Embed Size (px)
Citation preview

Wilhelm Roux's Archives 186, 51-64 (1979) Roux's Archives of Developmental Biology �9 by Springer-Verlag 1979
Differentiation Capacities of the Labial Imaginal Disc of Drosophila melanogaster
Kaushal Kumar* , Willem J. Ouweneel**, and Jacob Faber Hubrecht Laboratory (International Embryological Institute), Utrecht, The Netherlands
Summary. The mature labial disc, when implanted into a larva of the same age, undergoes metamorphosis along with the host and produces one lateral half of the medi- and distiproboscis. On the basis of results obtained from transplanted disc halves (including the separate peripodial membrane) a tentative fate map of the labial disc was constructed, which shows most of the presumptive mediproboscis to be located in the dorsal, and most of the presumptive distiproboscis in the ventral part of the disc. The distal port ion of the peripodial membrane also contains imaginal anlagen, viz. part of the mediproboscis, prementum, and labellar cap anlagen. The involve- ment of this part of the peripodial membrane was checked by a careful histological analysis of labial disc development during the first ten hours after prepupation. The results were compared with the situation described for Calliphora imaginal discs.
In addition, a detailed morphological analysis was made of the proboscis of the homoeotic mutant proboscipedia (pb). At 27 ~ C, pb changes the distipro- boscis into a " te lopodi te" (leg segments distal to the coxa); the (unchanged) prementum may therefore correspond to the coxa. At 15 ~ C, the tarsus of this homoeotic " te lopodi te" is replaced to a greater or lesser extent by an arista. The present analysis thus confirms (a) the fundamental morphological correspondence of the medi- and distiproboscis with the la- bium of other insects, and (b) the fundamental developmental correspondence of the labial, antennal, and leg discs.
Key words: Imaginal discs - Labial disc - Fate map - Drosophi la -
Homoeosis.
* Present address: Dept. of ZooIogy, Univ. of Gorakhpur, Gorakhpur-273001 India. (K.K. was a member of the 8th International Research Group in Developmental Biology, and was the recipient of a UNESCO travel grant.) ** Present address: Emmalaan 1, De Bilt, The Netherlands Offprint requests should be sent to Dr. J. Faber, Hubrecht Laboratory, Uppsalalaan 8, 3584 CT Utrecht, The Netherlands
0340-0794/79/0186/0051/$02.80

52 K. Kumar et al.
Introduction
By t r ansp lan t ing f ragments o f imagina l discs into ma tu re host la rvae and examin- ing the adu l t s t ructures they fo rm after undergo ing m e t a m o r p h o s i s a long with the host , de ta i led fate maps could be m a d e for mos t of the imagina l discs of Drosophila melanogaster. So far no fate m a p has been made for the labia l disc, poss ib ly because its very small size makes f r agmen ta t ion and t r ansp lan ta - t ion very difficult . In the present s tudy we descr ibe the results of exper iments in which halves o f lab ia l discs were t r ansp lan ted , and supply a fate m a p for this disc. A new aspect of this m a p for Drosophila is tha t we also separa te ly t r an sp l an t ed the so-cal led " p e r i p o d i a l m e m b r a n e " , which in the lab ia l disc is re la t ively th icker than in o ther discs and therefore t hough small can be t rans- p l an ted m o r e easily. It is shown here, bo th by means o f t r a n s p l a n t a t i o n and in a h is to logica l s tudy of the m e t a m o r p h o s i n g lab ia l disc in which the fate o f the pe r ipod ia l m e m b r a n e was examined , tha t the pe r ipod ia l m e m b r a n e ap- pears to con ta in imag ina l p r imord ia .
A ques t ion o f special in teres t concerns the re la t ionsh ip of the labia l disc m a p with the fate m a p s o f o ther imagina l discs. A close cor re la t ion is suggested by the homoeo t i c m u t a t i o n proboscipedia (pb), which replaces specific p robosc i s par t s by specific leg or an tenna l par t s (Bridges and D o b z h a n s k y , 1933), and which we have fur ther ana lysed in this study. The presen t s tudy shows a g o o d co r r e spondence between the fate m a p s o f the labia l , the an tenna l and the leg discs.
Material and Methods
To obtain labial discs a Sevelen wild-type stock was used. All strains (donors and hosts) were kept at 25~ on standard medium (sugar-maize-yeast-agar). Mature labial discs were dissected out in insect Ringer and implanted into the abdomina of full-grown female Sevelen larvae, according to the technique described by Ursprung (1967). The discs were either implanted intact or were first bisected with the help of tungsten needles. The metamorphosed implants were dissected from the flies and embedded in Faure's mixture.
To study the fate of the peripodial membrane during metamorphosis histological preparations of prepupae were made. Prepupae were fixed in Van Leeuwen's fixative (1% picric acid in abs. alcohol: 6 pts; chloroform: I pt; formalin: 1 pt; add 1:16 pts of glacial acetic acid just before use) for 2 h at 50 ~ C, embedded in paraffin wax, transversely sectioned at 7 gm, stained in haemalum- erythrosin, and mounted in Depex.
