Embed Size (px)
Citation preview

246 BIOCHIMICA ET B1OPHYSICA ACTA
BBA 97467
DNA R E P A I R A F T E R ULTRAVIOLET IRRADIATION IN SYNCHRONOUS
PLASMODIA OF P H Y S A R U M P O L Y C E P H A L U M
J. JUSTIN MCCORMICK*, CAROL MARKS AND H. 13. RUSCH McArdle Laboratory for Cancer Research, University of Wisconsin Medical Center, Madison, Wisc. ( U.S.A.)
(Received June 3oth, 1972)
SUMMARY
Plasmodia of Physarum polycephahtm were irradiated with 14 500 ergs/mm 2 of ultraviolet irradiation during early G2 period and the subsequent repair synthesis of DNA was studied. I t was found that the G2 period, which is normally 5 h in length, is prolonged to 18 h by such a dose of irradiation and that repair synthesis occurs almost entirely during the last 9 h of this period. Following the extended G2 period, the nuclei undergo mitosis in a normal manner and during the DNA synthesis period which follows mitosis all the nuclear DNA is replicated semiconservatively as shown by density label centrifugation studies. During the extended G2 period, normal replication of the nucleolar satellite DNA and trace amounts of nuclear DNA occurs and the total amount synthesized is equal to what would have been synthesized in a G2 period of normal length. Caffeine shows a strong inhibitory effect on DNA repair synthesis without interfering with DNA replication or the length of the intermitotic period.
INTRODUCTION
Extreme radioresistance in eucaryotic cells has been reported in free-living amoeba-like cells 1'~. Radioresistance has also been demonstrated in plasmodia of Physarum polycephalum which can withstand 25 000 rad of X-irradiation 3, 14 500 ergs/mm 2 of ultraviolet irradiation 4,s and 1.5 mg/ml of the radiomimetic agent, ni- trogen mustard 6. These results suggest that this organism may have extraordinary repair capabilities. The data on X-irradiation are most telling since factors such as plasmodial thickness (which might potentiate ultraviolet irradiation) or plasmodial permeability (which might potentiate nitrogen mustard damage) can be presumed to have little effect on X-rays.
P. polycephalum plasmodia are syncytia which exhibit synchronous nuclear division and synchronous synthesis of nuclear DNA in the 3-h period immediately following mitosis. Replicate plasmodia set up from the same pool of microplasmodia
" Present address: Michigan Cancer Foundation, 4811 John R. Street, Detroit, Mich. 482or. U.S.A.
Abbreviations: BrdUrd, 5-bromo-2'-deoxyuridine; FdUrd, 5-fIuoro-z'-deoxyuridine; SSC, o.15 M NaC1 containing o.o15 M sodium citrate (pH 7.2).
Biochirn. Biophys. Acta, 287 (1972) 246-255

DNA REPAIR AFTER ULTRAVIOLET IRRADIATION 247
are synchronous with each other. If ultraviolet irradiation is performed early in the G2 period when normal DNA synthesis is essentially complete, it is possible to deter- mine the extent of repair by measuring the amount of label incorporated without interference from normal synthesis.
In this study, the repair capabilities of P. polycephalum were exanfined follow- ing ultraviolet irradiation at 14 500 ergs/nlm 2, the dose used by Devi et al. 4 and by Sachsenmaier et al. 5 in their studies on the effect of ultraviolet irradiation upon mi- tosis in plasmodia of P. polycephalum. Repair synthesis was deternfined by the method of Painter and Cleaver 7 which involved the incorporation of I3HlBrdUrd into DNA and subsequent separation of DNA on alkaline cesium chloride gradients to detect non-semiconservative replication. I t has been reported that caffeine blocks DNA repair in bacteria 8 but not in mammalian cells in culture 9. Since caffeine has been shown to increase the sensitivity of P. polycephalum amoebae to ultraviolet irra- diation 1°, its effect upon DNA repair in this organism was also examined.
