremoved, placed in 2-ml cryovials, flash-frozen inliquid nitrogen, and stored at −80°C. Measurement ofblood glucose and insulin was performed by theHypertension Core Laboratory at the Wake ForestUniversity School of Medicine, using standardizedradioimmunoassay procedures, after solid-phase extrac-tion (Sep-Pak, Waters, Milford, MA). Data are pre-sented as nanogram per deciliter and nanogram permilliliter for glucose and insulin, respectively. A one-way ANOVA was used for testing the effects oftreatment (enalapril 20 mg/kg vs enalapril 40 mg/kg vslosartan 30 mg/kg vs placebo).
RNA extraction and Affymetrix gene array Genearray analyses were performed on rats from the40 mg/kg enalapril and placebo groups (n=4 pergroup) in order to develop initial hypotheses regard-ing enalapril’s effect on skeletal muscle quality. RNAwas isolated from the soleus muscle sample using theRNeasy Mini Kit (Qiagen, Germantown, MD). RNAconcentration was determined using UV spectropho-tometry (NanoDrop ND-1000), and RNA quality wasconfirmed using Agilent 2100 bioanalyzer (SantaClara, CA). Following confirmation of RNA quality,5 μg RNA was synthesized to cRNA using Affyme-trix amplification kit (Santa Clara, CA) following themanufacturer’s protocol. Hybridization of cRNA wascarried out by the University of Florida Interdisci-plinary Center for Biotechnology Research Micro-array Core. Hybridization of Affymetrix Rat 1.0arrays occurred for 17 h at 60°C in accordance withmanufacturer’s instructions, and arrays were scannedusing an Affymetrix Microarray scanner. Images wereanalyzed using Affymetrix Gene Chip OperatingSystem software (version 1.1) and scaled to 500.Raw data were normalized by filtering of the probesby “present” and “marginal” flags. A “present,”
“absent,” or “marginal” flag was designated to aprobe based on signal intensity and background noise.The number of “present” calls was determined acrossall chips, and the probe was removed if fewer than75% of the chips exhibited a “present” call for theprobe. The remaining probe sets were further filteredto remove expressed sequence tags and probes forhypothetical proteins and pseudo-genes that did nothave an indication of biological or molecular func-tion. Following this filtering process, a total of 9,017network eligible genes remained. Differential expres-sion between groups was determined using two-tailedt tests with an alpha level of 0.025. These differen-tially expressed transcripts were functionally annotat-ed using the Database for Annotation, Visualization,and Integrative Discovery (david.abcc.ncifcrf.gov)and submitted to Ingenuity Pathway Analysis (IPA,Ingenuity Systems, Redwood City, CA) to determinetheir roles in networks, cellular functions, andcanonical pathways. The IPA program uses a right-tailed Fisher’s exact test to compute the likelihoodthat the relationship between the list of submittedgenes and a set of genes representing a given pathwayis due to chance.
Subcellular fractionation of gastrocnemius musclesamples Isolation of cytosolic, mitochondrial, andnuclear fractions was performed as detailed elsewhere(Marzetti et al. 2008b). Protein concentration in thecytosolic and mitochondrial fractions was determinedby the method developed by Bradford (1976),whereas the detergent-compatible DC assay (Bio-Rad,Hercules, CA) was employed for nuclear extracts.Subcellular fractions were subsequently aliquoted andstored at −80°C until analysis.
Determination of the extent of skeletal muscleapoptosis Overall levels of apoptosis in the gastroc-nemius muscle were quantified by measuring theamount of cytosolic mononucleosomes and oligonu-cleosomes using an enzyme-linked immunosorbentassay (ELISA) kit (cell death detection ELISA; RocheDiagnostics, Mannheim, Germany), as previouslydescribed (Marzetti et al. 2008a). The assay relies onthe quantification of histone-complexed fragmentedDNA. Although the kit does not allow for thediscrimination between apoptotic and necrotic celldeath in cytosolic extracts, occurrence of significantnecrosis in skeletal muscle during normal aging has
AGE (2011) 33:167–183 171

not been reported. Therefore, the impact of necroticcell death in our system may be considered negligible.Absorbance was measured at 405 nm with a SynergyHT multidetection microplate reader (BioTek,Winooski, VT) and reported as arbitrary opticaldensity (OD) units per milligram of protein (apoptoticindex). A one-way ANOVA was used for testing theeffects of treatment (enalapril 20 mg/kg vs enalapril40 mg/kg vs losartan 30 mg/kg vs placebo) on allmeasures.
Western blot analysis for the determination of keymitochondrial apoptotic signaling proteins A vastliterature supports a central role for mitochondria-driven apoptosis during the development of sarcope-nia (Marzetti et al. 2009b). Therefore, we assessedseveral proteins integral to the regulation and execu-tion of mitochondrial apoptotic signaling via Westernimmunoblot analysis of gastrocnemius subcellularfractions. Specifically, we determined expressionlevels of both caspase-dependent (i.e., cytochrome c,active caspase 9, and cleaved caspase 3) and caspase-independent [apoptosis-inducing factor (AIF) andendonuclease G (EndoG)] mitochondrial apoptogenicmediators in specific subcellular compartments. Fur-thermore, we measured mitochondrial levels of theanti-apoptotic Bcl-2 and pro-apoptotic Bax, giventheir central role in controlling cell fate (Marzetti et al.2009b). Experiments were performed as detailedelsewhere (Marzetti et al. 2009a). The followingprimary antibodies and relative dilutions were used:rabbit monoclonal anti-cleaved cleaved-caspase-3(Cell Signaling Technology, Beverly, MA), 1:1,000;rabbit polyclonal anti-Bcl-2 (Santa Cruz Biotechnol-ogy, Santa Cruz, CA), 1:200; rabbit polyclonal anti-Bax (Santa Cruz Biotechnology), 1:200; rabbit poly-clonal anti-cytochrome c (Santa Cruz Biotechnology),1:200; rabbit polyclonal anti-active caspase-9 (SantaCruz Biotechnology), 1:200; rabbit polyclonal anti-EndoG (Abcam, Cambridge, MA), 1:1,000; rabbitpolyclonal anti-EndoG (Abcam), 1:200; and rabbitpolyclonal anti-AIF (BD Pharmingen, San Diego,CA), 1:500. Generation of the chemiluminescentsignal, digital acquisition, and densitometry analysiswere performed as previously described (Marzetti etal. 2008b). Spot density of target bands was normal-ized to the amount of protein loaded in each lane, asdetermined by densitometric analysis of thecorresponding Ponceau S-stained membranes, and
expressed as arbitrary OD units (Image Lab 2.0.1,Bio-Rad Laboratories). For each measure, a one-wayANOVA was used for testing the effects of treatment(enalapril 20 mg/kg vs enalapril 40 mg/kg vs losartan30 mg/kg vs placebo).