For analysis of the proboscipedia (pb) proboscis, larvae of the mutant strain pb/In(3LR)Cx were raised either at 27 ~ C (leading to replacement of proboscis structures by leg structures) or at 15 ~ C (leading to replacement of the homoeotic tarsus by an arista). Heads of adult flies were mounted in Faure's mixture.
Results
The Entire Disc
The no rma l inventory o f s t ructures fo rmed by the lab ia l disc was examined in 25 whole moun t s of t r ansp lan ted a n d m e t a m o r p h o s e d entire discs. The results
Fate Map of Drosophila Labial Disc 53
i i ~! +i~!! !~ii[ �84 ~i~iil,i :~ ~ i~,~iiiii!i~
!i
DP
a b
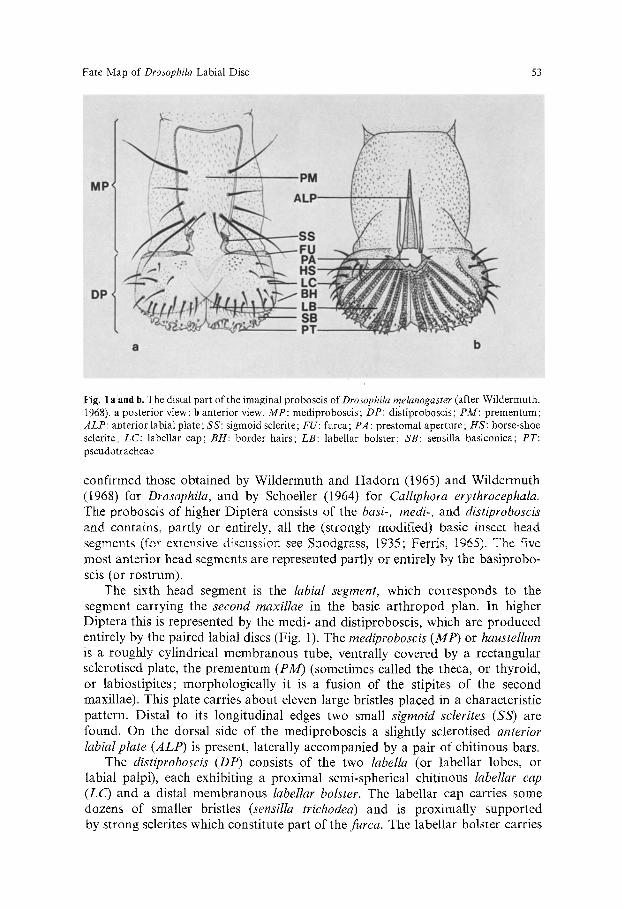
Fig. 1 aand b. The distal part of the imaginal proboscis of Drosophila melanogaster (after Wildermuth, 1968). a posterior view; b anterior view. MP: mediproboscis; DP: distiproboscis; PM: prementum; ALP: anterior labial plate; SS: sigmoid sclerite; FU: furca; PA: prestornal aperture; HS: horse-shoe sclerite: LC: labellar cap; BH: border hairs; LB: labellar bolster; SB: sensilla basiconica; PT: pseudotraeheae
confirmed those obtained by Wildermuth and Hadorn (1965) and Wildermuth (1968) for Drosophila, and by Schoeller (1964) for Calliphora erythrocephala. The proboscis of higher Diptera consists of the basi-, medi-, and distiproboscis and contains, partly or entirely, all the (strongly modified) basic insect head segments (for extensive discussion see Snodgrass, 1935; Ferris, i965). The five most anterior head segments are represenled partly or entirely by the basiprobo- scis (or rostrum),
The sixth head segment is the labial segment, which corresponds to the segment carrying the second maxillae in the basic arthropod plan. In higher Diptera this is represented by the medi- and distiproboscis, which are produced entirely by the paired labial discs (Fig. 1). The mediproboscis (MP) or haustellum is a roughly cylindrical membranous tube, ventrally covered by a rectangular sclerotised plate, the prementum (PM) (sometimes called the theca, or thyroid, or labiostipites; morphologically it is a fusion of the stipites of the second maxillae). This plate carries about eleven large bristles placed m a characteristic pattern. Distal to its longitudinal edges two small sigmoid sclerites (SS) are found. On the dorsal side of the mediproboscis a slightly sclerotised anterior labial plate (ALP) is present, laterally accompanied by a pair of chitinous bars.
The distiproboscis (DP) consists of the two labella (or labellar lobes, or labial palpi), each exhibiting a proximal semi-spherical chitinous labellar cap (LC) and a distal membranous labellar bolster. The labellar cap carries some dozens of smaller bristles (sensilla trichodea) and is proximally supported by strong sclerites which constitute part of the furca. The labellar bolster carries

54 K. Kumar et al.
S �9
L C b
Fig. 2. A metamorphosed implant of a whole labial disc. In the photograph the following structures can be recognised: MP: mediproboscis; PMb: prementum bristles; S: sclerites (unidentifiable in this picture); PT: pseudotracheae; LCb: labellar cap bristles
about fivepseudotracheae (PT) running radially from what we call the horse-shoe sclerite (HS), which borders the prestomal aperture (not to be confused, as Wildermuth and Hadorn (1965) do, with the mouth proper, which is situated at the base of the labrum). Irregularly dispersed between the pseudotracheae are a number of sensilla basiconica. The border between the cap and the bolster is marked by a regular row of slender trichomes, the border hairs (BH).
