Embed Size (px)
Citation preview

Ecology, 89(12), 2008, pp. 3275–3281� 2008 by the Ecological Society of America
DUCKING AS A MEANS OF RESISTANCE TO HERBIVORYIN TALL GOLDENROD, SOLIDAGO ALTISSIMA
MICHAEL J. WISE1
AND WARREN G. ABRAHAMSON
Department of Biology, Bucknell University, Lewisburg, Pennsylvania 17837 USA
Abstract. Many populations of goldenrod show a peculiar, genetically controlled stemdimorphism. In Solidago altissima, for instance, while most stems are erect, a sizable minority(the ‘‘candy-cane’’ stems) nod at the apex during growth. We used data from three studies totest the hypothesis that this candy-cane growth form confers resistance to herbivory. In acontrolled growth trial, we showed that nodding is a temporary phenomenon that coincideswith the oviposition period of at least two common apex-attacking herbivores: the tephritidgaller Eurosta solidaginis and the gall midge Rhopalomyia solidaginis. In a large field survey,stems of candy-cane genets were only half as likely to be ovipunctured by E. solidaginis. In acommon-garden study, candy-cane stems were less likely to be ovipunctured by E. solidaginis,and they were galled only half as often by R. solidaginis as erect stems. These results suggestthat the candy-cane stems of goldenrod possess a resistance strategy that allows them toessentially duck and hide from certain herbivores.
Key words: candy-cane stems; ducking; Eurosta solidaginis; galls; goldenrod; resistance to herbivory;Rhopalomyia solidaginis; Solidago altissima; stem morphology.
INTRODUCTION
While chemical toxins and mechanical deterrents
dominate the arsenal of known or suspected herbivore
defenses in plants, architectural features of plants may
also serve important roles in plant resistance (Lawton
1983, Kareiva and Sahakian 1990, Alonso and Herrera
1996, Espırito-Santo et al. 2007). Investigators have
identified a variety of architectural characteristics that
can affect resistance to herbivory, including number of
buds, branches, or flowers (Oghiakhe et al. 1993, Araujo
et al. 2006), size of internodes (Larson and Whitham
1997), spacing of leaves (Marquis et al. 2002), rolling of
young leaves (Grubb and Jackson 2007), pod angle
(Oghiakhe et al. 1992), growth habit (e.g., erect,
prostrate, or spreading [Oghiakhe et al. 1993, Rudgers
and Whitney 2006]), and architectural complexity in
general (Kaitaniemi et al. 2004). Perhaps the most basic
feature of plant architecture, however, is size. Plant
height, in particular, can have a major influence on a
plant’s susceptibility to herbivory.
Although being tall may provide trees and shrubs
protection against browsing (Palo et al. 1993), some
vertebrate (Merton et al. 1976) and many invertebrate
herbivores, particularly flying insects, tend to prefer
taller plants (Bach 1981, Karban and Courtney 1987,
Marquis 1992, Haysom and Coulson 1998). Plant height
may be an indication of host vigor, or taller plants may
just be more apparent and thus more likely to be found
by herbivores (Lawton 1983). As such, short stature can
reduce a plant’s susceptibility to herbivory, and, to the
extent that height is genetically controlled, shortness
might be favored by natural selection to reduce
herbivore damage. However, the evolution of shortness
as a resistance strategy is likely to be strongly
constrained by the disadvantages short plants face in
competing with taller neighbors for access to light or
pollinators. In this paper, we report on a trait that can
potentially enable plants to take advantage of short
stature to increase resistance while minimizing the
associated costs.
The stems of the common herbaceous perennial
Solidago altissima L. (‘‘tall goldenrod,’’ Asteraceae)
display a genetically controlled dimorphism. The stems
emerge from rhizomes in spring, and after a couple of
weeks each stem develops an apical-leaf bud that lasts
until the apical meristem begins to produce flower buds
in mid-summer. In most stems (the ‘‘erect’’ morpholo-
gy), the apical-leaf bud points straight up throughout
the growing season until the plants flower. In a
substantial minority of plant genets (i.e., genetic
individuals), however, the stem begins to bend near the
apex as the apical-leaf bud forms until the apex points
straight down. Although the stem appears wilted, the
apex is turgid. The stem remains recurved for weeks, and
the resemblance of the stem shape to a familiar candy
earned this nodding morphology the nickname ‘‘candy-
cane’’ stem (Fig. 1). The candy-cane stems straighten
back up just prior to flowering.