MATERIALS AND METHODS
Physarurn polycephalum, subline M3cV, was carried in shake flask cultures on semi-defined medium, as previously described n. Prior to each experiment, 3 1111 of a full grown shake flask culture was transferred to 20 ml of fresh medium to which 0.2 #Ci/ml of E14C]thymidine had been added. Cultures were shaken for approx. 24 h. Petri dish cultures were prepared on a Millipore HA membrane with I ml of water - microplasmodial suspension (2 : I, v/v). 1. 5 h were allowed for fusion of the micro- plasmodia before the addition of medium to the Petri dishes. They were then incu- bated at 26 °C. Mitosis I occurred approx. 4 to 5 h after the addition of the medium. The intermitotic period between mitosis I and mitosis I I was 8 h in control cultures. Onset of mitosis was determined by phase microscopy, as previously described 6.
After incubation in [3H]BrdUrd medium, cultures were harvested, quick-frozen in liquid nitrogen and stored at --20 °C for not more than 2 days. The I~HlBrdUrd medium used was a modification of that previously reported 12. Regular growth medi- um was prepared which contained 4 ° / ,g /ml BrdUrd, 2.5#Ci/ml [3H]BrdUrd, 5/zg/ml FdUrd, and IOO #g/ml of uridine. FdUrd was included because it blocks thymidine synthesis and thus increases the extent of incorporation of the thymidine analog, BrdUrd (J. J. McCormick, unpublished). Uridine was added to prevent a decrease in the rate of RNA synthesis, which is seemingly caused by a breakdown of FdUrd to 5-fluorouraci113.
Ultraviolet irradiation was performed using two I5-W Champion GI5T8 germicidal lamps positioned over the plasmodia in Petri dishes with covers removed so that an incident dose of 14 500 ergs/mm2was delivered in 2 min 29 s. Measurement of ultraviolet irradiation was performed with a J225 metering unit from Ultra-violet Products, San Gabriel, Calif.
At the time of irradiation, control cultures were transferred from the incubator along with the cultures to be irradiated and were held in an area immediately adjacent to, but protected from, the ultraviolet lamp during the period of irradiation (sham irradiation). After ultraviolet or sham irradiation, all cultures were protected against white light and incubated in the dark to prevent photoreactivation.
Biochim. Biophys. Acta, 287 (1972) 246-255

248 j . J . MCCORMICK e[ al.
L3H] BrdUrd (5 mCi/mole) and [14C]thymidine (5 Ci/mole) were purchased from New England Nuclear (Boston, Mass.). BrdUrd, uridine, Pronase and ribo- nuclease were purchased from Calbiochem (Los Angeles, Calif.). CsC1 was obtained from Kerr-McGee Chemicals (Chicago, Ill.). FdUrd was a gift from C. Heidelberger of this laboratory.
DNA was prepared from nuclei isolated according to the method of Mohberg and Rusch 14. Nuclei were incubated for 30 min in o.15 M NaC1 containing o.o15 M sodium citrate, pH 7.2, (SSC) containing IOO #g/ml of pancreatic ribonuclease and 5o units per ml of TI ribonuclease which had been preincubated at 80 °C for IO rain to denature any contaminating DNAase. Nuclei were lysed by the addition o! sodium lauryl sarkosine to a final concentration of o.I 0.2 o,;, fl)llowed by incubation at 60 °C for IO min. The solution was cooled to room temperature and Pronase was added to a final concentration of approx. I mg/ml. Prior to use, Pronase was predigested for a minimum of 3 h at 37 °C to inactivate any DNAase in the preparation. A drop of chloroform was added to each crude DNA solution to prevent bacterial growth and the preparation was transferred to a dialysis sack. Dialysis against 1 1 of SSC was carried out overnight at room temperature with at least 3 changes of solution. The DNA preparation was then transferred to 3-nil conical glass centrifuge tubes and centrifuged at 27 3o0 × g (13 ooo rev./min) in a HB- 4 swinging bucket rotor of a Sorvall refrigerated centrifuge to remove much of the contaminating nuclear poly- saccharide and thereby reduce the viscosity of the aqueous phase containing the DNA.