In metamorphosed whole-disc implants (see Fig. 2) we found the MP to be severely shrunken (also reported by Wildermuth and Hadorn, 1965). For the shrivelled PM this results in certain bristles sometimes being placed so close together that their sockets are partly in touch. The size of the bristles is normal, however. The DP elements are much more regular in implants. Only the PT are often aberrant; in many cases they are reduced in size and number and/or branched. The SS and furca are often difficult to distinguish in implants; hence they were scored together. Otherwise, we could trace in all implants the whole inventroy of structures known to be produced by the labial discs (see Table 1).
Transplanted Disc Halves
The labial discs were bisected in three different ways, as shown in Fig. 3 a -c ; right and left discs were bisected in symmetrical fashion. Because of the very small size of the discs a finer fragmentation was impracticable. The results of these six experimental series taken together provide information on the developmental capacities of the eight areas shown in Fig. 3 d. The labial struc- tures formed by the 116 metamorphosed disc halves are summarised in Table 1. For easier comparison the percentages of incidence of given structures in comple-
Fate Map of Drosophila Labial Disc 55
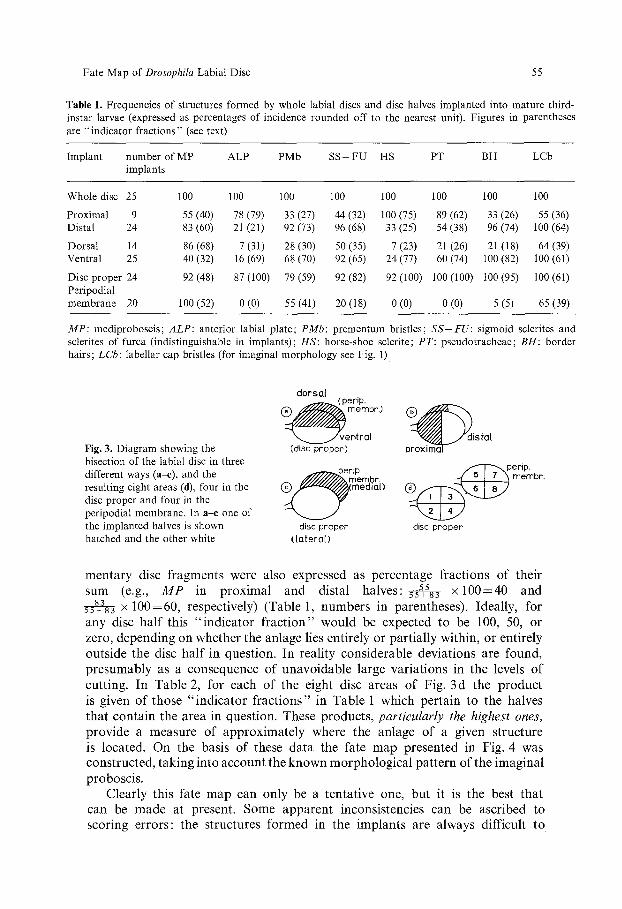
Table 1. Frequencies of structures formed by whole labial discs and disc halves implanted into mature third- instar larvae (expressed as percentages of incidence rounded off to the nearest unit). Figures in parentheses are "indicator fractions" (see text)
Implant number of MP ALP PMb SS+FU HS PT BH LCb implants
Whole disc 25 100 100 100 100 100 100 100 100
Proximal 9 55 (40) 78 (79) 33 (27) 44 (32) 100 (75) 89 (62) 33 (26) 55 (36) Distal 24 83 (60) 21 (21) 92 (73) 96 (68) 33 (25) 54 (38) 96 (74) 100 (64)
Dorsal 14 86 (68) 7 (31) 28 (30) 50 (35) 7 (23) 21 (26) 21 (18) 64 (39) Ventral 25 40 (32) 16 (69) 68 (70) 92 (65) 24 (77) 60 (74) 100 (82) 100 (61)
Disc proper 24 92 (48) 87 (100) 79 (59) 92 (82) 92 (100) 100 (100) 100 (95) 100 (61) Peripodial membrane 20 100 (52) 0 (0) 55 (41) 20 (18) 0 (0) 0 (0) 5 (5) 65 (39)
MP: mediproboscis; ALP: anterior labial plate; PMb: prementum bristles; SS+FU: sigmoid sclerites and sclerites of furca (indistinguishable in implants); HS: horse-shoe sclerite; PT: pseudotracheae; BH: border hairs; LCb: labellar cap bristles (for imaginal morphology see Fig. 1)
Fig. 3. Diagram showing the bisection of the labial disc in three different ways (a--e), and the resulting eight areas (d), four in the disc proper and four in the peripodial membrane. In a-e one of the implanted halves is shown hatched and the other white
dorsal
~ v e perip" membr.)
ntral (disc proper)
~ rip. membr,
@ ~ ( m e d m l )
disc proper ([oteraD
@ . ~ d i s t a t proxima~
membr.
