Embed Size (px)
Citation preview

of February 25, 2016.This information is current as
Responses in MicrogliaReceptor-Mediated Inhibition of TLR
2AReceptors Controls Adenosine A3Differential Expression of Adenosine A
Kondova, Ad P. IJzerman and Jeffrey J. BajramovicStraalen, Eveline D. de Geus, Leonie A. Boven, Ivanela Céline van der Putten, Ella A. Zuiderwijk-Sick, Linda van
http://www.jimmunol.org/content/182/12/7603doi: 10.4049/jimmunol.0803383
2009; 182:7603-7612; ;J Immunol
MaterialSupplementary
htmlhttp://www.jimmunol.org/content/suppl/2009/06/02/182.12.7603.DC1.
Referenceshttp://www.jimmunol.org/content/182/12/7603.full#ref-list-1
, 18 of which you can access for free at: cites 49 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 25, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

Differential Expression of Adenosine A3 Receptors ControlsAdenosine A2A Receptor-Mediated Inhibition of TLRResponses in Microglia
Celine van der Putten,* Ella A. Zuiderwijk-Sick,* Linda van Straalen,* Eveline D. de Geus,‡
Leonie A. Boven,‡ Ivanela Kondova,† Ad P. IJzerman,§ and Jeffrey J. Bajramovic1*
Microglia activation is a prominent feature in many neuroinflammatory disorders. Unrestrained activation can generate a chronicinflammatory environment that might lead to neurodegeneration and autoimmunity. Extracellular adenosine modulates cellularactivation through adenosine receptor (ADORA)-mediated signaling. There are four ADORA subtypes that can either increase(A2A and A2B receptors) or decrease (A1 and A3 receptors) intracellular cyclic AMP levels. The expression pattern of the subtypesthus orchestrates the cellular response to extracellular adenosine. We have investigated the expression of ADORA subtypes inunstimulated and TLR-activated primary rhesus monkey microglia. Activation induced an up-regulation of A2A and a down-regulation of A3 receptor (A3R) levels. The altered ADORA-expression pattern sensitized microglia to A2A receptor (A2AR)-mediated inhibition of subsequent TLR-induced cytokine responses. By using combinations of subtype-specific agonists andantagonists, we revealed that in unstimulated microglia, A2AR-mediated inhibitory signaling was effectively counteracted byA3R-mediated signaling. In activated microglia, the decrease in A3R-mediated signaling sensitized them to A2AR-mediated in-hibitory signaling. We report a differential, activation state-specific expression of ADORA in microglia and uncover a role for A3Ras dynamically regulated suppressors of A2AR-mediated inhibition of TLR-induced responses. This would suggest exploration ofcombinations of A2AR agonists and A3R antagonists to dampen microglial activation during chronic neuroinflammatoryconditions. The Journal of Immunology, 2009, 182: 7603–7612.
M icroglia are macrophage-like resident cells of thebrain. Under inflammatory conditions, microglia be-come activated, as indicated by an altered morphol-
ogy, the production of inflammatory mediators, and by the in-creased expression of molecules involved in Ag presentation (1,2). Activated microglia are a prominent feature in a wide variety ofneuroinflammatory disorders where they are attributed roles asAPCs as well as effector cells.
TLR are part of the innate immune system and recognize patho-gen-associated molecular patterns (3, 4). To date, ten members ofthe TLR family have been described for human and nonhumanprimates (5). Ligand binding to TLR leads to cellular activation byactivation of the transcription factor NF-�B, which in turn inducesthe production of cytokines such as TNF-� and IL-12 (4, 6–8).Human microglia express mRNA for a broad variety of TLR, andactivation of rhesus monkey microglia via TLR1/2, -2/6, -3, -4, -5and -8 was recently described to trigger a rapid inflammatory re-sponse (9–12). Such a response must be tightly regulated becauseunrestrained TLR signaling can generate a chronic inflammatoryenvironment that might lead to neurodegeneration.
Several studies have shown that extracellular adenosine modu-lates TLR-mediated activation through G-protein coupled, adeno-
sine receptor (ADORA)2-mediated signaling. Documented effectsof adenosine include the inhibition of LPS-induced production ofproinflammatory cytokines such as TNF-� and IL-12 by macro-phages and monocytes and the facilitated production of the anti-inflammatory cytokine IL-10 by LPS-treated macrophages (13–15). During periods of high metabolic activity such as ischemiaand inflammation, extracellular levels of adenosine are stronglyincreased, possibly forming an endogenous brake on inflammation(16, 17).
The cellular response to extracellular adenosine is orchestratedby the expression pattern of four different ADORA subtypes: A1,A2A, A2B, and A3, which are characterized by their capacity toeither increase or decrease intracellular cAMP levels (16). A1 andA3 receptors (A1R and A3R) are coupled to Gi protein signalingand mediate biological effects opposite to A2A and A2B receptors(A2AR and A2BR), which are coupled to Gs protein signaling. Ro-dent microglia have been reported to express functional A1, A2A,and A3, but not A2B receptors (18), whereas studies detailingADORA expression profiles on human or nonhuman primate mi-croglia are currently lacking. In addition, little is known about thedynamics of ADORA expression after TLR-mediated activationand the consequences thereof during chronic inflammatoryconditions.
*Alternatives Unit and †Animal Science Department, Biomedical Primate ResearchCentre, Rijswijk, The Netherlands; ‡Department of Immunology, Erasmus MC, Rot-terdam, The Netherlands; and §Division of Medicinal Chemistry, Leiden/AmsterdamCenter for Drug Research, Leiden, The Netherlands
Received for publication October 9, 2008. Accepted for publication April 14, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Jeffrey J. Bajramovic, Alterna-tives Unit, Biomedical Primate Research Centre, Lange Kleiweg 139, 2280 GH Ri-jswijk, The Netherlands. E-mail address: [email protected]
2 Abbreviations used in this paper: ADORA, adenosine receptor; A1R, A1 receptor;A3R, A3 receptor; A2AR, A2A receptor; A2BR, A2B receptor; CGS21680, 2-p-(2-carboxyethyl)phenethylamino-5�-N-ethylcarboxamidoadenosine hydrochloride;DPCPX, 8-cyclopentyl-1,3-dipropylxanthine; I�B, inhibitor of NF-�B proteins;NECA, 5�-(N-ethylcarboxamido)adenosine; SCH58261, 7-(2-phenylethyl)-5-amino-2-(2-furyl)-pyrazolo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine; VUF5574, N-(2-me-thoxyphenyl)-N�-[2-(3-pyridinyl)-4-quinazolinyl]-urea.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0803383
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

Using primary rhesus monkey microglia, we characterized theexpression of ADORA subtypes in unstimulated and TLR-acti-vated microglia. Microglia simultaneously up-regulated A2AR anddown-regulated A3R expression levels upon TLR-mediated acti-vation. As a consequence, ADORA-mediated inhibition of subse-quent TLR-induced TNF-� and IL-12p40/p70 production wasmuch more potent in activated than in unstimulated microglia. Thiscould be attributed to an increase in A2AR-mediated signaling andinvolved suppression of NF-�B activation. By using combinationsof subtype-specific agonists and antagonists, we revealed that inunstimulated microglia, A2AR-mediated inhibitory signaling waseffectively counteracted by A3R-mediated signaling. In activatedmicroglia, the decrease in A3R-mediated signaling shifted the bal-ance toward A2AR-mediated signaling, sensitizing the cells to in-hibitory signaling. We report a differential, activation state-specificexpression of ADORA in microglia and uncover a role for A3R asdynamically regulated suppressors of A2AR-mediated inhibition ofTLR-induced responses. These findings contribute significantly toour understanding of how inhibitors of cellular activation are reg-ulated themselves, and might have relevance for therapeutic inter-vention in neuroinflammatory disorders where chronic microgliaactivation often is an unwanted feature.
