Embed Size (px)
Citation preview

Neurourology and Urodynamics 11:225-238 (1992)
Early Molecular Response to Rabbit Bladder Outlet Obstruction Ralph Buttyan, Benjamin 2. Jacobs, Jerry G. Blaivas, and Robert M. Levin
Department of Urology, Columbia University, New York, New York (R.B., B.Z. J., J.G.B.); Department of Urology, School of Medicine, University of Pennsylvania, Philadelphia (R. M. L.)
Partial obstruction of the rabbit bladder outlet induces a rapid hypertrophy characterized by increased bladder weight and smooth muscle content in addition to increased collagen deposition. In experiments designed to examine the genetic basis of this response, we evaluated the expression of a multitude of gene products in the rabbit bladder during the first 48-hr period following placement of a urethral cuff blocking the rabbit bladder outlet by 90%. Individual bladders were obtained from unoperated rabbits, from sham-operated rabbits and from rabbits at 2, 6, 10, 24, and 48 hr after obstruction. Poly (A)+ mRNA was extracted from each of these tissues and Northern blots made from the RNAs were hybrid- ized repeatedly with a series of probes. Corrected densitometry was utilized to assign a value to each transcript and to follow the change in expression of a given gene product over time.
Although several of the gene products we analyzed for were undetectable or un- changed over this brief period, we were able to detect a substantial sustained increase in the 4.5 kb mRNA encoding basic fibroblust growth factor (bFGF) initiated by 2 hr after obstruction. Later, at 6-24 hr, we measured transient induction of transcripts encoding c-myc, heat shock 70K (hsp-70), Hu-, and N-rus. In contrast, the constitutively-expressed 2.4 kb transcript for transforming growth factor-p (TGF-p) transiently declined to 50% of control levels at 24 hr following obstruction. In summary, our results provide a picture of tissue in transition, with synthesis of certain growth-stimulating and stress-response prod- ucts (bFGF, myc, rus, and hsp-70) increasing and a growth-inhibitory substance (TGF-p) decreasing. 0 1992 Wiley-Liss, Inc
Key words: bladder hypertrophy, growth factors, oncogenes
INTRODUCTION
Benign human bladder pathology can develop subsequent to any chronic con- dition that restricts the outflow of urine, such as obstructive prostate disease. This bladder pathology is often characterized by an increase in bladder mass, decreases in urinary flow and compliance, and residual urine volume remaining following urina-
Received for publication October 28, 1991; accepted February 4, 1992.
Address reprint requests to Ralph Buttyan, Ph.D., Department of Urology, Columbia University, 630 W. 168th Street, New York, NY 10032.
0 1992 Wiley-Liss, Inc.

226 Buttyan et al.
tion [Sterling et al., 1983; Levin et al., 19901. Although it is clear from experimental studies that outlet obstruction induces marked alterations in both the structure and function of the bladder [Mattiasson et al., 1987; Steers and De Groat, 1988; Karim et al., 19901 there is little information concerning the biological factors that mediate this response.
The rabbit model of outlet obstruction has been studied extensively on both a physiologica1 and pharmacological level to gain a better understanding of the devel- opment of human bladder pathology [Mayo and Hinman, 1976; Ghoniem et al., 1986; Malkowicz et al., 19861. As demonstrated by numerous investigations of this model, surgical obstruction of the rabbit urethra results in rapid changes in bladder morphol- ogy, contractile function, and protein chemistry [Malkowicz et al., 1986; Kato et al., 1990; Cher et al., 1990; McConnell et al., 1990; Kim et al., 19901. Briefly, outlet obstruction induces a 5- to 10-fold increase in bladder mass (from 1.5 g to 8-15 g) within the first week [Malkowicz et al., 1986; Levin et al., 1990; Kato et al., 19901. The contractile response to both field stimulation and postsynaptic receptor stimula- tion decreases substantially within 1 day of obstruction and partially recovers by 1 week [Malkowicz et al., 1986; Levin et al., 19901. Additionally, there are significant alterations in the ratio of myosin isozymes and cytoskeletal proteins detected during the first week following outlet obstruction [Cher et al., 1990; McConnell et al., 1990; Kim et al., 19901. As we demonstrate here, the rabbit outlet obstruction model is also a useful system to begin to understand the genetic basis of bladder pathology devel- oping as a result of obstructive stimuli.