disc proper
mentary disc fragments were also expressed as percentage fractions of their sum (e.g., M P in proximal and distal halves: 5s 55+83 x 1 0 0 = 4 0 and 5s~a83 • 100=60, respectively) (Table 1, numbers in parentheses). Ideally, for any disc half this "indicator fract ion" would be expected to be 100, 50, or zero, depending on whether the anlage lies entirely or partially within, or entirely outside the disc half in question. In reality considerable deviations are found, presumably as a consequence of unavoidable large variations in the levels of cutting. In Table 2, for each of the eight disc areas of Fig. 3 d the product is given of those '~ fractions" in Table 1 which pertain to the halves that contain the area in question. These products, particularly the highest ones, provide a measure of approximately where the anlage of a given structure is located. On the basis of these data the fate map presented in Fig. 4 was constructed, taking into account the known morphological pattern of the imaginal proboscis.
Clearly this fate map can only be a tentative one, but it is the best that can be made at present. Some apparent inconsistencies can be ascribed to scoring errors: the structures formed in the implants are always difficult to

K. Knmar et al.
proximal
MP ALP PMb S S + F U HS PT BH LCb
1. (D, Pr, Dor) i3 24 5 9 17 16 4 9 2. (D, Pr, Ven) 6 54 l l 17 58 46 20 13 3. (D, Di, Dor) 20 7 13 20 6 10 13 15 4. (D, Di, Ven) 9 14 30 36 19 28 58 24
5. (PM, Pr, Dor) 14 0 3 2 0 0 0 5 6. (PM, Pr, Ven) 7 0 8 4 0 0 1 9 7. (PM, Di, Dor) 21 0 9 4 0 0 1 10 8. (PM, Di, Ven) 10 0 21 8 0 0 3 15
Abbreviations of structures as in Table 1. The numbers 1-8 denote the areas shown in Fig. 3d D: disc proper; PM: peripodial membrane; Pr: proximal; Di: distal; Dor: dorsal; Ven: ventral
dista(
Lateral Medial
(disc epithelium proper (peripodial membrane of right disc) of left disc)
dorsQ[ dorsal
56
Table 2. Products of the "indicator fractions" in Table 1 (see text)
Fig. 4. Tentative fate maps of the labial disc The anlagen are indicated by the abbreviations
ventral vent ra[
proper (left) and its peripodial membrane (right). used in Fig. 1
recognise and the various sclerites in particular are easily confused. This may explain the scorings for ALP and HS in distal halves, and for PMb in proximal halves of the disc proper (see Table 1).
For the first time in Drosophila, the peripodial membrane is here shown to contain imaginal anlagen. The location of the membrane portion assumed to contain imaginal anlagen (Fig. 4) was inferred from the fact that large amounts of MP, PM, and LC tissue were found in membrane implants, whereas certain other medial structures (ALP, HS, and PT) were entirely lacking (see Table 1). It was therefore assumed that only part of the periphery of the membrane participates in the formation of the MP and DP, that the larger part of the membrane disappears during metamorphosis, and that the edges of the remaining part of the membrane and the disc proper fuse with those of the contralateral labial disc. As a matter of fact, the imaginal anlagen bordering the postulated edges are exactly those giving rise to the proboscis structures bordering the medial symmetry plane of the MP and DP (each sagittal half of which is

Fate Map of Drosophila Labial Disc 57
b
fig.5 (2h after prepupation /
...... .'.~-~-.L.::: :,-~Y_-~:~ ~
.... V."'"'V;/" ................................. ;~,'"'i .............. a pm -----J--~ / ,' _ \
aw ~ ~ / ~ ..... I,~ .~1 I't"""~ //" ~ ,' .............. . - / ." : : '~K" ". .................. "'-. . I ~ ,
fig. 7 (5 h after prepupatlon)
aw
d p ~ d l a aw
fig.6 (4h after prepupation)
Figs. 5-7. Development of labial disc proper and peripodial membrane at 2, 4 and 5 h after prepupation. Each figure consists of a graphical reconstruction of the two labial discs and the atrium in ventral view (anterior on top), and two selected cross sections at the levels indicated by a and b (in normal dorso-ventral orientation). In the graphical reconstructions the atrial wall in surface view is hatched; unbroken lines indicate visible external tissue boundaries, broken lines hidden external boundaries and internal tissue boundaries. aw: atrical wall; a: atrium; dp: disc proper; dl: disc lumen;pro: peripodial membrane; dla: communi- cation of disc lumen with atrium; el): unidentified other epithelia; oe: oesophagus
formed by one labial disc). These considerations necessitated a closer morpholog- ical study of the peripodial membrane during early metamorphosis.
Fate of the Peripodial Membrane
On the basis of cross sections of 93 prepupae made during the first eight hours after prepupation we have attempted to follow the further development

58 K. Kumar et al.
o f t h e p e r i p o d i a l m e m b r a n e a n d t h e l a b i a l d isc p r o p e r . Figs . 5 - 7 a n d d e s c r i p t i o n s
o f t h e d e v e l o p m e n t a l s i t u a t i o n a t c e r t a i n t i m e s a f t e r p r e p u p a t i o n wil l p r o v i d e
a p i c t u r e o f t h i s p r o c e s s in s p a c e a n d t ime .