To prepare neutral CsC1 gradients, solid CsC1 was dissolved in a total of 4.5 ml SSC solution containing the DNA sample. Alkaline gradients were prepared as follows. Each centrifuge tube contained a total of 1.5 ml of SSC which included the DNA sample plus 3 ml of o.15 M Na2HPO 4, 0.2 M NaOH (pH 12.4) and solid CsC1. The refractive index of all solutions was determined with a Bausch and Lomb refracto- meter and was adjusted, if necessary, to a refractive index of 1.3997 1.4 °o0 for neutral gradients and 1.4o62-1.4o65 for alkaline gradients. Tubes were overlaid with paraffin oil and centrifuged for at least 60 h at 25 °C in a Model L or Model L2-65B Spinco ultracentrifuge equipped with a No. 4 ° rotor (Beckman Instruments, Palo Alto, Calif.). After centrifugation, tubes were punctured at the bottom with a 20- gauge needle and I2-drop fractions collected. CsC1 gradients were not strictly linear since traces of sodium lauryl sarkosine, which were not removed by dialysis, were found in the light region of the gradient and caused a reduction in the size of the drop. All comparisons were made within an individual experiment where dialysis conditions were identical to minimize this effect.
For determination of radioactivity, each fraction from the neutral gradients was added to scintillation vials containing o.2 ml of distilled water and IO ml Instagel (Packard Laboratories, Downers Grove, Ill.) and counted for IO min on a Packard Liquid Scintillation Spectrometer with absolute activity analyzer programmed to correct for double label samples. Samples from alkaline gradients were acidified by the addition of o.I ml of I.O M HCI to each scintillation vial to obtain a clear counting solution. The fractions from the alkaline CsC1 gradient centrifugation of each sample were diluted with 0. 5 inl of distilled water and o.I ml of the dilution was counted as above. After plotting the data from these fractions it was necessary to reband, in a second alkaline CsC1 gradient, the region of normal density DNA to show the extent
Biochim. Biophys. dcta, 287 (1972) 246-255

D N A REPAIR AFTER ULTRAVIOLET IRRADIATION 249
of [aH]BrdUrd incorporation. This was done by pooling the fractions from the region to be rebanded, lyophilizing them, and preparing them for centrifugation as describ- ed above. Recovery of the counts applied to the gradients was 85-95 %.
EXPERIMENTAL DESIGN
To examine repair synthesis of DNA at intervals after ultraviolet irradiation, the protocol shown in Fig. I. was used. A number of synchronous plasmodia were ultraviolet-irradiated or sham-irradiated 5 h after mitosis I. It was known from preli- minary experimentation that little repair synthesis occurred during the first 9 h after ultraviolet irradiation. Therefore, only one culture was harvested at this point (Fig. I, Culture C). Two cultures were harvested during the second 9-h period, Fig. I, Cultures D and E; Culture D was harvested 14 h after irradiation and Culture E during early prophase of the first post-irradiation mitosis, mitosis II. Taken together, these 3 cultures comprise the total DNA repair which took place during G2. Similar information about repair can be obtained from Culture F. It is possible to divide the time of repair into discrete periods in G2 since preliminary experiments showed that incubation of plasmodia in [3H]BrdUrd medium during G2 only minimally delays the next mitosis. The amount of DNA replication in the synthetic period immediately following mitosis II was determined from an analysis of Culture G.
Control cultures were incubated with [3HJBrdUrd medium at appropriate times and harvested either at prophase of mitosis II, Fig. I, Culture A; or 4 h after mitosis occurred, Fig. I, Culture B.
UV IRRADIATION 14 500 ERG*~'mm 2
EX~ER,.E.TAL 4 t ' I CULTURES " ' CULTURE C . =
CULTURE D
CULTURE E
CULTURE F
SHAM IRRADIATION CONTROL I ~' I
CULTURES M | M 1"r
CULTURE A
m m ~ m CULTURE B
o E T I U E I I
CULTURE G
16
I H O U R S
m
2 4
I
Fig. i . Pro tocol of expe r imen t . Cul tures C, D, E, F and G were i r r ad i a t ed a t the t ime ind ica t ed b y the arrow. Cul tures A and B were s h a m - i r r a d i a t e d a t t h a t same t ime, as ind ica ted . Cul tures were i n c u b a t e d w i t h [SH]lBrdUrd m e d i u m for the t ime per iod ind ica t ed by the solid line; dashed l ine ind ica t e s i n c u b a t i o n on regu la r g rowth medium. [aH]BrdUrd m e d i u m is def ined unde r Mate r i a l s and Methods. A second set of cu l tu res was t r e a t ed in the same m a n n e r bu t was in- c u b a t e d in m e d i u m c o n t a i n i n g 70o/~g/ml of caffeine from I h before u l t r a v i o l e t or sham irra- d i a t ion un t i l t he t ime of ha rves t . Such cu l tu res are ind ica t ed in the t e x t by the add i t i on of the subsc r ip t "c" to the des igna ted le t t e r ; e.g. Ac. Mitosis I is i nd ica t ed by M[; mi tos i s I I as MII .