Pathology After rats were necropsied for grosspathological lesions, organs and tissues were ex-cised and preserved in 10% buffered formalin.Organs and tissues analyzed included brain, pitui-tary gland, heart, lung, trachea, thymus, aorta,esophagus, stomach, small intestine, colon, liver,pancreas, spleen, kidneys, urinary bladder, repro-ductive system (prostate, testes, epididymis, andseminal vesicles), thyroid gland, adrenal glands,parathyroid glands, psoas muscle, knee joint,sternum, and vertebrae. Any other tissue with grosslesions was also excised. Fixed tissues wereprocessed conventionally, embedded in paraffin,sectioned at 5 μm, and stained with hematoxylin–eosin. Diagnosis of each histopathological changewas established based on histological classificationsin aging rats (Maeda et al. 1985; Iwasaki et al.1988). The prevalence of tumor-bearing rats andoverall specific incidence of disease was calculatedfor each experimental group. The percentage oftumor-bearing rats was calculated as the percentageof animals that had one or more neoplastic lesions.For this assessment, all neoplastic lesions werecounted regardless of the severity of tumors, i.e.,both incidental (not severe enough to be the cause ofdeath) and fatal (severe enough to be the cause ofdeath) tumors were considered. Chronic nephropathywas graded in the order of increasing severity basedon the grading system described by Yu et al. (1982)as follows: grade 0 (no lesions), grade 1, grade 2,grade 3, grade 4, and grade E (very severe). Gradingof neoplastic lesions was based on a modification ofpreviously reported criteria (Ikeno et al. 2003):grade 1, primary site only; grade 2, primary site andintra-organ or one other organ metastasis; grade 3,metastasis to two to three organs; and grade 4,metastasis to more than four organs or grade 3+additional pathology (e.g., pleural effusion, ascites,subcutaneous edema, etc.). Hydrothorax, ascites, andsubcutaneous edema were the most common com-plications associated with advanced neoplastic dis-eases. Pathological comparisons of tumor-bearingrats and number of neoplastic and non-neoplastic
172 AGE (2011) 33:167–183
diseases (i.e., chronic nephropathy) among groupswere examined by Mantel–Haenszel chi-squared test.The average number of tumors per rat, total number ofpathologies, and average severity of nephropathy werecompared among groups using the Kruskal–Wallis test.
Statistical analysis
Statistical analyses were performed using SAS 9.1 forWindows (Cary, NC). Multiple pairwise group com-parisons were performed using the Bonferroni proce-dure. The level of significance was set at p<0.05 forall analyses. All data are presented as means±standard error.
Results
Experiment 1
Body weight and composition
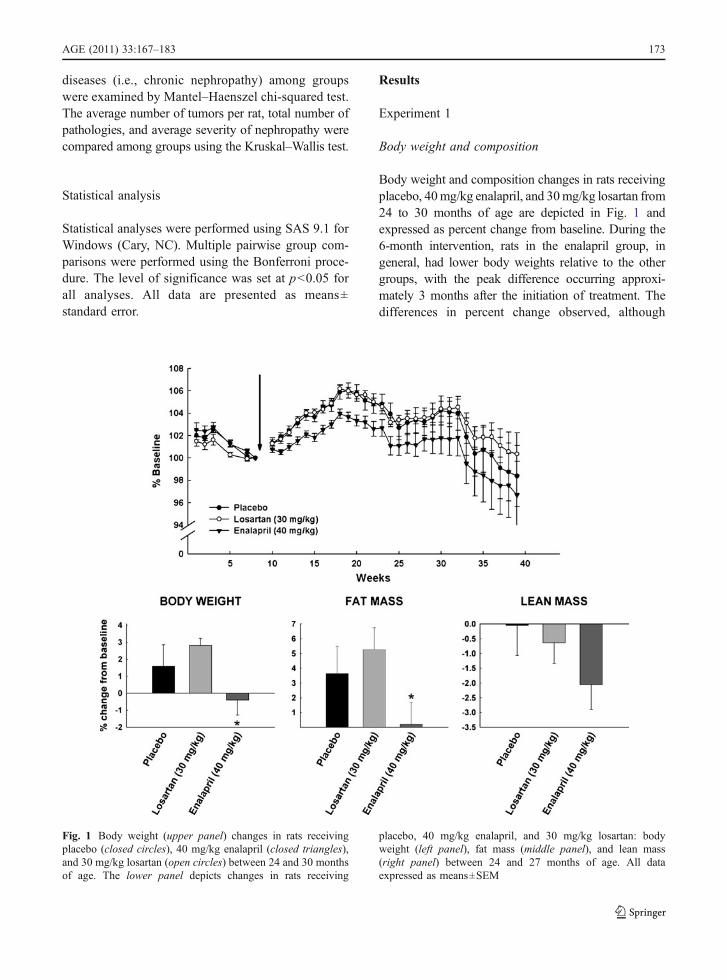
Body weight and composition changes in rats receivingplacebo, 40mg/kg enalapril, and 30mg/kg losartan from24 to 30 months of age are depicted in Fig. 1 andexpressed as percent change from baseline. During the6-month intervention, rats in the enalapril group, ingeneral, had lower body weights relative to the othergroups, with the peak difference occurring approxi-mately 3 months after the initiation of treatment. Thedifferences in percent change observed, although
Fig. 1 Body weight (upper panel) changes in rats receivingplacebo (closed circles), 40 mg/kg enalapril (closed triangles),and 30 mg/kg losartan (open circles) between 24 and 30 monthsof age. The lower panel depicts changes in rats receiving
placebo, 40 mg/kg enalapril, and 30 mg/kg losartan: bodyweight (left panel), fat mass (middle panel), and lean mass(right panel) between 24 and 27 months of age. All dataexpressed as means±SEM
AGE (2011) 33:167–183 173
relatively small (~4%), were statistically significant andreflect the larger gains in body weight observed duringthe first 3 months of the experiment in the placebo andlosartan groups relative to the enalapril-treated rats(p=0.027 and 0.015, respectively). Analysis ofchanges in body composition from baseline to 3 monthsshowed an increase in fat mass in the placebo (p=0.016)group and in rats treated with losartan (p=0.019),whereas the opposite pattern was detected in theenalapril group (p=0.017). At 3 months, there was atrend towards a decrease in lean mass loss in theenalapril group, but this change was not statisticallysignificant (p=0.06). By the end of the study, nostatistical differences were observed among groups inany measures of body weight or composition.
