Embed Size (px)
Citation preview

Early piglet mortality in loose-housed sows
related to sow and piglet behaviour and
to the progress of parturition
Lene Juul Pedersen a,*, Erik Jørgensen b, Teresia Heiskanen c,Birgitte I. Damm c
a Danish Institute of Agricultural Sciences, Department of Animal Health,
Welfare and Nutrition, P.O. Box 50, 8830 Tjele, Denmarkb Danish Institute of Agricultural Sciences, Department of Genetics and Biotechnology,
P.O. Box 50, 8830 Tjele, Denmarkc The Royal Veterinary and Agricultural University, Department of Large Animal Science,
Grønnegardsvej 8, 1870 Frederiksberg C, Denmark
Accepted 24 June 2005
Available online 16 August 2005
Abstract
The aim of the present study was to identify characteristics of sow behaviour and parturition
related to early piglet mortality in loose-housed farrowing sows. Data from 152 farrowings that
originated from three different herds with loose-housed sows during parturition were used. Graphical
chain models were used to model the relationships between perinatal behaviour, periparturient
individual conditions (time of day of parturition, rectal temperature 1–3 days postpartum) and causes
of early piglet mortality. Modelling was based on the correlation between variables within herd and
farrowing batch.
The analysis showed that different causes of mortality were linked to different behavioural
variables during the periparturient period and that they grouped into three independent categories.
The first category was associated with stillbirth and death due to other causes. Stillbirth was
positively related to the variation of the inter-birth interval and negatively related to the percentage of
piglets that suckled during the first 8 h after birth of first piglet (BFP). Death due to other causes was
negatively related to the suckling activity during the post-partal period (9–24 h after BFP). The
second category was associated with piglet crushing, which was positively related to much lateral
www.elsevier.com/locate/applanim
Applied Animal Behaviour Science 96 (2006) 215–232
* Corresponding author.
E-mail address: [email protected] (L.J. Pedersen).
0168-1591/$ – see front matter # 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.applanim.2005.06.016

lying the last 4 h before BFP. Finally, the third category was associated with death due to lack of
colostrums ingestion of colostrum, which was linked to the time of parturition and sow rectal
temperature on days 1 and 2 after farrowing. Sows that gave birth during the morning compared to
evening/night had more dead piglets due to lack of colostrums ingestion. Death due to lack of
colostrums ingestion was also higher in sows with high rectal temperature on the day after BFP. The
results further showed that litter size not only influenced mortality but also behavioural variables.
High litter size was related to high nest building activity before BFP, low but more variable inter-birth
intervals, and much lateral lying after BFP. The study points towards several areas of interest for
further research that can help understand early piglet mortality. These include causes of variation in
the progress of parturition, causes of individual differences in sow activity and responsiveness to
piglets, and early management of farrowing. In addition, the study suggests possible indicators of
problematic farrowings rendering liveborn piglets at risk. These include prepartum lateral lying in the
sow, low suckling activity and rate of stillbirth.
# 2005 Elsevier B.V. All rights reserved.
Keywords: Farrowing behaviour; Graphical models; Sus Scrofa
1. Introduction
In sow herds, one of the most important causes of production loss is piglet mortality,
especially in the first few days after farrowing. Data from management information
systems indicate differences between populations both between and within systems. A
recent report (Jultved, 2004) showed that an average of 13.3% of the liveborn piglets die
before weaning in Danish sow herds. Mortality in modern housing systems for loose-
housed sows is rather variable ranging from 9 to 25% of liveborn piglets (Marchant et al.,
2000; Grandinson et al., 2002; Pedersen et al., 2003; Damm et al., 2005b). Thus, early
piglet mortality is of great economic importance but it also constitutes a welfare problem
because the piglets often die of hunger and various traumas caused by thread and crushing
(Fraser, 1990; Vaillancourt and Tubbs, 1992).
Irrespective of production system, the majority of death among piglets occurs within the
first two days postpartum (Bille et al., 1974; English and Smith, 1975; Vaillancourt and
Tubbs, 1992; Kongsted and Larsen, 1999). Stillbirth reports from both crates and loose-
housing systems range from 3 to 6% of the total litter when using references from studies
where post-mortem examinations have been carried out (Kongsted and Larsen, 1999;
Fraser et al., 1997; Damm et al., 2005a; Tuchscherer et al., 2000). Crushing seems to be the
main cause of death among the liveborn piglets and starving and/or hypothermic piglets are
particularly at risk (Fraser, 1990; Vaillancourt and Tubbs, 1992; Marchant et al., 2000;
Damm et al., 2005b). Thus, these factors are also important in the understanding of piglet
mortality.
The variation in early piglet mortality between litters within the same herd is
considerably larger than would be expected with constant mortality rate. Since the
heritability of early piglet mortality is rather low (Knol et al., 2002; Su et al., 2004;
Grandinson et al., 2002), the variation between litters within a herd may to a great extent be
accounted for by non-genetic differences caused by, for example differences in climate,
management and/or sow conditions at the particular farrowing. For example, management
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232216

