Embed Size (px)
Citation preview

Ecological Applications, 24(4), 2014, pp. 663–679� 2014 by the Ecological Society of America
Disturbance, life history traits, and dynamics in an old-growth forestlandscape of southeastern Europe
THOMAS A. NAGEL,1,4 MIROSLAV SVOBODA,2 AND MILAN KOBAL3
1University of Ljubljana, Biotechnical Faculty, Department of Forestry and Renewable Forest Resources, Vecna pot 83,1000 Ljubljana, Slovenia
2Czech University of Life Sciences Prague, Faculty of Forestry and Wood Sciences, Kamycka 129,Praha 6 - Suchdol 16521 Czech Republic
3Slovenian Forestry Institute, Vecna pot 2, 1000 Ljubljana, Slovenia
Abstract. Much of our understanding of natural forest dynamics in the temperate regionof Europe is based on observational studies in old-growth remnants that have emphasizedsmall-scale gap dynamics and equilibrium stand structure and composition. Relatively littleattention has been given to the role of infrequent disturbance events in forest dynamics. In thisstudy, we analyzed dendroecological data from four stands and three windthrow patches in anold-growth landscape in the Dinaric Mountains of Bosnia and Herzegovina to examinedisturbance history, tree life history traits, and compositional dynamics. Over all stands, mostdecades during the past 340 years experienced less than 10% canopy loss, yet each standshowed evidence of periodic intermediate-severity disturbances that removed .40% of thecanopy, some of which were synchronized over the study area landscape. Analysis of radialgrowth patterns indicated several life history differences among the dominant canopy trees;beech was markedly older than fir, while growth patterns of dead and dying trees suggestedthat fir was able to tolerate longer periods of suppressed growth in shade. Maple had thefastest radial growth and accessed the canopy primarily through rapid early growth in canopygaps, whereas most beech and fir experienced a period of suppressed growth prior to canopyaccession. Peaks in disturbance were roughly linked to increased recruitment, but mainly ofshade-tolerant beech and fir; less tolerant species (i.e., maple, ash, and elm) recruitedsuccessfully on some of the windthown sites where advance regeneration of beech and fir wasless abundant. The results challenge the traditional notions of stability in temperate old-growth forests of Europe and highlight the nonequilibrial nature of canopy composition dueto unique histories of disturbance and tree life history differences. These findings providevaluable information for developing natural disturbance-based silvicultural systems, as well asinsight into maintaining less shade-tolerant, but valuable broadleaved trees in temperateforests of Europe.
Key words: Abies alba; dendroecology; Dinaric Mountains, Bosnia-Herzegovina; Fagus sylvatica;forest dynamics; intermediate severity disturbance; longevity; natural disturbance; old-growth forest; shadetolerance; species coexistence.
INTRODUCTION
After centuries of deforestation and soil degradation
in temperate forests of Europe, followed by widespread
planting of coniferous monocultures during the 20th
century, society is currently seeking ecologically based
forestry practices that maintain ecosystem services,
preserve native biodiversity, and enhance resilience to
climate change (Bengtsson et al. 2000, Spiecker 2003,
Schroter et al. 2005). Much emphasis has therefore been
placed on restoration of old-growth forest structure and
composition (Bauhus et al. 2009), the idea being to
create conditions under which native biodiversity
evolved, as well as promotion of complex age classes
and species mixtures at stand and landscape scales, a
strategy that may foster adaptation to future climate
changes (Noss 2001, Millar et al. 2007). Restoring such
conditions typically involves management approaches
that incorporate patterns and processes found in old-
growth forests, such as maintaining a sufficient supply of
dead wood (Christensen et al. 2005, Muller and Butler
2010) and harvesting schemes that emulate natural
disturbance processes and increase structural heteroge-
neity (Angelstam 1998, Seymour et al. 2002, Keeton
2006).
A major obstacle to developing such approaches for
the European temperate region, however, is the lack of
reference conditions where old-growth pattern and
process can be studied. Millennia of land use practices
have eliminated all but a few scattered remnants of old-
growth forest, which are primarily located in remote
mountain regions of eastern and southeastern Europe,
Manuscript received 5 April 2013; revised 11 September2013; accepted 13 September 2013. Corresponding Editor: E.Cienciala.
4 E-mail: [email protected]
663

where forests are typically dominated by mixtures of
beech (Fagus sylvatica) and fir (Abies alba). Much of our
current understanding of old-growth mountain forests
in Europe is based on early studies in these remaining
remnants (e.g., Leibundgut 1959, Zukrigl et al. 1963,
Korpel 1982, Mayer et al. 1989, Korpel 1995). The
conceptual model that emerged from this work suggests
that dynamics are driven by endogenous mortality of
canopy trees (i.e., small-scale gap dynamics), giving rise
to a steady-state system at stand scales, characterized by
a shifting, fine-scale mosaic of different stages of forest
development.
Recent work in many of these same old-growth forests
challenges this conceptual model, and highlights the role
of natural disturbances as a driver of ecosystem change.
Studies in old-growth beech and mixed beech–fir
communities of the Alps, Carpathian, and Dinaric
Mountains indicate that periodic intermediate-severity
damage from wind disturbances (i.e., single events that
cause stand-level damage ranging from scattered single
tree falls to larger openings several thousand square
meters in size) is an important component of the
disturbance regime in this region (Splechtna et al.
2005, Nagel and Diaci 2006, Nagel and Svoboda 2008,
Firm et al. 2009, Kucbel et al. 2010). Compared with the
continuous formation of small-scale gaps (e.g., ,100
m2), such disturbances play a key role in maintaining
biodiversity in temperate forest ecosystems by creating
biological legacies and increasing understory light levels
(Woods 2004, Hanson and Lorimer 2007, D’Amato et
al. 2008), and are likely to prevent forest stands from
reaching an equilibrium state (Frelich and Lorimer
1991, Woods 2000, 2004, Worrall et al. 2005). Although
empirical studies of disturbance regimes are increasing
in the temperate zone of Europe, it is still difficult to
make generalizations due to the lack of geographic
representation and methodological limitations. For
example, most recent research has examined disturbance
regimes by quantifying characteristics of recently formed
canopy gaps (Nagel and Svoboda 2008, Kucbel et al.
2010), an approach that covers a limited time span, or
with dendroecological data (Splechtna et al. 2005, Nagel
et al. 2007, Motta et al. 2011, Trotsiuk et al. 2012),
which allows reconstruction of disturbance patterns
several centuries past, but typically over a limited spatial
extent.
Furthermore, while these recent studies have yielded
valuable insight into disturbance regimes, few have
examined the role of disturbances on tree community
dynamics, particularly with regard to maintenance of
tree species diversity. In large part, our current
understanding of tree diversity in old-growth forests of
Europe is based on a floristic–phytosociological ap-
proach, whereby site conditions, rather than past
disturbances, are thought to control compositional
patterns (Braun-Blanquet 1964, Ellenberg 1988). Recent
research, however, suggests that intermediate-severity
disturbance in mixed beech–fir forests may be necessary
for canopy recruitment of more light-demanding species,
including maple (Acer pseudoplatanus), ash (Fraxinusexcelsior), and elm (Ulmus glabra) (Firm et al. 2009,
Nagel et al. 2010). What is lacking is a mechanisticunderstanding of community dynamics that links
natural disturbances and tree life history traits (e.g.,Lusk and Smith 1998, Bergeron 2000, Loehle 2000,Gutierrez et al. 2008). This is probably because there are
few empirical studies that quantify basic life historycharacteristics, such as shade tolerance, growth, and
longevity, of common temperate tree species in Europe(e.g., Kunstler et al. 2005, Petritan et al. 2007). Much of
what we know about shade tolerance and longevity, forexample, comes from early studies that relied mainly on
observational evidence (Ellenberg 1988, Korpel 1995).In this study we examined the disturbance regime, life
history traits, and compositional dynamics in an old-growth beech–fir forest landscape in the Dinaric
Mountains of Bosnia-Herzegovina. We used dendroeco-logical data to reconstruct patterns of disturbance in
four stands and three windthrow patches; by using aspatially explicit dendroecological approach that esti-
mates the area of past disturbances (Frelich andGraumlich 1994), we were able to reconstruct both the
frequency and severity of disturbances over the pastseveral centuries. Moreover, because our sampling wasdispersed over a relatively large area of old growth, the
results yield important information on stand andlandscape patterns of disturbance not captured by
previous dendroecological studies in the region. Weused the same dendroecological data set to provide
insight into interspecific differences in tree life historytraits, namely shade tolerance, growth rates, and
longevity. These data allowed us to address thefollowing questions relevant to the general understand-
ing of forest dynamics in the region. (1) Is thedisturbance regime consistent with the widely accepted
model of small-scale gap dynamics and fine-scaleuneven-aged stands of mixed mountain old-growth
forests in Europe? (2) Can interspecific differences intree life history traits, coupled with the reconstructed
history of disturbance, explain tree community compo-sition? Finally, we discuss how the results can informforest management in the temperate zone of Europe,
particularly in landscapes where maintenance of ecolog-ical functions is a management priority.
METHODS
Study area and sample sites
This study was conducted in the Peru�cica forestreserve in the Dinaric Mountains of Bosnia-Herzegovi-
na. With a size of ;1400 ha, Peru�cica is one of thelargest tracts of old-growth forest remaining in the
temperate zone of Europe. In the Dinaric Mountainregion, for example, most of the remaining old-growthforests are small (,100-ha) remnants (Diaci 1999),
making it difficult to capture the natural range ofvariability of disturbance processes. Thus, the large size
THOMAS A. NAGEL ET AL.664 Ecological ApplicationsVol. 24, No. 4

of Peru�cica allows a unique glimpse into natural
disturbances patterns and forest dynamics. The study
region is influenced by both mediterranean and conti-
nental climates, with a mean monthly temperature range
of �3.38C in January to 15.08C in July and a mean
annual precipitation of 1837 mm (Cemerno station,1305
m above sea level). Sampling was done between
approximately 1000 and 1400 m in elevation, where
beech and fir dominate the canopy layer. Several less
frequent species co-occur in the canopy, including
maple, elm, ash, and Norway spruce (Picea abies). The
forest has typical old-growth features, including large
amounts of standing and downed coarse woody debris,
heterogeneous stand structure, and large canopy trees
reaching heights between 40 and 50 m (Nagel and
Svoboda 2008, Nagel et al. 2010). Topography in the fir–
beech zone is characterized by deeply dissected, moder-
ate to steep slopes that surround the Peru�cica river
watershed. Soils are generally fertile and deep, derived
mainly from limestone and dolomite on the upper-
elevation slopes and a mixture of acidic sandstone and
shale at lower elevations (Fukarek and Stefanovic 1958).