Materials and MethodsAnimals
Adult brain donor rhesus monkeys (Macaca mulatta) without neurologicaldisease became available from the outbred breeding colony; no monkeyswere killed for the exclusive purpose of primary cell culture initiation.Individual details are listed in Supplementary Table I.3 Better use of ex-perimental animals contributes to the active refinement program within theBiomedical Primate Research Centre (Rijswijk, The Netherlands).
Reagents
The nonspecific adenosine receptor agonist 5�-(N-ethylcarboxamido)ad-enosine (NECA), adenosine A2AR agonist 2-p-(2-carboxyethyl)phenethyl-amino-5�-N-ethylcarboxamidoadenosine hydrochloride (CGS21680),adenosine A1R antagonist 8-cyclopentyl-1,3-dipropylxanthine (DPCPX),adenosine A2AR antagonist 7-(2-phenylethyl)-5-amino-2-(2-furyl)-pyra-zolo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine (SCH58261), adenosine A2BRantagonist benzo[g]pteridine-2,4(1H,3H)-dione (alloxazine) and the aden-osine A3R antagonist N-(2-methoxyphenyl)-N�-[2-(3-pyridinyl)-4-quina-zolinyl]-urea (VUF5574; all purchased from Sigma-Aldrich), were dilutedin DMSO, aliquotted and stored at �20°C. DMSO controls were includedin the functional assays.
TLR-ligands used were E. coli O26:B6 LPS (Sigma-Aldrich),Pam3CSK4, poly(I:C), Ultrapure LPS, Flagellin, FSL-1, R848, and CL075(Invivogen). During the course of this study, the distribution of R848 wasdiscontinued and replaced by CL075, a TLR8 agonist with similarproperties.
Primary cell isolation and culture
Primary microglia were obtained from adult rhesus monkeys at necropsy asdescribed previously (12). Briefly, tissue samples from prefrontal subcor-tical white matter were collected, divided into cubes of �3 g, and meningesand visible blood vessels were removed before mincing the tissue intocubes of less than 2 mm3. Tissue fragments were incubated at 37°C for 20min in HBSS (�8 ml/g tissue) containing 0.25% w/v porcine trypsin (Sig-ma-Aldrich), 0.2 mg/ml EDTA, 1 mg/ml glucose, and 0.1 mg/ml bovinepancreatic DNase I (Sigma-Aldrich). The supernatant (no centrifugation)was discarded and the pellet was resuspended in microglia medium, i.e.,1:1 v/v DMEM (high glucose)/HAMF10 (with L-glutamine) with 10% v/vFCS and antibiotic supplement (penicillin 100 U/ml and streptomycin 0.1mg/ml) (all purchased from Life Technologies), washed once, and passedthrough a 100-�m nylon cell strainer (BD Biosciences). Following cen-trifugation at 300 � g for 7 min, the resuspended pellet was subjected toPercoll gradient centrifugation and hypotonic shock to remove erythro-cytes. Cells were plated at a density of 2.2–2.5 � 105/ml in tissue culture-treated 6- or 24-well plates (Corning Costar). After 24 h incubation at 37°Cin a humidified atmosphere containing 5% CO2, unattached cells and my-elin debris were removed by washing and replaced by microglia mediumsupplemented with �4 U recombinant human M-CSF/ml (PeproTech).Half of the medium was replaced by fresh medium containing new growthfactors every 3–4 days.
RT-PCR and quantitative RT-PCR
Total cellular RNA was isolated using TriReagent (Sigma-Aldrich) accord-ing to manufacturer’s protocol. Subsequently, mRNA was reversely tran-scribed into cDNA using the Omniscript Reverse Transcription Systemaccording to manufacturer’s protocol (Qiagen) using 0.5 �g RNA as tem-plate and 0.25 �g oligo(dT)15 primers (Promega). RT-PCR were performedon the iCycler Thermal cycler (Bio-Rad). mRNA levels of ADORA sub-types and GAPDH as reference genes were determined by real-time quan-titative RT-PCR. Reactions were performed using the ABI PRISM 7700sequence detection system (Applied Biosystems) and the iQ5 multicolorPCR detection system (Bio-Rad). Sequences of primer and probe (Probe-Library, Roche) combinations are listed in Table I. ADORA mRNA ex-pression levels were standardized to GAPDH or �-actin mRNA expressionlevels using the Pfaffl method (19).
Cytokine analysis
TNF-� and IL-12p40/p70 levels were determined by ELISA according tomanufacturer’s protocol (U-CyTech Biosciences).
Western blot analysis
Protein extracts were prepared from untreated and R848-activated micro-glia as well as from A2AR-transfected human embryonic kidney 293 cells.The cells were lysed in lysis buffer (1% Triton X-100, 10 mM Tris-HCl, 50mM NaCl, 30 mM Na4P2O7, 50 mM NaF, 5 �M ZnCl2, 1 nM DTT, 100�M Na3VO4, and protease inhibitors; Roche). Lysates were heated at 85°Cfor 10 min before centrifugation at 14000 rpm for 10 min. Supernatantswere separated on 12% Bis-Tris gels in combination with MOPS bufferusing the Novex Xcell Surelock miniCell system (Invitrogen) and subse-quently transferred onto Hybond-ECL nitrocellulose membranes (Amer-sham Biosciences). The membranes were blocked for 30 min at room tem-perature in TBST containing 5% nonfat dry milk, and probed using goat3 The online version of this article contains supplemental material.
Table I. Primer and probe sequences
Target Forward Primer (5�–3�) Reverse Primer (5�–3�) Probe Amplicon Size (bp) Used in
A1R CTATGTTTGGCTGGAACAAT GCTGCTTGCGGATTAGGTAG 207 RT-PCRA2AR CTCATGCTGGGTGTCTATTT TGAAGCAGTTGATGATGTGT 193 RT-PCRA2BR ATTTCTTTGGGTGTGTTCTG ACTTGGGCTTATTTTTACCC 252 RT-PCRA3R CTGGAACATGAAACTGACCT GAGTTTGTTCCGAATGATGT 181 RT-PCR�-actin GGTCATCACCATTGGCAATGA ACGTCACACTTCATGATGGAGTTG 122 RT-PCRA1R GTCAAGATCCCTCTCCGGTA CCACCACGAAGGAGAGGA GGCTGCTG 91 Real-time RT-PCRA2AR CCAACTACTTCGTGGTGTCG GTAATGGCAAAGGGGATGG GGCGGCGG 73 Real-time RT-PCRA2BR TCTGTGTCCCGCTCAGGT GATGCCAAAGGCAAGGAC TGCTGTCC 89 Real-time RT-PCRA3R GGGTCAAGCTTACCGTCAGA ATGACACCAGCCAGCAAAG GGCCCTGG 84 Real-time RT-PCRGAPDH TCCACTGGCGTCTTCAC GGCAGATGATGACCCTTTT AGCCCCAG 78 Real-time RT-PCR
7604 DYNAMIC A3R EXPRESSION CONTROLS A2AR-MEDIATED TLR INHIBITION
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

anti-GAPDH (Imgenex) and goat anti-A2AR (Everest Biotech) Abs fol-lowed by anti-goat IgG-HRP-conjugated secondary Abs (Jackson Immu-noResearch Laboratories) for 1 h at room temperature. Blots were devel-oped using Femto ECL substrate (Pierce) and quantitated with theChemiDoc XRS system (Bio-Rad).
Immunofluorescence
Cells grown on glass coverslips were fixed for 20 min at 4°C with 2%paraformaldehyde, washed with PBS, and permeabilized for 15 min inPBS � 0.1% Triton X-100. After two washes with PBS, cells were incu-bated for 1 h at room temperature with normal rabbit serum (1 �g IgG/ml)and with rabbit polyclonal anti NF-�B p65 Abs (0.4 �g IgG/ml; SantaCruz) in PBS, washed with PBS � 0.01% Tween20, and followed by a 1 hincubation at room temperature with FITC-labeled goat anti-rabbit second-ary Abs (Jackson ImmunoResearch Laboratories). After extensive washes,coverslips were mounted using Vectashield � DAPI (Vector Laboratories,Burlingame, CA) and images were captured on a fluorescence microscope.