We hypothesized that the candy-cane morphology
acts as a resistance mechanism to reduce attack by
herbivores that oviposit or feed on the apical-leaf bud.
Specifically, we asked whether candy-cane stems in-
Manuscript received 8 February 2008; revised 16 June 2008;accepted 27 June 2008; final version received 23 July 2008.Corresponding Editor: R. J. Marquis.
1 E-mail: [email protected]
3275
Rep
orts

curred less damage than erect stems from two common
stem-gall-inducing dipterans: the fruit fly Eurosta solid-
aginis (Fitch) (Tephritidae) and the gall midge Rhopa-
lomyia solidaginis Lw. (Cecidomyiidae). This study
involved three components: (1) in greenhouse trials, we
compared the growth phenology of stems of erect and
candy-cane genets; (2) in a large field survey, we
measured ovipositions by E. solidaginis and subsequent
gall formation on stems of 110 erect and 11 candy-cane
genets; and (3) in a common garden study, we measured
ovipositions by E. solidaginis and galling rates of both E.
solidaginis and R. solidaginis on stems of 21 erect and
five candy-cane genets.
METHODS
Study species
Solidago altissima is a perennial herbaceous weed
abundant in roadsides, old fields, and waste places
throughout much of the eastern United States and
southern Canada. The herbivore fauna of S. altissima is
one of the best studied for any plant species (Maddox
and Root 1987, Root 1996, Abrahamson and Weis
1997), and 138 species of insects have been identified as
feeding on S. altissima in the Finger Lakes region of
New York state alone (Root and Cappuccino 1992).
Many of these insects attack the apices of goldenrod in
spring and early summer, during the window in which
the candy-cane stem morphology is expressed. Two of
the most common of these species are the gall-inducing
flies Eurosta solidaginis and Rhopalomyia solidaginis.
Previous studies have shown that both of these species
can negatively affect the growth and reproduction of S.
altissima (Hartnett and Abrahamson 1979, Stinner and
Abrahamson 1979, Abrahamson and Weis 1997, Wise et
al. 2006a).
Eurosta solidaginis is a univoltine specialist on S.
altissima in the study area (central Pennsylvania). Adults
emerge from galls in mid-to-late May and mate on
goldenrod ramets (Horner and Abrahamson 1992).
Females oviposit on apical-leaf buds, often after walking
up and down the stem several times and making test
punctures with their ovipositors in the bud (Uhler 1951,
Walton et al. 1990). These ovipunctures are visible as
one or more linear series of holes in the apical-leaf bud,
and later as an identical pattern of holes repeated on
successive leaves that were formerly overlapping in the
apical bud. Eggs hatch in about a week and spherical
stem galls become visible within three weeks (Uhler
1951, Craig et al. 2000). Each gall contains one larva,
but a single ramet occasionally forms up to three galls
(Hess et al. 1996, Wise et al. 2008). Full-grown larvae
diapause over winter inside galls and pupate the
following spring.
Overwintering first-instar larvae of the gall midge
Rhopalomyia solidaginis form relatively inconspicuous,
generally single-chambered galls on the apical meristems
of newly emerged goldenrod shoots in early spring: late
April in this study area (McEvoy 1988; N. Dorchin,
personal communication). Adults emerge in late May,
and gravid females lay eggs on S. altissima leaf buds.
The larvae initiate large, generally multi-chambered,
leafy ‘‘rosette’’ galls that can have major effects on
goldenrod morphology (Hartnett and Abrahamson
1979, Raman and Abrahamson 1995, Wise et al.
2006a). Second generation adults emerge from these
galls in late summer, and females lay eggs in soil or
seemingly haphazardly on plants (McEvoy 1988). Upon
hatching, larvae crawl underground to diapause over
winter, probably within goldenrod rhizomes (N. Dorch-
in, personal communication).
Stem-growth phenology
To compare stem-growth phenologies, we randomly
chose four candy-cane and four erect genets from a set
of 26 S. altissima genets originally collected as rhizomes
from a field in 2003 and grown in controlled conditions
each year (see methods in Common garden-experiment).
Rhizomes of these genets were removed from refriger-
ation on 26 June 2006 and cut into 2-mL segments,
which were measured using water displacement in a
graduated cylinder. The segments were planted into flats
in ProMix BX (Premier Horticulture Ltd., Dorval,
Quebec, Canada) in a greenhouse at Bucknell Univer-
sity. Shoots emerged in about one week, and eight
healthy ramets of each genet were transplanted into
16.5-cm plastic azalea pots on 24 July. Measurements
FIG. 1. Nodding, candy-cane stem of Solidago altissima.