Physical performance
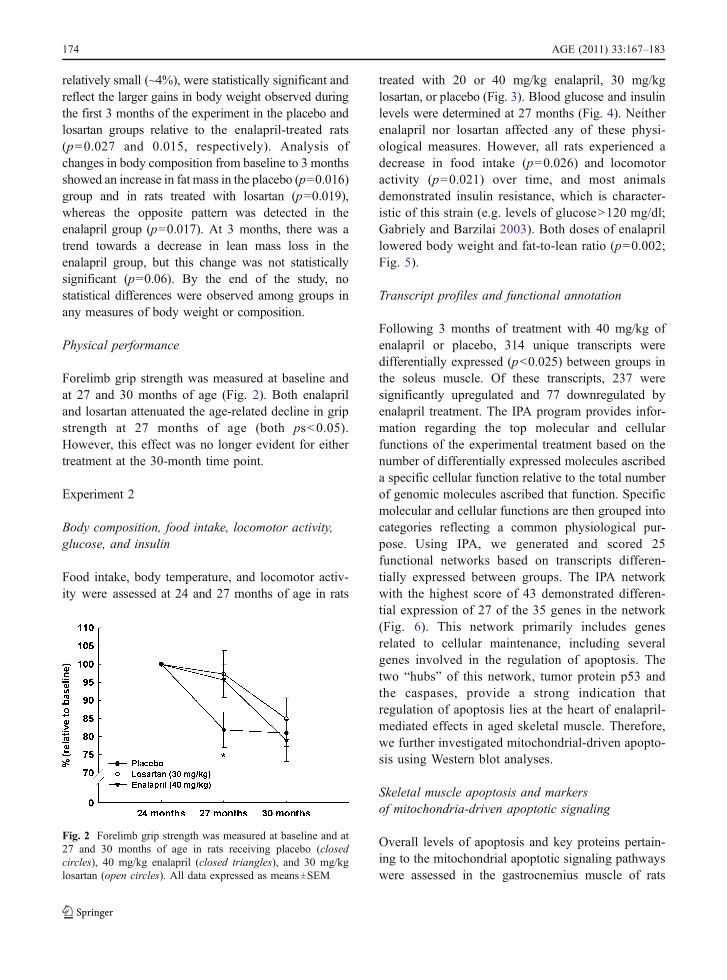
Forelimb grip strength was measured at baseline andat 27 and 30 months of age (Fig. 2). Both enalapriland losartan attenuated the age-related decline in gripstrength at 27 months of age (both ps<0.05).However, this effect was no longer evident for eithertreatment at the 30-month time point.
Experiment 2
Body composition, food intake, locomotor activity,glucose, and insulin
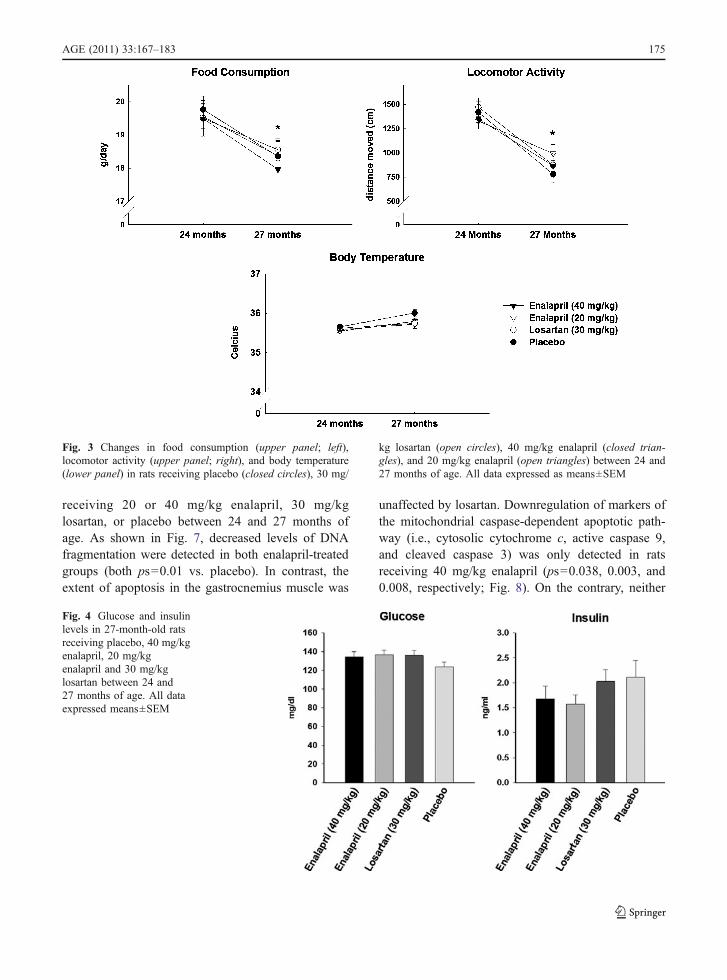
Food intake, body temperature, and locomotor activ-ity were assessed at 24 and 27 months of age in rats
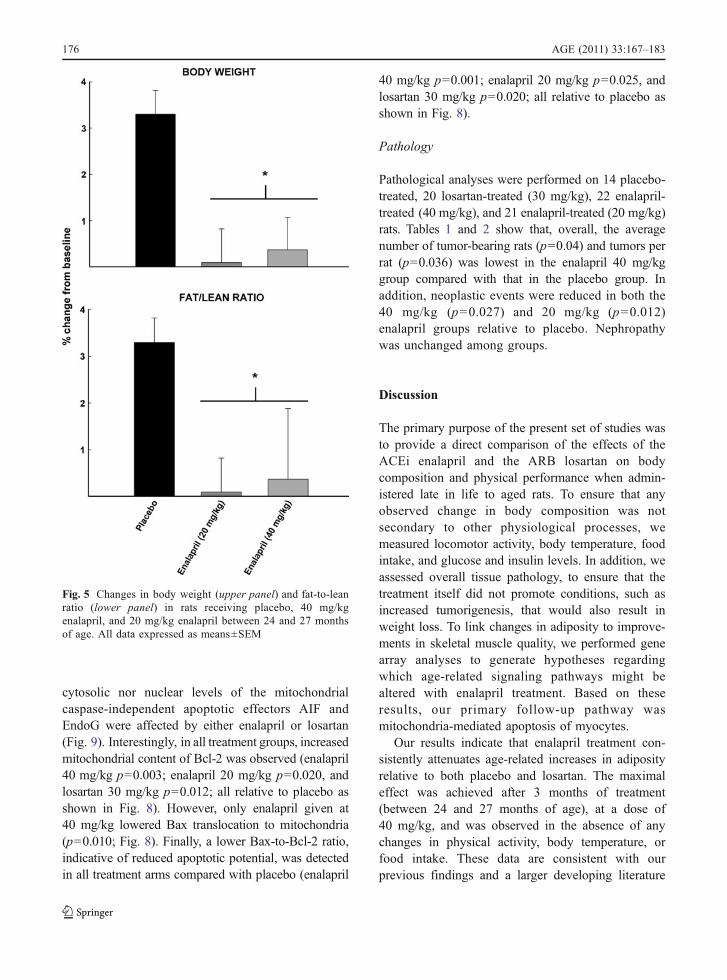
treated with 20 or 40 mg/kg enalapril, 30 mg/kglosartan, or placebo (Fig. 3). Blood glucose and insulinlevels were determined at 27 months (Fig. 4). Neitherenalapril nor losartan affected any of these physi-ological measures. However, all rats experienced adecrease in food intake (p=0.026) and locomotoractivity (p=0.021) over time, and most animalsdemonstrated insulin resistance, which is character-istic of this strain (e.g. levels of glucose>120 mg/dl;Gabriely and Barzilai 2003). Both doses of enalaprillowered body weight and fat-to-lean ratio (p=0.002;Fig. 5).
Transcript profiles and functional annotation
Following 3 months of treatment with 40 mg/kg ofenalapril or placebo, 314 unique transcripts weredifferentially expressed (p<0.025) between groups inthe soleus muscle. Of these transcripts, 237 weresignificantly upregulated and 77 downregulated byenalapril treatment. The IPA program provides infor-mation regarding the top molecular and cellularfunctions of the experimental treatment based on thenumber of differentially expressed molecules ascribeda specific cellular function relative to the total numberof genomic molecules ascribed that function. Specificmolecular and cellular functions are then grouped intocategories reflecting a common physiological pur-pose. Using IPA, we generated and scored 25functional networks based on transcripts differen-tially expressed between groups. The IPA networkwith the highest score of 43 demonstrated differen-tial expression of 27 of the 35 genes in the network(Fig. 6). This network primarily includes genesrelated to cellular maintenance, including severalgenes involved in the regulation of apoptosis. Thetwo “hubs” of this network, tumor protein p53 andthe caspases, provide a strong indication thatregulation of apoptosis lies at the heart of enalapril-mediated effects in aged skeletal muscle. Therefore,we further investigated mitochondrial-driven apopto-sis using Western blot analyses.