The study region has certainly been influenced by
historic land use practices, but there is no evidence that
they were responsible for any of the patterns document-
ed in this study. In Peru�cica, there is no archival
evidence of historical cutting in the reserve (Fukarek
and Stefanovic 1958). In fact, more than half of the total
2 million ha of forests in Bosnia were characterized as
old-growth prior to the Austro-Hungarian rule begin-
ning in 1878 (Frohlich 1954). Given the low population
density in the region during the study period (Palairet
1997) and inaccessible nature of the terrain prior to the
road that was constructed in the 1950s, we have no
reason to believe that widespread cutting would have
occurred in Peru�cica. However, transhumant grazing
has probably occurred for centuries, but was mainly
limited to the alpine pastures above the forest reserve.
Our interviews with the few remaining shepherds still
practicing transhumance in the area suggest that sheep
and cattle rarely entered the forest, except under very
dry conditions when animals would use the forest edge.
Given that our plots were placed in the interior area of
the reserve, far from the alpine pastures that were used
for grazing, it is unlikely that historical grazing practices
influenced the study plots.
To capture landscape-scale patterns of disturbance
and the gradient in canopy composition encountered in
the study area, we divided the fir–beech zone into two
broad areas (Tunjemir and Zanoglina) located on
opposite sides of the watershed for sampling. Forest
structure and composition in the Tunjemir and Zano-
glina areas were extensively surveyed in a previous study
(Nagel et al. 2010), and can be summarized as follows.
The upper canopy (i.e., trees with dbh .30 cm) in
Tunjemir was dominated by beech (103 trees/ha),
whereas fir was less abundant (47 trees/ha). The upper
canopy in Zanoglina was mainly composed of fir (168
trees/ha) with a lower density of beech (44 trees/ha).
Maple, elm, ash, and spruce were sporadically distrib-
uted in the upper canopy layer in both areas at low
densities (i.e., ,3 trees/ha). Throughout both areas,
regeneration was sparse and dominated by beech; other
species had much lower densities (Appendix A).
Within each area, we used random coordinates to
locate two 1-ha plots (100 3 100 m), but restricted the
distance between the plots to at least 400 m (Fig. 1,
Table 1). In several cases, plots were shifted along the
slope contour to avoid sampling on intermittent streams
with steep banks. Additionally, during reconnaissance
for the four disturbance history plots, we encountered
several larger (�0.5 ha) windthrows (relative to the
smaller gaps that form regularly in the forest). These
patches were characterized by abundant tip-up mounds,
downed wood, and scattered windfirm canopy trees. We
selected three of the largest patches for further study
(Fig. 1, Table 1). Finally, throughout the entire beech–fir
zone, whenever we encountered a tree in the upper
canopy (typically .30 m in height) of a less frequent tree
species (i.e., maple, ash, and elm), we extracted an
increment core at a height of 1 m for growth pattern
analysis. The mean dbh measurements of the maple, ash,
and elm trees sampled in this study were 79 cm, 96 cm,
and 108 cm, respectively.
Field procedures
Disturbance history.—Disturbance history sampling
was conducted in 2006 and 2007 in the 1-ha plots. Each
plot was divided into 10310 m grid squares and the tree
stem (with dbh �10 cm) closest to the center of each
quadrat was cored at a height of 1 m and parallel to the
slope contour to avoid reaction wood. These trees were
later used for age structure analysis. If the closest stem
was not in the canopy, defined as trees receiving direct
sunlight from above, regardless of height (Lorimer and
Frelich 1989), the closest canopy tree to the quadrat
center was also cored. Canopy trees were used to
reconstruct disturbance history. When a tree was too
rotten to extract a useable sample, we were usually able
to obtain an intact sample at a higher coring height (up
to 2 m). In the few rare cases when the sample tree was
very decayed, we cored another stem of the same species
and of similar size within the quadrat. In quadrats
located under recently formed gaps with no trees of dbh
�10 cm, no core was taken. Rather, to provide
information on the recent history of disturbance on
the plots, we mapped the shape and position of canopy
gaps and snapped and uprooted trees with the aid of the
103 10 m grid. We also recorded the position of old pits
and mounds to supplement the dendroecological recon-
struction of disturbance. In total, we collected 582 cores
over the four plots for the analysis of age structure and
growth patterns.
Windthrow patches.—After delineating the approxi-
mate boundary of each windthrow patch, we established
three 625 m2 (25 3 25 m) plots separated by 10-m
June 2014 665EUROPEAN OLD-GROWTH FOREST DISTURBANCE

intervals along a transect oriented through the center of
each patch. Similarly, three 625-m2 plots were also
placed along a transect in a control stand adjacent to
each windthrow that did not show evidence of wind
damage. In each plot, the species and dbh of all live and
dead (standing and down) trees with dbh �5 cm were
recorded, as well as the number of seedlings (.0.5 m tall
but ,1.3 m tall) and saplings (.1.3 m tall but ,5 cm
dbh) of each species. To provide additional insight into
post-windthrow successional processes, in each wind-
throw plot we extracted increment cores from three to
five of the trees with dbh �5 cm nearest to the plot
center for each tree species present in the plots. Cores
were extracted at ;10 cm in height to help determine the
timing of tree establishment relative to each windthrow
event. No cores were sampled in the control areas.
To help characterize the disturbance history of each
windthrow patch, we recorded the species, dbh, and
stage of decay (following Nagel and Svoboda 2008) of
all snapped and uprooted boles in both windthrow and
FIG. 1. Map of the Peru�cica forest reserve in the Sutjeska National Park, Bosnia-Herzegovina; circles show the location of thefour disturbance history plots in Tunjemir (T1, T2) and Zanoglina (Z1, Z2); triangles show the three windthrow patches (Osoje,Tunjemir, Skakavac).
TABLE 1. Site and stand characteristics for the four old-growth Fagus–Abies plots and three windthrow patches in the Peru�cicaforest reserve, Bosnia-Herzegovina.
Characteristic
Disturbance history plots Windthrow patches
Tunjemir-1 Tunjemir-2 Zanoglina-1 Zanoglina-2 Osoje Tunjemir Skakavac
Map ID T1 T2 Z1 Z2 1 2 3Elevation (m) 1150 1200 1340 1400 1200 1240 1100Slope (8) 25–30 10–25 0–25 5–25 15–20 15–20 15–20Aspect (8) 40 40 240 260 340 350 360Size (ha) 1 1 1 1 0.5 .1.0 0.6Gap fraction (%) 17 43 22 27Canopy composition (%)
Fagus sylvatica 83 44 31 43 53 67 68Abies alba 17 56 69 57 40 30 32
Notes: Only the dominant two species, F. sylvatica and A. alba, are shown for canopy composition. The composition of thewindthrow areas is based on undisturbed control plots (see Methods: Windthrow areas). Size refers to the plot size for thedisturbance history plots and patch size for the windthrows.
THOMAS A. NAGEL ET AL.666 Ecological ApplicationsVol. 24, No. 4

control plots. Finally, in addition to the cores collected
within plots, we extracted increment cores from at least
three large, windfirm trees in the upper canopy within or
near each windthrow plot (;10 trees per windthrow
patch) to help date the disturbances.
Tree growth histories.—In addition to the reconstruc-
tion of age structure and disturbance history, we
examined patterns of tree ring growth indicative of life
history differences among the common tree species
found in the study area. This data set included all of the
increment cores from the 1-ha plots (N ¼ 582), the
windthrow patches (N ¼ 108), the less frequent tree
species encountered throughout the study area (N¼ 43),
and samples from additional ‘‘winner’’ and ‘‘loser’’ trees
of beech and fir cored in 2009 (N ¼ 121). We targeted
winners and losers to compare the growth history of
trees that reach the canopy with that of trees that die in
the shaded understory. Winners were defined as trees in
the lower part of the upper canopy (;20 m tall) in the
process of filling a gap. In other words, these were
intermediate-sized trees with crowns exposed to full sun
from above that were likely to grow into the upper
canopy. In contrast, loser trees were defined as the
largest recently dead or morbid trees (i.e., trees with
plageotropic growth and only a small amount of living
foliage) growing in the shaded understory and presumed
to be dead or morbid due to shading from above rather
than overcrowding. These trees typically had a dbh .5
cm and were ,20 m tall. Sampling was carried out in
four belt transects (;40 3 200 m) placed parallel to the
slope contour starting from the edge of each 1-ha plot
used for the disturbance history reconstruction. In each
transect, we extracted increment cores and recorded the
species and dbh for all winner and loser trees of both fir
and beech. Cores were sampled at a height of 1 m and
parallel to the slope contour. Over the four transects, we
collected 60 cores of beech and 61 of fir, with
approximately equal sample sizes of winners and losers
for both species.
Data analysis
Dendroecological procedures.—In the laboratory,
cores were dried, mounted, and sanded to a high polish
using standard procedures. Samples were then digitized
with the ATRICS system (Levanic 2007), and annual
ring widths were measured to the nearest 0.01 mm using
WinDENDRO software (Regents Instruments, Quebec,
Canada). The tree ring series were visually cross-dated
using marker years (Yamaguchi 1991) and by compar-
ison with a master chronology developed from a
drought-sensitive black pine (Pinus nigra) stand within
the same watershed as the study site (Poljansek et al.
2012). Cross-dating accuracy was verified with the
COFECHA program (Holmes 1983). We were not able
to successfully cross-date early portions of some samples
because of extended periods of very suppressed initial
growth. Such samples were nevertheless retained in the
analysis because they contained important information
on disturbance history and growth patterns. Most of the
cores either included or were less than 3 cm from the
pith (60%, ,1 cm; 85%, ,3 cm). For samples that did
not include the pith, the number of missing rings was
extrapolated from the curvature and average growth
rate of the innermost 10 rings (Duncan 1989).
Reconstruction of past disturbances.—Radial growth
patterns of all increment cores collected in the study
were analyzed for evidence of past disturbance events.
This analysis was used to construct disturbance chro-
nologies for the 1-ha plots and windthrow patches, as
well as to examine interspecific differences in tree growth
histories. Specifically, each core was checked for (1)
abrupt increases in radial growth (i.e., releases) and (2)
rapid early growth rates (i.e., gap-recruited trees), both
of which indicate mortality of a former canopy tree
(Lorimer and Frelich 1989). To determine if trees
qualified for gap recruitment, we defined species-specific
growth-rate thresholds as the 95th percentile growth rate
of suppressed trees sampled in the disturbance history
plots (Splechtna et al. 2005, Firm et al. 2009). This gave
a growth-rate threshold of 1.16 mm/yr for fir and 1.0
mm/yr for beech. Trees were considered gap-recruited if
the mean ring width of the initial 10 years of growth
exceeded the threshold value. In a few cases, trees that
did not meet the growth rate threshold, but had a
lifelong declining growth pattern, were also categorized
as gap origin (Lorimer and Frelich 1989). Gap
recruitment was only determined for cores that missed
the pith by less than 3 cm.