One to Two Hours (Fig. 5). Proximally the discs are attached to the atrium over a considerable width, but fusion between the discs is still limited. The epithelia of the discs proper have become thicker and the discs are broader than in the larva. There are no open connections between the lumina of the discs and the atrium.
Three Hours. The peripodial membrane is strongly extended anteriorly and entirely fused with the atrium cranial to the region of fusion of the discs.
Four Hours (Fig. 6). The fusion of the two discs is complete along more than three quarters of their length. In a broad anterior zone the peripodial membrane is detached from the disc proper and is now attached to the atrial wall in front of the opening. The disc proper is quite firmly attached to the atrial wall along about the anterior third of its length, particularly on the dorsal side of the disc where the disc proper passes into the peripodial membrane.
Five Hours (Fig. 7). The detachment of the peripodial membrane from the disc proper gradually continues along the dorsal side of the anlage, the disc proper fusing with the ventral, and the peripodial membrane with the dorsal atrial wall. This spectacular simultaneous rupture in the atrial wall and the discs, and the subsequent fusion of the corresponding epithelia, at this stage extends about half the length of the discs. Anteriorly the two discs proper and the ventral atrial wall thus form a firm, indented tube.
Six Hours. Anteriorly the peripodial membrane is diminishing in area; it is continuous with the larval epithelial layers of the atrium that have the important but temporary function of fusing with the developing imaginal rudiments and drawing them together. As these larval epithelia diminish in area and finally disappear, the imaginal epithelia grow and finally fuse with each other.
Seven Hours. At this stage another important epithelial rupture and re-fusion takes place. The distal part of the peripodial membrane (i.e. the part away from the original disc stalk) detaches itself from the rest of the membrane and fuses with the palpus rudiment of the antennal disc, which now extends far anteriorly. More posteriorly this rudiment detaches itself from the rest of the antennal disc and fuses with the ventral half of the clypeo-labrum rudiment to form an epithelial tube. Thus the beginning of proboscis formation is already observable, its rudiment consisting of fused parts of the labial and antennal discs, the clypeo-labrum rudiment, and some connecting larval epithelia.
Eight Hours. The ventral atrial wall has nearly disappeared, so that the discs proper are almost completely fused ventrally. The antennal discs extend nearly the full length of the labial discs and fuse with them at about one third of their length. In front of the posterior end of each disc the palpal rudiment has already fused dorsally with the ventral half of the clypeo-labrum rudiment (the dorsal half of which fuses with the remainder of the antennal disc).
I t m a y b e c o n c l u d e d t h a t t h i s s t u d y b r o a d l y c o n f i r m s t he c o n f i g u r a t i o n
o f t h e d i sc f a t e m a p in t h a t i n d e e d t he d i s t a l p a r t o f t he p e r i p o d i a l m e m b r a n e
c o n t r i b u t e s to t h e f o r m a t i o n o f t h e p r o b o s c i s , w h e r e a s t h e p r o x i m a l p a r t h a s
a t e m p o r a r y m o r p h o g e n e t i c f u n c t i o n , a l o n g w i t h s eve ra l l a r v a l e p i t h e l i a , b u t
e v e n t u a l l y d i s a p p e a r s .
M o r p h o l o g y o f H o m o e o t i c Proboscises
A t o t a l o f 3 0 8 p b h a l f p r o b o s c i s e s w e r e e x a m i n e d , 260 g r o w n a t 1 5 ~ a n d 48
g r o w n a t 27 ~ C.
Fate Map of Drosophila Labial Disc 59
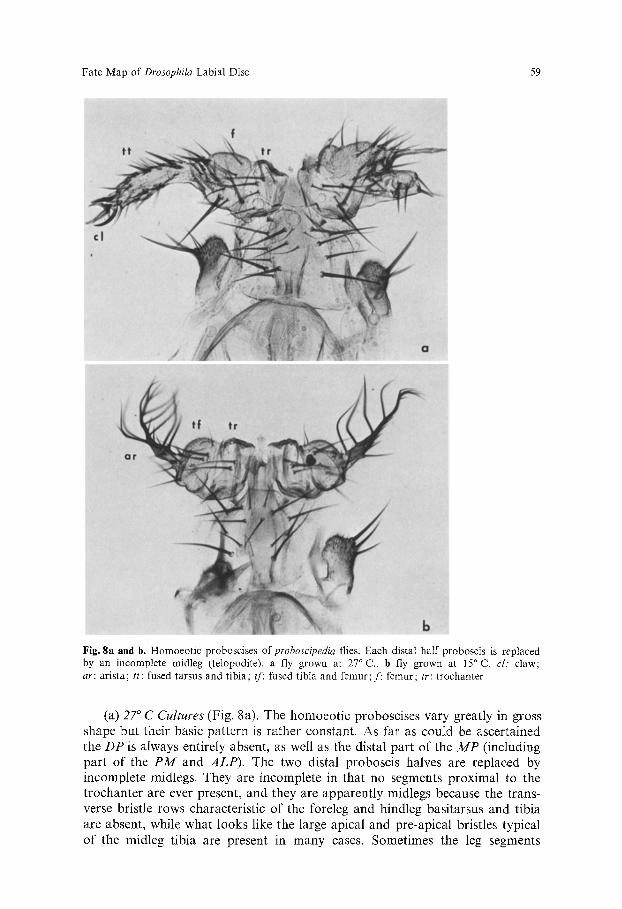
b Fig. 8a and b. Homoeotic proboscises ofproboscipedia flies. Each distal half proboscis is replaced by an incomplete midleg (telopodite). a fly grown at 27 ~ C., b fly grown at 15 ~ C. cl: claw; ar: arista; tt: fused tarsus and tibia; tf: fused tibia and femur; f: femur; tr: trochanter
(a) 27 ~ C Cul tures (Fig. 8a). The homoeot ic proboscises vary greatly in gross shape but their basic pat tern is rather constant. As far as could be ascertained the D P is always entirely absent, as well as the distal part o f the M P (including par t o f the P M and A L P ) . The two distal proboscis halves are replaced by incomplete midlegs. They are incomplete in that no segments proximal to the t rochanter are ever present, and they are apparent ly midlegs because the trans- verse bristle rows characteristic of the foreleg and hindleg basitarsus and tibia are absent, while what looks like the large apical and pre-apical bristles typical of the midleg tibia are present in many cases. Sometimes the leg segments