To this end, we have initiated a study to identify potential genetic components of the early response to outlet obstruction. Our approach to this problem is based on the rationale that a tissue’s response to stressful stimuli is driven by changes in the activity of specific genes, the products of which enable the cells in the tissue to properly adapt to the stress. Therefore, by measuring the expression of particular genes in this tissue which is undergoing remodeling, we might be able to establish both the temporal response parameters and the extent to which the tissue is initiating proliferative, hypertrophic, or degenerative processes in response to the obstruction.
We have chosen to study bladder gene activity during the first 48 hr following partial urethral obstruction with the use of Northern blot analytical techniques. This method allows us to estimate the relative expression of mRNA for any given gene product in a tissue such as the bladder. Northern blot analysis has several benefits over other forms of analysis. These benefits include its predictable specificity; bands detected by hybridization can be matched to the known size of the mRNA transcript. In addition, one has the ability to repeatedly hybridize a single Northern blot and thus we are able to analyze for the expression of over 20 gene products in a single experiment. This technique provides a quantifiable assay; densitometry of X-ray films allows us to assign a number to the hybridization and these numbers can be compared as a function of time following obstruction. Finally, probes are readily available for a wide variety of gene products; therefore we can examine for the expression of growth factors, proto-oncogenes, structural proteins, and stress response proteins in the same experiment. With the use of this technique, we have already identified certain gene products that might play an important role in the acute response of the bladder to obstructive stress.

Molecular Response to Bladder Obstruction 227
MATERIALS AND METHODS Animals and Tissues
Age-matched male New Zealand White rabbits (3-5 kg) were obtained from Ace Animals and were maintained with food and water ad libitum as recommended by the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. Rabbits were anesthetized with 50 mg/kg pentobarbitol prior to surgery. Under sterile conditions, the bladder was catheterized (via the urethra) with an 8 Fr foley catheter. The bladder and urethra were then surgically exposed through a mid-abdominal incision. A 0-silk suture was snugly tied around the catheterized urethra and the catheter was removed to produce moderate urethral obstruction. The incision was closed and the animal was allowed to recover. At the indicated times during the first 48 hr after obstruction, rabbits were sacrificed by pentobarbital over- dose (100 mg/kg) and the bladder was recovered, frozen under liquid nitrogen and then stored in a -80°C freezer prior to processing for RNA extraction. Some rabbits were sham-operated by temporarily receiving the catheter followed by surgical in- tervention into the peritoneal cavity, without urethral ligation. These rabbits were treated as for the experimental obstruction group.
Extraction and Analysis of RNA
Individual bladders were pulverized to a powder under liquid nitrogen. Total RNA was extracted from the frozen powder by a modification of a LiCl precipitation method [Buttyan et al., 19881. The poly(A)+ mRNA fraction was then selected from the total RNA by oligo (dT)-cellulose chromatography (Collaborative Research Inc., Bedford, MA) [Aviv and Leder, 19721 and was quantitated by spectrophotometry at 260 nm. Five aliquots of poly (A)+ RNA from each specimen were electrophoresed on a 1.2% agarose-formaldehyde gel. The RNA in the gel was transferred to a nylon filter by capillary blotting methods. The resulting Northern blot was sequentially hybridized to a series of 32P-labeled probes as are listed below. The probes were labeled with a-32P by the nick translation method using purified cDNA inserts [Rigby et al., 19771. Labeled DNA probe was denatured by boiling, then hybridized to the Northern blot as described [Buttyan et al., 19881. Following overnight hybridization at 42”C, the blots were washed in a successive series of solutions containing decreas- ing amounts of SSC (saline-sodium citrate buffer) in the presence of 0.1% SDS and 5 mM EDTA. The filter was exposed to Kodak XAR-5 film for autoradiography. The intensity of hybridization to any given probe was measured by quantitative densi- tometry of the transcript band. Following a short rinse in boiling water, Northern blots were repeatedly hybridized up to 5 times. In the 6th cycle, each blot was hybridized to a probe for 18s ribosomal RNA and the autoradiograph of this hybrid- ization provided a means to quantitate RNA loading.