of night-time farrowings may differ from that of day-time farrowings and lameness and/or
obesity may only occur in some sows.
The relationship between mortality and individual conditions around parturition is not
well known. Larger data sets on mortality exist both from experimental herds and from
production herds. Such data sets often lack information about individual conditions around
parturition, making it difficult to point at potential causal mechanisms underlying individual
differences in mortality. In order to focus the experimental design, there is a need beforehand
to identify potential causal mechanism. Multivariate data analysis may be used in an
exploratory phase to identify such potential mechanism underlying the variability. In this
context, graphical models, and especially the so-called chain-graph models are promising
techniques (Wermuth, 2003). The latter technique allows a partitioning of the variables based
on prior knowledge, corresponding to, e.g. Risk factors, Diseases and Symptoms.
In the present study, the techniques of graphical models were used to gain insight into
the relations between early piglet mortality in loose housing systems and perinatal sow and
piglet behaviour and individual conditions around parturition.
2. Materials and methods
2.1. Animals and housing
Data were collected from 152 farrowings. The 152 farrowings took place in three
different herds (Table 1), where sows were loose-housed during farrowing and lactation.
Farrowings within each herd were divided into batches. A batch was defined as a group of
sows that was introduced to the farrowing unit within a 2–4 days period. However, in about
a third of the cases (11 out of 30) it was necessary to include sows which farrowed further
apart in the same batch, in order to avoid batches with very few animals. In these cases the
batch was a compromise between closeness of day of introduction and the number of sows
in the batch.
The body condition of the sows at the time of introduction to the farrowing pens was
observed as well as the condition of the legs. All sows with abnormal body condition
(obese/emaciated sows) and/or with leg problems were excluded from the study.
The three herds differed with respect to pen design and management and the
characteristics of each herd are described below:
� Herd 1: The sows were pure-bred Danish Landrace sows. During farrowing the sows
were housed in Schmid pens (Schmid, 1991, 1993) with modifications that had been
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 217
Table 1
Summary of main production traits in the three herds (L: Danish Landrace, Y: Danish Yorkshire)
Herd Breed Litters Batches Total born
piglets per litter
Dead
per litter
Percentage
dead
Mean
parity
1 L � L 47 11 13.6 4.3 31.7 1.3
2 Y � L 55 15 12.5 3.1 24.3 2.4
3 Y � L 30 6 14.4 2.8 19.2 2.8
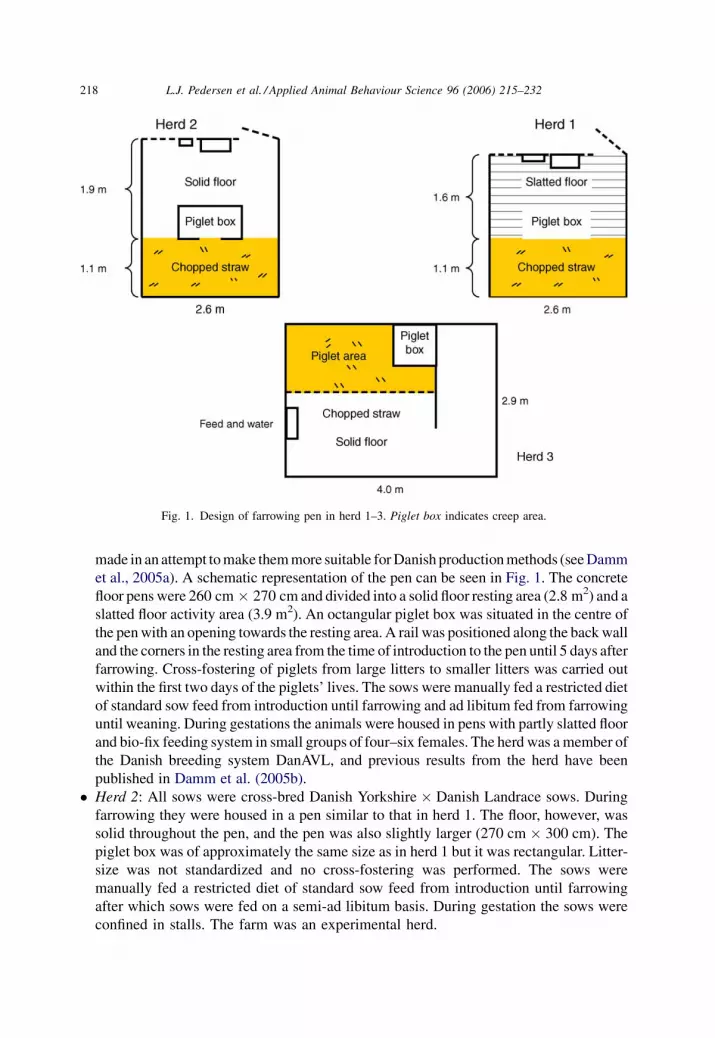
made in an attempt to make them more suitable for Danish production methods (see Damm
et al., 2005a). A schematic representation of the pen can be seen in Fig. 1. The concrete
floor pens were 260 cm � 270 cm and divided into a solid floor resting area (2.8 m2) and a
slatted floor activity area (3.9 m2). An octangular piglet box was situated in the centre of
the pen with an opening towards the resting area. A rail was positioned along the back wall
and the corners in the resting area from the time of introduction to the pen until 5 days after
farrowing. Cross-fostering of piglets from large litters to smaller litters was carried out
within the first two days of the piglets’ lives. The sows were manually fed a restricted diet
of standard sow feed from introduction until farrowing and ad libitum fed from farrowing
until weaning. During gestations the animals were housed in pens with partly slatted floor
and bio-fix feeding system in small groups of four–six females. The herd was a member of
the Danish breeding system DanAVL, and previous results from the herd have been
published in Damm et al. (2005b).
� Herd 2: All sows were cross-bred Danish Yorkshire � Danish Landrace sows. During
farrowing they were housed in a pen similar to that in herd 1. The floor, however, was
solid throughout the pen, and the pen was also slightly larger (270 cm � 300 cm). The
piglet box was of approximately the same size as in herd 1 but it was rectangular. Litter-
size was not standardized and no cross-fostering was performed. The sows were
manually fed a restricted diet of standard sow feed from introduction until farrowing
after which sows were fed on a semi-ad libitum basis. During gestation the sows were
confined in stalls. The farm was an experimental herd.
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232218
Fig. 1. Design of farrowing pen in herd 1–3. Piglet box indicates creep area.