Flow cytometry
Microglia were harvested by incubation with 4 mg lidocaine/ml (Sigma-Aldrich) for maximally 20 min at 37°C. All following incubations were
performed on ice. Microglia were washed in FACS buffer (PBS � 2%BSA) and incubated for 10 min with 1/10 diluted Fc receptor-block (Milte-nyi Biotec) in FACS buffer, to prevent aspecific binding of the Abs. Cellswere washed and incubated for 30 min with anti-adenosine A3 receptor Abs(SP055P, Acris Antibodies) or total rabbit IgG (AbD Serotec) as an isotypecontrol. Cells were washed and incubated for 30 min with PE-labeledCD11b Abs (clone D12; BD Biosciences) and allophycocyanin-labeledsecondary anti-rabbit Abs (Jackson ImmunoResearch Laboratories). Cellswere analyzed on the LSRII and data were analyzed using FACS Divasoftware (both purchased from BD Biosciences).
Statistics
GraphPad Prism 5 (GraphPad Software) for Macintosh was used for sta-tistical analysis. Statistical analysis of the differential effect of activation onNECA-induced suppression was analyzed using linear mixed effects mod-els (20) in the R environment (21). The following model was used:
Log cytokine level � b0 � b1 � stimulus � b2 � activation � b3 �stimulus � activation,
where stimulus was coded 0 for LPS only and 1 for LPS � NECAand activation was coded 0 for unstimulated and 1 for activated micro-glia. b0 represents the log value of the intercept, which is the mean of
FIGURE 1. TLR-mediated activation alters the ADORA expression profile in primary rhesus microglia. A, Analysis of ADORA subtype mRNAexpression by RT-PCR in unstimulated and TLR-activated microglia. B, ADORA mRNA expression levels were quantitated by real-time RT-PCR inunstimulated and in TLR-activated microglia. Cells were stimulated for 16 h with 1 �g R848/ml (open symbols) or with 100 ng LPS/ml (filled symbols).Different symbols represent different donors. �, p � 0.05; Wilcoxon signed-rank test. ADORA mRNA expression levels are expressed relative to referencegene (GAPDH) mRNA expression levels. A1R mRNA expression levels were below detection limits and low levels of A2BR prohibited reliable quanti-fication of all samples. C, Western blot analysis of cell lysates of unstimulated microglia and of microglia stimulated for 16 h with 1 �g R848/ml. A2ARexpression levels were quantitated and normalized to GAPDH protein expression levels. Data are shown for three donors. D, FACS analysis of A3Rexpression levels on unstimulated microglia and microglia stimulated for 16 h with 1 �g R848/ml. The filled curves in the histogram (left) represent isotypecontrols, the open curves A3R-staining. The solid lines represent unstimulated microglia and the dashed lines represent microglia that were stimulated for16 h with 1 �g R848/ml. The MFI (mean fluorescence intensity) indices given in the right panel were calculated as follows: (MFI A3R-staining � MFIisotype control)/MFI isotype control.
7605The Journal of Immunology
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

the log cytokine production levels when stimulated with LPS only (theantilog is the geometric mean). b1 represents the difference in log cy-tokine levels when NECA is added. Thus, the antilog of b1 representsthe ratio of cytokine levels induced by LPS � NECA/LPS; b2 repre-sents the difference in intercept for unstimulated and activated micro-glia, again the antilog represents a ratio; and b3 represents the interac-tion term for activation on NECA. The antilog of b3 is the ratio ofNECA � LPS-induced cytokine production in unstimulated and acti-vated microglia.
ResultsTLR-mediated activation alters the ADORA expression profile inrhesus monkey microglia
The expression profile of ADORA subtypes in human and non-human primate microglia had not been characterized before andit was unknown whether TLR-mediated activation would affecttheir expression levels. As published recently, signaling viaTLR1/2, -2/4, -3, -5, or -8 potently activates rhesus monkeymicroglia (12). We therefore exposed adult primary rhesusmonkey microglia from two different donors to TLR8 ligandsand analyzed the expression of all ADORA subtypes. mRNAtranscripts encoding A2AR, A2BR, and A3R were readily de-tected by RT-PCR, whereas transcripts for A1R were low orbelow detection levels (Fig. 1A). After TLR8-mediated activa-tion, A2AR-encoding mRNA levels appeared up-regulated,whereas A3R-encoding mRNA levels appeared down-regulated.To quantitate ADORA subtype mRNA expression levels, weexposed microglia from different donors to TLR2/4 and TLR8ligands and determined mRNA levels by quantitative RT-PCR.Consistent with abovementioned results, A1R mRNA levels
were below detection limits in both unstimulated and TLR-ac-tivated microglia from all donors, prohibiting quantitation. Fur-thermore, comparison of the threshold cycle values suggeststhat A2AR- and A3R-encoding mRNA levels were generallyhigher than those that encoded for A2BR. TLR-mediated acti-vation of microglia resulted in an 11-fold (95% CI 7.34 –15.28,p � 0.05) increase in A2AR-encoding mRNA expression levels.By contrast, A3R-encoding mRNA expression levels were3-fold decreased (95% CI 0.14 – 0.48, p � 0.05; Fig. 1B). A2BR-encoding mRNA levels were not significantly altered afterTLR-mediated activation. The predicted relative contribution ofstimulatory G protein signaling-coupled A2AR over Gi proteinsignaling-coupled A3R to ADORA-mediated signaling wouldthereby be enhanced 30-fold in TLR-activated microglia. Thepresented effects of TLR-mediated activation on A2AR and A3RmRNA levels are in line with earlier observations in LPS-acti-vated human and mouse macrophages (22), and preliminary re-sults indicate that they hold true for primary rhesus bone mar-row-derived macrophages as well (data not shown).
To examine whether the changes in mRNA expression levelswere reflected by changes in protein levels, A2A receptor proteinexpression levels were quantified by Western blotting. Resultsdemonstrate that A2AR protein levels in TLR8-activated microgliawere increased 2- to 4-fold (Fig. 1C), consistent with our data onmRNA expression levels. A3R protein expression levels werequantified by flow cytometry analysis. These data reveal thatTLR8-mediated activation indeed resulted in decreased A3R pro-tein expression levels on primary microglia (Fig. 1D).
FIGURE 2. ADORA-mediated in-hibition of LPS-induced TNF-� andIL-12p40/p70 is enhanced in acti-vated microglia. Unstimulated andactivated microglia were exposed to100 ng LPS/ml in the presence or ab-sence of 10 �M NECA. After 16 h,TNF-� (A) and IL-12p40/p70 (B) lev-els were measured in the culture su-pernatants using ELISA. �, p � 0.05;Student’s t test. TNF-� (C) and IL-12p40/p70 (D) production after expo-sure to 100 ng LPS/ml � 10 �MNECA is expressed relative to pro-duction after exposure to 100 ngLPS/ml alone for unstimulated andactivated microglia of six differentdonors. TNF-� (E) and IL-12p40/p70(F) production after exposure to 100ng LPS/ml � different concentrationsof NECA is expressed relative to pro-duction after exposure to 100 ngLPS/ml alone for unstimulated andactivated microglia. �, p � 0.05;2-way ANOVA. The data are repre-sentative for three independent exper-iments using different donors.