MICHAEL J. WISE AND WARREN G. ABRAHAMSON3276 Ecology, Vol. 89, No. 12R
epor
ts
were taken weekly until flowering (through 16 Septem-
ber) on (1) height from ground to highest point, (2)
height from ground to apex tip, and (3) total stem
length. For an erect stem, these three measurements are
equivalent.
Field survey
As part of a study to examine the stability of E.
solidaginis galling patterns in genets over time (McCrea
and Abrahamson 1987), we marked all the ramets of 121
spatially distinct genets (110 erect-stem and 11 candy-
cane-stem genets) in a roadside population along an 8.2-
km section of Route 147 in Northumberland County,
Pennsylvania, USA and measured galling rates from
1982 to 1984. In early June of 1984, immediately after
the period of E. solidaginis oviposition activity, we
counted all ramets from the 121 genets and loosely tied
small wire bands around ramets with E. solidaginis
ovipunctures. At the end of the growing season, we
revisited the plants to observe whether galls were formed
on each ramet.
Data analysis involved one-tailed t tests of the hypoth-
esis that candy-cane genets were more resistant than erect
genets. The response variables were the proportion of
rametsper genet (1)withE. solidaginisovipunctures and (2)
with mature E. solidaginis galls. Both proportions were
square-root transformed prior to analysis. A third, two-
tailed t-test was performed to test whether the proportion
of punctured ramets that ended up with mature galls
differed between stem types. These proportions were
arcsine-square-root transformed prior to analysis.
Common-garden experiment
In early April of 2003, rhizomes were collected from
26 S. altissima genets, which were separated by at least
30 m, from a 3-ha field population in Union County,
Pennsylvania (4085705400 N, 7685701800 W). (Rhizomes of
21 of these genets produced erect stems, and five
produced candy-cane stems.) The rhizomes were
cleaned, cut into 5-cm segments and planted into flats
in ProMix BX. After shoot emergence, the plants were
transplanted into 16.5-cm diameter plastic azalea pots.
In mid-June, 15 ramets of each genet (390 pots total)
were transplanted into a 300-m2 garden plot that had
been treated with herbicide, cleared, and plowed. The
garden plot was divided into a grid with planting
positions 90 cm apart within a row and 90 cm between
rows. Each sequence of 26 positions in the plot was
considered one statistical block, and one ramet from
each genet was placed in a randomized position in each
of the 15 blocks.
In mid-March of 2004, we collected over 1200 E.
solidaginis galls from the goldenrod field from which the
rhizomes were collected the previous spring. To inocu-
late the garden plot with E. solidaginis, we placed three
galls at each of the 390 planting positions. Rhopalomyia
solidaginis colonized the garden plot on its own. In mid-
to-late May, we tagged up to five of the nearest newly
emerged ramets to each planting position that had
clearly arisen from the original individual. Usually more
than five ramets were available, but there were always at
least two ramets per position (mean number tagged ¼4.83, SD¼0.53 ramets). Between 25 May and 3 June, we
examined each tagged ramet for E. solidaginis ovipunc-
tures). In late July and again in December, we revisited
each ramet and recorded the presence of E. solidaginis
and R. solidaginis galls.
Data analysis involved ANOVA and one-tailed t tests
of the hypothesis that candy-cane stems were more
resistant than erect stems. The original planting position
was considered the independent statistical unit, and the
response variables were the proportion of ramets at each
planting position with (1) E. solidaginis ovipunctures, (2)
mature E. solidaginis galls, and (3) R. solidaginis galls.
Four observers searched plants for ovipunctures in
different sections of the plot. To account for potential
spatial differences in ovipuncture frequencies in the field,
or differences in observers’ ability to spot ovipunctures,
observer identity was included in the ANOVA for
ovipunctures.
