Embed Size (px)
Citation preview

www.elsevier.com/locate/trim
Transplant Immunology 14
EBV-specific memory CD8+ T cell phenotype and function in stable solid
organ transplant patients
Camila Macedoa,b, Albert Donnenbergb,c, Iulia Popescua,b, Jorge Reyesa,b,
Kareem Abu-Elmagda,b, Ron Shapiroa,b, Adriana Zeevia,d, John J. Funga,b,
Walter J. Storkusc,e,f, Diana Metesa,b,c,f,*
aThomas E. Starzl Transplantation Institute, University of Pittsburgh, Pittsburgh, PA, United StatesbDepartment of Surgery, University of Pittsburgh, Pittsburgh, PA, United States
cUniversity of Pittsburgh Cancer Institute, University of Pittsburgh, Pittsburgh, PA, United StatesdDepartment of Pathology, University of Pittsburgh, Pittsburgh, PA, United States
eDepartment of Dermatology, University of Pittsburgh, Pittsburgh, PA, United StatesfDepartment of Immunology, University of Pittsburgh, Pittsburgh, PA, United States
Received 14 January 2005; received in revised form 9 February 2005; accepted 10 February 2005
Abstract
Immune responses to EBV in immunosuppressed (IS) solid organ transplant (SOTx) recipients have not been well characterized. Here we
evaluate the phenotype and function of EBV-specific CD8+ T cells in peripheral blood isolated from ‘‘stable’’ IS SOTx recipients. The EBV-
specific CD8+ T cell memory subset distribution in the peripheral blood of patients was examined by flow cytometric analysis using HLA-A2
tetramers incorporating BMLF1 (lytic), and LMP2 and EBNA3A (latent)-derived peptides, in conjunction with mAbs against the CD45RO,
CD45RA, and CD62L markers. The ability of CD8+ T cells to produce IFN-g in response to the same EBV-derived peptides was measured by
ELISPOT assay. Patients and healthy normal donors exhibited similar anti-EBV CD8+ T cell frequencies and specificities against the EBV
epitopes evaluated. When compared to healthy normal donors, an overall significant expansion of the CD8+ T cell ‘‘effector memory’’
(CD45RO+/CD62L�) pool, including that of EBV ‘‘latent’’ (LMP2 and EBNA3A)-specific CD8+ T cells was detected in IS SOTx patients.
However, the patients’ EBV-specific CD8+ T cells showed decreased IFN-g production to the EBV-peptide stimulation. These results indicate
that the impairment of EBV-specific CD8+ T cell activity is not due to clonal depletion, but is mainly due to impaired functional activation.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Solid organ transplantation; EBV; Memory CD8+ T cells; IFN-g
1. Introduction
Epstein Barr Virus (EBV) is a ubiquitous human g-
herpes virus with tropism for B lymphocytes [1,2]. More
than 90% of the adult world’s population is infected with
EBV, and carry the virus life-long, as an asymptomatic
0966-3274/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.trim.2005.02.001
* Corresponding author. Section of Cellular Transplantation, Thomas E.
Starzl Transplantation Institute, E1551 Biomedical Science Tower, 200
Lothrop Street, Pittsburgh, PA 15261, United States. Tel.: +1 412 648 3291;
fax: +1 412 624 6666.
E-mail address: [email protected] (D. Metes).
infection. EBV replication and persistence are maintained
through a fine balance of productive (‘‘lytic’’) and non-
productive (‘‘latent’’) infections [3,4]. In healthy individ-
uals, EBV-specific CD8+ T cells play a pivotal role in
controlling viral replication and latency establishment
during the primary infection, and throughout the life-long
carrier state, preventing EBV-associated diseases [5,6].
Since chronic administration of immunosuppressive (IS)
drugs to prevent graft rejection impairs cellular immune
surveillance, EBV is an important source of morbi-
mortality in solid organ transplant (SOTx) recipients,
rendering them susceptible to post-transplant lymphoproli-
ferative disorders (PTLD) [7]. These are typically B cell
(2005) 109 – 116

Table 1
Patient information
Patientsa,b (n =15) Regimenc
FK506 (mg/day) Prednisone (mg/day)
Liver (n =4) 2–6 5
Small Bowel (n =5) 2–12 5–15
Multivisceral (n =6) 4–8 5
Controlsb (n =9) N/A N/A
a Stable SOTx patients=patients with no recent rejection episodes or
EBV re-activations.b The age/gender distribution of subjects was as follows: Controls – 4F/
2M, median age 45; Patients – 4F/6M, median age 49.c All patients were on two-drug maintenance therapy with the exception
of two multivisceral recipients who were on FK506 only.