7606 DYNAMIC A3R EXPRESSION CONTROLS A2AR-MEDIATED TLR INHIBITION
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

ADORA-mediated inhibition of LPS-induced TNF-� andIL-12p40/p70 production is enhanced in activatedmicroglia
Adenosine-induced signaling through ADORA can modulateLPS-induced production of TNF-� and IL-12 (13, 23, 24). Toassess the functional relevance of the altered ADORA expres-sion profile on TLR-activated microglia, we used the followingexperimental set-up. Microglia were either unstimulated or ex-posed for 16 h to TLR8-mediated activation, yielding activatedmicroglia. Both populations were washed, rested for 4 h,washed again, and subsequently stimulated with LPS in thepresence or absence of NECA, a potent nonselective ADORAagonist that is more stable than adenosine. After 16 h, TNF-�and IL-12p40/p70 levels in the culture supernatants were mea-sured by ELISA.
Both unstimulated and activated microglia produced compara-ble amounts of TNF-� and IL-12p40/p70 upon stimulation withLPS (Fig. 2, A and B). The addition of NECA alone did not induceTNF-� or IL-12p40/p70 production in either unstimulated or ac-tivated microglia (data not shown). In unstimulated microglia,NECA inhibited the LPS-induced production of TNF-� and IL-12p40/p70 to 86 and 78%, respectively of control levels. Remark-ably, in activated microglia, NECA inhibited the LPS-induced pro-duction of TNF-� and IL-12p40/p70 to 9 and 39%, respectively ofcontrol levels (Fig. 2, A and B). Analysis of six additional donorsconfirmed that there were no significant differences in LPS-in-duced production of TNF-� or IL-12p40/p70 between unstimu-lated or activated microglia ( p � 0.39 and p � 0.47, respectively).As expected, the effects of NECA on LPS-induced TNF-� pro-duction differed significantly between unstimulated and activatedmicroglia (0.125-fold of unstimulated, 95% CI 0.061–0.255, p �0.001; Fig. 2C). In addition, inhibition of LPS-induced IL-12p40/p70 production by NECA was stronger in activated than in un-
stimulated microglia in all donors tested (0.448-fold of unstimu-lated, 95% CI 0.157–1.279, p � 0.123; Fig. 2D), but due to asmaller effect of activation and higher within-donor variability inIL-12p40/p70 production levels this difference was not statisticallysignificant.
To better characterize the ADORA-mediated inhibitory ef-fects, we measured LPS-induced cytokine production in thepresence of different concentrations of NECA. Under nonsat-urating conditions (NECA �1 �M), inhibition of LPS-inducedTNF-� and IL-12p40/p70 was stronger in activated microgliathan in unstimulated microglia at all concentrations used (Fig.2, E and F). LPS-induced cytokine production in activated mi-croglia was already inhibited by 0.02 �M NECA, whereas thisconcentration increased LPS-induced cytokine production inunstimulated microglia. Under saturating conditions (NECA�5 �M), inhibition of LPS-induced TNF-� and IL-12p40/p70in unstimulated microglia did not attain the same level as inactivated microglia (Fig. 2, E and F). Activated microglia werethus not only sensitized to ADORA-mediated inhibition of LPS-induced cytokine production, the potency of ADORA-mediatedinhibition was enhanced as well.
Enhanced inhibition of LPS-induced TNF-� production israpidly gained and lost in activated microglia, whereasinhibition of IL-12p40/p70 production remains enhanced for upto 72 h
Whereas it is conceivable that microglia receive multiple TLRstimuli during inflammatory conditions in vivo, the timing of suchstimuli will be variable. To investigate how fast after activationmicroglia became sensitized to ADORA-mediated inhibition, wevaried the duration of the activation period. Interestingly, therewas a discordance between inhibition of LPS-induced TNF-� andIL-12p40/p70 production. Where LPS-induced TNF-� production
FIGURE 3. Kinetics of the enhanced inhibitory phenotype of activated microglia. Microglia were either unstimulated or activated by exposure to 1 �gR848/ml for 2, 4, 8, or 16 h. After extensive washing and a 4-h recovery period, microglia were exposed to 100 ng LPS/ml in the presence or absence of10 �M NECA. After 16 h, TNF-� (A) and IL-12p40/p70 (B) levels were measured and expressed as relative values of exposure to 100 ng LPS/ml alone.Microglia were either unstimulated or activated by exposure to 1 �g R848/ml for 16 h. After extensive washing, microglia were allowed to recover for 4,8, 24, 48, or 72 h before exposure to 100 ng LPS/ml in the presence or absence of 10 �M NECA. After 16 h, TNF-� (C) and IL-12p40/p70 (D) levelswere measured and expressed as relative values of exposure to 100 ng LPS/ml alone. �, p � 0.05; Student’s t test. The data are representative for threeindependent experiments using different donors.
7607The Journal of Immunology
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

was already stronger when inhibited by NECA in microglia thatwere activated for 2 h, enhanced inhibition of IL-12p40/p70 pro-duction was observed only after activation for 8 h (Fig. 3Aand B).
Next we investigated how long activated microglia remainedsensitized by varying the duration between activation and LPSexposure. Again there was a discordance between the inhibitionof LPS-induced TNF-� and IL-12p40/p70 production. NECA-mediated inhibition of TNF-� production was comparable inunstimulated and activated microglia already within 8 h after
activation, whereas NECA-mediated inhibition of IL-12p40/p70 production remained enhanced for up to 72 h after activa-tion (Fig. 3, C and D).
Important roles for A2AR as well as for A3R
Although the altered ADORA expression pattern on activated mi-croglia was likely correlated to the enhanced inhibitory phenotype,the contribution of individual ADORA subtypes remained unclear.Because selective agonists for all four ADORA subtypes are not
FIGURE 4. The enhanced inhibitory effect of NECA is mediated by the enhanced contribution of A2AR-mediated signaling and the decreasedcontribution of A3R-mediated signaling. Unstimulated and activated microglia were exposed to 100 ng LPS/ml in the presence or absence of 10 �MNECA in combination with A1, A2A, A2B, and A3 receptor antagonists. After 16 h, TNF-� (A) and IL-12p40/p70 (B) levels were measured andexpressed as relative values of exposure to 100 ng LPS/ml alone. DPCPX was used as an A1R, SCH58261 as an A2AR, alloxazine as an A2BR andVUF5574 as an A3R antagonist. All antagonists were used at 10 �M concentrations. Solid lines in the graph indicate relative cytokine productionlevels of exposure to 100 ng LPS/ml � 10 �M NECA without antagonists of unstimulated microglia, the dashed lines indicate activated microglia.�, p � 0.05; Student’s t test. TNF-� (C) and IL-12p40/p70 (D) production after exposure to 100 ng LPS/ml in the presence or absence of 10 �MCGS21680, an A2AR-selective agonist, and/or of 10 �M VUF5574, an A3R antagonist. Cytokine production is expressed relative to production afterexposure to 100 ng LPS/ml alone for unstimulated and activated microglia. �, p � 0.05; Student’s t test. TNF-� (E) and IL-12p40/p70 (F) productionafter exposure to 100 ng LPS/ml � different concentrations of CGS21680, in the presence or absence of A3R antagonist, is expressed relative toproduction after exposure to 100 ng LPS/ml alone for unstimulated and activated microglia. All data are representative for three independentexperiments using different donors.
7608 DYNAMIC A3R EXPRESSION CONTROLS A2AR-MEDIATED TLR INHIBITION
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

available, we used the nonselective agonist NECA in combinationwith subtype-selective antagonists to address this issue.