Quantitation of mRNA Expression
X-ray autoradiographs were scanned on a Molecular Dynamics Scanning Laser Densitometer to produce an image. Selected areas of the computer image containing individual bands of hybridization were then identified for densitometric quantitation by means of a “grid” selection program. This program allowed us to assign equiv- alent areas to each band on the autoradiograph. The densities obtained were corrected

228 Buttyan et al.
for the film background as measured from an empty grid section adjacent to the transcript bands. Prior to any measurement, the densitometer was calibrated for linearity by scanning the autoradiograph of a slot blot containing increasing amounts of rat ventral prostate mRNA, hybridized to the radiolabeled probe for C3, a subunit of the major prostate secretory protein. This calibration provided the linear scale of film density and identified the film saturation density beyond which quantitative measurement was unreliable. The only transcript band that exceeded this film satu- ration was the 24 hr point of the hsp-70 hybridization experiment; thus our estimate of the induced expression of this gene (10-12-fold) is only a minimum estimate. The density of the 18s rRNA band for the first lane on any given blot was assigned 1 .O and adjacent lanes were then corrected for RNA loading inequities by comparison to the first lane. Densities of individual transcript bands were then multiplied by this correction factor to produce the “relative transcript band density” measurement, utilized for graphical presentation of our data.
Hybridization Probes
The following cDNA probes were utilized for hybridization of Northern blots: c-fos from Tom Curran, Roche Institute of Molecular Biology; c-myc from Frederick Alt, Columbia University, basic fibroblast growth factor (bFGF) from Judy Abra- hams, California Biotechnology, Inc., 18s rRNA from Dr. Ramreddy Guntaka, Uni- versity of Missouri at Columbia; heat shock-68 (hsp-70) from Larry Moran, Univer- sity of Toronto; acidicfibroblast growth factor (aFGF), nerve growth factor (NGF), epidermal growth factor (EGF), heavy chain myosin, and transforming growth fac- for+ (TGF-P) from the American Type Culture Collection, Rockville, MD; Ha-ras, Ki-ras, N-ras, c-sis, and I-myc from Oncor, Inc., Gaithersburg, MD.
RESULTS
Poly (A) + mRNA was extracted from unoperated individual rabbit bladders and from a series of individual rabbit bladders removed at 2, 6, 10, 24, or 48 hr following the surgical placement of a urethral cuff that reduced urine outflow from the bladder by 90%. Five microgram aliquots of RNA from each specimen were co-electro- phoresed in adjacent lanes of a denaturing agarose gel. Gels were blotted onto nylon filter paper and the resulting Northern blots were hybridized to 32P-labeled cDNA probes, as indicated. Following suitable exposure to X-ray film for autoradiography, the blots were washed to remove hybridized radiolabeled probe and then rehybridized to a different probe. Following a succession of up to 5 sequential hybridizations, the blots were washed and then hybridized one final time to a probe for 18s ribosomal RNA to provide a means to estimate potential RNA loading inequities.