C. Macedo et al. / Transplant Immunology 14 (2005) 109–116110
malignancies, ranging from polyclonal proliferations to
aggressive monomorphic lymphomas. They display a
broad spectrum of latency genes, also referred to as
latency III phenotype [8]. Recent reports also imply that
EBV lytic replication might actively participate in estab-
lishment and pathogenesis of latency.
Memory CD8+ T cells can be characterized by distinct
phenotypic, migratory, and effector functions. ‘‘Central
memory’’ T cells are CD45RO+ cells that constitutively
express high levels of the L-selectin CD62L marker, and
lack immediate effector function. They home to secondary
lymphoid organs, where they proliferate and differentiate
into effectors upon proper antigenic re-challenge [9,10]. In
contrast, ‘‘effector memory’’ cells exhibit a down-regulation
in CD62L expression, are found mostly in the peripheral
inflamed tissues, and can readily secrete IFN-g, IL-4, and/or
perforin in response to antigen stimulation [9,10]. Some
CD8+ T cells in this functional compartment are CD45RA+,
and are known as ‘‘stable effector memory’’ cells, whereas
others are CD45RO+, and are known as ‘‘effector memory’’
cells. Thus, in humans, the ‘‘effector’’ memory pool
contains the bonafide functional Th1, Th2, and CTL cell
populations [9,10].
The repertoire and activation status of EBV-specific
CD8+ T cells have been well documented in EBV+ healthy
individuals [11–13]. Interestingly, recent studies on
healthy EBV chronic carriers have revealed profound
differences in the composition of EBV-specific CD8+ T
cell memory populations and their functionality, suggesting
that immune response to ‘‘lytic’’ vs. ‘‘latent’’-associated
antigens is independent [14]. EBV ‘‘lytic’’-epitope specific
CD8+ T cells are CD45RO+ or CD45RA+ and CD62L+,
whereas EBV ‘‘latent’’-epitope specific CD8+ T cells
express mainly CD45RO+ and CD62L+ in normal subjects
[11,14].
To date, most studies regarding anti-EBV cellular
immunity in IS SOTx patients have focused on pediatric
transplant recipients with PTLD [15–17]. Little is known
about EBV-specific CD8+ T cell responses in the context of
impaired immunity seen with quiescent SOTx patients. Here
we have characterized the antigen specificities, phenotype,
and function of EBV-specific CD8+T cells in adult stable
transplant patients receiving two-drug maintenance therapy
(Prednisone and Tacrolimus). Our results provide a better
understanding of the anti-viral immune responses influenced
by this unique iatrogenic-biased virus–host relationship.
2. Materials and methods
2.1. Subjects
Fifteen stable IS SOTx patients and nine healthy
individuals (Table 1) were consented under IRB-approved
protocols. All subjects were EBV+ as determined by
serology. Ten stable patients and six healthy individuals
were HLA-A2+ as determined by molecular HLA-typing,
performed at the University of Pittsburgh Medical Center
(Table 1). At the time of the blood donation, all patients
were quiescent, without evidence of rejection or EBV
(re)activation. All patients had been receiving chronic
daily administration of Prednisone (5–15 mg/day) and
FK506 (2–12 mg/day) for more than 3 years. These
patients had never received Tymoglobulin or Campath1H
as preconditioning regimen or for rejection– rescue
intervention.
2.2. Cells
Ten milliliters of heparinized whole blood was collected
from each subject every 6 months over a period of 1 year.
Whole blood was used in flow cytometry analyses, while
peripheral blood mononuclear cells (PBMC) were isolated
by density gradient centrifugation for functional assays [18].
The TAP-deficient T2 cell line was pulsed with the relevant
peptides, and used as an antigen-presenting cell to stimulate
autologous PBMC in ELISPOT assays [19].
2.3. Peptides
The following EBV-derived peptides presented by HLA-
A2 were synthesized at the Peptide Synthesis Facility at
University of Pittsburgh: GLCTLVAML (BMLF1),
CLGGLLTMV (LMP2), and SVRDRLARL (EBNA3A)
[4,5]. These peptides were used in ELISPOT assays, as
well as for generation of streptavidin-PE conjugated HLA-
A2 tetramers at the NIAID MHC Tetramer Core Facility
(Emory University, Atlanta, GA).