RESULTS
Stem-growth phenology
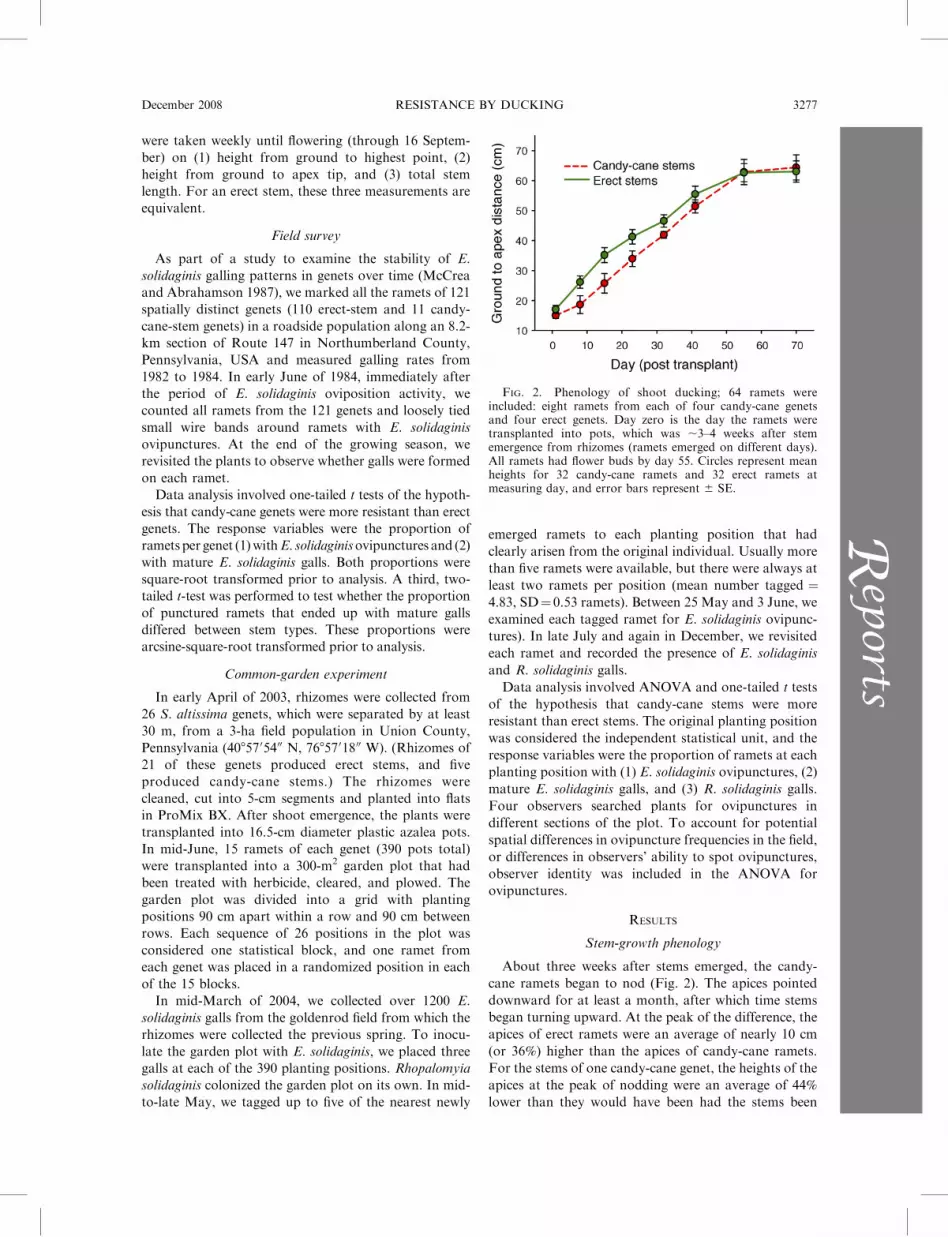
About three weeks after stems emerged, the candy-
cane ramets began to nod (Fig. 2). The apices pointed
downward for at least a month, after which time stems
began turning upward. At the peak of the difference, the
apices of erect ramets were an average of nearly 10 cm
(or 36%) higher than the apices of candy-cane ramets.
For the stems of one candy-cane genet, the heights of the
apices at the peak of nodding were an average of 44%
lower than they would have been had the stems been
FIG. 2. Phenology of shoot ducking; 64 ramets wereincluded: eight ramets from each of four candy-cane genetsand four erect genets. Day zero is the day the ramets weretransplanted into pots, which was ;3–4 weeks after stememergence from rhizomes (ramets emerged on different days).All ramets had flower buds by day 55. Circles represent meanheights for 32 candy-cane ramets and 32 erect ramets atmeasuring day, and error bars represent 6 SE.
December 2008 3277RESISTANCE BY DUCKINGR
eports

straight (based on total main-stem length). By the time
the plants were flowering (around 12 weeks post
emergence), the candy-cane stems had straightened
completely, and the heights of the stems of the two
types no longer differed.