� Herd 3: The sows were cross-bred Danish Yorkshire � Danish Landrace sows. During
farrowing the sows were housed in pens measuring 290 m � 400 m. The pens consisted
of a piglet area (150 m � 260 m) separated from the rest of the pen by vertical bars. In
the piglet area a heating panel was placed in the corner farthest away from the sow area
forming the creep area. The sow area was divided into a resting and a dunging area as
shown in Fig. 1. Cross-fostering of piglets from large litters to smaller litters was carried
out within the first two days of the piglets’ lives. Twice daily the sows were manually fed
a restricted ration before parturition. From farrowing until weaning they were fed on a
semi-ad libitum basis. During gestation the sows were confined in stalls. The farm was
an experimental herd.
The production traits in the three herds are shown in Table 1.
2.2. Maternal behaviour and reproduction
The behaviour of the sows and piglets was observed from 4 h prior to parturition until
24 h after Birth of first piglet (BFP) using 24 h time-lapse video recordings. One–zero
sampling (Martin and Bateson, 1986) at 15 min intervals was used to record whether the
sow performed nest building (pawing, rooting, carrying or arranging straw) and whether
she remained in lateral recumbency throughout the 15 min period. These recordings were
subsequently split into two time intervals: before BFP (from �4 to 0 h before BFP) and
after BFP (from 0 to 8 h after). The average number of 15 min periods where the sow
performed nest-building activities was calculated as well as the number of periods where
the sows lay in lateral recumbency during the whole 15 min period (LatBef, LatAft).
Furthermore, instantaneous scan sampling of the number of piglets suckling (teat in mouth
while performing sucking movements) as well as the number of piglets in contact with the
udder (piglet in contact with the udder without sucking on a teat) was carried out. The
average percentage of piglets suckling at the udder during the instantaneous samplings was
subsequently calculated from BFP until 8 h after (Suckl8) and from 9 h after BFP until 24 h
after (Suckl24). Similarly, the average percentage of piglets in contact with the udder was
calculated from BFP until 8 h (Udder8) after and from 9 h after BFP until 24 h after
(Udder24). In addition, the time from BFP to the time of birth of each subsequent piglet
was recorded. From this, the average inter-birth interval (IBInt) and the standard deviation
of the inter-birth interval (StdIBInt) were calculated.
On the day of parturition and the following two days, the rectal temperature of the sows
was measured (Temp0, Temp1, Temp2). The length of gestation (days), the total number of
piglets born by the same sow (TotBorn) and the number of the sows’ own piglets dying each
day after birth were recorded. All piglets that died while still being with their biological
mother (including still born piglets) were marked with date of death and sow number, and
then frozen. Each month the dead piglets were thawed and the cause of death was determined
by macroscopic examination and divided into four categories: still born (StillB), piglets dying
without injuries and without milk in their stomach (NoMilk), piglets dying from injuries
caused by the sow either with or without milk in their stomach (Crush) and piglets dying from
other causes including intestinal and infectious diseases and being culled by the stock person
due to low viability (OthCau). A detailed description of each category can be found in Damm
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 219

et al. (2005b). Note that some of the piglets that died from injuries (Crush) had not yet
ingested colostrum.
For use in the statistical analysis to model the effect of the diurnal variation of BFP the
sinus and cosinus of BFP were calculated, i.e. sin BFP = cos(BFP � 2p/24). Furthermore,
RelSuckl8 was calculated as the Proportion of piglets at the udder 0–8 h after BFP that
suckles, where RelSuckl8 = Suckl8/Udder8.
Variables included in the analysis are described and summarized in Table 2.
2.3. Statistical analysis
The model used was a graphical chain model (Lauritzen and Wermuth, 1989) also called
a block-recursive model (Højsgaard and Thiesson, 1995). A chain-graph model facilitates
the search for the optimal statistical model of a multivariate data set. In recent years,
standard software for estimation in these models have become available, such as MIM
(Edwards, 2000) and GraphFitI (Blauth et al., 2000). This has lead to a number of
applications especially within social science, Wermuth (2003) lists some examples and
Mohamed et al. (1998) illustrate the application in a study of infant mortality in humans. In
special cases, the directed links may be considered as modelling causal relations. We refer
to Lauritzen and Richardson (2002) for a detailed discussion of this aspect.
The independence structure of a multivariate data set can be shown as a so-called
conditional independence graph. Each variable in the data set is shown with a node (or
vertex), and edges (or links) between the nodes indicate whether the variables are
conditionally dependent. In chain-graph models the variables are partitioned into blocks. In
the present study, four blocks of variables were identified based on the temporal ordering of
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232220
Table 2
Variables included in the analysis of mortality
Description Abbreviation Block
Herd, batch, parity Herd, Batch, Parity Design
Total born piglets TotBorn Sow
Birth of first piglet BFP
Sinus component of BFP sin BFP
Cosinus component of BFP cos BFP Prenatal
Lateral lying before BFP LatBef
Intervals with nest building before BFP NestBef
Still born piglets StillB
Piglets that died without milk in stomach NoMilk
Piglets that died with injuries from the sow Crush
Piglets that died from other causes OthCau
Rectal temperature day 0, 1, 2 (8C) Temp0, Temp1, Temp2
Average inter-birth interval in the litter IBInt Postnatal
Standard deviation of the inter-birth interval (min) StdIBInt
Lateral lying the first 8 h after BFP LatAft
Intervals with nest building the first 8 h after BFP NestAft
Piglets suckling 0–8 and 9–24 h after BFP (%) Suckl8, Suckl24
Piglets at the udder 0–8 and 9–24 h after BFP (%) Udder8, Udder24

the variables. The first block consisted of variables related to the Design of the study with
the variables Herd, Batch and Parity. The second block was related to the individual Sow
and consisted only of Total number of piglets (TotBorn). The third block consisted of
variables observed in the Pre-natal period. Finally, the fourth block consisted of variables
observed in the Peri- and Postnatal period. The details of the blocking can be seen in
Table 2. This partitioning implies that links between variables within a block are required to
be undirected, whereas links between blocks are directed according to the ordering of the
blocks. A directed link indicates a dependency similar to the association between
explanatory and response variables in regression models and is indicated with an arrow
between the two nodes.
The Batch effect is a random effect, and mainly used to model the dependency between
observations within the same Batch. This introduces complications. First of all, no program
for analysis is readily available. Secondly, dependency between variables may exist at
several levels of the hierarchy. Therefore, we have chosen to correct the observations for
the effect of Batch before the chain-graph model analysis. The initial model runs showed
that many of the dependent variables were related to the herd, parity and total number of
born piglets. Therefore, the following model was estimated initially
YðkÞhi ¼ m
ðkÞh pðiÞ þ B
ðkÞhtðiÞ þ b1kXhi þ b2kX
2hi þ eðkÞhi (1)
where YðkÞhi is the i observed value of dependent variable k in the h herd, m
ðkÞh pðiÞ the ‘mean’
level of variable k in herd h and the parity p(i) corresponding to the ith observation. BðkÞhtðiÞ is
the random effect of batch t(i) within herd h, BðkÞhtðiÞ �Nð0; s2
BkÞ, b�k a regression coefficient
for the linear and the quadratic effect of total number of born piglets, XhieðkÞhi is the residual
assumed distributed as Nð0; s2ekÞ.
The ‘corrected’ values of the dependent variables YðkÞhi are thus calculated as
YðkÞhi ¼ Y
ðkÞhi � Y
ðkÞhi , where Y
ðkÞhi is the predicted values from the model in Eq. (1). The
calculations were made using the lme in the nlme package (Pinheiro et al., 2004) in R (R
Development Core Team, 2004).
The corrected values were analysed with a Block Recursive Model (Chain-graph) using
the MIM3.1-program (Edwards, 2000), where the variables are supplemented with a
discrete factor containing the herd � parity levels in block no. 1 (to investigate interactions
between herd � parity and the variables). The mimR-package (Højsgaard, 2003) was used
to call the MIM-program from R. The Bayesian Information Criteria (BIC) were used to
find the final model. The BIC usually leads to more parsimonious models than simple
iterative significance testing, where the number of tests severely invalidates the overall
level of significance. In order to avoid local minima in the model search, the search started
with a forward stepwise procedure from the model with only main effects. The ‘incoherent’
search strategy (i.e. repeated investigation of links) between decomposable graphs was
used. Three selection criteria were used: first the likelihood criteria, secondly the Akaike’s
information criteria (AIC) and third the BIC. Finally, a backward selection strategy with
BIC as the criterion was performed. This procedure was used because simple backwards
selection from the full saturated model consistently ended up with less satisfactory models
(higher BIC-values) than the forward search from a main effects only model. A detailed
description of the search procedures mentioned can be found in Edwards (2000). The
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 221