Biochim. Biophys. Acta, 287 (1972) 246-255

250 J . J . MCCORMICK et al.
To determine the effect of caffeine upon repair synthesis, a second set of six cul- tures were treated in the same manner except that they were incubated in medium containing 700/~g/ml caffeine from I h before the time of ultraviolet irradiation or sham irradiation until time of harvest. At this dose caffeine had no effect upon mitosis but some cultures rounded up into small mounds after extended incubation in this medium.
For both of these kinds of experiments, cultures were prelabeled with [14C~dThd so tha t the bulk of the nuclear DNA could be located without the need for 26o-nm absorbance readings. Any new synthesis of DNA or repair synthesis during a parti- eula, period would be indicated by incorporation of EZH~BrdUrd. Neutral gradients were run in order to determine the extent that E3H]BrdUrd had been incorporated into bihelical DNA molecules. Normal replication synthesis of DNA in the presence of [3H}BrdUrd will result in an easily detectable shift in the density of such DNA. Alkaline CsC1 gradients were used to detect the non-semiconservative incorporation of EaHlBrdUrd into DNA. At a pH of 12.4, the DNA becomes single stranded. Strands of DNA synthesized semiconservatively in the presence of E~H~BrdUrd will incorpo- rate long stretches of more dense material and band near the bot tom of the tube. The light parent DNA strands labeled with ElaCldThd will band near the top of the tube. [3H~BrdUrd found incorporated into the light [14CldThd region of the gradient indicates non-semiconservative synthesis which is at tr ibuted to repair of the DNA. In practice, it is necessary to reband the E14CldThd region in a second alkaline CsC1 gradient in order to determine whether I3HlBrdUrd incorporation in this region is real or artifactual.
RESULTS
Before investigating the effect of caffeine upon repair synthesis, it was neces- sary to determine the maximum dose of this agent which would be tolerated by plasmodia of P. polycephalum without interfering with mitosis or the steps leading to it. Preliminary studies showed that 250 #g/ml of this agent, which is the dose level used to sensitize P. polycephalum amoebae to ultraviolet irradiation, had no effect upon repair-type synthesis of DNA in plasmodia and when the dose was in- creased to 85 °/~g/ml, plasmodia were killed. However, plasmodia underwent mitosis at the normal 8-h intervals in the presence of caffeine at 700 #g/ml and this concen- tration was used in all experiments reported here. Plasmodia were transferred to caffeine medium i h before ultraviolet or sham irradiation.