Field survey
In the 1984 survey of a roadside population of 121 S.
altissima genets, the stems of the 11 candy-cane genets
were only about half as likely to have ovipunctures as
the stems of the 110 erect genets (one-tailed t¼ 2.43, P¼0.008; Fig. 3A). By the end of the season, 9% of the
candy-cane stems and 13% of the erect stems had mature
E. solidaginis galls (one-tailed t ¼ 1.08, P ¼ 0.14; Fig.
3B). This disparity in galling rates between stem types
was not as large as the disparity in ovipuncture rates
because candy-cane stems were slightly (but not
statistically significantly) more likely to form galls once
ovipunctured than were erect stems (43% vs. 33%; two-
tailed t¼�1.24, P ¼ 0.22; Fig. 3C).
Common-garden experiment
The stems of the five candy-cane genets in the
common-garden experiment were only 82% as likely to
be ovipunctured by E. solidaginis as erect stems (one-
tailed P¼ 0.0006, Fig. 4, Table 1). As in the field survey
of 1984, this significant difference in resistance to
ovipuncturing did not translate into a significant
difference in the presence of mature E. solidaginis ball
galls on the two stem types. Ball galls were found on an
average of 19.0% 6 3.3% of the stems of the candy-cane
genets and 21.4% 6 1.6% of the stems of erect genets
(one-tailed P ¼ 0.32). Again, there was no significant
difference in the probability of the two stem types
forming galls once oviposited: 39.1% 6 7.5% for candy-
cane stems vs. 32.8% 6 3.8% for erect stems (two-tailed
P ¼ 0.65).
There was a larger discrepancy between the stem types
in resistance to the gall midge R. solidaginis. By the end
of the summer, stems of candy-cane genets were only
half as likely to have R. solidaginis rosette galls as stems
of erect genets (P , 0.0001; Fig. 5, Table 1).
DISCUSSION
Stems of candy-cane genets of Solidago altissima were
less likely to be ovipunctured by the ball-galling fly
Eurosta solidaginis, and they were less likely to be form
rosette galls induced by the midge Rhopalomyia solid-
FIG. 3. Herbivory by Eurosta solidaginis on S. altissima in the field survey. Columns represent mean percentages (6 SE) oframets for 110 erect genets and 11 candy-cane genets: (A) ovipuncture rate (P¼ 0.008); (B) mature gall presence (P¼ 0.14); (C)gall-formation rate of ovipunctured ramets (P¼ 0.22).
FIG. 4. Herbivory by Eurosta solidaginis on S. altissima in the common-garden experiment. The experiment included fivecandy-cane and 21 erect genets, replicated in 15 spatial blocks. Columns represent least-squares means proportions (6 SE) oframets for each stem type from an ANOVA on stem type, block, and data collector: (A) ovipuncture rate (P¼ 0.0006); (B) maturegall presence (P¼ 0.32); (C) gall-formation rate of ovipunctured ramets (P¼ 0.65).
MICHAEL J. WISE AND WARREN G. ABRAHAMSON3278 Ecology, Vol. 89, No. 12R
epor
ts

aginis, than were stems of erect genets. Because both of
these gallers can severely impact the growth and
reproduction of S. altissima (Hartnett and Abrahamson
1979, Stinner and Abrahamson 1979, Abrahamson and
Weis 1997, Wise et al. 2006a), candy-cane stems should
enjoy a clear fitness advantage due to their greater
resistance.
With such an important resistance benefit, a big
question is why the candy-cane stems are always in the
minority, at least in the geographic area of this study.
One possibility is simply that the ducking trait has
evolved only recently, and selection will drive it to be
more common in the future. Alternatively, balancing
selection may act on stem morphology, and ducking
may entail costs to the plants.
The simplest evidence for a cost to the candy-cane
strategy would be lower seed production than erect
stems in the absence of gall flies. The results of a
separate study on the plants involved in the common-
garden study show no evidence for such a cost (M. J.
Wise and W. G. Abrahamson, unpublished data).
Specifically, 18 ramets of each of the five candy-cane
and 21 erect-stemmed genets were exposed to a range of
levels of spittlebug herbivory (but no galling) (Wise et al.
2006b). The genets differed in seed production (F25, 434¼24.49, P , 0.0001), but the difference had nothing to do
with stem morphology. The mean for the candy-cane
genets was 22,068 6 3503 seeds per ramet, while the
mean for the erect genets was 19 450 6 1709 seed per
ramet (t ¼ 0.67, P ¼ 0.51).