Many of the probes that we utilized showed no hybridization to our Northern blots. In this category were c-fos, aFGF, NGF, EGF, I-myc, Ki-ras, and c-sis. These results do not necessarily mean that these gene products are not expressed by the normal rabbit bladder or within 48 hr following obstruction. Rather it means that if they are made, they are below our sensitivity of detection using this technique. Heavy-chain myosin transcript (1.8 kb) was present at a low level in the normal rabbit bladder and never changed in intensity over the 48 hr of the experiment (not shown here). In contrast, several gene products showed distinct and marked changes in

Molecular Response to Bladder Obstruction 229
concentration during the first 48 hr following bladder outlet obstruction. Perhaps the most prominent change was in the expression of the 2.5 kb mRNA encoding the mammalian heat shock protein, hsp-70. As shown in Figure 1, this transcript was elevated in all specimens of obstructed rabbit bladder, compared to control, unoper- ated rabbit bladders. At the 10 hr obstruction, the transcript was approximately 10-fold more abundant than in control. Early elevation was also demonstrated for expression of mRNA encoding bFGF. This gene product is a mammalian growth factor, particularly noted as an agent that stimulates proliferation of fibroblasts and promotes angiogenesis. The single 4.8 kb transcript detected by this probe was not detected in RNA from control rabbit bladder, yet was readily identified in the RNAs of all the obstructed bladders, beginning at 2 hr following obstruction (Fig. 2). As with hsp-70, this transcript was most prominent in the 10 hr obstructed specimen. Two types of proto-oncogenes were more highly expressed during early obstruction. The 2.4 kb transcript encoding c-myc, a gene product often associated with the onset of cellular proliferation was noted to be elevated in the post-obstruction bladder RNAs (Fig. 3 ) . This transcript was not detected until 6 hrs post-obstruction and was maximally expressed in the 24 hr obstructed specimen. Finally, transcripts encoding the proto-oncogenes cHu-ras (1.2 kb) and N-rus (1.55 kb) were also conspicuously elevated post-obstruction (Fig. 4). It is of interest that these two members of the rus family demonstrate a significantly different temporal pattern of induction (highly elevated only in the 24 and 48 hr specimens) compared to the other genes in this study. Transcripts encoding the third noted number of this proto-oncogene family, cKi-ras, were never detected in any of these specimens.
Of all the gene products that we probed for only one, TGF-P, demonstrated an apparent primary decline in expression during the early post-obstruction period. The 2.5 kb transcript encoding this polypeptide growth factor was constitutively expressed in control rabbit bladder, and its concentration declined to approximately 50% control during the first 24 hr post-obstruction (Fig. 5). This early decline is transient and there was regain in concentration in the 48 hr specimen.