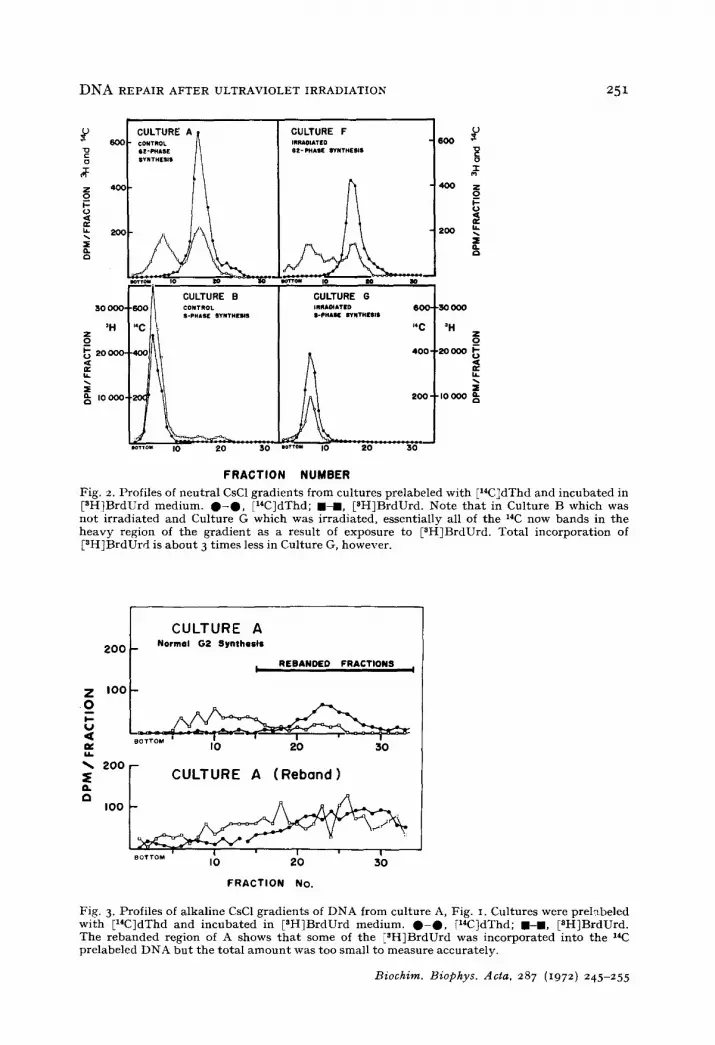
Neutral CsC1 gradients were run on the DNA extracted from Cultures B and G of Fig. I to determine whether plasmodia were capable of semiconservative repli- cation of all their nuclear DNA during the first post-irradiation synthetic period. A comparison of profiles B and G of Fig. 2 with profiles A and F of Fig. 2 shows that all of the E14ClDNA is shifted to a heavier density and that the incorporation of E3H~BrdUrd is greatly enhanced. The fact that no [14CIDNA of light density remains in B and G indicates that all of the DNA is replicated. Cultures treated caffeine were processed in the same manner as the ultraviolet-irradiated and control cultures and gave identical results. Profile G shows that in the S period following ultraviolet irradiation, only 1/3 as much pH}BrdUrd is incorporated. This may be because pycnotic nuclei which are present in such cultures failed to incorporate BrdUrd.
Biochim. Biophys. Acta, 287 (i972) 246-255
DNA REPAIR AFTER ULTRAVIOLET IRRADIATION 251
'~ 6OO
4oo
~o
30 000-
=H z
0 ~ 20 000.
b. :S
I0 000,
CULTURE A CONTROL NZ-~ASE SYNTHESIS
CULTURE B 60C
I* C
40(:
ao~
n
IOTTO~
CONTROL S-PHASE NYNTHE~S
I0 20 30
CULTURE F INIIAOIATED e t - PHASE I~tNTHE$1$
CULTURE G IRRAOIATED 600' tt-PHAK gYNTHNNI$
J4 C
V 6OO
2 O 0
.30000
=H Z _o
400 ' ,20000 1
200 ' " I0 000
lOTTO. I0 20 30
F R A C T I O N N U M B E R
Fig. 2. Profiles of neutral CsC1 gradients from cultures prelabeled wi th El*C~dThd and incubated in [3H]BrdUrd medium. 0 - 0 , IziC]dThd; I - m , [NH]BrdUrd. Note t ha t in Culture B which was not i rradiated and Culture G which was irradiated, essentially all of the 14C now bands in the heavy region of the gradient as a result of exposure to [NH]BrdUrd. Total incorporat ion of [NH~BrdUrd is abou t 3 t imes less in Culture G, however.
CULTURE A 2 0 0 - Normal G2 Synthesis
b R E B A N D E D F R A C T I O N S I
Z I O 0 - o m
m~ I O 20 30 kh
~. 2oor- CULTURE A (Rebond)
BOTTOM II ' II IO 20
FRACTION NO.
3 0
Fig. 3. Profiles of alkaline CsC1 gradients of DNA from culture A, Fig. I. Cultures were prel~beled wi th [z4C]dThd and incubated in [SH]BrdUrd medium. 0 - 0 , [x4C]dThd; I - D , [SH]BrdUrd. The rebanded region of A shows t h a t some of the ENH]BrdUrd was incorporated into the 1'C prelabeled DNA bu t the total a m o u n t was too small to measure accurately.