2.4. Flow cytometry
Whole blood (200 Al) was incubated with a given
tetramer and a mixture of 3 mAbs for 30 min in the dark
at room temperature [19]. Surface staining markers used
included: anti-CD8 Cy-Chrome and anti-CD45RO FITC
(BD Bioscience Pharmingen, San Jose, CA); anti-CD62L
FITC, and anti-CD45RA ECD (Immunotech, Miami, FL).
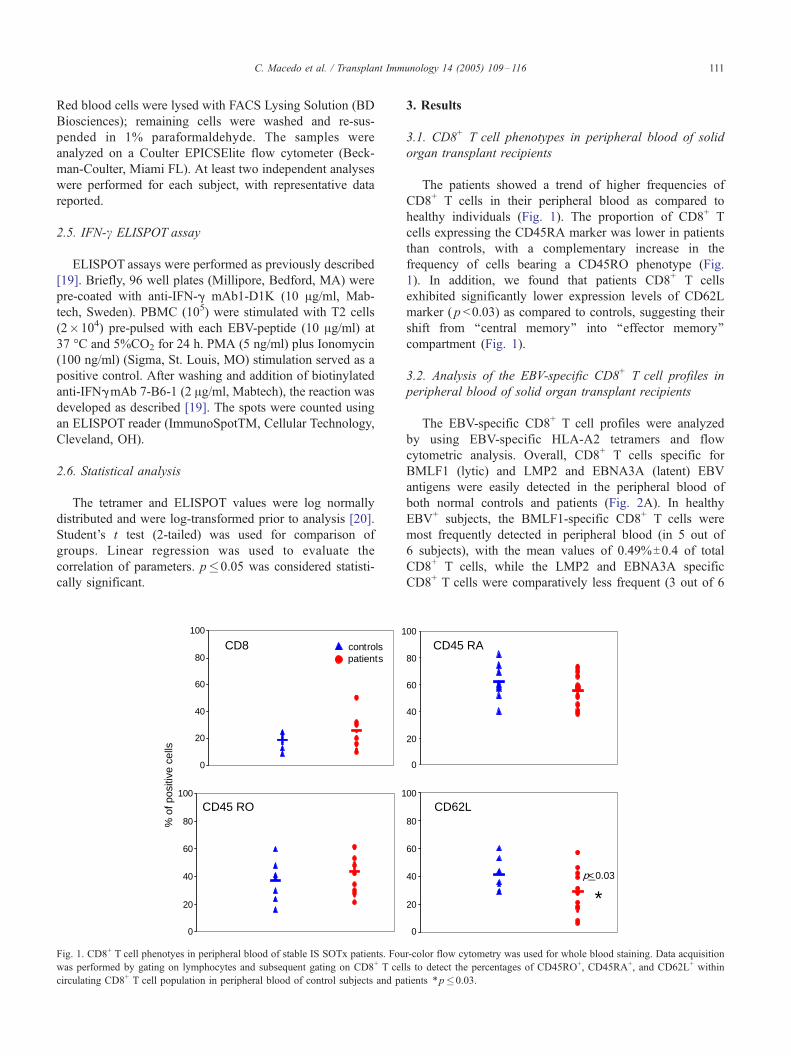
C. Macedo et al. / Transplant Immunology 14 (2005) 109–116 111
Red blood cells were lysed with FACS Lysing Solution (BD
Biosciences); remaining cells were washed and re-sus-
pended in 1% paraformaldehyde. The samples were
analyzed on a Coulter EPICSElite flow cytometer (Beck-
man-Coulter, Miami FL). At least two independent analyses
were performed for each subject, with representative data
reported.
2.5. IFN-c ELISPOT assay
ELISPOT assays were performed as previously described
[19]. Briefly, 96 well plates (Millipore, Bedford, MA) were
pre-coated with anti-IFN-g mAb1-D1K (10 Ag/ml, Mab-
tech, Sweden). PBMC (105) were stimulated with T2 cells
(2�104) pre-pulsed with each EBV-peptide (10 Ag/ml) at
37 -C and 5%CO2 for 24 h. PMA (5 ng/ml) plus Ionomycin
(100 ng/ml) (Sigma, St. Louis, MO) stimulation served as a
positive control. After washing and addition of biotinylated
anti-IFNgmAb 7-B6-1 (2 Ag/ml, Mabtech), the reaction was
developed as described [19]. The spots were counted using
an ELISPOT reader (ImmunoSpotTM, Cellular Technology,
Cleveland, OH).
2.6. Statistical analysis
The tetramer and ELISPOT values were log normally
distributed and were log-transformed prior to analysis [20].