Since the previous experiments were done with RNAs extracted from individual rabbit bladders, we wanted to determine the extent to which any of these findings were reproducible. Therefore two groups of rabbits were operated, 4 rabbits receiving a sham-operation in which the urethra was surgically exposed and manipulated, but was never ligated while 4 rabbits received surgery and the partial urethral restriction. Twenty-four hours later, all rabbits were sacrificed, the bladders were recovered and processed individually for extraction of poly (A) + mRNA. Five microgram aliquots of mRNA from each specimen were run in adjacent lanes of a formaldehyde-agarose gel. Northern blots made from these gels were probed to identify and quantitate transcripts for hsp-70 or TGF-P. A final hybridization to cDNA for 18s rRNA was utilized to standardize RNA loading on these blots. As shown in Figure 6, the expression of the 2.5 kb transcript encoding hsp-70 is elevated approximately 9- 10-fold in the obstructed bladder group relative to the sham-operated control blad- ders. In contrast, our analysis for TGF-P expression demonstrates the opposite trend, namely that the 2.5 kb transcript for TGF-P is reduced approximately 50% compared to the sham-operated specimens (Fig. 7). Both of these findings reiterate the results obtained by single-point analysis of individual specimens at hourly intervals follow- ing partial urethral obstruction.
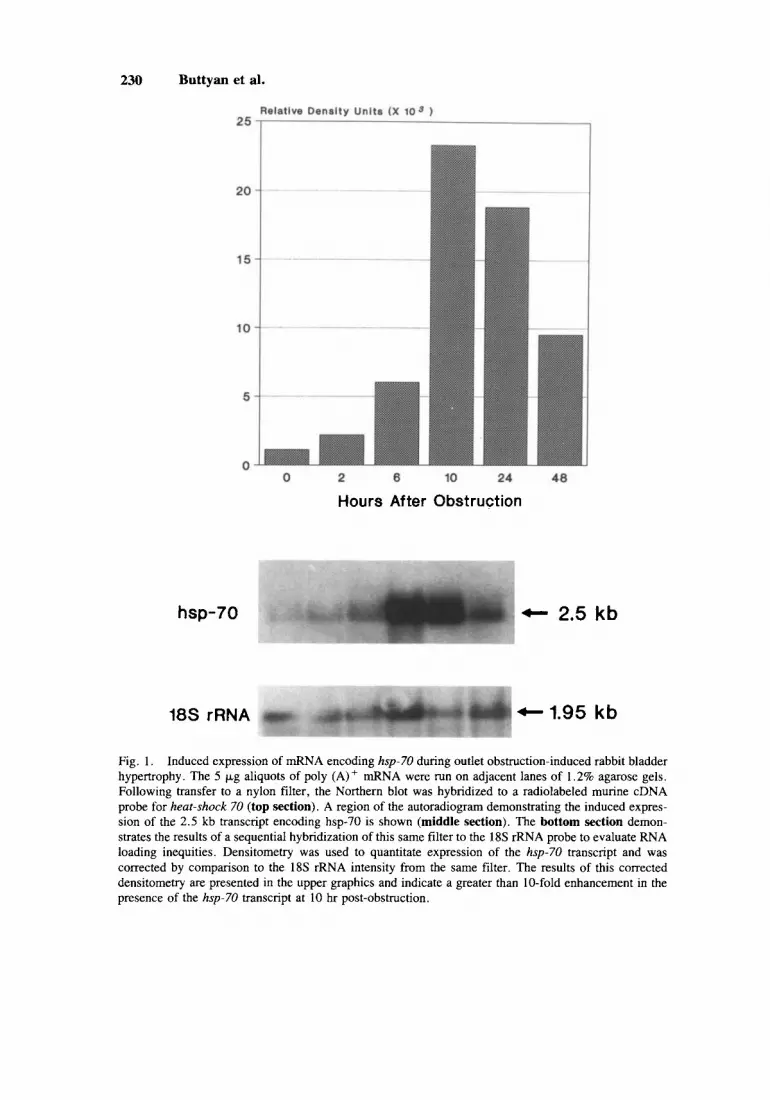
230 Buttyan et al.
Hours After Obstruction
hsp-70 + 2.5 kb
18s rRNA + 1.95 kb
Fig. 1. Induced expression of mRNA encoding hsp-70 during outlet obstruction-induced rabbit bladder hypertrophy. The 5 pg aliquots of poly (A)+ mRNA were run on adjacent lanes of 1.2% agarose gels. Following transfer to a nylon filter, the Northern blot was hybridized to a radiolabeled murine cDNA probe for heat-shock 70 (top section). A region of the autoradiogram demonstrating the induced expres- sion of the 2.5 kb transcript encoding hsp-70 is shown (middle section). The bottom section demon- strates the results of a sequential hybridization of this same filter to the 18s rRNA probe to evaluate RNA loading inequities. Densitometry was used to quantitate expression of the hsp-70 transcript and was corrected by comparison to the 18s rRNA intensity from the same filter. The results of this corrected densitometry are presented in the upper graphics and indicate a greater than 10-fold enhancement in the presence of the hsp-70 transcript at 10 hr post-obstruction.

bFGF
Molecular Response to Bladder Obstruction 231
Hours After Obstruction
- 4.8 kb
0 2 6 10 24 48 Fig. 2. Expression of bFGF mRNA during outlet-obstruction-induced rabbit bladder hypertrophy. The lower section demonstrates autoradiograph of rabbit bladder hypertrophy Northern blot following hy- bridization with bFGF cDNA probe. A 4.8 kb transcript encoding this growth factor is not detected in RNA from unobstructed rabbit bladder (0) but is present in all specimens subsequent to obstruction (2-48 hr). The upper section is a graphic presentation of quantitative densitometry for bFGF expression corrected for hybridization to 18s rRNA.
SUMMARY AND CONCLUSIONS
As measured by our Northern blot assay, partial outlet obstruction leads to distinct, rapid and reproducible changes in the genetic activity of the rabbit bladder.

232 Buttyan et al.
c-myc
Hours After Obstruction
1 2.4 kb
0 2 6 10 24 48 Fig. 3. Expression of c-myc mRNA during outlet obstruction-induced rabbit bladder hypertrophy. The lower section demonstrates autoradiograph of rabbit bladder hypertrophy Northern blot following hy- bridization with c-myc cDNA probe. The 2.5 kb transcript encoding this proto-oncogene is detected only subsequent to 6 hr obstruction. The upper section is a graphic presentation of quantitative densitometry for c-myc expression corrected for hybridization to 18s rRNA.