Addition of an A1R antagonist slightly enhanced the inhibitorypotential of NECA in unstimulated microglia as measured by theeffect on LPS-induced TNF-� and IL-12p40/p70 production (Fig.4, A and B). Although analysis of A1R mRNA expression levelsindicate that they were low at best (Fig. 1A), A1R expression levelsappear sufficient to affect LPS-induced cytokine production in un-stimulated microglia. In activated microglia however, NECA-me-diated inhibition of LPS-induced cytokine production was similarin the presence and absence of the antagonist, indicating that A1Rare not responsible for the enhanced inhibition. Addition of anA2AR antagonist did not affect the inhibitory effect of NECA onLPS-induced TNF-� production in unstimulated microglia, but didannul NECA-mediated inhibition of IL-12p40/p70. In activatedmicroglia, NECA-mediated inhibition of both TNF-� and IL-12p40/p70 production was completely abrogated, revealing an im-portant role for A2AR. Addition of an A2BR antagonist did notaffect the inhibitory effect of NECA on LPS-induced cytokine pro-duction in either unstimulated or activated microglia, indicatingthat the role of A2BR in the inhibition of LPS-induced cytokineproduction is probably limited. Most interestingly, addition of anA3R antagonist enhanced the inhibitory effect of NECA in both
unstimulated and activated microglia, in line with the idea thatA2AR-mediated signaling generates opposite effects from A3R-me-diated signaling.
To further investigate the contribution of A2AR and A3R, we per-formed experiments using A2AR-specific agonists (CGS21680), A3Rantagonists (VUF5574), and a combination of both. Addition ofCGS21680 alone mimicked our earlier observations using NECA,confirming an important role for A2AR-mediated signaling in ac-tivated microglia (Fig. 4, C and D). Addition of A3R antagonistalone inhibited LPS-induced cytokine responses in both nonacti-vated and activated microglia. Because no exogenous adenosinewas added, the inhibitory effects are most likely attributable toendogenously produced extracellular adenosine in response to LPS(25–27) and demonstrate that A3R-mediated signaling indeedcounteracts adenosine-mediated inhibitory signaling. When A2ARagonists were added in combination with A3R antagonists, inhibi-tion was maximal in both nonactivated and activated microglia,confirming this idea.
Additional experiments using different concentrations ofCGS21680 either alone or in combination with A3R antagonistsindicate that the contribution of A3R-mediated signaling is moreimportant in unstimulated microglia than in activated microglia
FIGURE 5. The enhanced inhibitory phenotype of activated microglia inhibits proinflammatory cytokine production induced via suppression of NF-�Bactivation. A, Unstimulated and activated microglia were exposed to 100 ng LPS/ml in the presence or absence of 10 �M NECA for 1 h. Binding of normalrabbit serum and of Abs recognizing NF-�B p65 was visualized by FITC-conjugated anti-rabbit Ab (left panels). Cell nuclei were visualized using4�,6-diamidino-2-phenylindole (DAPI; right panels). Original magnifications, �800 (upper panels) and �400 (lower panels). B, Quantification of theintracellular localization of NF-�B. Intracellular localization was scored as exclusively cytosolic, cytosolic and nuclear or exclusively nuclear. Total cellnumbers were determined by counting DAPI-stained nuclei and data are represented as a percentage of total cell numbers. A minimum of 500 cells pervariable were counted.
7609The Journal of Immunology
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

(Fig. 4, E and F). In addition, they indicate that, even in the ab-sence of A3R-mediated signaling, activated microglia are moresensitive to A2AR-mediated inhibition and are more potently in-hibited at saturating concentrations than unstimulated microglia.
Together, these results demonstrate that the potential to inhibitLPS-induced cytokine production via A2AR is already present inunstimulated microglia to a considerable extent, but counteractedby A3R-mediated signaling. Combined with our data on ADORAexpression levels, these data are most consistent with the idea thatthe simultaneous up-regulation of A2AR and down-regulation ofA3R sensitize activated microglia to ADORA-mediated inhibitorysignaling.
Cross-inhibition of proinflammatory cytokine production inducedby different TLR involves suppression of NF-�B activation
Next we investigated whether the altered ADORA profile on ac-tivated microglia also resulted in a stronger inhibition of cytokineproduction induced by other TLR ligands. We measured cytokineproduction induced by TLR1/2, -2/6, -2/4, -3, and -4-mediatedactivation and all these responses were stronger when inhibited byNECA in activated microglia than in unstimulated microglia (Sup-plementary Fig. 1).
Because activation of microglia enhanced the ADORA-medi-ated inhibition of cytokine production as induced by a variety ofTLR ligands, the mechanism is likely to involve a shared elementof TLR-mediated signaling. All TLR-induced intracellular signal-ing cascades finally lead to the activation of NF-�B, a transcriptionfactor that controls among others. The mRNA transcription levelsof TNF-� and IL-12p40/p70, followed by its translocation fromthe cytosol to the nucleus. We therefore evaluated the effect ofADORA-mediated signaling on TLR-induced NF-�B transloca-tion in rhesus microglia by using immunofluorescence (Fig. 5A).Our results demonstrate that exposure of microglia to LPS is fol-lowed by a rapid translocation of NF-�B from the cytosol to thenucleus. Simultaneous triggering of ADORA-mediated signalingby the addition of NECA partially blocked translocation of NF-�B,as exemplified by the enhanced proportion of cells where NF-�Bwas localized in the cytosol as well as in the nucleus. Quantitativeanalysis of the images (Fig. 5B) demonstrates that NECA inhibitedthe LPS-induced nuclear translocation of NF-�B in both unstimu-lated as well as in activated microglia. Exposure of unstimulatedand activated microglia to LPS resulted in 81 and 80%, respec-tively of total cell numbers with an exclusive nuclear localizationof NF-�B. Addition of NECA decreased these percentages to 54%in unstimulated microglia and to 23% in activated microglia, inline with the enhanced inhibition in activated microglia.
DiscussionIn this study we explored how ADORA-mediated signaling mod-ulates microglial innate immune responses during chronic inflam-matory conditions. To do so, we characterized the ADORA ex-pression profile of primary adult rhesus monkey microglia beforeand after TLR-mediated activation. We show that activation altersthe ADORA expression pattern and that ADORA-mediated inhi-bition of LPS-induced TNF-� and IL-12p40/p70 is much strongerin activated than in unstimulated microglia. We demonstrate thatboth enhanced A2AR-mediated signaling as well as decreasedA3R-mediated signaling are responsible for this effect. Whereasthe inhibitory potential of A2AR-mediated signaling on LPS-in-duced cytokine production is widely acknowledged (13, 14, 16, 18,23, 28–32), the dynamically regulated counteracting role of A3Ris new.