Biochim. Biophys. Acta, 287 (1972) 245-255

252 J . J . MCCORMICK et al.
Under ordinary circumstances high-density nucleolar satellite DNA and a trace of normal density nuclear DNA are synthesized during G2. I t was of interest to see whether the extension of G2 period by ultraviolet irradiation would affect such syn- thesis. Neutral CsC1 gradient profiles of Culture A and Culture F (Fig. ~) show almost identical patterns. This indicates that, in spite of the extremely extended G2 period after ultraviolet irradiation, normal DNA synthesis during G~ does occur. The heavy [aH]BrdUrd peak which appears to the left in these two gradients is the nucleolar- satellite DNA and trace amounts of nuclear DNA replicated semiconservatively in the presence of BrdUrd and thus shifted to a heavier density than normal. This peak is about the same density as the BrdUrd peak in Fig. z, B and O. This is to be expected since BrdUrd substitution in the nucleolar satellite DNA (51 °/" o G + C) will be less than in the nuclear DNA (41%) G + C) causing the density to be almost identical as has been noted 15. There is no 14C peak under the heavy [3H~BrdUrd peak in Fig. 2,
o,. of the total DNA 15. The [aH1BrdUrd A and F since the satellite represents only 2 ..o radioactivity which appears under the [14C!DNA peak probably represents a combi- nation of normal repair-type synthesis, mechanical entrapment of heavier [~H]Brd- Urd in the 14C peak, and synthesis of trace amounts of [aH!BrdUrd DNA ioined end- to-end to normal density nuclear DNA (indicated by a slight shift toward the denser region of the gradient).
To determine whether low levels of repair-type synthesis of DNA occur normally during the G2 period in P. polycephalum, the following experiment was carried out. Alkaline CsC1 gradients were run on DNA which was extracted from cultures desig- nated A and Ac in the legend of Fig. I. Fig. 3 shows the profile of a CsCI gradient from Culture A as well as a reband of the 14C region from this gradient. These data suggest small amounts of aHlBrdUrd are incorporated into the ~4C-labeled DNA, but the radioactivity counts are too near background to provide any degree of confidence
I CULTURE C CULTURE C (Rebond) 1 400
/ \~ 200
• I 0 2b 3 0 . T ~ 0 2 0 ~$0 /a- . . . . 400 "~ I O,TORE Co f COUTURE Co ,R,°o°°, , I
I~RA~ATION + CAFFEI~ I IRRADIATION + CAFFEINe IK x~ ',
,oo I- I . ~ • "'~,: 4 2oo
. , I BOTXO~ eOTTO IO ZO 30 IO 20 :30
F R A C T I O N N U M B E R
Fig. 4- Profi les of alkal ine CsC1 g rad ien t s of D N A f rom cul tures des igned C and Cc in Fig. i . Cu l tu res were prelabeled wi th [14CldThd and i ncuba t ed in [3HlBrdUrd med ium. O - O , [14CMThd; I - B , [3H]BrdUrd. Note change of scale in Cc. Incorpora t ion of isotopes in cu l tures t rea ted wi th caffeine is approx, equal to t h a t of the controls .
Biochim. Biophys. dcta, 287 (t972) z46 z55

DNA REPAIR AFTER ULTRAVIOLET IRRADIATION 253
in this conclusion. Corresponding cultures exposed to caffeine show similar profiles. Thus, there is no clear evidence for repair during the normal G2 period.
Fig. 4 shows profiles of alkaline CsC1 gradients and rebands of the ~*C region of cultures incubated with [~H~BrdUrd for the first 9 h after irradiation. These are designated C and Ce in the legend of Fig. I. There is no appreciable difference in [~H!BrdUrd incorporation (repair synthesis) between those cultures which were treated with caffeine and those which were incubated in "caffeine-free" medium.