Though differences in seed production between stem
morphs were not observed in the spittlebug study, the
candy-cane morphology may still entail costs that would
not have been evident in that study. For instance, it is
possible that there are tradeoffs among different
components of defense against the gall inducing flies.
In the field and garden studies, there was a slight
indication that ovipunctured candy-cane ramets might
be more likely to form galls than ovipunctured erect
ramets. Such a pattern would indicate that the
antixenosis (i.e., avoidance) resistance advantage of
candy-cane stems could cost the plants in terms of
lower antibiosis resistance. In addition, because resis-
tance and tolerance are sometimes negatively correlated
(Leimu and Koricheva 2006), it is possible that candy-
cane stems have lower tolerance of galling than erect
stems. It is also possible that candy-cane stems are less
resistant than erect stems to species of herbivores otherthan these gall inducing flies. While we have no data
indicating that such tradeoffs exist, we plan to look
more closely into these possibilities in the future.
Another potential cost of the candy-cane morphologymay be apparent in more light-limiting environments.
FIG. 5. Rhopalomyia galling rates on S. altissima in thecommon-garden experiment. The experiment included fivecandy-cane and 21 erect genets, replicated in 15 spatial blocks.Columns represent least-squares means (6 SE) of the percent-age of ramets ovipunctured for each stem type from anANOVA on stem type and block (P , 0.0001.)
TABLE 1. ANOVA results for resistance of candy-cane- and erect-stem morphs of Solidago to gallflies (Eurosta solidaginis) and gall midges (Rhopalomyia solidaginis).
Resistance measure Source of variation df MS F P
Percentage of ramets with stem morph 1 1.5582 10.6802 0.0012E. solidaginis ovipunctures� block 14 0.2896 1.9849 0.018
observer 3 1.9459 13.3371 ,0.0001error 371 0.1459
Mature E. solidaginis galls� stem morph 1 0.1625 0.2317 0.63block 14 1.0132 1.4443 0.13error 372 0.7015
E. solidaginis galls per stem morph 1 0.2071 0.1947 0.66ovipunctured ramet� block 14 1.7590 1.6989 0.054
observer 3 3.8285 3.6958 0.012error 331 1.0359
R. solidaginis galls� stem morph 1 3.364 21.9222 ,0.0001block 14 0.2583 1.6823 0.057error 372 0.1536
Note: Analyses were performed on transformed data to better meet assumptions of ANOVA.� Data were arcsine-square-root transformed.� Data were natural-log transformed.
December 2008 3279RESISTANCE BY DUCKINGR
eports

During the period in which they are nodding, it is likely
that candy-cane stems are less able to compete for light
with the erect stems. In fact, we have observed that
candy-cane stems will generally straighten out (become
erect) within a few days of being placed in heavy shade.
This phenomenon would seem to indicate that light
limitation can be a factor in candy-cane stems. Finally,
the resistance advantage of candy-cane stems may be
frequency dependent. Ducking may provide a large
advantage when the majority of stems do not duck, but
the advantage may attenuate as the frequency of
ducking plants rises. If candy-cane stems were to
approach a majority in a population, then the costs
(whatever they may be) may overshadow the shrinking
resistance benefits.
Several architectural resistance mechanisms have been
identified in other plant species (Marquis et al. 2002,
Kaitaniemi et al. 2004, Araujo et al. 2006). However, the
candy-cane strategy of goldenrod is different in that it is
a more active, almost behavioral trait. A candy-cane
stem starts out straight, then bends at just the time that
susceptibility to galling insects (and probably other
species) is the greatest. After the danger has passed, the
stems straighten back out, and the candy-cane stems are
just as tall as the erect stems by the time of flowering.
This growth pattern is tantamount to slow-motion
ducking, and to our knowledge, it is the first document-
ed instance of resistance by means of ducking.
The ducking strategy does not appear to be unique to
S. altissima, but we do not know how widespread it may
be. We have observed candy-cane stems in Pennsylvania
in two other old-field goldenrods: S. gigantea and S.
juncea. Both of these species host a community of apex-
attacking herbivores and would presumably also benefit
from a ducking strategy. However, we have never
observed candy-cane stems of S. rugosa, another old-
field goldenrod with apex-attacking herbivores. It seems
likely that the ducking strategy may be found in other
fast-growing, herbaceous plants in open habitats, where
individual stems are relatively conspicuous to flying
herbivores.