Student’s t test (2-tailed) was used for comparison of
groups. Linear regression was used to evaluate the
correlation of parameters. p�0.05 was considered statisti-
cally significant.
0
20
40
60
80
100
0
20
40
60
80
100 1
1
% o
f pos
itive
cel
ls
CD8
CD45 RO
controlspatients
Fig. 1. CD8+ T cell phenotyes in peripheral blood of stable IS SOTx patients. Fou
was performed by gating on lymphocytes and subsequent gating on CD8+ T cel
circulating CD8+ T cell population in peripheral blood of control subjects and pa
3. Results
3.1. CD8+ T cell phenotypes in peripheral blood of solid
organ transplant recipients
The patients showed a trend of higher frequencies of
CD8+ T cells in their peripheral blood as compared to
healthy individuals (Fig. 1). The proportion of CD8+ T
cells expressing the CD45RA marker was lower in patients
than controls, with a complementary increase in the
frequency of cells bearing a CD45RO phenotype (Fig.
1). In addition, we found that patients CD8+ T cells
exhibited significantly lower expression levels of CD62L
marker ( p <0.03) as compared to controls, suggesting their
shift from ‘‘central memory’’ into ‘‘effector memory’’
compartment (Fig. 1).
3.2. Analysis of the EBV-specific CD8+ T cell profiles in
peripheral blood of solid organ transplant recipients
The EBV-specific CD8+ T cell profiles were analyzed
by using EBV-specific HLA-A2 tetramers and flow
cytometric analysis. Overall, CD8+ T cells specific for
BMLF1 (lytic) and LMP2 and EBNA3A (latent) EBV
antigens were easily detected in the peripheral blood of
both normal controls and patients (Fig. 2A). In healthy
EBV+ subjects, the BMLF1-specific CD8+ T cells were
most frequently detected in peripheral blood (in 5 out of
6 subjects), with the mean values of 0.49%T0.4 of total
CD8+ T cells, while the LMP2 and EBNA3A specific
CD8+ T cells were comparatively less frequent (3 out of 6
0
20
40
60
80
00
0
20
40
60
80
00
CD45 RA
CD62L
*p< 0.03
r-color flow cytometry was used for whole blood staining. Data acquisition
ls to detect the percentages of CD45RO+, CD45RA+, and CD62L+ within
tients *p�0.03.
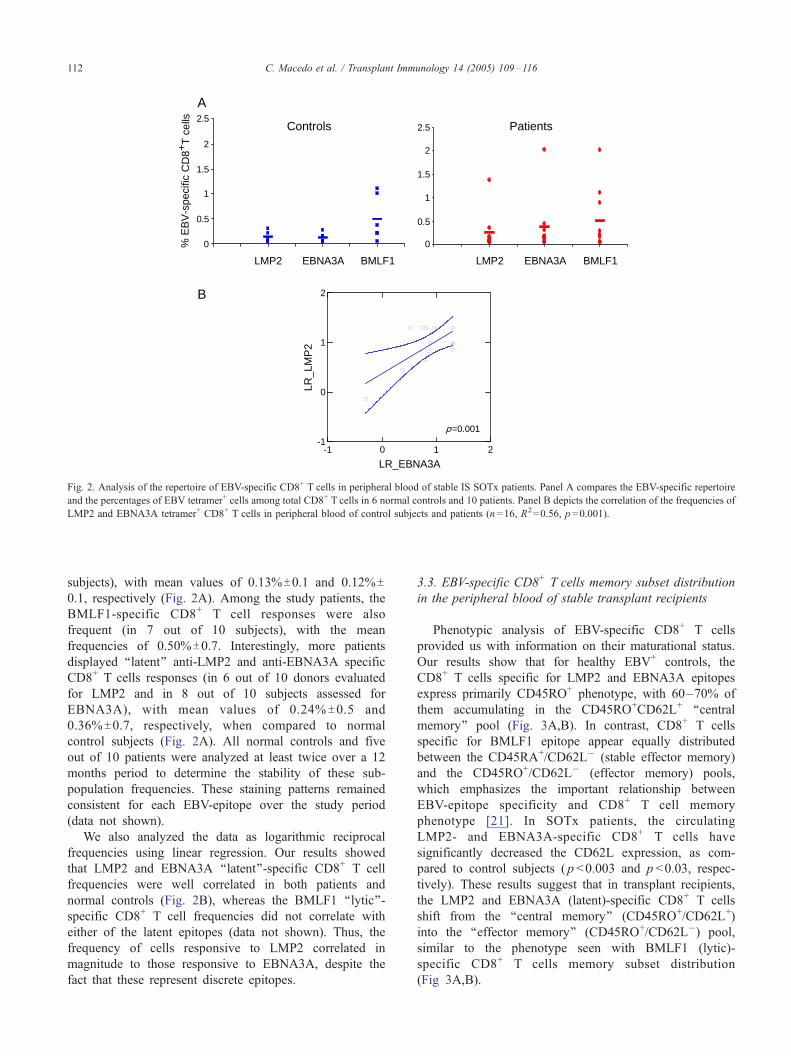
0
0.5
1
1.5
2
2.5
.5 1 1.5 2 2.50
0.5
1
1.5
2
2.5
0 0.5 1 1.5LMP2 EBNA3A BMLF1 LMP2 EBNA3A BMLF1
Controls Patients
% E
BV
-spe
cific
CD
8+T
cel
ls
A
-1 0 1 2
LR_EBNA3A
-1
0
1
2
LR_L
MP
2B
p=0.001
Fig. 2. Analysis of the repertoire of EBV-specific CD8+ T cells in peripheral blood of stable IS SOTx patients. Panel A compares the EBV-specific repertoire
and the percentages of EBV tetramer+ cells among total CD8+ T cells in 6 normal controls and 10 patients. Panel B depicts the correlation of the frequencies of
LMP2 and EBNA3A tetramer+ CD8+ T cells in peripheral blood of control subjects and patients (n =16, R2=0.56, p =0.001).