search strategy resulted in an overall fit better than both the backward and the forward
selection alone. Subsequently, the sub model candidates identified via the MIM program
were reformulated for analysis using standard mixed model formulation, and analysed
using the lme and GLMM function of R (Bates and Sarkar, 2004). In this final step
interactions and main design factors such as herd and parity were sequentially omitted
from the model if they were not significant. In most cases, the effect of these factors were
additive and results have been presented as least square means, using the average of the
herd and parity effect. In one case, the effect of herd interacted with the relation between
two behaviour variables, in this case we have presented the relationship for each herd.
The magnitude of the relationships between two variables was calculated as marginal
changes in the dependent variables as a function of a change in each independent variables
from the average level. Such calculations should, however, be treated with caution because
of the collinearity of the variables due to the non-experimental design of the data
collection.
Due to missing values for some of the temperature measurements the model found for
the full dataset without temperature, was supplemented with an additional model search
with the data with temperature included.
3. Results
Herd, Parity and Batch all influenced Total number of piglets born (TotBorn) and the
first step of the analysis was therefore to correct data for these design factors. TotBorn also
influenced most behavioural variables. The percentage of intervals with nest building
before birth (NestBef) increased from an average of 64.9 to 65.9% when litter size
increased from 13 to 14. An increase in TotBorn of one piglet resulted in a 9% decrease of
the Average inter-birth interval in the litter (IBInt), and an increase of Standard deviation
of the inter-birth interval (StdIBInt) with 6%. Lateral lying the first 8 h after BFP (LatAft)
increased slightly with 1.6% when litter size increased from 13 to 14 piglets.
The influence on specific causes of mortality is described in detail under each
subsection.
3.1. The total model: the structural relationship
After correction for Batch, the model selection resulted in the graphical model shown in
Fig. 2. To reduce the complexity of the figure the links from variables in the Design and
Sow block have been omitted.
The BIC of the final model was 4682 using 1640 degrees of freedom. The change in BIC
(which measures the effect of reducing the model with a particular variable) was larger
when links between two behavioural variables compared to a behavioural variable and a
mortality variable were omitted (see Fig. 2). The final graphical model showed that the four
different causes of death grouped into three groups related to different behavioural
variables. One group comprised Piglets that died without milk in stomach (NoMilk) and
variables related to sow rectal temperature after parturition and to time of birth of the first
piglet. Another comprised Still born piglets (StillB) and Piglets that died from other causes
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232222

(OthCau) and was related to suckling activity, birth interval and time spent in lateral
recumbency after birth. Finally, the third comprised Piglets that died with injuries from the
sow (Crush) and was related to time spent in lateral recumbency before birth, nest building
activity and sow rectal temperature on the day of parturition. The three groups were almost
independent of each other. The relationships between variables within each group are
described in detail in the following subsections.
3.2. Quantification of the submodels
3.2.1. Still born piglets (StillB)
The most common cause of death was stillbirth. The estimated mean value of Still born
piglets (StillB) was 7.2% of the total born piglets. The influence of Total number of piglets
born (TotBorn) was nonlinear with a marginal increase of 0.7% when TotBorn increased
from 13 to 14 piglets (see Fig. 3). As also seen in Fig. 3 StillB was directly linked to the
Standard deviation of the inter-birth interval (StdIBInt) in that increased StdIBInt resulted
in increased StillB.
StdIBInt was further positively correlated to Average inter-birth interval in the litter
(IBInt) in that a 1% increase in IBInt increased StdIBInt with 1.4%.
Stillbirth was also linked to Percentage of piglets suckling 0–8 h after BFP (Suckl8) and
Percentage of piglets at the udder 0–8 h after BFP (Udder8). However, these two variables
were highly correlated, as Udder8 is the maximum level of Suckl8. This resulted in
complicated models of the relationship. Therefore, in the detailed analysis Proportion of
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 223
Fig. 2. Selected block recursive model. See Table 2 for description of variable names. The nodes with a double
line indicate the four different mortality causes. The numbers at the links indicate marginal increase in BIC from
removing the link from the model. Removal of links indicated by (*) would lead to non-decomposable models
and have not been validated. BIC values at links marked with (*) were not calculated due to missing values. Links
from the Design and Sow are omitted to reduce the complexity of the figure.