ACKNOWLEDGMENTS
We thank C. Blair and N. Dorchin for discussions andcomments on the manuscript. S. Anderson, K. McCrea, and A.Weis assisted with data collection in the 1984 field survey. B.Davis and A. Turbett assisted with planting the gardenexperiment in 2003; R. Fox, P. March, J. Partelow, and A.Snyder collected ovipuncture data in 2004; and P. Marchcollected rosette-gall data in 2004. Rolf and Annick Helbigkindly granted permission to collect goldenrod rhizomes ontheir property for the garden experiment. We acknowledge theDavid Burpee Chair of Plant Genetics endowment of BucknellUniversity for supporting this research. The National ScienceFoundation also provided financial support for parts of thestudy under the following grants: DEB-8205856 to W. G.Abrahamson, DEB-0343633 to W. G. Abrahamson and J. T.Irwin, and DEB-0515483 to W. G. Abrahamson and M. J.Wise. Any opinions, findings, and conclusions expressed in thispaper are those of the authors and do not necessarily reflect theviews of the National Science Foundation.
LITERATURE CITED
Abrahamson, W. G., and A. E. Weis. 1997. Evolutionaryecology across three trophic levels: goldenrods, gallmakers,and natural enemies. Princeton University Press, Princeton,New Jersey, USA.
Alonso, C., and C. M. Herrera. 1996. Variation in herbivorywithin and among plants of Daphne laureola (Thymelaea-ceae): correlation with plant size and architecture. Journal ofEcology 84:495–502.
Araujo, A. P. A., J. D. A. De Paula, M. A. A. Carneiro, andJ. H. Schoereder. 2006. Effects of host plant architecture oncolonization by galling insects. Austral Ecology 31:343–348.
Bach, C. E. 1981. Host plant growth form and diversity: effectson abundance and feeding preference of a specialistherbivore, Acalymma vittata (Coleoptera: Chrysomelidae).Oecologia 50:370–375.
Craig, T. P., J. K. Itami, C. Shantz, W. G. Abrahamson, J. D.Horner, and J. V. Craig. 2000. The influence of host plantvariation and intraspecific competition on ovipositionpreference and offspring performance in the host races ofEurosta solidaginis. Ecological Entomology 25:7–18.
Espırito-Santo, M. M., F. d. S. Neves, F. R. Andrade-Neto,and G. W. Fernandes. 2007. Plant architecture and meristemdynamics as the mechanisms determining the diversity ofgall-inducing insects. Oecologia 153:353–364.
Grubb, P. J., and J. B. C. Jackson. 2007. The adaptive value ofyoung leaves being tightly folded or rolled on monocotyle-dons in tropical lowland rain forest: an hypothesis in twoparts. Plant Ecology 192:317–327.
Hartnett, D. C., and W. G. Abrahamson. 1979. The effects ofstem gall insects on life history patterns in Solidagocanadensis. Ecology 60:910–917.
Haysom, K. A., and C. Coulson. 1998. The Lepidoptera faunaassociated with Calluna vulgaris: effects of plant architectureon abundance and diversity. Ecological Entomology 23:377–385.
Hess, M. D., W. G. Abrahamson, and J. M. Brown. 1996.Intraspecific competition in the goldenrod ball-gallmaker(Eurosta solidaginis): larval mortality, adult fitness, oviposi-tional and host-plant response. American Midland Naturalist136:121–133.
Horner, J. D., and W. G. Abrahamson. 1992. Influence of plantgenotype and environment on oviposition preference andoffspring survival in a gallmaking herbivore. Oecologia 90:323–332.
Kaitaniemi, P., H. Vehvilainen, and K. Ruohomaki. 2004.Movement and disappearance of mountain birch defoliatorsare influenced by the interactive effects of plant architectureand induced resistance. Ecological Entomology 29:437–446.
Karban, R., and S. Courtney. 1987. Intraspecific host plantchoice: lack of consequences for Streptanthus tortuosus(Cruciferae) and Euchloe hyantis (Lepidoptera: Pieridae).Oikos 48:243–248.
Kareiva, P., and R. Sahakian. 1990. Tritrophic effects of asimple architectural mutation in pea plants. Nature 345:433–434.
Larson, K. C., and T. G. Whitham. 1997. Competition betweengall aphids and natural plant sinks: plant architecture affectsresistance to galling. Oecologia 109:575–582.
Lawton, J. H. 1983. Plant architecture and the diversity ofphytophagous insects. Annual Review of Entomology 28:23–39.
Leimu, R., and J. Koricheva. 2006. A meta-analysis of tradeoffsbetween plant tolerance and resistance to herbivores:combining the evidence from ecological and agriculturalstudies. Oikos 112:1–9.