C. Macedo et al. / Transplant Immunology 14 (2005) 109–116112
subjects), with mean values of 0.13%T0.1 and 0.12%T0.1, respectively (Fig. 2A). Among the study patients, the
BMLF1-specific CD8+ T cell responses were also
frequent (in 7 out of 10 subjects), with the mean
frequencies of 0.50%T0.7. Interestingly, more patients
displayed ‘‘latent’’ anti-LMP2 and anti-EBNA3A specific
CD8+ T cells responses (in 6 out of 10 donors evaluated
for LMP2 and in 8 out of 10 subjects assessed for
EBNA3A), with mean values of 0.24% T0.5 and
0.36%T0.7, respectively, when compared to normal
control subjects (Fig. 2A). All normal controls and five
out of 10 patients were analyzed at least twice over a 12
months period to determine the stability of these sub-
population frequencies. These staining patterns remained
consistent for each EBV-epitope over the study period
(data not shown).
We also analyzed the data as logarithmic reciprocal
frequencies using linear regression. Our results showed
that LMP2 and EBNA3A ‘‘latent’’-specific CD8+ T cell
frequencies were well correlated in both patients and
normal controls (Fig. 2B), whereas the BMLF1 ‘‘lytic’’-
specific CD8+ T cell frequencies did not correlate with
either of the latent epitopes (data not shown). Thus, the
frequency of cells responsive to LMP2 correlated in
magnitude to those responsive to EBNA3A, despite the
fact that these represent discrete epitopes.
3.3. EBV-specific CD8+ T cells memory subset distribution
in the peripheral blood of stable transplant recipients
Phenotypic analysis of EBV-specific CD8+ T cells
provided us with information on their maturational status.
Our results show that for healthy EBV+ controls, the
CD8+ T cells specific for LMP2 and EBNA3A epitopes
express primarily CD45RO+ phenotype, with 60–70% of
them accumulating in the CD45RO+CD62L+ ‘‘central
memory’’ pool (Fig. 3A,B). In contrast, CD8+ T cells
specific for BMLF1 epitope appear equally distributed
between the CD45RA+/CD62L� (stable effector memory)
and the CD45RO+/CD62L� (effector memory) pools,
which emphasizes the important relationship between
EBV-epitope specificity and CD8+ T cell memory
phenotype [21]. In SOTx patients, the circulating
LMP2- and EBNA3A-specific CD8+ T cells have
significantly decreased the CD62L expression, as com-
pared to control subjects ( p <0.003 and p <0.03, respec-
tively). These results suggest that in transplant recipients,
the LMP2 and EBNA3A (latent)-specific CD8+ T cells
shift from the ‘‘central memory’’ (CD45RO+/CD62L+)
into the ‘‘effector memory’’ (CD45RO+/CD62L�) pool,
similar to the phenotype seen with BMLF1 (lytic)-
specific CD8+ T cells memory subset distribution
(Fig 3A,B).