piglets at the udder 0–8 h after BFP that suckles (RelSuckl8) was analysed instead of
Suckl8, and a more parsimonious model was found. As shown in Fig. 4, the expected level
of RelSuckl8 decrease with increasing number of StillB.
Sows with litters with low Suckl8 also showed little Lateral lying the first 8 h after BFP
(LatAft). However, there was no direct link between StillB and LatAft.
3.2.2. Piglets that died from other causes (OthCau)
On average 4.6% of the Total number of piglets born died of OthCau. OthCau increased
nonlinearly with a marginal increase of 0.6% when TotBorn increased from 13 to 14 piglets as
shown in Fig. 5. Death from OthCau primarily included piglets that died of disease or were
culled by the stock person due to poor viability. As can be seen in Fig. 2, this death cause was
related to thesamebehaviouralvariablesasstillbirth.OthCauwasdirectly linkedtoPercentage
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232224
Fig. 3. Estimated mortality (solid line) due to Still born piglets (StillB) as a function of Total number of piglets
born (TotBorn) (left) and Standard deviation of the inter-birth interval (StdIBInt) (right). The dotted lines
indicates 0.95 confidence interval. The rug indicates observed values.
Fig. 4. Relationship between number of stillborn piglets in the litter and the proportion of time spent suckling.
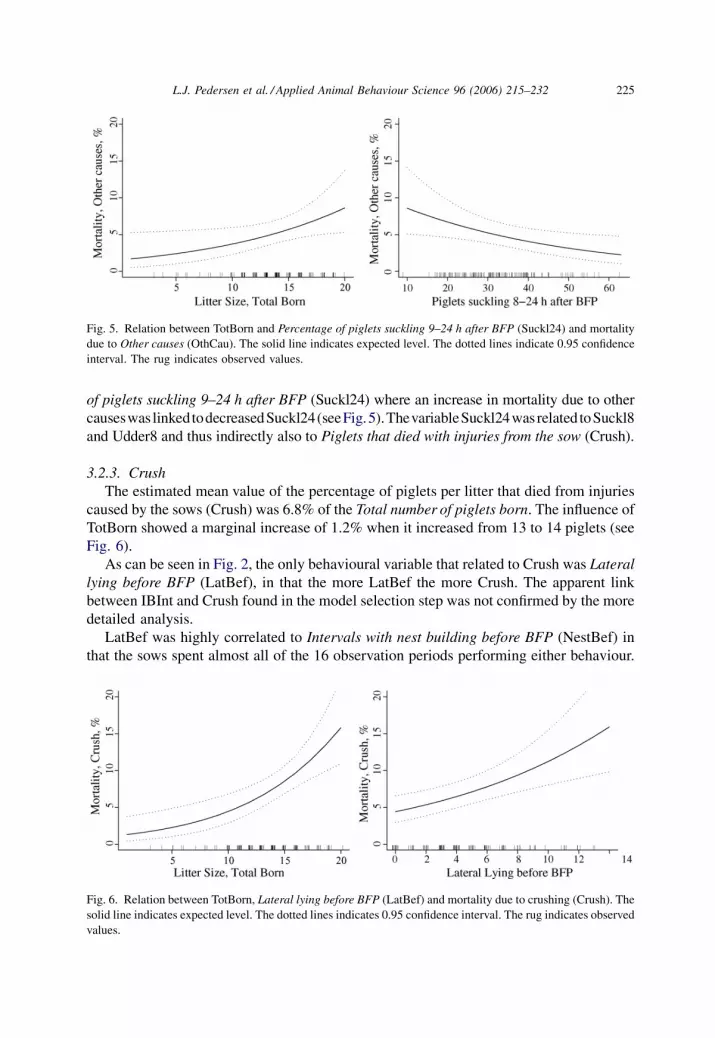
of piglets suckling 9–24 h after BFP (Suckl24) where an increase in mortality due to other
causeswas linkedtodecreasedSuckl24(seeFig.5).ThevariableSuckl24wasrelated toSuckl8
and Udder8 and thus indirectly also to Piglets that died with injuries from the sow (Crush).
3.2.3. Crush
The estimated mean value of the percentage of piglets per litter that died from injuries
caused by the sows (Crush) was 6.8% of the Total number of piglets born. The influence of
TotBorn showed a marginal increase of 1.2% when it increased from 13 to 14 piglets (see
Fig. 6).
As can be seen in Fig. 2, the only behavioural variable that related to Crush was Lateral
lying before BFP (LatBef), in that the more LatBef the more Crush. The apparent link
between IBInt and Crush found in the model selection step was not confirmed by the more
detailed analysis.
LatBef was highly correlated to Intervals with nest building before BFP (NestBef) in
that the sows spent almost all of the 16 observation periods performing either behaviour.
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 225
Fig. 5. Relation between TotBorn and Percentage of piglets suckling 9–24 h after BFP (Suckl24) and mortality
due to Other causes (OthCau). The solid line indicates expected level. The dotted lines indicate 0.95 confidence
interval. The rug indicates observed values.
Fig. 6. Relation between TotBorn, Lateral lying before BFP (LatBef) and mortality due to crushing (Crush). The
solid line indicates expected level. The dotted lines indicates 0.95 confidence interval. The rug indicates observed
values.

Thus, NestBef was related to Intervals with nest building the first 8 h after BFP (NestAft),
whereas there was no link between LatBef and LatAft. Sows with high NestBef also had
high NestAft (see Fig. 7) and high Rectal temperature day 0 (Temp0).
3.2.4. Piglets that died without milk in stomach (NoMilk)
The estimated mean percentage of piglet that died solely due to lack of colostrum uptake
(NoMilk) was 0.8% of the total number of piglets born. The detailed analysis (Fig. 2)
showed that NoMilk was related to BFP as described by a sinoid function and to Rectal
temperature day 1 (Temp1) as shown in Fig. 8.
Temp1 and Rectal temperature day 2 (Temp2) were strongly related whereas none of
them were related to Temp0.
4. Discussion
Several studies, including the present one, have shown that increased total number of
born piglets is followed by an increased rate of mortality (Bille et al., 1974; Roehe and
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232226
Fig. 7. Relation between nest building behaviour and lateral lying.
Fig. 8. Effect of farrowing start (BFP) (left) and temperature (Temp1) (right) on mortality due to no milk in the
stomach (NoMilk). The solid line indicates expected level. The dotted lines indicate 0.95 confidence interval. The
rug indicates observed values.