Releases were identified using the boundary line
technique (Black and Abrams 2003). The advantage of
this method is that it scales release detection according
to the pre-disturbance growth rate. Potential releases,
which are later scaled to species-specific boundary lines,
were identified by first calculating percentage growth
change values in each tree ring series with the running
mean method of Nowacki and Abrams (1997), which
compares consecutive 10-year growth intervals. The
maximum value of each growth pulse (pulses were
defined as positive, sequential percentage growth chang-
es values) was then selected as the potential release year.
To minimize the detection of short-term growth
responses caused by factors other than loss of canopy
trees, such as climate extremes or mast years, we only
used potential releases from pulses with percentage
growth change exceeding 50% (Splechtna et al. 2005,
Svoboda et al. 2012). These potential releases were
scaled relative to a boundary line constructed for both
fir and beech using tree ring measurements from all of
the increment cores sampled in the study (see Appendix
B for more detail on the species-specific boundary line
functions). Based on the original threshold of Black and
Abrams (2003), all releases within 20–100% of the
boundary line were tallied. Finally, to reduce the
number of falsely identified releases detected by the
boundary line, all releases for each sample were
subjected to additional visual inspection. Releases were
June 2014 667EUROPEAN OLD-GROWTH FOREST DISTURBANCE

deemed false if they lasted less than 10 years or were
caused by several narrow rings in a 10-year series, such
as growth reductions due to mast years of beech
(Drobyshev et al. 2010).
The release and gap recruitment events were used to
construct disturbance chronologies for each disturbance
history plot and windthrow patch. For the disturbance
history plots, the chronologies only included gap
recruitment events and growth releases indicative of
canopy accession (Lorimer and Frelich 1989). Namely,
only disturbance events that allowed trees access to the
canopy were included. The goal of this approach was to
exclude releases in existing canopy trees due to crown
expansion into a gap and moderate releases of under-
story trees caused by nearby gaps, because including
such events would overestimate the amount of canopy
area removed during a disturbance event. To distinguish
canopy accession events from releases in existing canopy
trees, we used species-specific diameter thresholds to
determine if a tree was in the canopy at the time of a
given disturbance event. Based on the approach of
Lorimer and Frelich (1989), data from the size structure
of canopy and understory trees in the disturbance
history plots were used to calculate the thresholds.
Releases were not counted for fir with dbh �28 cm and
beech with dbh �18 cm at the time of disturbance.
Following visual inspection of each tree ring series,
releases below these thresholds were counted as evidence
of canopy accession. When samples had multiple
releases of similar magnitude that met our criteria, each
release was tallied. We allowed for multiple releases
because understory trees often require several distur-
bance events to reach the canopy (Lorimer and Frelich
1989, Canham 1990).
Because canopy trees were cored at regular grid points
in the disturbance history plots, disturbance chronolo-
gies represent the area of the plots affected by new gap
formation in each decade, and were estimated by (x/
n)(100), where x was the number of trees showing a
canopy accession during a given decade and n was the
number of trees in the sample (Frelich and Graumlich
1994). To partially reduce the uncertainty in the
disturbance history interpretation due to the loss of
information farther back in time, we truncated chronol-
ogies when the sample size dropped below 10 trees
(Fraver et al. 2009). Furthermore, disturbance rates in
the last decade of the chronologies, 1990–1999, may be
underestimates because the method used to detect
releases requires a 10-year post-disturbance window,
and because trees recruited by disturbance events may
not have been large enough to core during the time of
sampling. For the windthrow patches, where we
collected a smaller number of cores, all gap recruitment
events and growth releases were included in disturbance
chronologies, regardless of canopy status.
Tree growth histories.—We quantified a variety of
lifetime growth characteristics that are indicative of tree
life history characteristics, namely, shade tolerance,
radial growth rates, and longevity. These included
statistics on periods of minimum and maximum growth
calculated from 10-year average growth periods (Orwig
and Abrams 1994) and the maximum duration of
sequential fast and slow growth per core, defined as
the number of consecutive years in which radial growth
was above (fast growth) or below (slow growth) species-
specific growth rate thresholds, calculated as the 75th
and 25th percentile ring widths, respectively, from all of
the ring width data for each species in the study (Baker
and Bunyavejchewin 2006). We also examined release
frequency and canopy accession patterns (i.e., gap origin
without a subsequent release; gap origin followed by one
or several releases; and non-gap origin followed by one
or several releases) among the dominant species. Life
span of the dominant species was estimated from the
90th percentile of age distributions of trees with dbh
.50 cm. When appropriate, calculations were only
carried out with complete cores, that is, cores that
included or missed the pith by less than 3 cm. All
dendroecological and statistical analyses were per-
formed in the statistical environment R (R Development
Core Team 2011).
RESULTS
Disturbance history.—Disturbance chronologies for
the four disturbance history plots show considerable
temporal variability in canopy loss over the past three
centuries (Fig. 2; Appendix C). Although there was no
clear evidence of stand-replacing disturbance, all of the
chronologies showed marked peaks, with maximum
canopy loss in a given decade ranging from 45% to 77%across the four plots. The mean rate of disturbance
ranged between 8.5% and 10% canopy area loss per
decade among the plots, yet the distribution of decadal
disturbance rates was skewed; with all plots pooled, 68%of decadal rates were below 10% canopy loss, 26%between 10% and 30%, 4% between 30% and 50%, and
2% greater than 50%. Therefore, the overall median
decadal rate of disturbance (5%) across the four plots
may be a better indication of the disturbance rate.
Rotation periods calculated from the pooled decadal
rates of all four plots (following Fraver et al. 2009)
provide an indication of the time needed for all plots to
experience a given disturbance rate: rates of �20%, 30%,
40%, and 50% canopy loss recur approximately every 90,
150, 200, and 460 years, respectively.
The age structures of the four plots were uneven-aged
and irregular, with pronounced peaks that roughly
followed periods with higher rates of disturbance (Fig.
3). For example, notable recruitment pulses followed
disturbance peaks in the 1660s and 1760s in Tunjemir 1,
the 1950s in Tunjemir 2, and the 1700s and 1820s in
Zanoglina-2. Recruitment dynamics of beech and fir
also show markedly different patterns among the four
plots. Some disturbance peaks were followed by
pronounced recruitment pulses of beech (e.g., the
1660s event in Tunjemir-1 and the 1950s event in
THOMAS A. NAGEL ET AL.668 Ecological ApplicationsVol. 24, No. 4
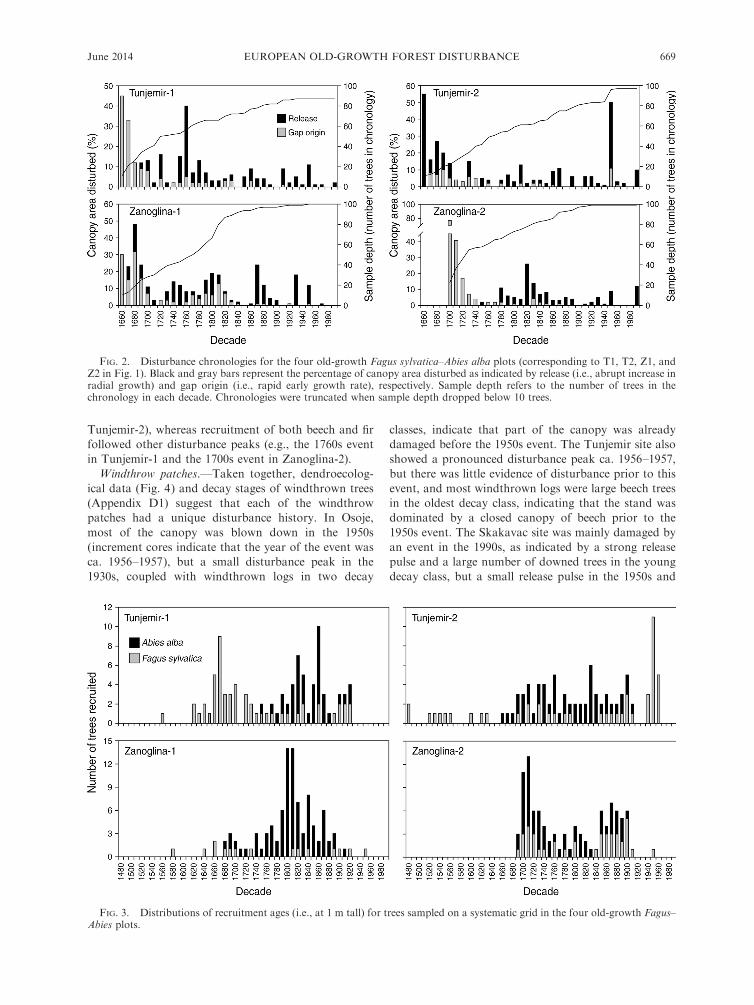
Tunjemir-2), whereas recruitment of both beech and fir
followed other disturbance peaks (e.g., the 1760s event
in Tunjemir-1 and the 1700s event in Zanoglina-2).
Windthrow patches.—Taken together, dendroecolog-
ical data (Fig. 4) and decay stages of windthrown trees
(Appendix D1) suggest that each of the windthrow
patches had a unique disturbance history. In Osoje,
most of the canopy was blown down in the 1950s
(increment cores indicate that the year of the event was
ca. 1956–1957), but a small disturbance peak in the
1930s, coupled with windthrown logs in two decay
classes, indicate that part of the canopy was already
damaged before the 1950s event. The Tunjemir site also
showed a pronounced disturbance peak ca. 1956–1957,
but there was little evidence of disturbance prior to this
event, and most windthrown logs were large beech trees
in the oldest decay class, indicating that the stand was
dominated by a closed canopy of beech prior to the
1950s event. The Skakavac site was mainly damaged by
an event in the 1990s, as indicated by a strong release
pulse and a large number of downed trees in the young
decay class, but a small release pulse in the 1950s and
FIG. 3. Distributions of recruitment ages (i.e., at 1 m tall) for trees sampled on a systematic grid in the four old-growth Fagus–Abies plots.
FIG. 2. Disturbance chronologies for the four old-growth Fagus sylvatica–Abies alba plots (corresponding to T1, T2, Z1, andZ2 in Fig. 1). Black and gray bars represent the percentage of canopy area disturbed as indicated by release (i.e., abrupt increase inradial growth) and gap origin (i.e., rapid early growth rate), respectively. Sample depth refers to the number of trees in thechronology in each decade. Chronologies were truncated when sample depth dropped below 10 trees.