Since it is these genetic changes that propagate the physical response to the obstruc- tive stress, it is now our goal to resolve the potential meaning of the genetic changes that we have already identified. By comparing well-characterized alterations in the physiological, biochemical, and morphological state of the rabbit bladder following obstruction with our observations concerning genetic changes, it is possible to make some preliminary suggestions concerning the relevance of our observations.
Partial outlet obstruction induces a rapid and substantial increase in rabbit

Molecular Response to Bladder Obstruction 233
Fig. 4. Expression of two rus-family genes during outlet obstruction-induced rabbit bladder hypertro- phy. The lower sections demonstrate autoradiographs of rabbit bladder hypertrophy following hybrid- ization with either the Ha-rus or N-rus cDNA probes. Transcripts encoding the Ki-rus were never detected by this method. The top section shows graphic presentation of quantitative densitometry for expression of these rus-genes corrected for hybridization to the 18s rRNA probe.
bladder mass which is characterized by marked augmentation of virtually all tissue compartments: epithelial, smooth muscle, interstitial connective, and serosal [see review by Levin et al., 19901. Clearly some of this increase is the result of cellular hypertrophy, especially of the smooth muscle compartment [Levin et al., 19901. Therefore it is of note that two of the ras-family genes (Ha- and N - ) are highly induced during the second day of our experiment, as has been previously observed for

234 Buttyan et al.
TGF-0
Hours After Obstruction
- 2.5 kb
0 2 10 24 48 Fig. 5 . Expression of TGF-P during outlet obstruction-induced rabbit bladder hypertrophy. The lower section demonstrates autoradiograph of rabbit bladder hypertrophy Northern bIot foIIowing hybridization to the TGF-P cDNA probe. The 2.5 kb transcript encoding this growth factor decreases in concentration following obstruction. The upper section shows graphic presentation of quantitative densitometry for expression of TGF-P corrected for hybridization to the 18s rRNA probe.
the cHa-ras gene from studies of the early period of compensatory renal hypertrophy in rats [Sawczuk et al., 19891. Moreover, a recent study [Monson et al., 19901 gives reason to believe that at least some of this increase is the result of hyperplasia. In this study, it was demonstrated that 3H-thymidine incorporation was substantially in- creased in the rabbit bladder at 24 hr following obstruction, reached a maximum at 72 hr and returned to control levels at 14 days. Autoradiography of 3H-thymidine labeled

Molecular Response to Bladder Obstruction 235
Control 24 Hour Obstructed
Fig. 6. Induced expression of hsp-70 in 24 hr obstructed rabbit bladders. RNA was extracted from 4 specimens of unobstructed rabbit bladders at 24 hr following a sham-operation (Control) and from 4 specimens of 24 hr obstructed rabbit bladders. The 5 pg aliquots of RNA from individual rabbit bladders were co-electrophoresed on agarose gels and were blotted onto a nylon filter to produce a Nylon blot. The blot was hybridized initially to the cDNA probe for hsp-70 and subsequently to a probe for 18s rRNA. The graph shows comparative corrected densitometry measurements for these two groups. Bars demon- strate standard error of group measurements.
bladders verified that during the first day following obstruction, virtually all the increased DNA synthetic activity was located within the urothelium [Monson et al., 19901. Our detection of highly induced c-myc expression within the first 48 hr following obstruction is, in fact, consistent with the onset of an early proliferative response in some cellular compartment of the bladder and it will be of interest to learn whether in situ analysis techniques will localize c-myc expression to the urothelium at this time.
More significant is our observation of the induced expression of the bFGF transcript within 2 hr following outlet obstruction. The elevated expression of this growth factor is sustained throughout the 48 hr of the experiment. Polypeptide growth factors, such as bFGF, are under increasing scrutiny for their role in initiating cellular response pathways such as proliferation. Since this growth factor is induced in the apparent absence of induced expression of other growth factors such as aFGF, EGF, NGF, or PDGF, it is possible that this substance is one of the primary motivators of outlet obstruction-induced bladder growth. One additional polypeptide growth factor that was examined for in this experiment, TGF-p, was striking because it behaved in

236 Buttyan et al.
300
250
200
150
100
50
0 Control 24 Hour
Obstructed
Fig. 7. Reduced expression of TGF-P in 24 hr obstructed rabbit bladders. The Northern blot described in Figure 6 was subsequently hybridized to cDNA probe for TGF-P. The graph shows comparative corrected densitometry measurements for unobstructed, 24 hr sham-operated bladders (Control), and for 24 hr obstructed rabbit bladders. Bars demonstrate standard error of group measurements.