Kalm, 2000; Su et al., 2004). The present study further demonstrates that also behavioural
variables describing prepartum nesting behaviour and progress of parturition are
influenced by litter size. With increasing litter size sows performed more prepartum
nesting, had shorter but more variable inter-birth intervals and more partal–postpartal
lateral lying. The increase in prepartum nesting in sows with large litters may be a
consequence of stronger physiological signals in large litters than in small litters. This is
supported by Castren et al. (1993), who found negative correlations between litter size and
plasma concentrations of somatostatin during the last 3 days prepartum as well as between
plasma somatostatin and the time spent carrying straw on day �2 in relation to farrowing
(i.e. the period from 48 to 24 h before birth of the first piglet). The decreased inter-birth
interval may also be explained by increased physiological signals from the unborn piglets.
The birth of a piglet stimulates a pulsatile release of oxytocin in response to the mechanical
stimulation of the birth canal (Gilbert et al., 1994). Oxytocin release in response to the new
born piglets’ stimulation of the mammary glands may also be seen (Castren, 1993). Such
mechanisms may contribute to the decreased inter-birth intervals but do not explain the
increased variability in the inter-birth interval also seen in larger litters. Thus, a more
complex relationship between litter size, physiological signals and birth progress may exist
that on one hand speeds up the birth process but one the other hand also induces more
variability. The greater stimulation from large litters of the mammary glands may motivate
the sows for nursing, thus explaining the prolonged postpartum lateral lying.
The final graphical model (Fig. 2) of the present study showed that the four different
causes of death separated into three categories that each were related to different
behavioural variables. The three groups were unrelated to each other indicating that
different causal relationships may be involved in each cause of death. An exception,
however, was stillbirth and other causes, that both were related to behavioural variables
describing early suckling activity by the piglets.
Crushing (with or without lack of colostrums) was the most common cause of death
among the liveborn piglets. Crushing was related to lateral lying before parturition. Sows
that were in lateral recumbency (without standing or sitting) in most 15 min periods during
the last 4 h before parturition started, crushed more piglets after farrowing. In fact, sows
with low crushing only had few 15 min periods with constant lateral lying during the last
4 h before birth. The most obvious explanation is that extensive lateral lying is caused by
illness, bad leg constitution or obesity in the sow. These conditions may make movement
less frequent, but also make the few movements that occur highly dangerous for the piglets,
either because the sow is less motivated for carefulness when changing posture, or because
her ability to control the movements is reduced (see Damm et al., 2005b). Illness may also
cause lack of milk making the piglets less viable and thereby more at the risk of being
crushed. However, several aspects of the study design and results contradict this. Firstly,
sows with poor leg constitution and extreme body conditions were excluded from the study.
Secondly, there was no association between prepartum and partal–postpartal lying. This
would have been expected, as the mentioned conditions (leg problems and extreme body
condition) are not relieved simply through giving birth. Finally, there was no association
between prepartum lateral lying and body temperature, which would also have been
expected if lateral lying was caused by illness. A more likely explanation may therefore be
that prepartum lateral lying reflects general passivity. The passivity was related to low
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 227

nesting activity in the present study and may also be related to low postpartum motivation
for behavioural patterns that make postural changes safer for the piglets, e.g. pre-lying
behaviour (Clough and Baxter, 1984; Marchant et al., 2001) and responsiveness to screams
of piglets trapped underneath the sow’s body (Wechsler and Hegglin, 1997; Janczak et al.,
2003). Therefore, in solving the problem of piglet crushing it might be advantageous to
focus on stimulating sows to change posture safely rather than on mere reduction of the
occurrence of postural changes. One way may be through the provision of nesting materials
and the succeeding feed-back from a completed nest (Herskin et al., 1998, 1999; Pedersen
et al., 2003).
It has previously been shown that piglets are crushed when the sow rolls or lies down
from standing (Weary et al., 1996, 1998; Marchant et al., 2001). This would suggest that
much partal–postpartal lateral lying would be beneficial for piglet survival (Jarvis et al.,
1999; Pedersen et al., 2003) due to reduced number of postural changes. However, in the
present study no such link was found between crushing and partal–postpartal lateral lying,
which support our previous suggestion of focusing on making postural changes safer rather
than on reducing their frequency. Neither pre-partal nor partal–postpartal nesting activity
was related to partal–postpartal lateral lying, whereas partal–postpartal lateral lying was
related to the piglets’ suckling activity. Thus, the results suggests that partal–postpartal
lateral lying is mainly a reflection of piglets suckling activity or vice verse and not related
to pre-partal passivity.
In the present study, a great proportion of the crushed piglets were dead without
ingestion of colostrum and similar results have been found in previous studies (see
Edwards (2002) for review). Furthermore, Weary et al. (1996) has shown that starving
piglets are more at risk of being crushed. This suggests a link between crushing and piglet
suckling activities. The lack of relationship between crushing and measures of piglet
viability in the present data, may be explained by the fact that death due to other causes also
involved piglets culled by the stock person due to pure growth. Thus, some of the piglets
that suffered from starvation and therefore were in risk of being crushed may have been
culled by the stock person and therefore not crushed later by the sow. This may have
weakened the expected link between piglet viability and crushing and strengthened the link
between piglet viability and death due to other causes.
Low piglet activities were solely related to litters with much Stillbirth and with many
deaths due to other causes. In turn, stillbirth was related to variation in the inter-birth
interval, which suggests that liveborn piglets in litters suffering from much stillbirth were
subjected to hypoxia during birth due to the occurrence of some long birth intervals (Fraser
et al., 1997). Thus, birth problems may render liveborn piglets less viable after birth and
more prone to death due to other causes (Herpin et al., 1996; Leenhouwers et al., 1999).
Therefore, the consequences of problematic births could be viewed as a continuum with
stillbirth as the most extreme outcome and varying degrees of reduced viability and vitality
as less extreme outcome. A high occurrence of stillbirth might therefore be used as an
indicator of litters with particular risk of death amongst the liveborn and hence of litters
requiring extra attention and care.
Death due to lack of colostrum ingestion was more common when parturition started in
the morning than when it started in the evening. There was on the contrary, no link to
suckling activities, suggesting that piglets dying from lack of colostrum in general come
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232228