June 2014 669EUROPEAN OLD-GROWTH FOREST DISTURBANCE
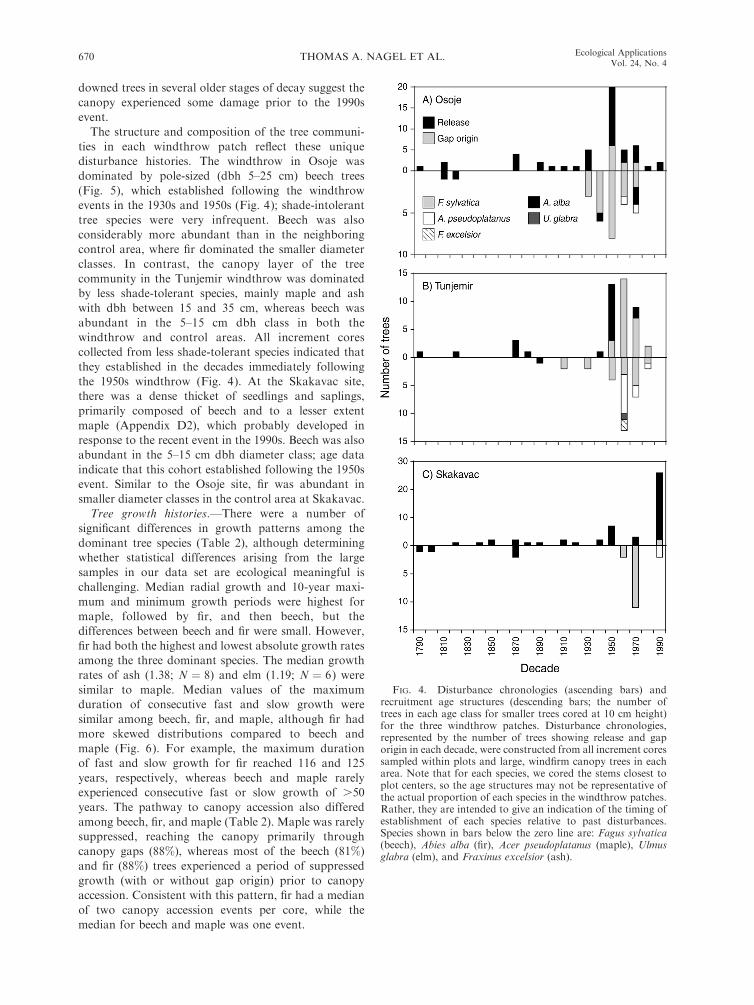
downed trees in several older stages of decay suggest the
canopy experienced some damage prior to the 1990s
event.
The structure and composition of the tree communi-
ties in each windthrow patch reflect these unique
disturbance histories. The windthrow in Osoje was
dominated by pole-sized (dbh 5–25 cm) beech trees
(Fig. 5), which established following the windthrow
events in the 1930s and 1950s (Fig. 4); shade-intolerant
tree species were very infrequent. Beech was also
considerably more abundant than in the neighboring
control area, where fir dominated the smaller diameter
classes. In contrast, the canopy layer of the tree
community in the Tunjemir windthrow was dominated
by less shade-tolerant species, mainly maple and ash
with dbh between 15 and 35 cm, whereas beech was
abundant in the 5–15 cm dbh class in both the
windthrow and control areas. All increment cores
collected from less shade-tolerant species indicated that
they established in the decades immediately following
the 1950s windthrow (Fig. 4). At the Skakavac site,
there was a dense thicket of seedlings and saplings,
primarily composed of beech and to a lesser extent
maple (Appendix D2), which probably developed in
response to the recent event in the 1990s. Beech was also
abundant in the 5–15 cm dbh diameter class; age data
indicate that this cohort established following the 1950s
event. Similar to the Osoje site, fir was abundant in
smaller diameter classes in the control area at Skakavac.
Tree growth histories.—There were a number of
significant differences in growth patterns among the
dominant tree species (Table 2), although determining
whether statistical differences arising from the large
samples in our data set are ecological meaningful is
challenging. Median radial growth and 10-year maxi-
mum and minimum growth periods were highest for
maple, followed by fir, and then beech, but the
differences between beech and fir were small. However,
fir had both the highest and lowest absolute growth rates
among the three dominant species. The median growth
rates of ash (1.38; N ¼ 8) and elm (1.19; N ¼ 6) were
similar to maple. Median values of the maximum
duration of consecutive fast and slow growth were
similar among beech, fir, and maple, although fir had
more skewed distributions compared to beech and
maple (Fig. 6). For example, the maximum duration
of fast and slow growth for fir reached 116 and 125
years, respectively, whereas beech and maple rarely
experienced consecutive fast or slow growth of .50
years. The pathway to canopy accession also differed
among beech, fir, and maple (Table 2). Maple was rarely
suppressed, reaching the canopy primarily through
canopy gaps (88%), whereas most of the beech (81%)
and fir (88%) trees experienced a period of suppressed
growth (with or without gap origin) prior to canopy
accession. Consistent with this pattern, fir had a median
of two canopy accession events per core, while the
median for beech and maple was one event.
FIG. 4. Disturbance chronologies (ascending bars) andrecruitment age structures (descending bars; the number oftrees in each age class for smaller trees cored at 10 cm height)for the three windthrow patches. Disturbance chronologies,represented by the number of trees showing release and gaporigin in each decade, were constructed from all increment coressampled within plots and large, windfirm canopy trees in eacharea. Note that for each species, we cored the stems closest toplot centers, so the age structures may not be representative ofthe actual proportion of each species in the windthrow patches.Rather, they are intended to give an indication of the timing ofestablishment of each species relative to past disturbances.Species shown in bars below the zero line are: Fagus sylvatica(beech), Abies alba (fir), Acer pseudoplatanus (maple), Ulmusglabra (elm), and Fraxinus excelsior (ash).
THOMAS A. NAGEL ET AL.670 Ecological ApplicationsVol. 24, No. 4
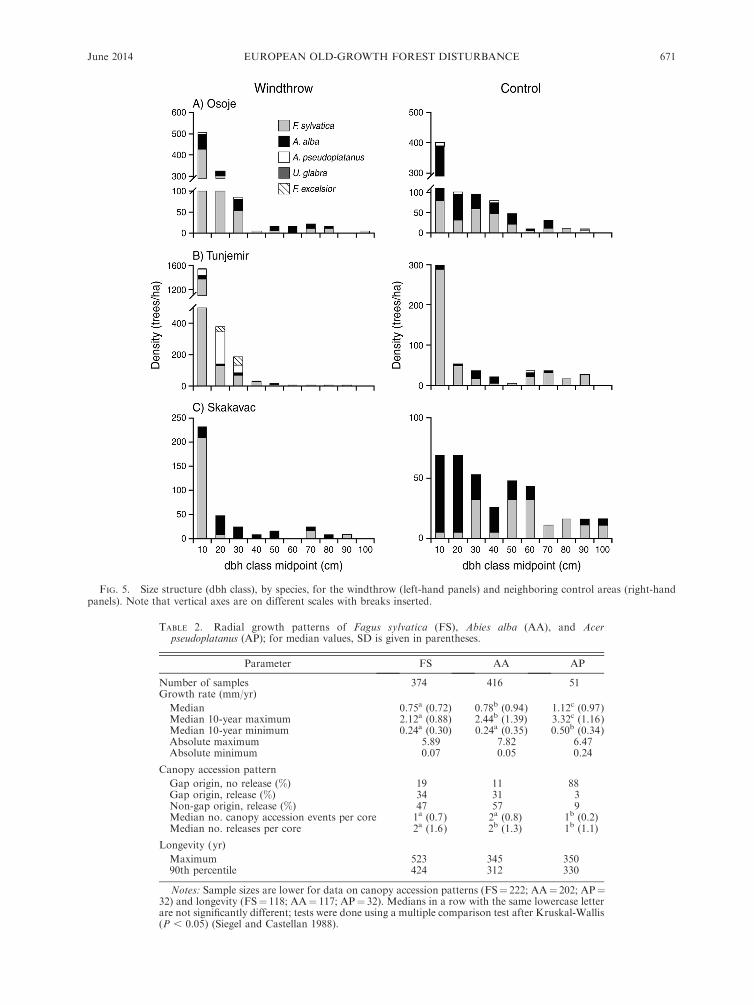
FIG. 5. Size structure (dbh class), by species, for the windthrow (left-hand panels) and neighboring control areas (right-handpanels). Note that vertical axes are on different scales with breaks inserted.
TABLE 2. Radial growth patterns of Fagus sylvatica (FS), Abies alba (AA), and Acerpseudoplatanus (AP); for median values, SD is given in parentheses.
Parameter FS AA AP
Number of samples 374 416 51Growth rate (mm/yr)
Median 0.75a (0.72) 0.78b (0.94) 1.12c (0.97)Median 10-year maximum 2.12a (0.88) 2.44b (1.39) 3.32c (1.16)Median 10-year minimum 0.24a (0.30) 0.24a (0.35) 0.50b (0.34)Absolute maximum 5.89 7.82 6.47Absolute minimum 0.07 0.05 0.24
Canopy accession pattern
Gap origin, no release (%) 19 11 88Gap origin, release (%) 34 31 3Non-gap origin, release (%) 47 57 9Median no. canopy accession events per core 1a (0.7) 2a (0.8) 1b (0.2)Median no. releases per core 2a (1.6) 2b (1.3) 1b (1.1)
Longevity (yr)
Maximum 523 345 35090th percentile 424 312 330
Notes: Sample sizes are lower for data on canopy accession patterns (FS¼ 222; AA¼ 202; AP¼32) and longevity (FS¼ 118; AA¼ 117; AP¼ 32). Medians in a row with the same lowercase letterare not significantly different; tests were done using a multiple comparison test after Kruskal-Wallis(P , 0.05) (Siegel and Castellan 1988).