Skeletal muscle apoptosis and markersof mitochondria-driven apoptotic signaling
Overall levels of apoptosis and key proteins pertain-ing to the mitochondrial apoptotic signaling pathwayswere assessed in the gastrocnemius muscle of rats
Fig. 2 Forelimb grip strength was measured at baseline and at27 and 30 months of age in rats receiving placebo (closedcircles), 40 mg/kg enalapril (closed triangles), and 30 mg/kglosartan (open circles). All data expressed as means±SEM
174 AGE (2011) 33:167–183
receiving 20 or 40 mg/kg enalapril, 30 mg/kglosartan, or placebo between 24 and 27 months ofage. As shown in Fig. 7, decreased levels of DNAfragmentation were detected in both enalapril-treatedgroups (both ps=0.01 vs. placebo). In contrast, theextent of apoptosis in the gastrocnemius muscle was
unaffected by losartan. Downregulation of markers ofthe mitochondrial caspase-dependent apoptotic path-way (i.e., cytosolic cytochrome c, active caspase 9,and cleaved caspase 3) was only detected in ratsreceiving 40 mg/kg enalapril (ps=0.038, 0.003, and0.008, respectively; Fig. 8). On the contrary, neither
Fig. 3 Changes in food consumption (upper panel; left),locomotor activity (upper panel; right), and body temperature(lower panel) in rats receiving placebo (closed circles), 30 mg/
kg losartan (open circles), 40 mg/kg enalapril (closed trian-gles), and 20 mg/kg enalapril (open triangles) between 24 and27 months of age. All data expressed as means±SEM
Fig. 4 Glucose and insulinlevels in 27-month-old ratsreceiving placebo, 40 mg/kgenalapril, 20 mg/kgenalapril and 30 mg/kglosartan between 24 and27 months of age. All dataexpressed means±SEM
AGE (2011) 33:167–183 175
cytosolic nor nuclear levels of the mitochondrialcaspase-independent apoptotic effectors AIF andEndoG were affected by either enalapril or losartan(Fig. 9). Interestingly, in all treatment groups, increasedmitochondrial content of Bcl-2 was observed (enalapril40 mg/kg p=0.003; enalapril 20 mg/kg p=0.020, andlosartan 30 mg/kg p=0.012; all relative to placebo asshown in Fig. 8). However, only enalapril given at40 mg/kg lowered Bax translocation to mitochondria(p=0.010; Fig. 8). Finally, a lower Bax-to-Bcl-2 ratio,indicative of reduced apoptotic potential, was detectedin all treatment arms compared with placebo (enalapril
40 mg/kg p=0.001; enalapril 20 mg/kg p=0.025, andlosartan 30 mg/kg p=0.020; all relative to placebo asshown in Fig. 8).