Fig. 5 shows the profiles of DNA extracted from Cultures D and Dc which were incubated with ~H~BrdUrd medium during the next 5-h period. Repair-type syn-
400
200 Z o I.-- 0 <[
I¢. CULTURE Dc • %. 9-14 HOURS AFTER IRRADIATION + CAFFEINE
"% I REBANDED FRACTION! I Q- I00
, o
CULTURE D CULTURE D (R~nd)~" ~ 9-14 HOURS AFTER 9-14 HOURS AFTER
IRRADIATION IRRADIATION ~ ~ y ~ | RIE~NDEO FR~TIOI~I |
~TTO. I0 20 80 ~orrou I0 20 30
~rT0. I0 20 30
F R A C T I O N N U M B E R
4oo z 0 V- O
400 b.
t" G.
~'00
Fig. 5. Profiles of alkaline CsC1 gradients of DNA from cultures designated D and Dc in Fig. I. Cultures were prelabeled wi th [l*C~dThd and incubated in [3HlBrdUrd medium. 0 - 0 , [14C]dThd; m-m, [3H]BrdUrd. Note change of scale for Cultures D and De. (Caffeine inhibits repair synthesis of DNA approx. 80 % in this period.)
CULTURE E
I REIIANDIED FRACTIONS I Z~ O
, . . -
u.
n ~ ioo~-CULTURE EC I~'WL"~P F~¢TIONS t t too
mo
CU~TU2L A ,~R'b~"2~
p ~ , CAFFE~N!
FRACTION NUMBER
Fig. 6. Profiles of a lkal ine CsC1 g rad i en t s of D N A from cul tures des igna ted E and Ec in Fig. i . Cu l tu res were pre labeled wi th [14C~dThd and i ncuba t ed in IRH]BrdUrd m e d i u m . 0 - 0 , II*C~dThd; B - I , [3H~BrdUrd. Note scale change in Ec. (Caffeine inh ib i t s repair syn thes i s of D N A more t h a n 80 % du r ing th i s period.)
Biochim. Biophys. Acta, 287 (1972) 246-255

254 j . j . MCCORMICK e~ al.
thesis, as shown by the alkaline rebands, is much greater than in the earlier 9-h period and is inhibited approximately 4-fold by treatment with caffeine.
Fig. 6 shows the alkaline CsC1 gradient profiles of DNA extracted from Cultures E and Ec which had been incubated with F3H]BrdUrd medium during the last 4 h of the extended G2 period. These cultures were harvested at prophase of mitosis II. Repair synthesis of DNA remains at high levels though less than that seen in Fig. 5. Treatment with caffeine inhibits repair synthesis to about the same extent as seen in Fig. 5.
DISCUSSION
These results show that repair-type synthesis of DNA, similar to that reported for other types of celW, also occurs in the plasmodia of P. polycephalum after high doses of ultraviolet irradiation. The major portion of this repair-type DNA synthesis occurs between 9 and 18 h after irradiation during the G2 period. In addition, this repair-type synthesis is largely blocked by caffeine. Other cell systems undergo maxi- mal repair-type synthesis of DNA in the period immediately following ultraviolet irradiation 7 while P. polycephalum shows a time lag before maximal repair. Whether this lag relates to induction or activation of repair enzymes or to other factors is now under study. I t is also of interest to examine whether repair synthesis continues in the first post-irradiation DNA synthetic period when normal DNA synthesis is block- ed.
Devi et al. 4 exposed P. polycephalum plasmodia to 14 500 ergs/mm ~ of ultra- violet irradiation and observed up to 17. 3 °/o degenerating or pycnotic nuclei both before and after the first post-irradiation mitosis. In contrast, only 0.76 o,L of the nuclei in the unirradiated controls were pycnotic. They suggest that the large number of degenerating nuclei in the irradiated cells may be the result of unequal exposure of the nuclei to ultraviolet irradiation caused by the thickness of the plasmodia (approx. o.o5-o.lo mm). Since we have shown that all the DNA replicates during the first post-irradiation synthetic period and since, by definition, pycnotic nuclei do not undergo normal DNA synthesis, it seems likely the DNA of pycnotic nuclei is degra- ded. Sachsenmaier et al. 5 have shown by microspectrophotometry that after irradi- ation, surviving nuclei contain the same amount of DNA as in controls.