June 2014 671EUROPEAN OLD-GROWTH FOREST DISTURBANCE
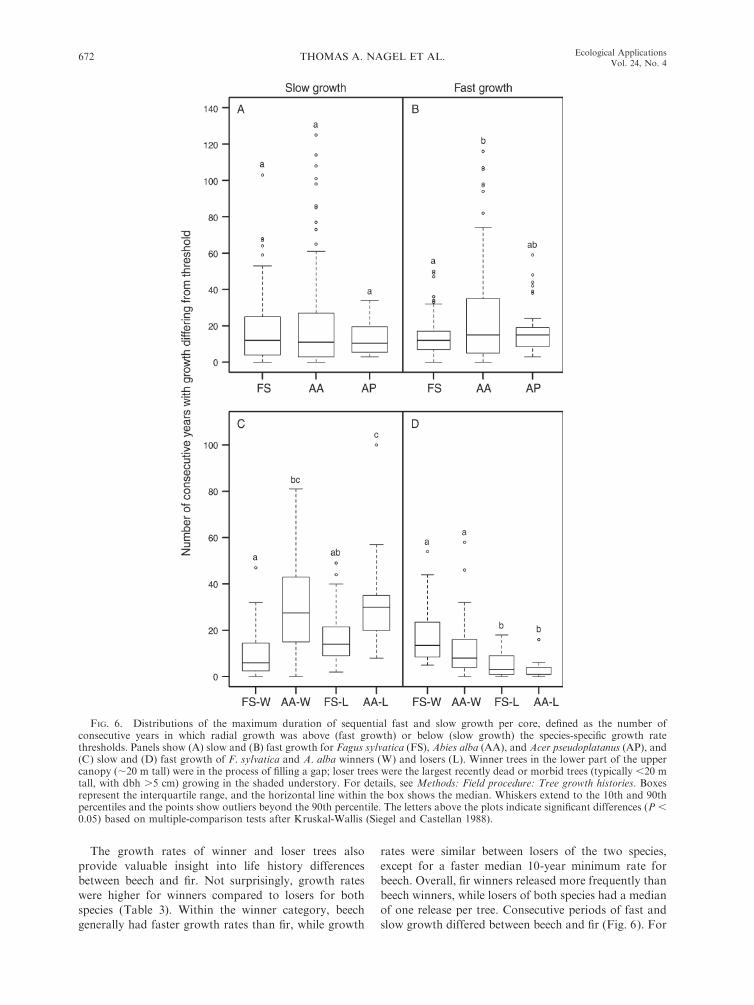
The growth rates of winner and loser trees also
provide valuable insight into life history differences
between beech and fir. Not surprisingly, growth rates
were higher for winners compared to losers for both
species (Table 3). Within the winner category, beech
generally had faster growth rates than fir, while growth
rates were similar between losers of the two species,
except for a faster median 10-year minimum rate for
beech. Overall, fir winners released more frequently than
beech winners, while losers of both species had a median
of one release per tree. Consecutive periods of fast and
slow growth differed between beech and fir (Fig. 6). For
FIG. 6. Distributions of the maximum duration of sequential fast and slow growth per core, defined as the number ofconsecutive years in which radial growth was above (fast growth) or below (slow growth) the species-specific growth ratethresholds. Panels show (A) slow and (B) fast growth for Fagus sylvatica (FS), Abies alba (AA), and Acer pseudoplatanus (AP), and(C) slow and (D) fast growth of F. sylvatica and A. alba winners (W) and losers (L). Winner trees in the lower part of the uppercanopy (;20 m tall) were in the process of filling a gap; loser trees were the largest recently dead or morbid trees (typically ,20 mtall, with dbh .5 cm) growing in the shaded understory. For details, see Methods: Field procedure: Tree growth histories. Boxesrepresent the interquartile range, and the horizontal line within the box shows the median. Whiskers extend to the 10th and 90thpercentiles and the points show outliers beyond the 90th percentile. The letters above the plots indicate significant differences (P ,0.05) based on multiple-comparison tests after Kruskal-Wallis (Siegel and Castellan 1988).
THOMAS A. NAGEL ET AL.672 Ecological ApplicationsVol. 24, No. 4

suppressed growth, the median period for fir winners
was nearly five times longer than for beech winners, and
suppression periods for fir losers were twice that of
beech losers. Among the losers, the longest period of
slow growth was 100 years for fir and 49 years for beech.
Interspecific differences were less pronounced for
consecutive periods of fast growth. Winners of both
species experienced longer periods of fast growth
compared to losers, and beech had longer periods of
fast growth than fir, although these differences were not
significant.
Estimated longevities were markedly different for
beech and fir (Table 2). The oldest beech trees were
.500 years old and large canopy dominants had a 90th
percentile age of 424 years, whereas fir did not exceed
350 years and had a 90th percentile age of 312 years. The
oldest maple was 350 years old (90th percentile, 330
years). Large canopy trees of ash and elm ranged from
253 to 331 (N ¼ 6) and 326 to 369 (N ¼ 3) years old,
respectively.
DISCUSSION
Disturbance regime.—The history of disturbance
during the past three centuries in the beech–fir stands
documented in this work exhibits substantial variability.
Although most decades experienced less than 10%
canopy loss, indicative of small-scale canopy distur-
bances (i.e., gap dynamics), all of the stands were
affected by periodic, intermediate-severity disturbances,
removing 20–50% of the canopy. Our data indicate that
rotation periods of disturbances spanning this range of
severity are 90 to 500 years, respectively. Although this
can only be treated as a rough estimate, as either a very
long time window or a large geographical area is needed
for accurate estimates (Frelich and Graumlich 1994),
these results suggest that intermediate-severity distur-
bances occur within the expected life span of a tree
cohort. Such disturbances are most likely to be related
to strong winds associated with intense, local thunder-
storms, one of the primary agents of disturbance in the
region (Nagel and Diaci 2006, Nagel et al. 2006, Nagel
and Svoboda 2008).
Although some of these events were only identified
within individual stands, there is evidence that somedisturbances were synchronized over the larger studyarea landscape. The four stands sampled for disturbance
history all show high rates of canopy loss from 1660 to1700, when most of the trees in the disturbancechronologies show evidence of gap origin (Fig. 2). This
was particularly pronounced in Zanoglina-2, wherethere were no trees that predated this period (Fig. 3).Although the sample depth was relatively low at this
time, the fact that all plots show a similar patternsuggests that there was an intense disturbance thatdamaged stands over the broader landscape. A wind-storm ca. 1956–1957 also caused widespread damage to
the broader study area, as indicated by the largedisturbance peak in Tunjemir-2 (damage to the canopyfrom this event is consistent with the high gap fraction in
this stand; Table 1 and Appendix E), several smallerpeaks in Tunjemir-1 and Zanoglina-1, and disturbancepeaks in the three windthrow patches.
Mortality patterns resulting from intermediate-sever-ity disturbances ranged from scattered, small gaps tolarger multi-tree openings and small patches of cata-
strophic damage (Appendices C and E). The 1950sstorm event well exemplifies the range of damage foundin the disturbance chronologies; this event removed
about half of the canopy trees throughout Tunjemir-1,creating a mosaic of interconnected gaps (Appendix E),and most of the canopy at the Tunjemir windthrow,
leaving only scattered, windfirm canopy trees. Theperiodic occurrence of intermediate-severity disturbanc-es found in this study is consistent with the emergingview that intermediate-severity wind disturbances are an
important component of the disturbance regime intemperate forests of Europe (Wolf et al. 2004, Nageland Diaci 2006, Nagel et al. 2007, Kucbel et al. 2010)
and North America (Frelich and Lorimer 1991, Green-berg and McNab 1998, Woods 2004, Fraver and White2005, Hanson and Lorimer 2007, D’Amato and Orwig
2008, Fraver et al. 2009, Stueve et al. 2011). Given thatevents in this range of severity probably have returntimes of about one to five centuries (Frelich and Lorimer
1991, Fraver et al. 2009) and damage trees at stand
TABLE 3. Radial growth patterns of winner and loser trees of Fagus sylvatica (FS) and Abies alba(AA); for median values, SD is given in parentheses.
Parameter
Winners Losers
FS AA FS AA
Number of samples 32 28 28 33Growth rate (mm/yr)
Median 0.80a (0.75) 0.55b (0.66) 0.40a (0.55) 0.40a (0.38)Median 10-year max 2.01a (0.68) 1.83a (0.84) 1.06a (0.68) 1.16a (0.34)Median 10-year min 0.34a (0.34) 0.17b (0.46) 0.18a (0.16) 0.13b (0.04)Absolute max 4.02 4.09 2.66 2.30Absolute min 0.10 0.09 0.08 0.09
Median no. releases per core 1a (0.9) 2b (1.2) 1a (0.8) 1a (0.9)
Note: See Table 2 for explanation of statistical tests. Different superscript lowercase lettersindicate significant differences.
June 2014 673EUROPEAN OLD-GROWTH FOREST DISTURBANCE

scales, we might expect a substantial portion of
temperate forest landscapes to be in some stage of
recovery from intermediate-severity wind damage
(Stueve et al. 2011).
Life history traits and community dynamics.—Our
dendroecological data are generally consistent with
earlier rankings of life history traits for European trees,
such as those from Ellenberg’s (1988, 1992) classic
studies, but there are some important differences that
warrant discussion. For example, Ellenberg character-
ized juvenile shade tolerance of beech and fir as very
high and maple as high, which is partly in accordance
with our findings; maple’s fast growth rate, limited
tolerance to suppression, and accession to the canopy
via gaps indicate that it is less shade tolerant than beech
and fir, which is in agreement with a recent study that
quantified the effects of shade on growth and mortality
of beech and maple (Petritan et al. 2007). However,
although the relative ranking of beech and fir regener-
ation as ‘‘very shade tolerant’’ may be reasonable, there
are several lines of evidence indicating that there may be
differences in their tolerance to shade during the pole-
size life stage. First, based on the size structure of trees
in the understory and those receiving direct light from
above in the four disturbance history plots, beech trees
with dbh ;10–20 cm were rarely growing in the shaded
understory, whereas fir trees in this size range were
frequently encountered beneath canopy cover. Second,
the growth pattern analysis of winner and loser trees
indicates that fir losers can tolerate substantially longer
periods of suppressed growth compared to beech,
whereas beech winners showed a trend toward faster
growth, longer periods of fast growth, and fewer releases
per core than fir winners. These patterns suggest that
there may be differences in the trade-off between
survival in shade and growth in gaps for these two
species, a trade-off often invoked to explain tree
coexistence in temperate forests (e.g., Kobe et al. 1995,
Pacala et al. 1996, Clark et al. 2010, Gravel et al. 2010).
However, without data on light levels or survival,
dendroecological data alone can only provide limited
insight into interspecific differences in shade tolerance.
We also found notable discrepancies between our
estimates of longevity and values reported in the
literature. Ellenberg (1988), for example, categorizes
the life span of beech and fir as intermediate (150–400
years) and maple as long lived (.400 years). Korpel
(1995), in his classic work on the ecology of old-growth
forests in the Carpathian region, gives estimates of 230
and 350 years for beech and fir, respectively, but these
were based on a small sample size. More recent studies
that report age structure data from old-growth beech–fir
stands in Bosnia-Herzegovina and Slovenia suggest
similar longevities for both species, at ;400 years (Firm
et al. 2009, Motta et al. 2011), and studies from old-
growth beech forests in the Italian Apennines and
Ukrainian Carpathians reported ages exceeding 500
years (Piovesan et al. 2005, Trotsiuk et al. 2012). Taken
together, our results and those of recent studies suggest
that beech may be longer lived than fir, which is very
much in contrast to the traditional forestry dogma in
Europe, which maintains that fir is substantially older
than beech. This thinking may stem from the large
diameter and height of adult fir trees; individuals .50 m
tall and with dbh .100 cm are not uncommon in old-
growth stands on productive sites, yet beech trees with
these dimensions would be exceptional.