opposition to bFGF expression. As with many other tissues, constitutive expression of this gene product is detectable as a 2.5 kb transcript in control rabbit bladder. However, in contrast to many of the other gene products studied in these experiments, the expression of TGF-P decreases during the first 24 hr after obstruction. Since TGF-P is most well known as a growth-inhibitor substance [Sporn and Roberts, 19901, this observation is probably not trivial. It is certainly notable that the expres- sion of a growth inhibitor (TGF-P) apparently declines during the same period in which expression of a growth stimulator (bFGF) increases.
A third interesting and potentially important finding relates to the marked in- crease in the mRNA for hsp-70. Hsp-70 encodes a protein that has proven to have many cellular actions. For example, the elevated expression of hsp-70 is known to coincide with entry into S-phase of cells in culture [Milarski and Morimoto, 19861, thus possibly being related to the above mentioned proliferative phenomenon in the obstructed rabbit bladder. In addition, it is of interest that this gene product partici- pates in cellular stress responses and the distention caused by bladder outlet obstruc- tion is sure to stress the cells of the bladder. Moreover, the protein encoded by hsp-70 has been implicated in alterations of the cytoskeletal intermediate-sized filaments [Welch et al., 19851 which have been shown via protein chemistry to be one of the

Molecular Response to Bladder Obstruction 237
bladder proteins modified during outlet obstruction [Kim et al., 19911. Increased hsp-70 expression might also affect the bladder mitochondria since it has been dem- onstrated that this protein is involved in a macromolecular transport through the outer mitochondrial membrane [Hartl and Neupert, 1991; Mizzen et al., 19911. This action, which would seemingly alter mitochondrial membrane permeability, may be related to the 50% decrease in the activity of specific mitochondrial enzymes, malate dehy- drogenase and citrate synthetase previously reported to occur during outlet obstruc- tion-induced bladder hypertrophy [Haugaard et al., in press]. The role of this gene in the bladder obstructive response deserves further analysis.
Although there were several gene products that we could not identify in any of the bladder mRNAs specimens that we examined, it should be emphasized that other techniques, such as S1 nuclease mapping and quantitative PCR methods, can be more sensitive than Northern blotting techniques. Therefore, until these methods are uti- lized in a study such as ours, we cannot yet rule out a role for these gene products in the response of the rabbit bladder to obstruction. In addition it is important to consider that the changes in bladder gene expression that we are measuring, are based on determining the concentrations of individual mRNA transcripts during outlet obstruc- tion. There is no guarantee without corroborating studies, that these changes in d N A will result in concomitant changes in the expression of the corresponding protein. Since it is the protein moiety encoded by these genes that will ultimately be responsible for any physical alterations of the bladder, it is imperative to follow up this study with studies of the individual proteins. We are currently initiating exper- iments to establish that the protein products of these genes are correspondingly modulated during early bladder hypertrophy to further demonstrate the significance of these findings.
ACKNOWLEDGMENTS
The authors acknowledge the expert technical assistance of Lisa Potter and Po-Ying Ng and the editorial assistance of Sheila Levin. This study was aided, in part, through the Urology Research Fund of Columbia University and Funding ob- tained through the National Institutes of Health (RO 1-DK26508, RO1-DK33559, and RO 1 -DK39740).
REFERENCES
Aviv H, Leder P (1972): Purification of biologically active globin messenger RNA by chromatography on oligo-thyrnidilic acid-cellulose. Proc Natl Acad Sci U.S.A. 77: 1408-1413.
Buttyan R, Zakeri Z, Lockshin R, Wolgernuth D (1988): Cascade induction of c-fos. c-myc and hear- shock 70K transcripts during regression of the rat ventral prostate gland. Mol Endocrinol 2:650-657.