60 K. Kumar et al.
are fused to such an extent that they cannot be distinguished and identified. When identifiable, the segments are always very much smaller than in mormal legs. Usually the number of tarsal segments is reduced or their segmentation is defective. Sometimes certain leg segments are transversely duplicated.
The appendage nearly always bears a terminal claw. The tarsal segments can be recognised because they are narrower and carry short, stiff bristles with bracts. The tibia is a somewhat broader short segment, carrying longer, more slender bristles with bracts. The femur is a much broader segment, conically shaped, with longer bristles, the distal ones but not the proximal ones with bracts, as on the normal femur. Sometimes the characteristic proximal group of sensilla campaniformia is partially present. The most proximal leg segment present, the trochanter, is broad but quite short and carries fewer hairs and a few bristles, often unusually long. It also usually carries one or several groups of sensilla trichodea but sensilla campaniformia, though common on the normal trochanter, are seldom found. The bases of the homoeotic appendages are me- dially separated by an unidentifiable strip of hypodermis. The absence of coxae seems striking, but the proximal part of the MP is apparently also slightly affected by pb in that the number of P M bristles is increased (up to 14 16), so that it is possible that the P M corresponds partly or entirely to the coxa.
(b) /5 ~ C Cultures (Fig. 8 b). Many more flies cultured at this temperature were examined because the overall pattern here is much more irregular. First, the amount of tissue replaced varies. It is often less than at 27 ~ C, leading to one or more of the following features: (a) short stretches of PT are present; (b) the trochanter material is inconspicuous (or absent?), or (c) the P M is unaffected (the bristle number being normal). In the other cases the DP and the distal part of the MP are absent, the two distal proboscis halves being replaced entirely by incomplete midlegs. Instead of the tarsus usually a distinct arista is present, although often one or more terminal appendage segments show a mixture of arista features (thinness, branches) and tarsus features (thickness, stiff bristles); sometimes claws are still observed.
No antennal segments other than aristae are ever observed, although in quite a few cases the gross shape of the whole appendage seems to be more that of an antenna than a leg. In such cases the narrow terminal segments seem to correspond to the arista/tarsus and are carried by a large, more or less spheroid segment showing tibia and femur characteristics. This suggests and antennal pattern because the basitarsus, tibia and femur have been shown to correspond to the large third antennal segment (Postlethwait and Schneider- man, 1971). Often however the tibia is narrower, more like that at 27 ~ C. Distinct borders are hardly ever found between any of the leg "segments" . The only distinct segment (if present) is the trochanter, but usually this is also fused with the tibia/femur segment. All the "segments" are characterised by the same features as described for the flies cultured at 27 ~ C.
Discussion
Every imaginal disc is in fact a pouch consisting of a layer of high columnar epithelium, the "disc proper" , and a layer of usually thinner epithelium, the

Fate Map of Drosophila Labial Disc 61
"peripodial membrane". The latter layer has often been thought to disappear entirely during metamorphosis (Poodry and Schneiderman, 1970; Ursprung, 1972). However, the recent work of Mandaron (1978) has shown that the peripo- dial membrane of the leg disc of Drosophila, cultured in vitro, is able to form imaginal cuticle. Investigations with Musca domestica (Dfibendorfer, 1971) and CalIiphora erythrocephala (Emmert, 1972; Spreij and Oldenhave, 1974), where the peripodial membrane of various discs was separately transplanted, had yielded similar results.
These findings and our results have interesting implications for earlier fate maps of other imaginal discs in Drosophila. Spreij and Oldenhave (1974) dis- covered that the peripodial membrane of the wing disc in Calliphora erythroce- phala contributes extensively to the imaginal wing and suggested that this might also hold in some measure for Drosophila discs. They mention unpublished results from time-lapse films suggesting that the peripodial membrane of the wing disc does not disappear but is torn open along a predetermined line and spreads out. Kim and Yang (1972) found that the peripodial membrane of the wing disc of Pieris rapae ruptures in vivo along a similar line and fuses with the larval hypodermis.