Community dynamics observed in our study are
partly consistent with our results on disturbance history
and life history differences among the dominant tree
species, but not always. Based on shade tolerance alone
(and no dispersal limitation), we would expect higher
severity disturbances to promote recruitment of maple,
ash, and elm. Although we found partial support for this
in the three windthrow sites, the dynamics seem to be
more complex. For example, much of the current tree
canopy at the Tunjemir windthrow is dominated by
maple and ash that established after the 1950s wind-
throw event. In contrast, beech is much more abundant
than less shade-tolerant species at the Osoje and
Skakavac sites. Furthermore, fir was much less abun-
dant in the windthrow than in the neighboring control
areas at Osoje and Skakavac, where it dominated the
smaller diameter classes, lending further evidence that it
is capable of tolerating shaded understory conditions.
The different successional pathways found in the
windthrow patches seem to largely be the result of prior
disturbance, whereby low-severity canopy disturbance
before the last windthrow event (i.e., at Osoje and
Skakavac) enabled a bank of beech regeneration to
establish, which saturated the site and prevented the
recruitment of more light-demanding species even after
more widespread canopy removal. Such processes are
well documented in the stem exclusion stage of stand
development (Oliver and Larson 1996). Release of
advance, shade-tolerant regeneration following interme-
diate-severity disturbances, causing structural, but not
compositional change, has been documented in a
number of temperate forests (Webb and Scanga 2001,
Fraver and White 2005, Nagel et al. 2006, Beaudet et al.
2007, Fraver et al. 2009).
The presence of advance regeneration of beech and fir
may also explain the recruitment failure of less shade-
tolerant species within the four disturbance history plots
following the periods of heavy canopy removal.
Alternative explanations could be linked to a lack of
nearby adults (e.g., in the Tunjemir and Zanoglina
areas, average density of less tolerant species in the
canopy layer was ,3 trees/ha; Nagel et al. 2010). Our
sampling approach could have missed trees that
recruited to the canopy following past disturbances,
but died prior to the time of sampling, a common
problem with reconstructive studies that rely on static
age distributions (Johnson et al. 1994). Recruitment
patterns of beech and fir following the major distur-
bance peaks are also difficult to interpret; disturbance
THOMAS A. NAGEL ET AL.674 Ecological ApplicationsVol. 24, No. 4

peaks were followed by recruitment pulses dominated by
either beech, fir, or both species. It seems likely that
stand structure and composition at the time of these past
disturbances, coupled with the unique history of
disturbance in each stand, led to these variable
recruitment patterns. These results highlight the idio-
syncratic nature of forest dynamics at stand scales due to
the long-term legacy of past disturbances.
Among niche-based explanations of tree species
coexistence, differences in multiple tree life history
traits, expressed throughout all stages of a tree’s life,
are thought to contribute to maintenance of canopy
diversity (Nakashizuka 2001). In temperate forests, this
has been supported by both modeling work (Loehle
2000) and empirical studies (Veblen 1986, Lusk and
Smith 1998, Taylor et al. 2006, Gutierrez et al. 2008).
Several of these studies suggest that interspecific
differences in juvenile performance in gap–understory
gradients and adult life span contribute to tree
coexistence, whereby low recruitment rates of juvenile
trees into the canopy may be compensated by lower
mortality rates of adults. Although our analysis of radial
growth patterns only provides indirect evidence of
juvenile tree shade tolerance, the results suggest that
beech’s lower tolerance to shaded understory conditions
compared to that of fir may be balanced by the longer
life span of beech. In addition to life span, growing tall
may also convey certain advantages, namely, permanent
access to light, while casting shade on shorter neighbors
(Loehle 2000). Adult fir trees, which often reach heights
.50 m in mature stands, form an emergent canopy over
neighboring beech trees, thus adding another dimension
to the possible life history strategies that contribute to
maintaining tree diversity in beech–fir forests (Szwagrzyk
et al. 2012).
Although various life history differences may contrib-
ute to equilibrium coexistence of beech–fir forests at
stand scales, the relatively frequent occurrence of
intermediate-severity disturbances found in this study
suggests that canopy composition is probably non-
equilibrial. Certainly, the persistence of less shade-
tolerant species in our study system, such as maple,
ash, and elm, is linked to disturbances that create
canopy openings larger than those formed from
‘‘background’’ mortality of single canopy trees. Previous
work in Peru�cica indicated that maple, for example,
requires gaps of at least 400 m2 to recruit to the canopy
(Nagel et al. 2010). Once these less shade-tolerant
species reach the canopy, their relatively long life span
and seed dispersal ability may ensure their persistence in
the stand, particularly if intermediate-severity distur-
bance recurs at intervals suggested by our data. The
presence of these less shade-tolerant species in beech-
dominated forests of Europe has typically been associ-
ated with more mesic site conditions (Ellenberg 1988);
our results emphasize that disturbance, not only site
conditions, may be an important mechanism by which
these species coexist. Finally, our results and those of
other recent studies highlight that gap-phase models of
community dynamics, whereby tree diversity is main-
tained via tree replacement under a regime of low
intensity, relatively continuous canopy mortality (e.g.,
Forcier 1975, Barden 1980, Runkle 1982, Woods 1984),
may not be sufficient to explain the dynamics of tree
communities in temperate forests without considering
the influence of periodic, intermediate-severity distur-
bances (Woods 2000, Beaudet et al. 2007, Firm et al.
2009).
Management implications.—Throughout mountain-
ous regions of Europe, where beech and mixed beech–
fir forests are common, forests are managed with various
even- and uneven-aged silvicultural systems, including
single-tree selection, group selection, and shelterwood
systems (Madsen and Hahn 2008, Brunet et al. 2010,
Boncina 2011b). Used individually, none of these
systems would be sufficient to emulate the spatial and
temporal variability of canopy mortality documented in
this study. Rather, harvesting that employs several
silvicultural systems at appropriate spatial and temporal
scales within a forest landscape would be required to
reasonably mimic natural mortality patterns and to help
foster structural and compositional diversity at land-
scape scales (Seymour et al. 2002, Hanson and Lorimer
2007). For example, in mixed beech–fir forests managed
with small-scale selection silviculture, an approach that
is often advocated in such forests types in Europe
(Boncina 2011b), periodic, spatially variable harvests
that remove ;20–50% of the upper canopy layer in
patches several hectares in size could mimic the
intermediate-severity disturbances documented in this
study. Such structures may be best achieved with an
irregular shelterwood system (Raymond et al. 2009). In
other temperate forest regions where intermediate-
severity wind disturbances periodically interrupt forest
dynamics driven by small-scale, ‘‘background’’ mortal-
ity, similar approaches have been recommended to
curtail diversity declines that may result from only using
small-scale selection systems (Woods 2004, Hanson and
Lorimer 2007). Clearly, care needs to be taken not to
over-disturb managed landscapes, given that some
amount of natural canopy disturbance is inevitable,
such as damage from catastrophic windstorms (Seymour
et al. 2002).
Although long-term use of single-tree selection is
likely to result in a loss of less shade-tolerant species
(Schutz 1999), periodic higher intensity cutting, through
group selection, shelterwood cuts, or other more
irregular multi-cohort systems (Raymond et al. 2009),
should help to maintain tree diversity. However, our
results caution that advance regeneration of shade-
tolerant species, in our case beech, may inhibit
recruitment of less shade-tolerant species following
heavier harvests. This is particularly relevant for the
application of group selection and shelterwood systems,
which tend to favor shade-tolerant advance regeneration
by establishment and expansion of group openings in
June 2014 675EUROPEAN OLD-GROWTH FOREST DISTURBANCE

areas where beech regeneration is already established or,
in the case of shelterwood systems, by waiting for a mast
year to promote the establishment of dense beech
regeneration prior to the final overstory harvest. To
facilitate recruitment of less shade-tolerant species, it
may be necessary to use periodic, heavier cuts in closed-
canopy areas devoid of shade-tolerant advance regener-
ation, but where adequate seed sources of less shade-
tolerant species are present.
In addition to silvicultural prescriptions that account
for the size and frequency of natural disturbances,
incorporating other biological legacies resulting from
disturbances, particularly coarse woody debris, is central
for developing ecologically based management systems.
In forests dominated by beech, for example, the
importance of large old trees and sufficient amounts of
dead wood for maintaining myriad species dependent on
these structures is well documented (Brunet et al. 2010,
Gossner et al. 2013). Periodic, intermediate-severity
disturbances, similar to those documented in this work,
result in pulses of dead wood input in old-growth forests
(D’Amato et al. 2008), and are likely to play an
important role in the population dynamics of species
dependent on these structural elements (Jonsson et al.
2011). For example, a conservative estimate of the
amount of dead wood added to the Tunjemir-2 plot
following the single windthrow event in the 1950s totals
about 240 m3/ha (i.e., 50 trees with mean dbh of 64 cm),
more than an order of magnitude greater than the
average amount found in managed forests of Europe
(Forest Europe, UNECE and FAO 2011). Creating
pulses of dead wood input in managed forests, however,
is challenging, but could involve retaining a portion of
felled trees during a harvest, creation of dead wood
islands within a matrix of managed forest (Jakoby et al.
2010), or limiting the amount of salvage logging
following natural disturbances in managed areas.
Indeed, many of the findings documented in this work
would be challenging to incorporate in forests where
wood production is a primary function. Large amounts
of dead wood retention, long rotation periods (e.g., the
canopy residence time, calculated from the inverse of the
overall median disturbance rate, is 200 years in our
study area), and landscape-scale structural heterogeneity
may be unrealistic goals throughout much of the
European temperate region, where forest areas are often
small and heavily utilized. Despite these challenges, the
idea that managing forested landscapes with ecologically
based forestry (often termed ‘‘close-to-nature’’ forestry
in Europe) supplants the need for a segregated approach
to forest management, whereby forest land is divided
into areas focused on intensive wood production and
protected areas focused on ecological functions, is
gaining increased attention in Europe (Boncina 2011a).
However, we caution that close-to-nature forestry
should not be treated as a panacea for forest manage-
ment; in addition to forest land managed with ecolog-
ically based approaches, there is still a need to protect
and expand unmanaged forest reserves in the temperate
region of Europe to create large areas of old-growth
forest where landscape-scale structural and composi-
tional heterogeneity can develop. Recent work, in fact,
suggests that triad forest management (Seymour and
Hunter 1999), consisting of an intensive management
zone focused on wood production, an integrated
management zone that balances ecological and econom-
ic functions (e.g., similar to close-to-nature forestry in
Europe), and a reserve zone for restoration or protection
of existing old-growth forest, may be a better approach
to biodiversity protection than a less intensive integra-
tion approach spread out over a forested landscape
(Montigny and MacLean 2006, Tittler et al. 2012). The
more balanced approach of the triad model should also
provide more flexibility to foster adaptation to climate
change (Millar et al. 2007).