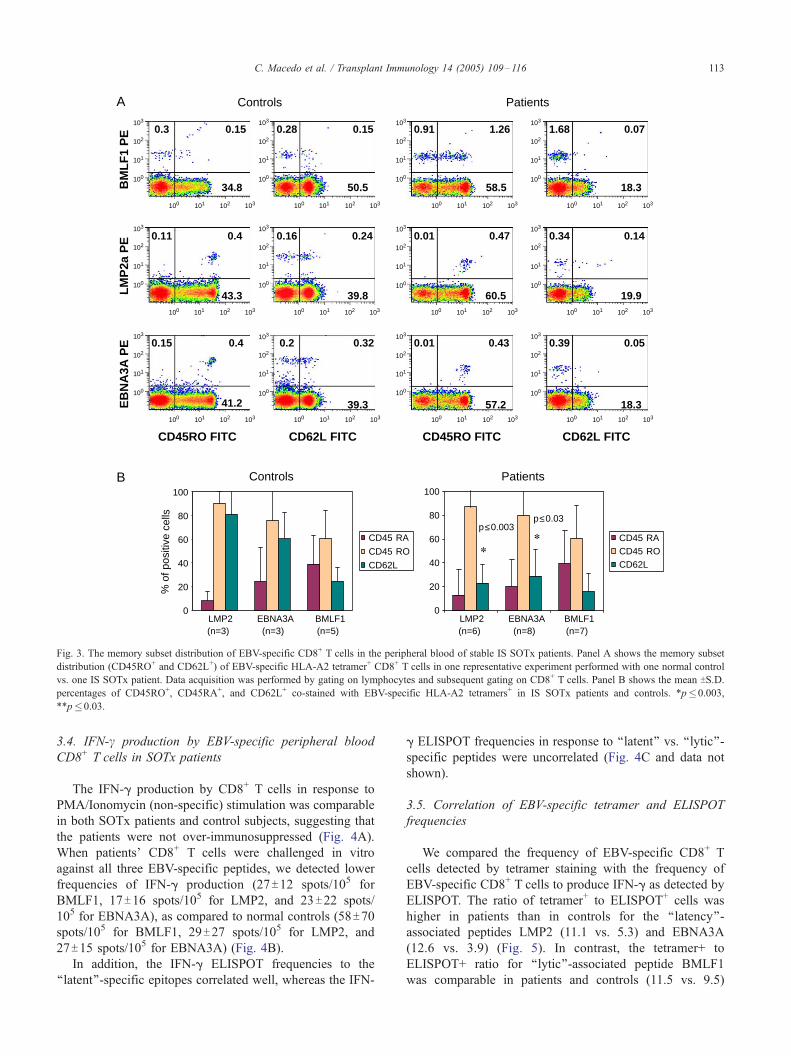
A
0.01 0.47
60.5
0.34 0.14
19.9
0.01 0.43
57.2
0.39 0.05
18.3
0.91 1.26
58.5
1.68 0.07
18.3
0.11 0.4
43.3
0.16 0.24
39.8
0.15 0.4
41.2
0.2 0.32
39.3
0.3 0.15
34.8
0.28 0.15
50.5
LM
P2a
PE
BM
LF
1 P
EE
BN
A3A
PE
CD45RO FITC CD62L FITC CD45RO FITC CD62L FITC
PatientsControls
Controls Patients
0
20
40
60
80
100
CD45 RA
CD45 RO
CD62L
0
20
40
60
80
100
CD45 RACD45 ROCD62L
% o
f pos
itive
cel
ls
LMP2 (n=3)
EBNA3A (n=3)
BMLF1 (n=5)
LMP2 (n=6)
EBNA3A (n=8)
BMLF1 (n=7)
p≤0.03p≤0.003
**
B
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
103
102
101
100
103102101100
Fig. 3. The memory subset distribution of EBV-specific CD8+ T cells in the peripheral blood of stable IS SOTx patients. Panel A shows the memory subset
distribution (CD45RO+ and CD62L+) of EBV-specific HLA-A2 tetramer+ CD8+ T cells in one representative experiment performed with one normal control
vs. one IS SOTx patient. Data acquisition was performed by gating on lymphocytes and subsequent gating on CD8+ T cells. Panel B shows the mean TS.D.
percentages of CD45RO+, CD45RA+, and CD62L+ co-stained with EBV-specific HLA-A2 tetramers+ in IS SOTx patients and controls. *p�0.003,
**p�0.03.
C. Macedo et al. / Transplant Immunology 14 (2005) 109–116 113
3.4. IFN-c production by EBV-specific peripheral blood
CD8+ T cells in SOTx patients
The IFN-g production by CD8+ T cells in response to
PMA/Ionomycin (non-specific) stimulation was comparable
in both SOTx patients and control subjects, suggesting that
the patients were not over-immunosuppressed (Fig. 4A).