We can now also see a difference between the discs mentioned, in Calliphora and Drosophila. Whereas in Calliphora the whole peripodiai membrane seems to be involved in wing formation, so that a single medial rupture line is sufficient to spread out the membrane, in the labial disc of Drosophila the larger portion of the membrane disappears, so that here the rupture line is roughly and ellipsoid surrounding the disappearing part. If other Drosophila discs behaved in either of these two ways, several older fate maps might have to be revised. As a matter of fact, Spreij and Oldenhave (1.c.), in consultation with one of us (W.J.O.), already suggested that certain imaginal anlagen might be located in the peripodial membranes of various discs.
In an earlier paper (Ouweneel and Van der Meer, 1973) the basically concen- tric arrangement of anlagen in most or all imaginal discs was emphasised. The concentric anlagen, from the periphery inwards, correspond to the various segments of the adult appendage in a proximo-distal direction. The labial disc does not exhibit an obviously concentric fate map, but then a proboscis half is not an obviously segmented appendage. Nevertheless, the effects ofproboscipe- dia immediately show that there is a correspondence between the labial disc and such strictly concentrically patterned discs as the antennal and leg disc. The position of the homoeotic appendages suggests that the central part of the labial disc (i.e., roughly the central portion of the LC anlage) corresponds to the central part of the antennal disc (the arista anlage) and to that of the leg disc (the tarsus anlage), and that in the homoeotic labial disc more peripher- ally localised areas produce the more "proximal" segments of the homoeotic appendage. It could be shown that the homoeotic appendages include all the leg segments from trochanter to tarsus, and that the prementum probably corre- sponds partly or entirely to the coxa.
It is striking that pb produces a leg-like appendage from each labial disc according to the pattern of the generalised insect labium (see Snodgrass, 1935). The whole DP (the united labellar lobes) is changed into two "telopodites"

62 K. Kumar et al.
(all leg segments beyond the coxa), so that the conspicuous division into MP and DP might roughly correspond to the basic division into "coxopodite" and "telopodite". The first maxillae are represented in Drosophila by the "pa lp i" , which however resemble the usual maxillary palpi just as little as the labellar lobes resemble the labial palpi.
The difference in homoeotic alteration occurring at different temperatures has been shown not to be simply a switch from leg to antenna, because at both temperatures essentially a leg-like appendage is produced. The only differ- ence is that at 15 ~ C the tarsus of the leg is changed into an arista (just as dachsous in certain genotypes produces aristae instead of tarsi on normal legs; see Ouweneel, 1976).
It is evident that pb affects only a specific region of the labial disc while the rest of the disc develops normally. It is plausible to assume, as Postlethwait and Schneiderman (1971) did for the leg and antennal discs and one of us for the wing and haltere discs (Ouweneel, 1973, 1974, 1976), that the labial, antennal, and leg discs each respond to the same set of influences providing "positional information" (see Wolpert, 1971). The specification of positional values would be identical in the three discs, the only difference between the discs being their interpretation of these values. "Correspondence" between spe- cific labial, antennal and leg structures would then mean that their anlagen are specified by the same positional cues. The interpretation of these cues would depend on the determinative state of the whole disc. The labial, antennal or leg quality of the disc seems to be determined as early as the blastoderm stage (see Ouweneel, 1976). In the case of the homoeotic labial disc it is not known whether the determination of the homoeotic disc region also occurs at this early stage; there are no data on the phenocritical phase or the phenocopy- sensitive or temperature-sensitive period(s).
It is tempting to relate the site of the boundary between the normal and the homoeotic disc region with the "disc compartmentalisation hypothesis" (see review by Garcia-Bellido, 1975). That compartments exist in the developing proboscis is shown by the recent clonal analysis study of Struhl (1977). However, the compartment boundary he found runs lengthwise along the proboscis and divides each half of the medi- and distiproboscis into a medial and a lateral compartment. It thus runs more or less at right angles to the boundary between MP and DP (the presumed coxo- and telopodite, see above).
It seems possible that in normal development pb § is involved in subdividing the anlage of the proboscis (and perhaps those of other appendages) into a coxopodite and a telopodite compartment, although it is equally possible that the incomplete homoeotic transformation is due to hypomorphism of pb. If the first alternative is true, this boundary must represent a different compartmen- talisation step from that demonstrated by Struhl.
Comparisons between the different appendages are rendered difficult by the fact that the spatial pattern of a distal proboscis half is very complicated compared with that of a leg or antenna, and the respective disc maps are quite different. For instance, in the labial disc map the MP and PM anlagen do not surround the anlagen of the more distal structures, as the coxa and antenna I anlagen do in the leg and antennal disc map, respectively: the MP

Fate Map of Drosophila Labial Disc 63
and DP (PT+LC) anlagen each seem to occupy more or less half of the disc and to be separated by a more or less straight boundary. Only clonal analysis of proboscis development beyond the data of Struhl (1977) can clarify the problems of determination and compartmentalisation in the labial disc.