ACKNOWLEDGMENTS
Funding was provided by the Slovenian Research Agency(bilateral, program financing) and the Pahernik Foundation. M.Svoboda received support from the project ESF and MSMTCZ.1.07/2.3.00/30.0040. We thank the staff at Sutjeska NationalPark for allowing us to carry out research in the Peru�cicareserve. For help in the field and laboratory, we thank MatejTajnikar, Urban Oroz, Janez Miklavcic, Katarina Flajsman,Gal Fidej, Urska Bradesko, Urska Klepec, Tihomir Rugani, andJan Nagel. We thank Jeri Peck, Robert S. Seymour, and twoanonymous reviewers for providing valuable comments on aprevious version of the manuscript. Finally, we thank KerryWoods for many stimulating discussions on disturbance andcommunity dynamics in temperate forests.
LITERATURE CITED
Angelstam, P. K. 1998. Maintaining and restoring biodiversityin European boreal forests by developing natural disturbanceregimes. Journal of Vegetation Science 9:593–602.
Baker, P. J., and S. Bunyavejchewin. 2006. Suppression, releaseand canopy recruitment in five tree species from a seasonaltropical forest in western Thailand. Journal of TropicalEcology 22:521–529.
Barden, L. S. 1980. Tree replacement in cove hardwood forestof the southern Appalachians. Oikos 35:16–19.
Bauhus, J., K. Puettmann, and C. Messier. 2009. Silviculturefor old-growth attributes. Forest Ecology and Management258:525–537.
Beaudet, M., J. Brisson, D. Gravel, and C. Messier. 2007. Effectof a major canopy disturbance on the coexistence of Acersaccharum and Fagus grandifolia in the understorey of an old-growth forest. Journal of Ecology 95:458–467.
Bengtsson, J., S. G. Nilsson, A. Franc, and P. Menozzi. 2000.Biodiversity, disturbances, ecosystem function and manage-ment of European forests. Forest Ecology and Management132:39–50.
Bergeron, Y. 2000. Species and stand dynamics in the mixedwoods of Quebec’s southern boreal forest. Ecology 81:1500–1516.
Black, B. A., and M. D. Abrams. 2003. Use of boundary-linegrowth patterns as a basis for dendroecological releasecriteria. Ecological Applications 13:1733–1749.
Boncina, A. 2011a. Conceptual approaches to integrate natureconservation into forest management: a Central Europeanperspective. International Forestry Review 13:13–22.
Boncina, A. 2011b. History, current status and future prospectsof uneven-aged forest management in the Dinaric region: anoverview. Forestry 84:467–478.
THOMAS A. NAGEL ET AL.676 Ecological ApplicationsVol. 24, No. 4

Braun-Blanquet, J. 1964. Pflanzensoziologie, Grundzuge derVegetationskunde. Third edition. Springer Verlag, Vienna,Austria.
Brunet, J., O. Fritz, and G. Richnau. 2010. Biodiversity inEuropean beech forests: a review with recommendations forsustainable forest management. Ecological Bulletins 53:77–94.
Canham, C. D. 1990. Suppression and release during canopyrecruitment in Fagus grandifolia. Bulletin of the TorreyBotanical Club 117:1–7.
Christensen, M., et al. 2005. Dead wood in European beech(Fagus sylvatica) forest reserves. Forest Ecology and Man-agement 210:267–282.
Clark, J. S., et al. 2010. High-dimensional coexistence based onindividual variation: a synthesis of evidence. EcologicalMonographs 80:569–608.
D’Amato, A. W., and D. A. Orwig. 2008. Stand and landscape-level disturbance dynamics in old-growth forests in westernMassachusetts. Ecological Monographs 78:507–522.
D’Amato, A. W., D. A. Orwig, and D. R. Foster. 2008. Theinfluence of successional processes and disturbance on thestructure of Tsuga canadensis forests. Ecological Applications18:1182–1199.
Diaci, J., editor. 1999. Virgin forests and forest reserves inCentral and East European countries: history, present statusand future development. Proceedings of the COST E4Management Committee and Working Groups Meeting,25–28 April 1998, Ljubljana, Slovenia.
Drobyshev, I., R. Overgaard, I. Saygin, M. Niklasson, T.Hickler, M. Karlsson, and M. T. Sykes. 2010. Mastingbehavior and dendrochronology of European beech (Fagussylvatica L.) in southern Sweden. Forest Ecology andManagement 259:2160–2171.
Duncan, R. P. 1989. An evaluation of errors in tree ageestimates based on increment cores in kahikatea (Dacrycar-pus dacrydioides). New Zealand Natural Sciences 16:31–37.
Ellenberg, H. 1988. Vegetation ecology of central Europe.Fourth edition. Cambridge University Press, Cambridge,UK.
Ellenberg, H. 1992. Indicator values of the vascular plants(except Rubus). Pages 9–166 in H. Ellenberg, H. E. Weber, R.Dull, V. Wirth, W. Werner, and D. Paulißen, editors.Indicator values of plants in Central Europe. Erich GoltzeKG, Gottingen, Germany.
Firm, D., T. A. Nagel, and J. Diaci. 2009. Disturbance historyand dynamics of an old-growth mixed species mountainforest in the Slovenian Alps. Forest Ecology and Manage-ment 257:1893–1901.
Forcier, L. K. 1975. Reproductive strategies and the co-occurrence of climax tree species. Science 189:808–810.
Forest Europe, UNECE, and FAO. 2011. State of Europe’sForests 2011. Status and Trends in Sustainable ForestManagement in Europe. http://www.unece.org/forests/fr/outputs/soef2011.html
Fraver, S., and A. S. White. 2005. Disturbance dynamics of old-growth Picea rubens forests of northern Maine. Journal ofVegetation Science 16:597–610.
Fraver, S., A. S. White, and R. S. Seymour. 2009. Naturaldisturbance in an old-growth landscape of northern Maine,USA. Journal of Ecology 97:289–298.
Frelich, L. E., and L. J. Graumlich. 1994. Age-class distributionand spatial patterns in an old-growth hemlock–hardwoodforest. Canadian Journal of Forest Research 24:1939–1947.
Frelich, L. E., and C. G. Lorimer. 1991. Natural disturbanceregimes in hemlock–hardwood forests of the Upper GreatLakes Region. Ecological Monographs 61:145–164.
Frohlich, J. 1954. Urwaldpraxis. 40 jahrige Erfahrungen undLehren. Neumann Verlag, Berlin, Germany.
Fukarek, P., and V. Stefanovic. 1958. Prasuma Peru�cica i njenavegetacija. Sarajevo, Bosnia-Herzegovina.
Gossner, M. M., T. Lachat, J. Brunet, G. Isacsson, C. Bouget,H. Brustel, R. Brandl, W. W. Weisser, and J. Muller. 2013.Current near-to-nature forest management effects on func-tional trait composition of saproxylic beetles in beech forests.Conservation Biology 27:605–614.
Gravel, D., C. D. Canham, M. Beaudet, and C. Messier. 2010.Shade tolerance, canopy gaps and mechanisms of coexistenceof forest trees. Oikos 119:475–484.
Greenberg, C. H., and W. H. McNab. 1998. Forest disturbancein hurricane-related downbursts in the Appalachian moun-tains of North Carolina. Forest Ecology and Management104:179–191.
Gutierrez, A. G., J. C. Aravena, N. V. Carrasco-Farias, D. A.Christie, M. Fuentes, and J. J. Armesto. 2008. Gap-phasedynamics and coexistence of a long-lived pioneer and shade-tolerant tree species in the canopy of an old-growth coastaltemperate rain forest of Chiloe Island, Chile. Journal ofBiogeography 35:1674–1687.
Hanson, J. J., and C. G. Lorimer. 2007. Forest structure andlight regimes following moderate wind storms: implicationsfor multi-cohort management. Ecological Applications17:1325–1340.
Holmes, R. L. 1983. Computer-assisted quality control in tree-ring dating and measurements. Tree-Ring Bulletin 44:69–75.
Jakoby, O., C. Rademacher, and V. Grimm. 2010. Modellingdead wood islands in European beech forests: how much andhow reliably would they provide dead wood? EuropeanJournal of Forest Research 129:659–668.
Johnson, E. A., K. Miyanishi, and H. Kleb. 1994. The hazardsof interpretation of static age structures as shown by standreconstructions in a Pinus contorta–Picea engelmaanii forest.Journal of Ecology 82:923–931.
Jonsson, M. T., S. Fraver, and B. G. Jonsson. 2011. Spatio-temporal variation of coarse woody debris input in woodlandkey habitats in Central Sweden. Silva Fennica 45:957–967.
Keeton, W. S. 2006. Managing for late-successional/old-growthcharacteristics in northern hardwood–conifer forests. ForestEcology and Management 235:129–142.
Kobe, R. K., S. W. Pacala, J. A. Silander, and C. D. Canham.1995. Juvenile tree survivorship as a component of shadetolerance. Ecological Applications 5:517–532.
Korpel, S. 1982. Degree of equilibrium and dynamical changesof the forest on example of natural forests of Slowakia. ActaFacutatis Forestalis, Zvolen, Czechoslovakia 24:9–30.
Korpel, S. 1995. Die Urwalder der Westkarpaten. GustavFischer Verlag, Stuttgart, Germany.
Kucbel, S., P. Jaloviar, M. Saniga, J. Vencurik, and V. Klimas.2010. Canopy gaps in an old-growth fir–beech forest remnantof Western Carpathians. European Journal of ForestResearch 129:249–259.
Kunstler, G., T. Curt, M. Bouchaud, and J. Lepart. 2005.Growth, mortality, and morphological response of Europeanbeech and downy oak along a light gradient in sub-Mediterranean forest. Canadian Journal of Forest Research35:1657–1668.
Leibundgut, H. 1959. Uber Zweck und Methodik der Strukturund Zuwachsanalyse von Urwaldern. Schweizerische Zeit-schrift Forstwes 110:111–124.
Levanic, T. 2007. ATRICS—A new system for image acquisi-tion in dendrochronology. Tree-Ring Research 63:117–122.
Loehle, C. 2000. Strategy space and the disturbance spectrum: alife-history model for tree species coexistence. AmericanNaturalist 156:14–33.
Lorimer, C. G., and L. E. Frelich. 1989. A methodology forestimating canopy disturbance frequency and intensity indense temperate forests. Canadian Journal of ForestResearch 19:651–663.