Pathology
Pathological analyses were performed on 14 placebo-treated, 20 losartan-treated (30 mg/kg), 22 enalapril-treated (40 mg/kg), and 21 enalapril-treated (20 mg/kg)rats. Tables 1 and 2 show that, overall, the averagenumber of tumor-bearing rats (p=0.04) and tumors perrat (p=0.036) was lowest in the enalapril 40 mg/kggroup compared with that in the placebo group. Inaddition, neoplastic events were reduced in both the40 mg/kg (p=0.027) and 20 mg/kg (p=0.012)enalapril groups relative to placebo. Nephropathywas unchanged among groups.
Discussion
The primary purpose of the present set of studies wasto provide a direct comparison of the effects of theACEi enalapril and the ARB losartan on bodycomposition and physical performance when admin-istered late in life to aged rats. To ensure that anyobserved change in body composition was notsecondary to other physiological processes, wemeasured locomotor activity, body temperature, foodintake, and glucose and insulin levels. In addition, weassessed overall tissue pathology, to ensure that thetreatment itself did not promote conditions, such asincreased tumorigenesis, that would also result inweight loss. To link changes in adiposity to improve-ments in skeletal muscle quality, we performed genearray analyses to generate hypotheses regardingwhich age-related signaling pathways might bealtered with enalapril treatment. Based on theseresults, our primary follow-up pathway wasmitochondria-mediated apoptosis of myocytes.
Our results indicate that enalapril treatment con-sistently attenuates age-related increases in adiposityrelative to both placebo and losartan. The maximaleffect was achieved after 3 months of treatment(between 24 and 27 months of age), at a dose of40 mg/kg, and was observed in the absence of anychanges in physical activity, body temperature, orfood intake. These data are consistent with ourprevious findings and a larger developing literature
Fig. 5 Changes in body weight (upper panel) and fat-to-leanratio (lower panel) in rats receiving placebo, 40 mg/kgenalapril, and 20 mg/kg enalapril between 24 and 27 monthsof age. All data expressed as means±SEM
176 AGE (2011) 33:167–183

demonstrating selective loss of body fat compartmentwith ACEi treatment in a variety of species, undervarious feeding regimens and across various timepoints in the life span (Weisinger et al. 2009a, b;Santos et al. 2009). However, one exception we noteis that most studies, especially those in younganimals, report decreased food intake. For example,enalapril administration (10 mg/kg) to young adultrats, fed either regular or a high-fat chow, wasaccompanied by a decrease in food intake (Santos etal. 2009), although when adjusted for changing bodyweight, food intake is equivalent or increased. Asimilar finding has been reported in captopril-treatedyoung mice maintained on a high-fat diet (Weisingeret al. 2009b). One potential difference between ourstudy and those using young rats and mice as subjectsis that young rodents are highly leptin sensitive.Leptin is a primary modulator of ingestive behavior,and with age, the F344BN rat becomes leptin resistant(Zhang and Scarpace 2006; Scarpace and Zhang2009). It is therefore likely that the large losses in
fat mass observed in those studies using younganimals resulted in decreased leptin levels, initiatingan anorectic response. However, it is unlikely that thisloss of fat was a direct action of enalapril on leptinsignaling given that in the study of Santos et al.(2009), young rats fed regular chow or a high-fat diethad a similar anorectic response to ICV leptinadministration regardless of enalapril treatment. Ourgroup has previously demonstrated that enalapriltreatment in old rats did indeed result in a reductionin leptin levels commensurate with the loss of fatmass; however, this effect did not translate in anychange in food intake (Carter et al. 2004). In fact, inthe present study, all animals decrease food intakebetween 24 and 27 months of age, even though fatmass is increasing. Therefore, enalapril most likelyis working through other pathways which modulatefat loss (enhanced fatty acid oxidation, increasedadiponectin levels, and improved insulin and per-oxisome proliferator-activated receptor (PPAR)-γsignaling; Santos et al. 2009; Weisinger et al.
Fig. 6 Functional network with the highest score of 43 following IPA analysis. Red indicates upregulation; green indicatesdownregulation. Note that the two central hubs, TP53 and the caspase group, are associated with myonuclear apoptosis
AGE (2011) 33:167–183 177
2009a, b) rather than through decreased food intake.Future studies in older animals are needed to addressthese mechanisms.
The weight loss observed was also not due tochanges in pathology. In fact, quite the oppositeoccurred in that enalapril, at the highest dose,attenuated the age-related increase in tumor develop-
ment relative to placebo- and losartan-treated animals.This finding is especially remarkable in light of arecent meta-analysis showing an increased risk forcancer associated with chronic ARB treatment (Sipahiet al. 2010). However, lifespan was not an outcome inour studies given that enalapril administration whichoccurred was not administered over the entire life ofthese rats (only between 24 and 30 months of age).However, in a recent study, as part of the NIAIntervention Testing Program, 15 mg/kg enalapriladministered to mice from middle age throughoutthe lifespan did not impact maximal life span(Harrison et al. 2009). In contrast to these findings,others reported increased life span in laboratoryrodents treated with enalapril. For example, Santoset al. showed that enalapril, administered for26 months to Wistar rats, increased mean life span(Santos et al. 2009). The effect on maximum life spanwas not ascertainable given that the study wascensored at 26 months. In addition, Basso et al. foundthat inhibition of the renin–angiotensin system (RAS)via either enalapril (10 mg/kg) or losartan (30 mg/kg)increased life span in normotensive Wistar rats (Bassoet al. 2007). However, in this study, many animalswere removed during the experiment for otherexperimental purposes, thereby changing the compo-sition of the original population and making a
Fig. 8 Changes in caspase-dependent apoptotic signaling in rats receiving placebo, 40 mg/kg enalapril, 20 mg/kg enalapril, and30 mg/kg losartan between 24 and 27 months of age. All data expressed means±SEM
Fig. 7 Changes in apoptotic index as measured by DNAfragmentation in rats receiving placebo), 40 mg/kg enalapril,20 mg/kg enalapril, and 30 mg/kg losartan between 24 and27 months of age. All data expressed means±SEM
178 AGE (2011) 33:167–183
conclusive statement regarding life span problematic.Increased longevity has been observed in ACEi-treated mice and rats fed high-fat diets, althoughthese studies were also censored at particular ages(Weisinger et al. 2009b). Hence, additional research isneeded to establish the impact of pharmacological
RAS inhibition on life span, taking into considerationthe dose and duration of the intervention.