When patients’ CD8+ T cells were challenged in vitro
against all three EBV-specific peptides, we detected lower
frequencies of IFN-g production (27T12 spots/105 for
BMLF1, 17T16 spots/105 for LMP2, and 23T22 spots/
105 for EBNA3A), as compared to normal controls (58T70spots/105 for BMLF1, 29T27 spots/105 for LMP2, and
27T15 spots/105 for EBNA3A) (Fig. 4B).
In addition, the IFN-g ELISPOT frequencies to the
‘‘latent’’-specific epitopes correlated well, whereas the IFN-
g ELISPOT frequencies in response to ‘‘latent’’ vs. ‘‘lytic’’-
specific peptides were uncorrelated (Fig. 4C and data not
shown).
3.5. Correlation of EBV-specific tetramer and ELISPOT
frequencies
We compared the frequency of EBV-specific CD8+ T
cells detected by tetramer staining with the frequency of
EBV-specific CD8+ T cells to produce IFN-g as detected by
ELISPOT. The ratio of tetramer+ to ELISPOT+ cells was
higher in patients than in controls for the ‘‘latency’’-
associated peptides LMP2 (11.1 vs. 5.3) and EBNA3A
(12.6 vs. 3.9) (Fig. 5). In contrast, the tetramer+ to
ELISPOT+ ratio for ‘‘lytic’’-associated peptide BMLF1
was comparable in patients and controls (11.5 vs. 9.5)
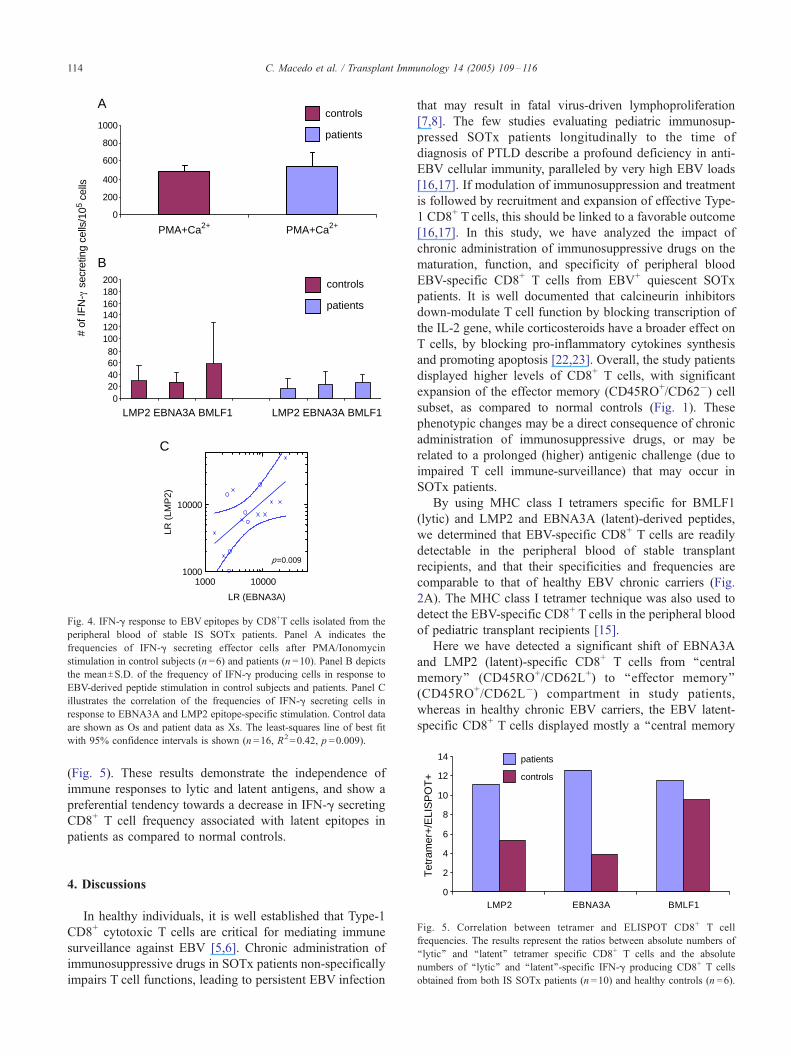
6
8
10
12
14
er+
/ELI
SP
OT
+ controls
patients
0
200
400
600
800
1000
1 2
A
B
020406080
100120140160180200
1 2 3 4LMP2 EBNA3A BMLF1 LMP2 EBNA3A BMLF1
PMA+Ca2+ PMA+Ca2+
# of
IFN
- s
ecre
ting
cells
/105
cells
controls
patients
controls
patients
1000 100001000
10000
LR (EBNA3A)
C
p=0.009
LR (
LMP
2)
Fig. 4. IFN-g response to EBV epitopes by CD8+T cells isolated from the
peripheral blood of stable IS SOTx patients. Panel A indicates the
frequencies of IFN-g secreting effector cells after PMA/Ionomycin
stimulation in control subjects (n =6) and patients (n =10). Panel B depicts
the meanTS.D. of the frequency of IFN-g producing cells in response to
EBV-derived peptide stimulation in control subjects and patients. Panel C
illustrates the correlation of the frequencies of IFN-g secreting cells in
response to EBNA3A and LMP2 epitope-specific stimulation. Control data
are shown as Os and patient data as Xs. The least-squares line of best fit
with 95% confidence intervals is shown (n =16, R2=0.42, p =0.009).