Acknowledgements. We thank Professor P.D. Nieuwkoop and Dr. Anne Fausto-Sterling for critical reading of the manuscript, and the latter for editorial help. The technical assistance of Miss W. Vonk is appreciated. The help of Mrs. E. Wolters and Mr. L. Boom with the illustrations is acknowledged.
References
Bridges, C.B., Dobzhansky, T. : The mutant "proboscipedia" in Drosophila melanogaster - a case of hereditary homo6sis. Wiihelm Roux' Arch. Entwickl.-Mech. Org. 127, 575-590 (t933)
Diibendorfer, A. : Untersuchungen zum Anlageplan und Determinationszustand der weiblichen Genital- und Analprimordien yon Musca domestiea L. Wilhelm Roux' Arch. Entwickl.-Mech.- Org. 168, 142-168 (1971)
Emmert, W. : Entwicklungsleistungen abdominaler Imaginalscheiben von Calliphora erythroeephala (Insecta, Diptera). Experimentelle Untersuchungen zur Morphologie des Abdomens, Wilhelm Roux' Arch. Entwickl.-Mech. Org. 169, 87 133 (1972)
Ferris, G.: External morphology of the adult. In: Biology of Drosophila (M. Demerec, ed.), pp. 364-419. 2nd Printing, London: Hafner 1965
Garcia-Bellido, A.: Genetic control of wing disc development in Drosophila. In: Cell Patterning (R. Porter, ed.), Ciba Found. Syrup. 29 (new set.), pp. i61-182. Amsterdam, Oxford, New York: Elsevier, Excerpta Medica, North Holland 1975
Kim, C.-W., Yang, H.Y. : Post-embryonic development of the wing from larva to adult in Pieris rapae L. Korean J. Entorn. 2, 13~0 (1972)
Mandaron, P. : Formation of presumptive territories in the wing and leg of Drosophila during in vitro evagination. Abstracts of the International Conference on Molecular and Developmental Biology of Insects. Heraclion - Crete (Greece) (1978)
Ouweneel, W.J. : Developmental and genetic homology of the wing and haltere imaginal discs in Drosophila melanogaster. Proc. XIIIth Intern. Congr. Genet., Genetics 74, 207 (1973)
Ouweneel, W.J.: Replacement patterns of homoeotic wing structures in halteres. Dros. Inform. Serv. 50, 102 (1974)
Ouweneel, W.J. : Developmental genetics of homoeosis. Adv. Genetics 18, 179~48 (1976) Ouweneel, W.J., Meer, J.M., Van der: Differentiation capacities of the dorsal metathoracic (haltere)
disc of Drosophila melanogaster. I. Normal organ map. Wilhelm Roux' Arch. Entwickl.-Mech. Org. 172, 149-16I (1973)
Poodry, C.A., Schneiderman, H.A. : The ultrastructure of the developing leg of Drosophila melano- gaster. Wilhelm Roux' Arch. Entwickl.-Mech. Org. 166, 1-44 (1970)
Postlethwait, J.H., Schneiderman, H.A.: Pattern formation and determination in the antenna of the homoeotic mutant Antennapedia of Drosophila melanogaster. Dev. Biol. 25, 606-640 (1971)
Schoeller, J. : Recherches descriptives et exp6rimentales sur la c6phalogen+se de Calliphora eryth- rocephala (Meigen), au cours des d~veloppements embryonnaire et postembryonnaire. Arch. Zool. Exp. g6n. 103, 1-216 (1964)
Snodgrass, R.E.: Principles of Insect Morphology. New York and London: McGraw-Hill Book Co. 1935
Spreij, Th.E., Oldenhave, M. : A detailed map of the wing disk of Calliphora erythrocephala. Neth. J. Zool. 24, 291-310 (1974)
Struhl, G.: Developmental compartments in the proboscis of Drosophila. Nature 270, 723-725 (1977)
Ursprung, H. : In vivo culture of Drosophila imaginal discs. In: Methods in Developmental Biology (F.H. Wilt and N.K. Wessels, eds.), pp. 485492. New York: Crowell 1967

64 K. Kumar et al.
Ursprung, H. : The fine structure of imaginal disks. In: The Biology of Imaginal Disks (H. Ursprung and R. N6thiger, eds). Results and Problems in Cell Differentiation 5, 93-107 (1972)
Wildermuth, H. : Differenzierungsleistungen, Mustergliederung und Transdeterminationsmechanis- men in hetero- und homoplastischen Transplantaten der R~sselprimordien von Drosophila. Wil- helm Roux' Arch. Entwickl.-Mech. Org. 160, 41-75 (1968)
Wildermuth, H., Hadorn, E. : Differenzierungsleistungen der LabiaMmaginalscheibe yon Drosophila melanogaster. Rev. Suisse Zool. 72, 686-694 (1965)
Wolpert, L. : Positional information and pattern formation. Curr. Top. Develop. Biol. 6, 183-224 (1971)
Received April 19, 1978 / Accepted in revised form October 30, 1978





