In the current study, losartan had no impact onbody composition. One interpretation is that blockadeof the actions of ANGII at the AT1 receptor has noanti-adiposity action. However, recent reports suggest
Fig. 9 Changes in caspase-independent apoptotic signaling in rats receiving placebo, 40 mg/kg enalapril, 20 mg/kg enalapril, and30 mg/kg losartan between 24 and 27 months of age. All data expressed means±SEM
Table 1 Number of tumor-bearing rats, average number of tumors per rat, total number of pathologies, and average severity ofnephropathy of each treatment group
Placebo Losartan (30 mg/kg) Enalapril (40 mg/kg) Enalapril (20 mg/kg)
Tumor-bearing rats 11 (78.6%) 12 (60%) 9 (39.1%)a 10 (45.5%)
Average number of tumors per rat 1 0.75 0.55a 0.62
Total number of pathologies 5.93 6.25 6.55 6.38
Average severity of nephropathy 1.07 0.6 0.86 1
Average number of tumors per rat: p=0.0366 vs group Pa Tumor-bearing rats: p=0.0407 vs group P
AGE (2011) 33:167–183 179

that ARBs other than losartan, such as telmisartan,prevent weight gain in normally fed and high-fat-fedmice through PPAR/AMPK signaling pathways (Fenget al. 2010; He et al. 2010). Indeed, the same series ofstudies showed, using an in vitro preparation (3 T3-L1 preadipocytes), that only telmisartan activatedPPAR- γ signaling when compared directly withlosartan. Furthermore, PPAR- γ and PPAR-deltaknockout mice do not respond with weight loss totelmisartan treatment. Most interestingly, mice fedtelmisartan had improved exercise endurance, accom-panied by a shift to increased slow-twitch musclefiber profile (Feng et al. 2010). Whether or not thiseffect of telmisartan is directly regulated at the AT1receptor or some other direct action of this drug onalternative signaling pathways is yet unknown.However, there is emerging evidence that telmisartanoperates as a partial PPAR-γ agonist and that itinhibits the proliferative capacity of some cells thatlack ANGII receptors (Benson et al. 2004). Thesedata highlight the need for comparative effectivenessof ARBs in late-life intervention studies, usingpreclinical models.
Importantly, at 27 months of age, both enalapriland losartan attenuated the age-dependent decline inphysical performance. However, in contrast to theenalapril group, in losartan-treated rats, this effectoccurred in the absence of changes in body weight orcomposition. This finding suggests that both com-pounds might improve physical performance througha direct action on the skeletal muscle, independent ofchanges in body mass. Therefore, we performed genearray analyses in order to develop working hypothe-ses regarding potential molecular signaling pathwayswhich might be altered with enalapril treatment, andapoptosis emerged as a leading candidate. This
hypothesis is supported by previous studies showingthat interventions reducing the severity of skeletalmuscle apoptosis improve physical function (Song etal. 2006; Marzetti et al. 2008a). Apoptosis, a processof programmed cell death, is a highly conserved andtightly regulated systematic set of events resulting incellular self-destruction without the induction ofinflammation or damage to the surrounding tissue(Kerr et al. 1972). Cysteine-aspartic proteases (cas-pases) are the executioner enzymes that carry out thedismantling of the cell and are normally present asinactive zymogens (procaspases) (Danial and Kors-meyer 2004). Upon apoptotic stimuli, initiator cas-pases (i.e., caspase 8, caspase 9, caspase 12) areactivated, subsequently leading to the activation ofeffector caspases (i.e., caspase 3, caspase 6, caspase7) that perform the actual cellular degradation (Danialand Korsmeyer 2004). Effector caspases can beactivated through extrinsic and intrinsic pathways(Hengartner 2000). The extrinsic apoptotic signalingis initiated by the activation of death receptors presenton the cell surface, such as the Fas receptor and tumornecrosis factor receptor (Danial and Korsmeyer2004). Intrinsic pathways of caspase activation in-clude several internal cellular stimuli mediated by theendoplasmic reticulum or the mitochondria (Danialand Korsmeyer 2004). Mitochondria-mediated intrin-sic signaling pathway of apoptosis is probably themore prominent means of programmed cell death andhas been the subject of intense scientific scrutiny inthe aging community (Jeong and Seol 2008).
In the present study, decreases in DNA fragmen-tation were only observed in enalapril-treated rats,regardless of the dosage. Interestingly, the Bax-to-Bcl-2 ratio, a critical checkpoint in the apoptoticsignaling, was lowered by both enalapril and losartan.
Histopathology/group P (n=14) L (n=20) EH (n=22) EL (n=21)
Neoplastic 14 15 12* 13**
Pituitary adenoma 2 4 2 3
Adenocarcinoma 4 4 2 2
Pheochromocytoma 3 4 3 4
Islet cell tumor, pancreas 4 1 2 1
Others 1 2 3 3
Non-neoplastic
Chronic nephropathy 9 10 15 14
Others: subcutaneous tumor, skin tumor, and tumor in reproductive system
Table 2 Number of tumorsin each treatment groups
*p=0.027 (EH vs P); ** p=0.0118 (EL vs P)
180 AGE (2011) 33:167–183

Our data do not allow inferring the mechanismswhereby the decreased Bax/Bcl-2 induced by losartandid not translate into a mitigation of muscle apoptoticDNA fragmentation. One explanation for this findingmay be that enalapril treatment, besides increasingmitochondrial Bcl-2 expression, was also able topromote Bcl-2 phosphorylation, which is requiredfor its anti-apoptotic activity (Horiuchi et al. 1997).Importantly, phosphorylation of Bcl-2 induced bymitogen-activated protein kinase (MAPK) is inhibitedby the activation of ANGII type 2 (AT2) receptor(Horiuchi et al. 1997). It is therefore conceivable thatthe reduced AT2 receptor signaling elicited byenalapril might have resulted in increased MAPKactivation and, hence, enhanced Bcl-2 phosphoryla-tion. On the other hand, the lack of AT2 receptorinhibition by losartan might have induced an incom-plete anti-apoptotic response, with increased mito-chondrial Bcl-2 levels not supported by adequate Bcl-2 phosphorylation. This possibility warrants furtherinvestigation.