Our data were obtained by using combinations of agonists andantagonists and show that even on unstimulated microglia the ex-
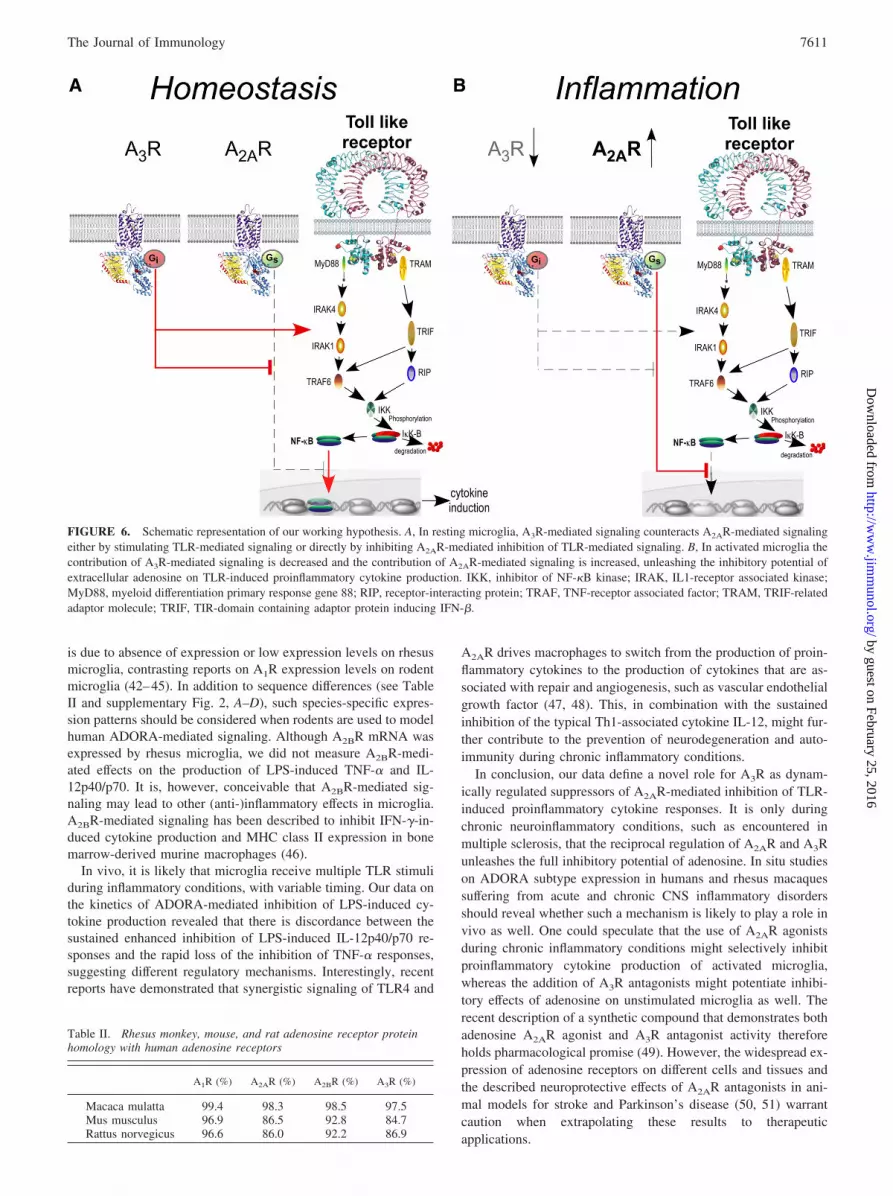
pression levels of A2AR are sufficient to inhibit LPS-induced cy-tokine levels to a considerable extent. However, it is only uponTLR-mediated activation that the full inhibitory capacity of A2ARis unleashed and this is primarily due to the decreased contributionof A3R-mediated signaling in the response to adenosine. Becausethese effects were also found when saturating amounts of NECAwere used, they cannot be explained by mere competition ofADORAs for available NECA. Based on our data, we propose amodel in which, under homeostatic conditions, A2AR-mediated in-hibitory responses are kept under tight control by A3R-mediated,counter-regulatory mechanisms. When microglia have become ac-tivated and inhibition has become necessary, A2AR-mediated in-hibitory responses are rapidly unleashed by removal of A3R. It isunclear yet whether A3R-mediated signaling suppresses A2AR-me-diated inhibition of TLR-induced cytokine responses by directlyinhibiting A2AR-mediated signaling or rather by stimulating TLR-induced signaling (Fig. 6). A3R-mediated signaling has been re-ported to induce p38 MAPK and ERK1/2 phosphorylation in pri-mary murine microglia and in the N13 microglial cell line (33, 34),while other studies have reported on A3R-mediated suppression ofLPS-induced TNF-� in murine BV2 microglial cells (24) and LPS-induced IL-12 in human monocytes (35). Biochemical studies us-ing reporter cell lines transfected with different ADORAs mightreveal possible interactions between A2AR- and A3R-mediated sig-naling but are currently lacking. Although our data strongly sug-gest that the changes in ADORA expression patterns are respon-sible for the enhanced inhibition of TLR-induced cytokineproduction in activated microglia, we cannot exclude the possibil-ity that other effects, such as altered coupling of ADORAs to in-tracellular signaling cascades (36, 37), TNF-�-induced preventionof A2AR desensitization (38), or the altered expression of down-stream signaling elements (39), play a role as well.
Different groups have reported that A2AR-mediated signalingcan suppress TLR-mediated activation of the NF-�B pathway andwe demonstrate that the A2AR-mediated inhibitory effects on LPS-induced cytokine production in primate microglia involvessuppression of NF-�B activation as well. Although we describedifferent effects of adenosine receptor-mediated signaling on LPS-induced TNF-� and IL-12p40/p70 production, both in kinetics andin strength of the inhibitory effect, this dissociation is not incon-sistent with our description of NF-�B signaling as a likely targetfor A2AR-mediated signaling. It would rather indicate that, in ad-dition to NF-�B, there are other factors that differentially modulatethe synthesis and secretion of TNF-� and IL-12 (e.g., AP-1). Themolecular basis for the suppression of NF-�B activation appears tobe cell type-specific. In rat glioma cells, A2AR-mediated signalingblocked the phosphorylation and subsequent degradation of inhib-itor of NF-�B proteins (I�B) in contrast to the I�B-independentsuppression of the NF-�B pathway in vascular endothelial cells(40). In addition, recent reports have shown that ADORA-medi-ated signaling can inactivate the ubiquitination of I�B by promot-ing the deneddylation of cullin-1 in lung tissue from mice thatwere exposed to hypoxia (41). Preliminary experiments using p38MAPK inhibitors, MAPK/ERK kinase inhibitors, and protein ki-nase A inhibitors to assess the relative importance of these path-ways for A2AR-mediated suppression of NF-�B activation in rhe-sus microglia have thus far yielded inconclusive results (ourunpublished observations), suggesting the involvement of redun-dant, multiple, or other signaling cascades in microglia.
Whereas both A2AR- and A3R-mediated signaling contribute tothe effects of adenosine on LPS-induced cytokine responses, wefound little evidence for contributions of A1R- or A2BR-mediatedsignaling. Our results on A1R expression levels are most consistentwith the idea that the limited contribution of A1R-mediated effects
7610 DYNAMIC A3R EXPRESSION CONTROLS A2AR-MEDIATED TLR INHIBITION
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
is due to absence of expression or low expression levels on rhesusmicroglia, contrasting reports on A1R expression levels on rodentmicroglia (42–45). In addition to sequence differences (see TableII and supplementary Fig. 2, A–D), such species-specific expres-sion patterns should be considered when rodents are used to modelhuman ADORA-mediated signaling. Although A2BR mRNA wasexpressed by rhesus microglia, we did not measure A2BR-medi-ated effects on the production of LPS-induced TNF-� and IL-12p40/p70. It is, however, conceivable that A2BR-mediated sig-naling may lead to other (anti-)inflammatory effects in microglia.A2BR-mediated signaling has been described to inhibit IFN-�-in-duced cytokine production and MHC class II expression in bonemarrow-derived murine macrophages (46).
In vivo, it is likely that microglia receive multiple TLR stimuliduring inflammatory conditions, with variable timing. Our data onthe kinetics of ADORA-mediated inhibition of LPS-induced cy-tokine production revealed that there is discordance between thesustained enhanced inhibition of LPS-induced IL-12p40/p70 re-sponses and the rapid loss of the inhibition of TNF-� responses,suggesting different regulatory mechanisms. Interestingly, recentreports have demonstrated that synergistic signaling of TLR4 and
A2AR drives macrophages to switch from the production of proin-flammatory cytokines to the production of cytokines that are as-sociated with repair and angiogenesis, such as vascular endothelialgrowth factor (47, 48). This, in combination with the sustainedinhibition of the typical Th1-associated cytokine IL-12, might fur-ther contribute to the prevention of neurodegeneration and auto-immunity during chronic inflammatory conditions.