Lusk, C. H., and B. Smith. 1998. Life history differences andtree species coexistence in an old-growth New Zealand rainforest. Ecology 79:795–806.
June 2014 677EUROPEAN OLD-GROWTH FOREST DISTURBANCE

Madsen, P., and K. Hahn. 2008. Natural regeneration in abeech-dominated forest managed by close-to-nature princi-ples—a gap cutting based experiment. Canadian Journal ofForest Research 38:1716–1729.
Mayer, H., K. Zukrigl, W. Schrempf, and G. Schlager. 1989.Urwaldreste, Naturwaldreservate und schutzenswerte Natur-walder in Osterreich. Second edition. Institut fur Waldbau,Universitat fur Bodenkultur, Wien, Austria.
Millar, C. I., N. L. Stephenson, and S. L. Stephens. 2007.Climate change and forests of the future: Managing in theface of uncertainty. Ecological Applications 17:2145–2151.
Montigny, M. K., and D. A. MacLean. 2006. Triad forestmanagement: Scenario analysis of forest zoning effects ontimber and non-timber values in New Brunswick, Canada.Forestry Chronicle 82:496–511.
Motta, R., R. Berretti, D. Castagneri, V. Dukic, M. Garbarino,Z. Govedar, E. Lingua, Z. Maunaga, and F. Meloni. 2011.Toward a definition of the range of variability of centralEuropean mixed Fagus–Abies–Picea forests: the nearlysteady-state forest of Lom (Bosnia and Herzegovina).Canadian Journal of Forest Research 41:1871–1884.
Muller, J., and R. Butler. 2010. A review of habitat thresholdsfor dead wood: a baseline for management recommendationsin European forests. European Journal of Forest Research129:981–992.
Nagel, T. A., and J. Diaci. 2006. Intermediate wind disturbancein an old-growth beech–fir forest in southeastern Slovenia.Canadian Journal of Forest Research 36:629–638.
Nagel, T. A., T. Levanic, and J. Diaci. 2007. A dendroeco-logical reconstruction of disturbance in an old-growth Fagus–Abies forest in Slovenia. Annals of Forest Science 64:891–897.
Nagel, T. A., and M. Svoboda. 2008. Gap disturbance regime inan old-growth Fagus–Abies forest in the Dinaric Mountains,Bosnia-Herzegovina. Canadian Journal of Forest Research38:2728–2737.
Nagel, T. A., M. Svoboda, and J. Diaci. 2006. Regenerationpatterns after intermediate wind disturbance in an old-growth Fagus–Abies forest in southeastern Slovenia. ForestEcology and Management 226:268–278.
Nagel, T. A., M. Svoboda, T. Rugani, and J. Diaci. 2010. Gapregeneration and replacement patterns in an old-growthFagus–Abies forest of Bosnia-Herzegovina. Plant Ecology208:307–318.
Nakashizuka, T. 2001. Species coexistence in temperate, mixeddeciduous forests. Trends in Ecology and Evolution 16:205–210.
Noss, R. F. 2001. Beyond Kyoto: Forest management in a timeof rapid climate change. Conservation Biology 15:578–590.
Nowacki, G. J., and M. D. Abrams. 1997. Radial-growthaveraging criteria for reconstructing disturbance historiesfrom presettlement-origin oaks. Ecological Monographs67:225–249.
Oliver, C. D., and B. C. Larson. 1996. Forest stand dynamics.Updated edition. John Wiley, New York, New York, USA.
Orwig, D. A., and M. D. Abrams. 1994. Contrasting radialgrowth and canopy recruitment patterns in Liriodendrontulipifera and Nyssa sylvatica: gap-obligate versus gap-facultative tree species. Canadian Journal of Forest Research24:2141–2149.
Pacala, S. W., C. D. Canham, J. Saponara, J. A. Silander, R. K.Kobe, and E. Ribbens. 1996. Forest models defined by fieldmeasurements: Estimation, error analysis and dynamics.Ecological Monographs 66:1–43.
Palairet, M. 1997. The Balkan economies c. 1800–1914:evolution without development. Cambridge University Press,Cambridge, UK.
Petritan, A. M., B. Von Luepke, and I. C. Petritan. 2007.Effects of shade on growth and mortality of maple (Acerpseudoplatanus), ash (Fraxinus excelsior) and beech (Fagussylvatica) saplings. Forestry 80:397–412.
Piovesan, G., A. Di Filippo, A. Alessandrini, F. Biondi, and B.Schirone. 2005. Structure, dynamics and dendroecology of anold-growth Fagus forest in the Apennines. Journal ofVegetation Science 16:13–28.
Poljansek, S., D. Ballian, T. A. Nagel, and T. Levanic. 2012. A435-year-long European black pine (Pinus nigra) chronologyfor the central-western Balkan region. Tree-Ring Research68:31–44.
R Development Core Team. 2011. R: A Language andEnvironment for Statistical Computing. R Foundation forStatistical Computing, Vienna, Austria. http://www.r-project.org
Raymond, P., S. Bedard, V. Roy, C. Larouche, and S.Tremblay. 2009. The irregular shelterwood system: review,classification, and potential application to forests affected bypartial disturbances. Journal of Forestry 107:405–413.
Runkle, J. R. 1982. Patterns of disturbance in some old-growthmesic forests of eastern North America. Ecology 63:1533–1546.
Schroter, D., et al. 2005. Ecosystem service supply andvulnerability to global change in Europe. Science 310:1333–1337.
Schutz, J. P. 1999. Close-to-nature silviculture: is this conceptcompatible with species diversity? Forestry 72:359–366.
Seymour, R. S., and M. L. Hunter, Jr. 1999. Principles ofecological forestry. Pages 22–61 in: M. L. Hunter, Jr., editor.Managing biodiversity in forest ecosystems. CambridgeUniversity Press, Cambridge, UK.
Seymour, R. S., A. S. White, and P. G. deMaynadier. 2002.Natural disturbance regimes in northeastern North Amer-ica—evaluating silvicultural systems using natural scales andfrequencies. Forest Ecology and Management 155:357–367.
Siegel, S., and N. J. Castellan. 1988. Nonparametric statisticsfor the behavioral sciences. MacGraw Hill, New York, NewYork, USA.
Spiecker, H. 2003. Silvicultural management in maintainingbiodiversity and resistance of forests in Europe (temperatezone). Journal of Environmental Management 67:55–65.
Splechtna, B. E., G. Gratzer, and B. A. Black. 2005.Disturbance history of a European old-growth mixed-speciesforest—A spatial dendro-ecological analysis. Journal ofVegetation Science 16:511–522.
Stueve, K. M., C. H. Perry, M. D. Nelson, S. P. Healey, A. D.Hill, G. G. Moisen, W. B. Cohen, D. D. Gormanson, and C.Huang. 2011. Ecological importance of intermediate wind-storms rivals large, infrequent disturbances in the northernGreat Lakes. Ecosphere 2:1–21.
Svoboda, M., P. Janda, T. A. Nagel, S. Fraver, J. Rejzek, andR. Bace. 2012. Disturbance history of an old-growth sub-alpine Picea abies stand in the Bohemian Forest, CzechRepublic. Journal of Vegetation Science 23:86–97.
Szwagrzyk, J., J. Szewczyk, and Z. Maciejewski. 2012. Shade-tolerant tree species from temperate forests differ in theircompetitive abilities: A case study from Roztocze, south-eastern Poland. Forest Ecology and Management 282:28–35.
Taylor, A. H., S. W. Jang, L. J. Zhao, C. P. Liang, C. J. Miao,and J. Y. Huang. 2006. Regeneration patterns and treespecies coexistence in old-growth Abies–Picea forests insouthwestern China. Forest Ecology and Management223:303–317.
Tittler, R., C. Messier, and A. Fall. 2012. Concentratinganthropogenic disturbance to balance ecological and eco-nomic values: applications to forest management. EcologicalApplications 22:1268–1277.
Trotsiuk, V., M. L. Hobi, and B. Commarmot. 2012. Agestructure and disturbance dynamics of the relic virgin beechforest Uholka (Ukrainian Carpathians). Forest Ecology andManagement 265:181–190.
Veblen, T. T. 1986. Treefalls and the coexistence of conifers insubalpine forests of the central Rockies. Ecology 67:644–649.
THOMAS A. NAGEL ET AL.678 Ecological ApplicationsVol. 24, No. 4

Webb, S. L., and S. E. Scanga. 2001. Windstorm disturbance
without patch dynamics: twelve years of change in a
Minnesota forest. Ecology 82:893–897.
Wolf, A., P. F. Moller, R. H. W. Bradshaw, and J. Bigler. 2004.
Storm damage and long-term mortality in a semi-natural,
temperate deciduous forest. Forest Ecology and Manage-
ment 188:197–210.
Woods, K. D. 1984. Patterns of tree replacement: canopy
effects on understory pattern in hemlock–northern hardwood
forests. Vegetatio 56:87–107.
Woods, K. D. 2000. Dynamics in late-successional hemlock–
hardwood forests over three decades. Ecology 81:110–126.
Woods, K. D. 2004. Intermediate disturbance in a late-successional hemlock–northern hardwood forest. Journal ofEcology 92:464–476.
Worrall, J. J., T. D. Lee, and T. C. Harrington. 2005. Forestdynamics and agents that initiate and expand canopy gaps inPicea–Abies forests of Crawford Notch, New Hampshire,USA. Journal of Ecology 93:178–190.
Yamaguchi, D. K. 1991. A simple method for cross-datingincrement cores from living trees. Canadian Journal of ForestResearch 21:414–416.
Zukrigl, K., G. Eckhart, and J. Nather. 1963. Standortskund-liche und waldbauliche Untersuchungen in Urwaldresten derniederosterreichischen Kalkalpern. Osterreichischer Agrar-verlag, Vienna, Austria.
SUPPLEMENTAL MATERIAL
Appendix A
Forest structure and composition of the Tunjemir and Zanoglina areas in the Peru�cica forest reserve (Ecological ArchivesA024-039-A1).
Appendix B
Plots of percentage growth change values with respect to prior growth for annual growth increments of Abies alba and Fagussylvatica, including the boundary line functions for both species (Ecological Archives A024-039-A2).
Appendix C
Animations showing the spatial and temporal patterns of canopy accession (including both gap origin and release) in the fourdisturbance history plots (Ecological Archives A024-039-A3).
Appendix D
Number of gapmakers in different decay classes for Fagus sylvatica and Abies alba and density of seedlings and saplings in thethree windthrow and neighboring control areas (Ecological Archives A024-039-A4).
Appendix E
Maps of canopy gaps for the four disturbance history plots (Ecological Archives A024-039-A5).
June 2014 679EUROPEAN OLD-GROWTH FOREST DISTURBANCE




