Maddox, G. D., and R. B. Root. 1987. Resistance to 16 diversespecies of herbivorous insects within a population ofgoldenrod, Solidago altissima: genetic variation and herita-bility. Oecologia 72:8–14.
MICHAEL J. WISE AND WARREN G. ABRAHAMSON3280 Ecology, Vol. 89, No. 12R
epor
ts

Marquis, R. J. 1992. Selective impact of herbivores. Pages 301–325 in R. S. Fritz and E. L. Simms, editors. Plant resistanceto herbivores and pathogens. University of Chicago Press,Chicago, Illinois, USA.
Marquis, R. J., J. T. Lill, and A. Piccinni. 2002. Effect of plantarchitecture on colonization and damage by leaftyingcaterpillars of Quercus alba. Oikos 99:531–537.
McCrea, K. D., and W. G. Abrahamson. 1987. Variation inherbivore infestation: historical vs. genetic factors. Ecology68:822–827.
McEvoy, M. V. 1988. The gall insects of goldenrod (Compo-sitae: Solidago) with a revision of the species Rhopalomyia(Diptera: Cecidomyiidae). Thesis. Cornell University, Ithaca,New York, USA.
Merton, L. F. H., D. M. Bourn, and R. J. Hnatiuk. 1976. Gianttortoise and vegetation interactions on Aldabra Atoll. Part 1:inland. Biological Conservation 9:293–304.
Oghiakhe, S., L. E. N. Jackai, and W. A. Makanjuola. 1992.Cowpea plant architecture in relation to infestation anddamage by the legume pod borer, Maruca testulalis Geyer(Lepidoptera: Pyralidae). 2. Effect of pod angle. InsectScience and Its Application 13:339–344.
Oghiakhe, S., L. E. N. Jackai, and W. A. Makanjuola. 1993.Cowpea plant architecture in relation to infestation anddamage by the legume pod borer, Maruca testulalis Geyer(Lepidoptera: Pyralidae). 3. Effects of plant growth habit.Insect Science and Its Application 14:199–203.
Palo, R. T., J. Gowda, and P. Hogberg. 1993. Species heightand root symbiosis, two factors influencing antiherbivoredefense of woody plants in East African savanna. Oecologia93:322–326.
Raman, A., and W. G. Abrahamson. 1995. Morphometricrelationships and energy allocation in the apical rosette gallsof Solidago altissima (Asteraceae) induced by Rhopalomyia
solidaginis (Diptera: Cecidomyiidae). Environmental Ento-mology 24:635–639.
Root, R. B. 1996. Herbivore pressure on goldenrods (Solidagoaltissima): its variation and cumulative effects. Ecology 77:1074–1087.
Root, R. B., and N. Cappuccino. 1992. Patterns in populationchange and the organization of the insect communityassociated with goldenrod. Ecological Monographs 62:393–420.
Rudgers, J. A., and K. D. Whitney. 2006. Interactions betweeninsect herbivores and a plant architectural dimorphism.Journal of Ecology 94:1249–1260.
Stinner, B. R., and W. G. Abrahamson. 1979. Energetics of theSolidago canadensis stem gall insect–parasitoid guild interac-tion. Ecology 60:918–926.
Uhler, L. D. 1951. Biology and ecology of the goldenrod gallfly, Eurosta solidaginis (Fitch). Memoir 300. Cornell Univer-sity Agricultural Experiment Station, Ithaca, New York,USA.
Walton, R., A. E. Weis, and J. P. Lichter. 1990. Ovipositionbehavior and response to plant height by Eurosta solidaginisFitch (Diptera: Tephritidae). Annals of the EntomologicalSociety of America 83:509–514.
Wise, M. J., W. G. Abrahamson, and K. Landis. 2006a.Edaphic environment, gall midges, and goldenrod clonalexpansion in a mid-successional old-field. Acta Oecologica30:365–373.
Wise, M. J., D. L. Kieffer, and W. G. Abrahamson. 2006b.Costs and benefits of gregarious feeding in the meadowspittlebug, Philaenus spumarius. Ecological Entomology 31:548–555.
Wise, M. J., J. M. Partelow, K. J. Everson, M. K. Anselmo, andW. G. Abrahamson. 2008. Good mothers, bad mothers, andresistance to herbivory in Solidago altissima. Oecologia 155:257–266.
December 2008 3281RESISTANCE BY DUCKINGR
eports