In conclusion, our data define a novel role for A3R as dynam-ically regulated suppressors of A2AR-mediated inhibition of TLR-induced proinflammatory cytokine responses. It is only duringchronic neuroinflammatory conditions, such as encountered inmultiple sclerosis, that the reciprocal regulation of A2AR and A3Runleashes the full inhibitory potential of adenosine. In situ studieson ADORA subtype expression in humans and rhesus macaquessuffering from acute and chronic CNS inflammatory disordersshould reveal whether such a mechanism is likely to play a role invivo as well. One could speculate that the use of A2AR agonistsduring chronic inflammatory conditions might selectively inhibitproinflammatory cytokine production of activated microglia,whereas the addition of A3R antagonists might potentiate inhibi-tory effects of adenosine on unstimulated microglia as well. Therecent description of a synthetic compound that demonstrates bothadenosine A2AR agonist and A3R antagonist activity thereforeholds pharmacological promise (49). However, the widespread ex-pression of adenosine receptors on different cells and tissues andthe described neuroprotective effects of A2AR antagonists in ani-mal models for stroke and Parkinson’s disease (50, 51) warrantcaution when extrapolating these results to therapeuticapplications.
FIGURE 6. Schematic representation of our working hypothesis. A, In resting microglia, A3R-mediated signaling counteracts A2AR-mediated signalingeither by stimulating TLR-mediated signaling or directly by inhibiting A2AR-mediated inhibition of TLR-mediated signaling. B, In activated microglia thecontribution of A3R-mediated signaling is decreased and the contribution of A2AR-mediated signaling is increased, unleashing the inhibitory potential ofextracellular adenosine on TLR-induced proinflammatory cytokine production. IKK, inhibitor of NF-�B kinase; IRAK, IL1-receptor associated kinase;MyD88, myeloid differentiation primary response gene 88; RIP, receptor-interacting protein; TRAF, TNF-receptor associated factor; TRAM, TRIF-relatedadaptor molecule; TRIF, TIR-domain containing adaptor protein inducing IFN-�.
Table II. Rhesus monkey, mouse, and rat adenosine receptor proteinhomology with human adenosine receptors
A1R (%) A2AR (%) A2BR (%) A3R (%)
Macaca mulatta 99.4 98.3 98.5 97.5Mus musculus 96.9 86.5 92.8 84.7Rattus norvegicus 96.6 86.0 92.2 86.9
7611The Journal of Immunology
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

AcknowledgmentsWe thank T. Haaksma and T. Mulder for expert technical assistance. Wethank Dr. E. Remarque for expert assistance with the statistical analysesand Drs. R.E. Bontrop, S.B. Geutskens, L.A. ‘t Hart, and J.M. van Noortfor critically reading the manuscript.
DisclosuresThe authors have no financial conflict of interest.
References1. Aloisi, F., F. Ria, G. Penna, and L. Adorini. 1998. Microglia are more efficient
than astrocytes in antigen processing and in Th1 but not Th2 cell activation.J. Immunol. 160: 4671–4680.
2. Santambrogio, L., S. L. Belyanskaya, F. R. Fischer, B. Cipriani, C. F. Brosnan,P. Ricciardi-Castagnoli, L. J. Stern, J. L. Strominger, and R. Riese. 2001. De-velopmental plasticity of CNS microglia. Proc. Natl. Acad. Sci. USA 98:6295–6300.
3. Janeway, C. A., Jr., and R. Medzhitov. 2002. Innate immune recognition. Annu.Rev. Immunol. 20: 197–216.
4. Kawai, T., and S. Akira. 2005. Pathogen recognition with Toll-like receptors.Curr. Opin. Immunol. 17: 338–344.
5. Iwasaki, A., and R. Medzhitov. 2004. Toll-like receptor control of the adaptiveimmune responses. Nat. Immunol. 5: 987–995.
6. Akira, S. 2006. TLR signaling. Curr. Top. Microbiol. Immunol. 311: 1–16.7. O’Neill, L. A. 2006. How Toll-like receptors signal: what we know and what we
don’t know. Curr. Opin. Immunol. 18: 3–9.8. Takeda, K., and S. Akira. 2005. Toll-like receptors in innate immunity. Int. Im-
munol. 17: 1–14.9. Bsibsi, M., R. Ravid, D. Gveric, and J. M. van Noort. 2002. Broad expression of
Toll-like receptors in the human central nervous system. J. Neuropathol. Exp.Neurol. 61: 1013–1021.
10. Jack, C. S., N. Arbour, J. Manusow, V. Montgrain, M. Blain, E. McCrea,A. Shapiro, and J. P. Antel. 2005. TLR signaling tailors innate immune responsesin human microglia and astrocytes. J. Immunol. 175: 4320–4330.
11. Olson, J. K., and S. D. Miller. 2004. Microglia initiate central nervous systeminnate and adaptive immune responses through multiple TLRs. J. Immunol. 173:3916–3924.
12. Zuiderwijk-Sick, E. A., C. van der Putten, M. Bsibsi, I. P. Deuzing, W. de Boer,C. Persoon-Deen, I. Kondova, L. A. Boven, J. M. van Noort, B. A. t Hart,S. Amor, and J. J. Bajramovic. 2007. Differentiation of primary adult microgliaalters their response to TLR8-mediated activation but not their capacity as APC.Glia 55: 1589–1600.
13. Hasko, G., P. Pacher, E. A. Deitch, and E. S. Vizi. 2007. Shaping of monocyteand macrophage function by adenosine receptors. Pharmacol. Ther. 113:264–275.
14. Link, A. A., T. Kino, J. A. Worth, J. L. McGuire, M. L. Crane, G. P. Chrousos,R. L. Wilder, and I. J. Elenkov. 2000. Ligand-activation of the adenosine A2areceptors inhibits IL-12 production by human monocytes. J. Immunol. 164:436–442.
15. Nemeth, Z. H., C. S. Lutz, B. Csoka, E. A. Deitch, S. J. Leibovich, W. C. Gause,M. Tone, P. Pacher, E. S. Vizi, and G. Hasko. 2005. Adenosine augments IL-10production by macrophages through an A2B receptor-mediated posttranscrip-tional mechanism. J. Immunol. 175: 8260–8270.
16. Fredholm, B. B., A. P. IJzerman, K. A. Jacobson, K. N. Klotz, and J. Linden.2001. International Union of Pharmacology. XXV. Nomenclature and classifi-cation of adenosine receptors. Pharmacol. Rev. 53: 527–552.
17. Jacobson, K. A., and Z. G. Gao. 2006. Adenosine receptors as therapeutic targets.Nat. Rev. Drug Discov. 5: 247–264.
18. Hasko, G., P. Pacher, E. S. Vizi, and P. Illes. 2005. Adenosine receptor signalingin the brain immune system. Trends Pharmacol. Sci. 26: 511–516.
19. Pfaffl, M. W. 2001. A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res. 29: e45.
20. Pinheiro, J. C., and D. M. Bates. 2000. Mixed Effects Models in S and S-plus.Springer-Verlag, New York.
21. R Development Core Team. 2007. R: a language and environment for statisticalcomputing. R Foundation for Statistical Computing, Vienna, Austria.
22. Murphree, L. J., G. W. Sullivan, M. A. Marshall, and J. Linden. 2005. Lipopoly-saccharide rapidly modifies adenosine receptor transcripts in murine and humanmacrophages: role of NF-�B in A(2A) adenosine receptor induction. Biochem. J.391: 575–580.
23. Hasko, G., D. G. Kuhel, J. F. Chen, M. A. Schwarzschild, E. A. Deitch,J. G. Mabley, A. Marton, and C. Szabo. 2000. Adenosine inhibits IL-12 andTNF-� production via adenosine A2a receptor-dependent and independent mech-anisms. FASEB J. 14: 2065–2074.
24. Lee, J. Y., B. S. Jhun, Y. T. Oh, J. H. Lee, W. Choe, H. H. Baik, J. Ha,K. S. Yoon, S. S. Kim, and I. Kang. 2006. Activation of adenosine A3 receptorsuppresses lipopolysaccharide-induced TNF-� production through inhibition ofPI 3-kinase/Akt and NF-�B activation in murine BV2 microglial cells. Neurosci.Lett. 396: 1–6.