In addition, although both enalapril doses wereable to reduce the extent of apoptosis in thegastrocnemius muscle, only the highest dose de-creased the activation of the mitochondrial caspase-dependent apoptotic pathway. Furthermore, neitherenalapril dose affected the caspase-independent path-way. Based on our findings, it is therefore unclearhow low-dose enalapril attenuated apoptosis. Onepossibility is that enalapril given at the lowest dosedid indeed result in downregulation of mitochondrialcaspase-dependent apoptotic signaling. However,changes in the expression levels of apoptogenicmediators might have been below the detection limitof our Western blot analysis. Alternatively, it may behypothesized that low-dose enalapril was still able toattenuate the caspase catalytic activity, possibly viaupregulation of caspase inhibitors (e.g., cIAPs andcFLIPs). The impact of caspase activity and caspaseinhibitors should be addressed in future studies.
In conclusion, enalapril treatment, between 20 and40 mg/kg, consistently lowers body weight in olderanimals, even when initiated late in life, with amaximal efficacy achieved after 3 months of treat-ment. This effect was not observed in losartan-treated animals, suggesting that blocking the AT1receptor pathway does not mediate these benefits.Changes in body weight and composition appeardissociated from improvements in physical function
and may reflect a differential impact of enalapriland losartan on muscle quality. These data suggestthat attenuation of the severity of skeletal muscleapoptosis promoted by enalapril may represent adistinct mechanism through which this compoundimproves muscle strength/quality.
Acknowledgments This study was supported by the NationalInstitute on Aging Grant AG24526, the University of FloridaInstitute on Aging, The Claude D. Pepper Older AmericansIndependence Center Grant NIH P30 AG028740, and theMcKnight Foundation.
References
Bahi L, Koulmann N, Sanchez H, Momken I, Veksler V, BigardAX et al (2004) Does ACE inhibition enhance enduranceperformance and muscle energy metabolism in rats? JAppl Physiol 96:59–64
Basso N, Cini R, Pietrelli A, Ferder L, Terragno NA, Inserra F(2007) Protective effect of long-term angiotensin IIinhibition. Am J Physiol Heart Circ Physiol 293:H1351–H1358
Benson SC, Pershadsingh HA, Ho CI, Chittiboyina A, Desai P,Pravenec M et al (2004) Identification of telmisartan as aunique angiotensin II receptor antagonist with selectivePPAR gamma-modulating activity. Hypertension 43:993–1002
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of protein-dye binding. Anal Biochem72:248–254
Carter CS, Sonntag WE, Onder G, Pahor M (2002) Physicalperformance and longevity in aged rats. J Gerontol A BiolSci Med Sci 57:B193–B197
Carter CS, Cesari M, Ambrosius WT, Hu N, Diz D, Oden S etal (2004) Angiotensin-converting enzyme inhibition, bodycomposition, and physical performance in aged rats. JGerontol A Biol Sci Med Sci 59:416–423
Carter CS, Onder G, Kritchevsky SB, Pahor M (2005)Angiotensin-converting enzyme inhibition intervention inelderly persons: effects on body composition and physicalperformance. J Gerontol A Biol Sci Med Sci 60:437–1446
Danial NN, Korsmeyer SJ (2004) Cell death: critical controlpoints. Cell 116:205–219
de Cavanagh EM, Inserra F, Ferder M, Ferder L (2007) Frommitochondria to disease: role of the renin–angiotensinsystem. Am J Nephrol 27:545–553
de Grey AD (2007) The case for prioritizing research on late-onset life-extension interventions in mammals. Rejuvena-tion Res 10:257–259
Feng X, Luo Z, Ma L, Ma S, Yang D, Zhao Z et al (2010)Angiotensin II receptor blocker telmisartan enhances runningendurance of skeletal muscle through activation of the PPARdelta/AMPK pathway. J Cell Mol Med (in press)
Foianini KR, Steen MS, Kinnick TR, Schmit MB, YoungbloodEB, Henriksen EJ (2000) Effects of exercise training and
AGE (2011) 33:167–183 181

ACE inhibition on insulin action in rat skeletal muscle. JAppl Physiol 89:687–694
Gabriely I, Barzilai N (2003) Surgical removal of visceraladipose tissue: effects on insulin action. Curr Diab Rep3:201–206
Gayagay G, Yu B, Hambly B, Boston T, Hahn A, CelermajerDS et al (1998) Elite endurance athletes and the ACE Iallele—the role of genes in athletic performance. HumGenet 103:48–50
Goodpaster BH, Carlson CL, Visser M, Kelley DE, ScherzingerA, Harris TB et al (2001) Attenuation of skeletal muscleand strength in the elderly: the health ABC Study. J ApplPhysiol 90:2157–2165
Goodpaster BH, Park SW, Harris TB, Kritchevsky SB, NevittM, Schwartz AV et al (2006) The loss of skeletal musclestrength, mass, and quality in older adults: the health,aging and body composition study. J Gerontol A Biol SciMed Sci 61:1059–1064
Guralnik JM, Simonsick EM, Ferrucci L, Glynn RJ, BerkmanLF, Blazer DG et al (1994) A short physical performancebattery assessing lower extremity function: associationwith self-reported disability and prediction of mortalityand nursing home admission. J Gerontol 49:M85–M94
Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, FlurkeyK et al (2009) Rapamycin fed late in life extends lifespan ingenetically heterogeneous mice. Nature 460:392–395
He H, Yang D, Ma L, Luo Z, Ma S, Feng X et al (2010)Telmisartan prevents weight gain and obesity throughactivation of peroxisome proliferator-activated receptor-delta-dependent pathways. Hypertension 55:869–879