from healthy viable litters. Thus, the probability that management was involved rather than
piglet viability seems therefore likely. Certain management procedures may be carried out
sooner in relation to birth when parturition starts in the morning rather than in the evening
or nights. Piglets may be cross-fostered sooner, possibly before having obtained adequate
amounts of colostrum. Piglets may also be isolated in the piglet creep area during the first
morning feeding with the intention of providing them with heat and securing that they are
not crushed, but at the same time preventing their colostrum intake. Such morning
procedures were unsystematically observed in all herds during the video analysis of the
present study. No studies have investigated the effects and timing of such procedures on
piglet survival, and controlled experimental studies relating the procedures to piglet
mortality needs to be carried out.
Sows with a high percentage of piglets dying due to lack of colostrum ingestion also had
higher rectal temperature days 1 and 2 after parturition. These sows may have suffered
from the agalactia that is part of the MMA (Mastitis–Metritis–Agalactia) syndrome
(Furniss, 1987). Rectal temperature on the day of parturition was not associated with
starvation of the piglets, but with nesting before parturition. Temperature at this time
probably reflects the physical strain of nesting and giving birth, and not yet any developing
disease.
The approach of using chain-graph models to analyse multivariate data in an
exploratory phase, to identify potential causal mechanisms that may later be investigated in
control experimental designs, turned out to be valuable. The major advantage over the more
commonly used factor analysis, was that the chain-graph models allowed estimation of
quantitative relationships between original variables that would not have been possible
using factor analysis. Finally, it should be mentioned that the present analysis only
investigated the association between variables at the sow/litter level of the hierarchy.
Similar analysis could have been performed on the herd and batch level. For example, the
estimates of the batch values from the random component of each of the linear models,
could have been analysed using the MIM-program, to identify the structure of the
covariance in batch effects. However, to obtain consistent estimates, this would require a
larger dataset than the present one.
5. Conclusion
In conclusion, the analysis showed that different causes of mortality were linked to
different behaviour variables during the periparturient period and that they grouped into
three categories. The first category was associated with stillbirth, which was linked to the
progress of parturition and to piglet viability/suckling activity. The second category was
associated with piglet crushing, which was linked to lateral lying and nesting activity
before parturition. And finally, the third category was associated with starvation, which
was linked to the time of parturition and sow rectal temperature on days 1 and 2 after
farrowing. The results further showed that litter size not only influenced mortality but also
most behavioural variables, even those occurring before parturition.
The study points towards several areas of interest for further research that can help
understand early piglet mortality. These include causes of variation in the progress of
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 229

parturition, causes of individual differences in sow activity and responsiveness to piglets,
and early management of farrowing.
In addition, the study suggests possible indicators of problematic farrowings rendering
liveborn piglets at risk. These include prepartum lateral lying in the sow, suckling
behaviour and rate of stillbirth.
References
Bates, D., Sarkar, D., 2004. lme4: Linear mixed-effects models using S4 classes. R package version 0.6-9.
Bille, N., Nielsen, N.C., Larsen, J.L., Svendsen, J., 1974. Preweaning mortality in pigs. 2. The perinatal period.
Nord. Vet. Med. 26, 294–313.
Blauth, A., Caputo, A., Didelez, V., Pigeot, I., 2000. GraphFitI—manual, discussion paper 225. Statistische
Analyse Diskreter Strukturen.
Castren, H., Algers, B., de Passille, A.-M., Rushen, J., Uvnæs-Moberg, K., 1993. Periparturient variation in
progesterone, prolactin, oxytocin and somatostatin in relation to nest building in sows. Appl. Anim. Behav.
Sci. 38, 91–102.
Castren, H., 1993. Suckling behaviour, milk consumption and hormone release in the sow relative to nest building
and early milk ejections. Ph.D. thesis, College of Veterinary Medicine, Helsinki, Finland.
Clough, C., Baxter, M., 1984. Has the crate had its day? Pig Farm. 32, 49.
Damm, B.I., Forkman, B., Pedersen, L.J., 2005a. Lying down and rolling behaviour in sows in relation to piglet
crushing: a review. Appl. Anim. Behav. Sci. 90, 3–20.
Damm, B.I., Pedersen, L.J., Heiskanen, T., Nielsen, N.P., 2005b. The effects of straw on periparturient sow
behaviour, and piglet mortality and growth in modified schmid pens in a commercial breeding unit. Appl.
Anim. Behav. Sci. 92, 45–60.
Edwards, D., 2000. Introduction to Graphical Modelling, second ed. Springer–Verlag, New York Inc..
Edwards, S.A., 2002. Perinatal mortality in the pig: environmental or physiological solutions? Livest. Prod. Sci.
78 (1), 3–12. URL:http://www.sciencedirect.com/science/article/B6T9B-475BB8C-5/2/4bf1439a5e585a
1fd1b08ed372a58765.
English, P.R., Smith, W.J., 1975. Some causes of death in neonatal piglets. Vet. Ann. 15, 95–104.
Fraser, D., Phillips, P.A., Thompson, B.K., 1997. Farrowing behaviour and stillbirth in two environments: an
evaluation of the restraint-stillbirth hypothesis. Appl. Anim. Behav. Sci. 55, 51–66.
Fraser, D., 1990. Crushing of piglets by sows: effects of litter features, pen features and sow behaviour. Appl.
Anim. Behav. Sci. 61, 103–111.
Furniss, S.J., 1987. Measurement of rectal temperature to predict mastitis, metritis and agalactia (MMA) in sows
after farrowing. Prev. Vet. Med. 5 (2), 133–139.
Gilbert, C.L., Goode, J.A., McGrath, T.J., 1994. Pulsatile secretion of oxytocin during parturition in the pig:
temporal relationship with fetal expulsion. J. Physiol. 475, 129–137.
Grandinson, K., Lund, M.S., Rydhmer, L., Strandberg, E., 2002. Genetic parameters for the piglet mortality traits
crushing, stillbirth and total mortality, and their relation to birth weight. Acta Agric. Scand., Sect. A: Anim.
Sci. 52, 167–173.
Herpin, P., Le Dividich, J., Hulin, J.C., Fillaut, M., De Marco, F., Bertin, R., 1996. Effects of the level of asphyxia
during delivery on viability at birth and early postnatal vitality of newborn pigs. J. Anim. Sci. 74, 2067–
2075.
Herskin, M.S., Jensen, K.H., Thodberg, K., 1998. Influence of environmental stimuli on maternal behaviour
related to bonding, reactivity and crushing of piglets in domestic sows. Appl. Anim. Behav. Sci. 58, 241–
254.
Herskin, M.S., Jensen, K.H., Thodberg, K., 1999. Influence of environmental stimuli on nursing and suckling
behaviour in domestic sows and piglets. Anim. Sci. 68, 27–34.
Højsgaard, S., Thiesson, B., 1995. Bifrost—block recursive models induced from relevant knowledge, observa-
tions, and statistical techniques. J. Comput. Stat. Data Anal. 19, 155–175.
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232230