If there is a sizable pool of free nucleotides available from degenerating nuclei, it is possible that the E14C]thymidine used in these experiments is re-utilized. A recent study by Cleaver and Trosko 16 has shown that in Chinese hamster cells reutilization, if it occurs, is less than 0.05 %. Such a study, however, may not reflect what happens in a multinuclear "cell" such as the plasmodium of P. polycephalum. The amount of reutilization which occurs in P. polycephalum has not been determined, but the large amount of BrdUrd present in the medium in the present experiments should minimize the reincorporation of any [~4C]thymidine made available by degradation of the DNA. The possible re-utilization of ~4C label for DNA synthesis is an important consideration in our experiments since repair is measured as the ratio of ~4C to aH counts in the normal density DNA peak of alkaline CsC1 gradients. In any event, re-utilization of ~4C label would cause an underestimation of the extent of repair.
The blockage of repair synthesis by caffeine that has been noted here with
Biochim. Biophys. Acta, 287 (1972) 246-255

DNA REPAIR AFTER ULTRAVIOLET IRRADIATION 255
P. po lycephalum is s i m i l a r to t h a t w h i c h is r e p o r t e d in b a c t e r i a l s y s t e m s s a n d u n l i k e
t h a t f o u n d in m a m m a l i a n cel ls w h e r e ca f f e ine does n o t i n t e r f e r e w i t h r e p a i r 9 b u t
r a t h e r w i t h a s t e p in s e m i c o n s e r v a t i v e D N A r e p l i c a t i o n l L
ACKNOWLEDGMENTS
T h i s w o r k was s u p p o r t e d b y P u b l i c H e a l t h Se rv i ce G r a n t CA-o7175 a n d CA-
5002 f r o m t h e N a t i o n a l C a n c e r I n s t i t u t e , a n d b y a g r a n t f r o m t h e A l e x a n d e r a n d
M a r g a r e t S t e w a r t T r u s t F u n d .
W e t h a n k L a V i l a W i n n i e a n d J u d y B l o m q u i s t for t e c h n i c a l a s s i s t ance .
REFERENCES
I R. A. Deering, M. S. Smith, B. K. Thompson and A. C. Adolf, Radiat. Res., 43 (197 o) 7 TM
2 J. B. Litt le and R. S. Chang, Radiat. Res., 35 (1968) 132. 3 R. A. McGrath, R. W. Williams and R. B. Setlow, Int. J. Radiat. Biol., 8 (1964) 373. 4 V. R. Devi, E. Guttes and S. Guttes, Exp. Cell Res., 5 ° (1968) 589 . 5 W. Sachsenmaier, K. H. Donges and H. Rupff, Z. Naturforsch., 25b (197 o) 866. 6 J. J. McCormick and R. M. Nardone, Exp. Cell Res., 6o (197 o) 247. 7 R. B. Painter and J. E. Cleaver, Radiat. Res., 37 (1969) 451. 8 A. S. Sideropoulos and D. M. Shankel, J. Bacteriol., 96 (1968) 198. 9 J. E. Cleaver, Radiat. Res., 37 (1969) 334.
IO F. B. Haugli and W. F. Dove, Mol. Gen. Genet., in the press. i I J. W. Daniel and H. H. Baldwin, in D. Prescott, Methods in Cell Physiology, Vol. I, Academic
Press, New York, 1964, p. 9. 12 R. Braun, C. Mit termayer and H. P. Rusch, Proc. Natl. Acad. Sci. U.S., 53 (1965) 912. 13 W. Sachsenmaier and H. P. Rusch, Exp. Cell Res., 36 (1964) 124. 14 J. Mohberg and H. P. Rusch, Exp. Cell Res., 66 (1971) 305. 15 A. Zellweger, U. Ryser and R. Braun, J. Mol. Biol., 64 (1972) 681. 16 J. E. Cleaver and J. E. Trosko, Int. J. Radiat. Biol., 15 (1969) 411. 17 J. E. Cleaver and G. H. Thomas, Biochem. Biophys. Res. Commun., 36 (1969) 203.
Biochim. Biophys. Acta, 287 (1972) 246-255





