Hengartner MO (2000) The biochemistry of apoptosis. Nature407:770–776
Henriksen EJ, Jacob S (2003) Modulation of metabolic controlby angiotensin converting enzyme (ACE) inhibition. J CellPhysiol 196:171–179
Horiuchi M, Hayashida W, Kambe T, Yamada T, Dzau VJ(1997) Angiotensin type 2 receptor dephosphorylates Bcl-2 by activating mitogen-activated protein kinasephosphatase-1 and induces apoptosis. J Biol Chem272:19022–19026
Ikeno Y, Bronson RT, Hubbard GB, Lee S, Bartke A (2003)Delayed occurrence of fatal neoplastic diseases in Amesdwarf mice: correlation to extended longevity. J GerontolA Biol Sci Med Sci 58:291–296
Iwasaki K, Gleiser CA, Masoro EJ, McMahan CA, Seo EJ, YuBP (1988) The influence of dietary protein source onlongevity and age-related disease processes of Fischer rats.J Gerontol 43:B5–B12
Jeong SY, Seol DW (2008) The role of mitochondria inapoptosis. BMB Rep 41:11–22
Kerr JF, Wyllie AH, Currie AR (1972) Apoptosis: a basicbiological phenomenon with wide-ranging implications intissue kinetics. Br J Cancer 26:239–257
Maeda H, Gleiser CA, Masoro EJ, Murata I, McMahan CA, YuBP (1985) Nutritional influences on aging of Fischer 344rats: II. Pathology. J Gerontol 40:671–688
Marzetti E, Groban L, Wohlgemuth SE, Lees HA, Lin M, JobeH et al (2008a) Effects of short-term GH supplementationand treadmill exercise training on physical performanceand skeletal muscle apoptosis in old rats. Am J PhysiolRegul Integr Comp Physiol 294:R558–R567
Marzetti E, Wohlgemuth SE, Lees HA, Chung HY, GiovanniniS, Leeuwenburgh C (2008b) Age-related activation ofmitochondrial caspase-independent apoptotic signaling inrat gastrocnemius muscle. Mech Ageing Dev 129:542–549
Marzetti E, Carter CS, Wohlgemuth SE, Lees HA, GiovanniniS, Anderson B et al (2009a) Changes in IL-15 expressionand death-receptor apoptotic signaling in rat gastrocnemi-us muscle with aging and life-long calorie restriction.Mech Ageing Dev 130:272–280
Marzetti E, Hwang JC, Lees HA, Wohlgemuth SE, Dupont-Versteegden EE, Carter CS et al (2009b) Mitochondrialdeath effectors: relevance to sarcopenia and disuse muscleatrophy. Biochim Biophys Acta 1800:235–244
Newman AB, Yanez D, Harris T, Duxbury A, Enright PL, FriedLP (2001) Weight change in old age and its associationwith mortality. J Am Geriatr Soc 49:1309–1318
Newman AB, Haggerty CL, Goodpaster B, Harris T, Kritch-evsky S, Nevitt M et al (2003) Strength and muscle qualityin a well-functioning cohort of older adults: the Health,Aging and Body Composition Study. J Am Geriatr Soc51:323–330
Olshansky SJ, Goldman DP, Zheng Y, Rowe JW (2009) Agingin America in the twenty-first century: demographicforecasts from the MacArthur Foundation Research Net-work on an Aging Society. Milbank Q 87:842–862
Onder G, Penninx BW, Balkrishnan R, Fried LP, Chaves PH,Williamson J et al (2002) Relation between use ofangiotensin-converting enzyme inhibitors and musclestrength and physical function in older women: anobservational study. Lancet 359:926–930
Rae MJ, Butler RN, Campisi J, de Grey AD, Finch CE,Gough M et al (2010) The demographic and biomedicalcase for late-life interventions in aging. Sci Transl Med2:40cm21
Rice KM, Linderman JK, Kinnard RS, Blough ER (2005) TheFischer 344/NNiaHSd X Brown Norway/BiNia is a bettermodel of sarcopenia than the Fischer 344/NNiaHSd: acomparative analysis of muscle mass and contractileproperties in aging male rat models. Biogerontology6:335–343
Santos EL, de Picoli SK, da Silva ED, Batista EC, Martins PJF,D'Almeida V et al (2009) Long term treatment with ACEinhibitor enalapril decreases body weight gain and increaseslife span in rats. Biochem Pharmacol 78:951–958
Scarpace PJ, Zhang Y (2009) Leptin resistance: a predisposingfactor for diet-induced obesity. Am J Physiol Regul IntegrComp Physiol 296:R493–R500
Schwartz RS (1998) Obesity in the elderly. In: Bray GA,Bouchard C, James WPT (eds) Handbook of obesity.Marcel Dekker, New York, pp 103–114
Sipahi I, Debanne SM, Rowland DY, Simon DI, Fang JC(2010) Angiotensin-receptor blockade and risk of cancer:meta-analysis of randomised controlled trials. LancetOncol 11:627–636
Song W, Kwak HB, Lawler JM (2006) Exercise trainingattenuates age-induced changes in apoptotic signaling inrat skeletal muscle. Antioxid Redox Signal 8:517–528
Visser M, Langlois J, Guralnik JM, Cauley JA, KronmalRA, Robbins J et al (1998) High body fatness, but notlow fat-free mass, predicts disability in older men and
182 AGE (2011) 33:167–183

women: the Cardiovascular Health Study. Am J ClinNutr 68:584–590
Weisinger RS, Begg DP, Jois M (2009a) Antagonists of therenin–angiotensin system and the prevention of obesity.Curr Opin Investig Drugs 10:1069–1077
Weisinger RS, Stanley TK, Begg DP, Weisinger HS, Spark KJ,Jois M (2009b) Angiotensin converting enzyme inhibitionlowers body weight and improves glucose tolerance in
C57BL/6 J mice maintained on a high fat diet. PhysiolBehav 98:192–197
Yu BP, Masoro EJ, Murata I, Bertrand HA, Lynd FT (1982)Life span study of SPF Fischer 344 male rats fed adlibitum or restricted diets: longevity, growth, lean bodymass and disease. J Gerontol 37:130–141
Zhang Y, Scarpace PJ (2006) The role of leptin in leptinresistance and obesity. Physiol Behav 88:249–256
AGE (2011) 33:167–183 183





