Cher ML, Kamrn KE, McConnell JD (1990): Stress generation and myosin phosphorylation in the obstructed bladder. J Urol 143:355A (abstract).
Ghoniern GM, Regnier CH, Biancani P, Johnson L, Susset JG (1986): Effect of vesical outlet obstruction on detrusor contractility and passive properties in rabbits. J Urol 135: 1284-1289.
Hartl F-U, Neupert W (1991): Protein sorting to mitochondria: Evolutionary conservation of folding and assembly. Science 247:930-938, 1991.
Haugaard N, Potter L, Wein AJ, Levin RM (1990): Effect of partial obstruction of the rabbit urinary bladder on malate dehydrogenase and citrate synthetase activity. J Urol (in press).
Karirn 0, Van Koeveringe G, Mostwin J (1990): Development of abnormal voiding patterns following partial obstruction of the guinea pig urethra. J Urol 143:355A.

238 Buttyan et al.
Kato K, Wein AJ, Longhurst PA, Haugaard N, Levin RM (1990): The functional effects of long-term outlet obstruction on the rabbit urinary bladder. J Urol 143:600-606.
Kim Y, Samuel M, Levin RM, Chacko S (1990a): Stress generation and myosin phosphorylation in the obstructed bladder. J Urol 143:354A (abstract).
Kim Y, Samuel M, Levin RM, Chacko S (1991): Alterations of proteins associated with cytoplasmic filaments in the hypertrophied urinary bladder. Biochim Biophys Acta (in press).
Levin RM, Longhurst PA, Monson FC, Kato K, Wein AJ (1990): Effect of bladder outlet obstruction on the morphology, physiology and pharmacology of the bladder. Prostate (Suppl) 3:9-26.
Malkowicz SB, Wein AJ, Elbadawi A, Van Arsdalen K, Ruggieri MR, Levin RM (1986): Acute biochemical and functional alterations in the partially obstructed rabbit urinary bladder. J Urol 136: 1324-1 329.
Mattiasson A, Ekstrorn J, Larsson B, Uvelius B (1987): Changes in the nervous control of the rat urinary bladder induced by outflow obstruction. Neurourol Urodyn 6:37-45.
Mayo ME, Hinman F (1976): Structure and function of the rabbit bladder altered by chronic obstruction or cystitis. Invest Urol 14:6-9.
McConnell JD, Lee IL, Cher ML (1990): Expression of nonmuscle caldesmon in obstruction-induced detrusor hypertrophy. J Urol 143:367A (abstract).
Milarski KL, Morimoto RI (1986): Expression of human HSP 70 during the synthetic phase of the cell cycle. Proc Natl Acad Sci USA 83:95 17-9521.
Mizzen LA, Chang C, Garrels JG, Welch WJ (1991): Identification, characterization and purification of two mammalian stress proteins present in mitochondria: grp 75, a member of the hsp 70 family and hsp 58, a homolog of the bacterial groEL protein. J Biol Chem 264:20664-20675.
Monson FC, McKenna BA, Wein AJ, Levin RM (1990): Effect of outlet obstruction on ’H-thymidine uptake and metabolism: A radiographic and biochemical study. J Urol.
Rigby PWJ, Dieckmann M, Rhodes C, Berg P (1977): Labeling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I. J Mol Biol 113:237-242.
Sawczuk IS, Hoke G, Olsson CA, Connor J, Buttyan R (1989): Gene expression in response to acute unilateral ureteral obstruction. Kidney Int 35:1315-1319.
Sporn MB, Roberts AB (1990): TGF-P: Problems and prospects. Cell Regul 153754382. Steers WD, De Groat WC (1988): Effect of bladder outlet obstruction on micturation reflex pathways in
Sterling AM, Ritter RC, Zinner NR (1983): The physical basis of obstructive uropathy. In Hinman Jr. F
Welch WJ, Feramisco JR, Bloise SH (1985): The mammalian stress response and the cytoskeleton:
the rat. J Urol 140:864-871.
(ed): “Benign Prostatic Hypertrophy. ” New York: Springer-Verlag, pp 433-442.
Alterations in intermediate filaments. Ann NY Acad Sci 45557-67.





