25. Eigler, A., T. F. Greten, B. Sinha, C. Haslberger, G. W. Sullivan, and S. Endres.1997. Endogenous adenosine curtails lipopolysaccharide-stimulated tumour ne-crosis factor synthesis. Scand. J. Immunol. 45: 132–139.
26. Sperlagh, B., M. Baranyi, G. Hasko, and E. S. Vizi. 2004. Potent effect of inter-leukin-1� to evoke ATP and adenosine release from rat hippocampal slices.J. Neuroimmunol. 151: 33–39.
27. Fredholm, B. B. 2007. Adenosine, an endogenous distress signal, modulates tis-sue damage and repair. Cell Death Differ. 14: 1315–1323.
28. Boucsein, C., R. Zacharias, K. Farber, S. Pavlovic, U. K. Hanisch, andH. Kettenmann. 2003. Purinergic receptors on microglial cells: functional ex-pression in acute brain slices and modulation of microglial activation in vitro.Eur. J. Neurosci. 17: 2267–2276.
29. Cunha, R. A. 2005. Neuroprotection by adenosine in the brain: from A(1) recep-tor activation to A (2A) receptor blockade. Purinergic Signal. 1: 111–134.
30. Dare, E., G. Schulte, O. Karovic, C. Hammarberg, and B. B. Fredholm. 2007.Modulation of glial cell functions by adenosine receptors. Physiol. Behav. 92:15–20.
31. Jijon, H. B., J. Walker, F. Hoentjen, H. Diaz, J. Ewaschuk, C. Jobin, andK. L. Madsen. 2005. Adenosine is a negative regulator of NF-�B and MAPKsignaling in human intestinal epithelial cells. Cell. Immunol. 237: 86–95.
32. Palmer, T. M., and M. A. Trevethick. 2008. Suppression of inflammatory andimmune responses by the A(2A) adenosine receptor: an introduction.Br. J. Pharmacol. 153(Suppl 1): S27–S34.
33. Hammarberg, C., B. B. Fredholm, and G. Schulte. 2004. Adenosine A3 receptor-mediated regulation of p38 and extracellular-regulated kinase ERK1/2 via phos-phatidylinositol-3�-kinase. Biochem. Pharmacol. 67: 129–134.
34. Hammarberg, C., G. Schulte, and B. B. Fredholm. 2003. Evidence for functionaladenosine A3 receptors in microglia cells. J. Neurochem. 86: 1051–1054.
35. la Sala, A., M. Gadina, and B. L. Kelsall. 2005. G(i)-protein-dependent inhibitionof IL-12 production is mediated by activation of the phosphatidylinositol 3-ki-nase-protein 3 kinase B/Akt pathway and JNK. J. Immunol. 175: 2994–2999.
36. Gsandtner, I., C. Charalambous, E. Stefan, E. Ogris, M. Freissmuth, andJ. Zezula. 2005. Heterotrimeric G protein-independent signaling of a G protein-coupled receptor: direct binding of ARNO/cytohesin-2 to the carboxyl terminusof the A2A adenosine receptor is necessary for sustained activation of the ERK/MAP kinase pathway. J. Biol. Chem. 280: 31898–31905.
37. Gsandtner, I., and M. Freissmuth. 2006. A tail of two signals: the C terminus ofthe A(2A)-adenosine receptor recruits alternative signaling pathways. Mol. Phar-macol. 70: 447–449.
38. Khoa, N. D., M. Postow, J. Danielsson, and B. N. Cronstein. 2006. Tumor ne-crosis factor-� prevents desensitization of G�s-coupled receptors by regulatingGRK2 association with the plasma membrane. Mol. Pharmacol. 69: 1311–1319.
39. Nguyen, D. K., M. C. Montesinos, A. J. Williams, M. Kelly, and B. N. Cronstein.2003. Th1 cytokines regulate adenosine receptors and their downstream signalingelements in human microvascular endothelial cells. J. Immunol. 171: 3991–3998.
40. Sands, W. A., A. F. Martin, E. W. Strong, and T. M. Palmer. 2004. Specificinhibition of nuclear factor-�B-dependent inflammatory responses by cell type-specific mechanisms upon A2A adenosine receptor gene transfer. Mol. Pharma-col. 66: 1147–1159.
41. Khoury, J., J. C. Ibla, A. S. Neish, and S. P. Colgan. 2007. Antiinflammatoryadaptation to hypoxia through adenosine-mediated cullin-1 deneddylation.J. Clin. Invest. 117: 703–711.
42. Wittendorp, M. C., H. W. Boddeke, and K. Biber. 2004. Adenosine A3 receptor-induced CCL2 synthesis in cultured mouse astrocytes. Glia 46: 410–418.
43. Wittendorp, M. C., J. von Frijtag Drabbe Kunzel, A. P. Ijzerman, H. W. Boddeke,and K. Biber. 2004. The mouse brain adenosine A1 receptor: functional expres-sion and pharmacology. Eur. J. Pharmacol. 487: 73–79.
44. Fiebich, B. L., K. Biber, K. Gyufko, M. Berger, J. Bauer, and D. van Calker.1996. Adenosine A2b receptors mediate an increase in interleukin (IL)-6 mRNAand IL-6 protein synthesis in human astroglioma cells. J. Neurochem. 66:1426–1431.
45. Fiebich, B. L., K. Biber, K. Lieb, D. van Calker, M. Berger, J. Bauer, andP. J. Gebicke-Haerter. 1996. Cyclooxygenase-2 expression in rat microglia isinduced by adenosine A2a-receptors. Glia 18: 152–160.
46. Xaus, J., M. Mirabet, J. Lloberas, C. Soler, C. Lluis, R. Franco, and A. Celada.1999. IFN-� up-regulates the A2B adenosine receptor expression in macro-phages: a mechanism of macrophage deactivation. J. Immunol. 162: 3607–3614.
47. Macedo, L., G. Pinhal-Enfield, V. Alshits, G. Elson, B. N. Cronstein, andS. J. Leibovich. 2007. Wound healing is impaired in MyD88-deficient mice: arole for MyD88 in the regulation of wound healing by adenosine A2A receptors.Am. J. Pathol. 171: 1774–1788.
48. Pinhal-Enfield, G., M. Ramanathan, G. Hasko, S. N. Vogel, A. L. Salzman,G. J. Boons, and S. J. Leibovich. 2003. An angiogenic switch in macrophagesinvolving synergy between Toll-like receptors 2, 4, 7, and 9 and adenosine A(2A)receptors. Am. J. Pathol. 163: 711–721.
49. Bevan, N., P. R. Butchers, R. Cousins, J. Coates, E. V. Edgar, V. Morrison,M. J. Sheehan, J. Reeves, and D. J. Wilson. 2007. Pharmacological characteri-sation and inhibitory effects of (2R,3R,4S,5R)-2-(6-amino-2-{[(1S)-2-hydroxy-1-(phenylmethyl)ethyl]amino}-9 H-purin-9-yl)-5-(2-ethyl-2H-tetrazol-5-yl)tetra-hydro-3,4-furandiol, a novel ligand that demonstrates both adenosine A(2A)receptor agonist and adenosine A(3) receptor antagonist activity. Eur. J. Phar-macol. 564: 219–225.
50. Chen, J. F., and F. Pedata. 2008. Modulation of ischemic brain injury and neu-roinflammation by adenosine A2A receptors. Curr. Pharm. Des. 14: 1490–1499.
51. Kalda, A., L. Yu, E. Oztas, and J. F. Chen. 2006. Novel neuroprotection bycaffeine and adenosine A(2A) receptor antagonists in animal models of Parkin-son’s disease. J. Neurol. Sci. 248: 9–15.
7612 DYNAMIC A3R EXPRESSION CONTROLS A2AR-MEDIATED TLR INHIBITION
by guest on February 25, 2016http://w
ww
.jimm
unol.org/D
ownloaded from





