Højsgaard, S, 2003. mimR—a package for graphical modelling in R. In: Hornik, K., Leisch, F. (Eds.), Proceedings
of the Third International Workshop on Distributed Statistical Computing (DSC2003), 20–22 March. Vienna,
Austria. URL: http://www.ci.tuwien.ac.at/Conferences/DSC-2003.
Janczak, A.M., Pedersen, L.J., Rydhmer, L., Bakken, M., 2003. Relation between early fear- and anxiety-related
behaviour and maternal ability in sows. Appl. Anim. Behav. Sci. 82, 121–135.
Jarvis, S., McLean, K., Calvert, S.K., Deans, L.A., Chirnside, J., Lawrence, A.B., 1999. The responsiveness of
sows to their piglets in relation to the length of parturition and the involvement of endogenous opioids. Appl.
Anim. Behav. Sci. 63, 195–207.
Jultved, C.R., 2004. Rapport over P-rapporternes resultater April 2004. (Results from the P-reports, April 2004).
No. Notat. 0421. Landsudvalget for Svin, Dansk Landbrugsradgivning, Landscentret Svin. URL: http://
www.infosvin.dk/index.aspx?id=d2a18db5-b488-4802-ae4b-02971933fb0e.
Knol, E.F., Ducro, B.J., van Arendonk, J.A.M., van der Lende, T., 2002. Direct, maternal and nurse sow genetic
effects on farrowing-, pre-weaning- and total piglet survival. Livest. Prod. Sci. 73, 153–164.
Kongsted, A.G., Larsen, V. Aa., 1999. Pattegrisedødelighed i frilandssohold. (Piglet mortality in outdoor sows).
No. DJF-rapport nr. 11. Ministeriet for Fødevarer, Landbrug og Fiskeri, Danmarks Jordbrugsforskning, P.O.
Box 50, 8830 Tjele.
Lauritzen, S.L., Wermuth, N., 1989. Graphical models for associations between variables, some of which are
qualitative and some quantitative. Ann. Stat. 17, 31–57. URL:http://links.jstor.org/sici?sici=0090-
5364%28198903%2917%3A1%3C31%3AGMFABV%3E2.0.CO%3B2-O.
Lauritzen, S.L., Richardson, T.S., 2002. Chain graph models and their causal interpretations. J. R. Stat. Soc. B
64(3), 321–348. URL: http://www.blackwell-synergy.com/links/doi/10.1111/1467-9868.00340/abs.
Leenhouwers, J.I., van der Lende, T., Knol, E.F., 1999. Analysis of stillbirth in different lines of pig. Livest. Prod.
Sci. 57, 243–253.
Marchant, J.N., Broom, D.M., Corning, S., 2001. The influence of sow behaviour on piglet mortality due to
crushing in an open farrowing system. Anim. Sci. 72, 19–28.
Marchant, J.N., Rudd, A.R., Mendl, M.T., Broom, D.M., Meredith, M.J., Corning, S., Simmens, P.H., 2000. The
timing and causes of piglet mortality in alternative and conventional farrowing systems. Vet. Rec. 147, 209–
214.
Martin, P., Bateson, P., 1986. Measuring Behaviour—An Introductory Guide.. Cambridge University Press,
Cambridge, UK.
Mohamed, W.N., Diamond, I., Smith, P.W.F., 1998. The determinants of infant mortality in Malaysia: a graphical
chain modelling approach. J. R. Stat. Soc. B 161, 349–366.
Pedersen, L.J., Damm, B.I., Marchant-Forde, J.N., Jensen, K.H., 2003. Effects of feed-back from the nest on
maternal responsiveness and postural changes in primiparous sows during the first 24 hours after farrowing
onset. Appl. Anim. Behav. Sci. 83 (2), 109–124.
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D, 2004. nlme: Linear and nonlinear mixed effects models. R package
version 3.1-53.
Roehe, R., Kalm, E., 2000. Estimation of genetic and environmental risk factors associated with pre-weaning
mortality in piglets using generalized linear mixed models. Anim. Sci. 70, 227–240.
R Development Core Team, 2004. R: A language and environment for statistical computing. R Foundation for
Statistical Computing, Vienna, Austria, ISBN 3-900051-07-0. URL: http://www.R-project.org.
Schmid, H., 1991. Naturliche Verhaltenssicherungen der Hausschweine (Sus Scrofa) gegen das Erdrucken der
Ferkel durch die Muttersau und die Auswirkun-gen haltungsbedingter Storungen. Ph.D. thesis. Philoso-
phischen Fakultat II, Universitat Zurich, Schwitzerland.
Schmid, H., 1993. Influence of domestication and housing conditions on the behaviour of lying down in sows. In:
Proceeding of the International Congress on Applied Ethology, K.T.B.L. Darmstadt, Germany.
Su, G., Lund, M.S., Sorensen, D., 2004. Variance components for litter size and survival in Danish landrace and
Yorkshire pigs. In: 55th EAAP, vol. G4.7, Bled, Slovenia, 5th–9th September 2004.
Tuchscherer, M., Puppe, B., Tuchscherer, A., Tieman, U., 2000. Identification of neonates at risk: traits of newborn
piglets with respect to survival. Theriogenology 54, 371–388.
Vaillancourt, J., Tubbs, R.C., 1992. Preweaning mortality. Swine Reprod. 8 (3), 685–706.
Weary, D.M., Pajor, E.A., Thompson, B.K., Fraser, D., 1996. Sow body movements that crush piglets: a
comparison between two types of farrowing accommodation. Appl. Anim. Behav. Sci. 49, 149–158.
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232 231

Weary, D.M., Phillips, P.A., Pajor, E.A., Fraser, D., Thompson, B.K., 1998. Crushing of piglets by sows: effects of
litter features, pen features and sow behaviour. Appl. Anim. Behav. Sci. 61, 103–111.
Wechsler, B., Hegglin, D., 1997. Individual differences in the behaviour of sows at the nest-site and the crushing of
piglets. Appl. Anim. Behav. Sci. 51, 39–49.
Wermuth, N., 2003. Analysing social science data with graphical Markov models. In: Green, P., Hjort, N., Ri-
chardson, S. (Eds.), Highly Structured Stochastic Systems. University Press, Oxford, pp. 47–52.
L.J. Pedersen et al. / Applied Animal Behaviour Science 96 (2006) 215–232232





