C. Macedo et al. / Transplant Immunology 14 (2005) 109–116114
(Fig. 5). These results demonstrate the independence of
immune responses to lytic and latent antigens, and show a
preferential tendency towards a decrease in IFN-g secreting
CD8+ T cell frequency associated with latent epitopes in
patients as compared to normal controls.
0
2
4
LMP2 EBNA3A BMLF1
Tet
ram
Fig. 5. Correlation between tetramer and ELISPOT CD8+ T cell
frequencies. The results represent the ratios between absolute numbers of
‘‘lytic’’ and ‘‘latent’’ tetramer specific CD8+ T cells and the absolute
numbers of ‘‘lytic’’ and ‘‘latent’’-specific IFN-g producing CD8+ T cells
obtained from both IS SOTx patients (n =10) and healthy controls (n =6).
4. Discussions
In healthy individuals, it is well established that Type-1
CD8+ cytotoxic T cells are critical for mediating immune
surveillance against EBV [5,6]. Chronic administration of
immunosuppressive drugs in SOTx patients non-specifically
impairs T cell functions, leading to persistent EBV infection
that may result in fatal virus-driven lymphoproliferation
[7,8]. The few studies evaluating pediatric immunosup-
pressed SOTx patients longitudinally to the time of
diagnosis of PTLD describe a profound deficiency in anti-
EBV cellular immunity, paralleled by very high EBV loads
[16,17]. If modulation of immunosuppression and treatment
is followed by recruitment and expansion of effective Type-
1 CD8+ T cells, this should be linked to a favorable outcome
[16,17]. In this study, we have analyzed the impact of
chronic administration of immunosuppressive drugs on the
maturation, function, and specificity of peripheral blood
EBV-specific CD8+ T cells from EBV+ quiescent SOTx
patients. It is well documented that calcineurin inhibitors
down-modulate T cell function by blocking transcription of
the IL-2 gene, while corticosteroids have a broader effect on
T cells, by blocking pro-inflammatory cytokines synthesis
and promoting apoptosis [22,23]. Overall, the study patients
displayed higher levels of CD8+ T cells, with significant
expansion of the effector memory (CD45RO+/CD62�) cell
subset, as compared to normal controls (Fig. 1). These
phenotypic changes may be a direct consequence of chronic
administration of immunosuppressive drugs, or may be
related to a prolonged (higher) antigenic challenge (due to
impaired T cell immune-surveillance) that may occur in
SOTx patients.
By using MHC class I tetramers specific for BMLF1
(lytic) and LMP2 and EBNA3A (latent)-derived peptides,
we determined that EBV-specific CD8+ T cells are readily
detectable in the peripheral blood of stable transplant
recipients, and that their specificities and frequencies are
comparable to that of healthy EBV chronic carriers (Fig.
2A). The MHC class I tetramer technique was also used to
detect the EBV-specific CD8+ T cells in the peripheral blood
of pediatric transplant recipients [15].
Here we have detected a significant shift of EBNA3A
and LMP2 (latent)-specific CD8+ T cells from ‘‘central
memory’’ (CD45RO+/CD62L+) to ‘‘effector memory’’
(CD45RO+/CD62L�) compartment in study patients,
whereas in healthy chronic EBV carriers, the EBV latent-
specific CD8+ T cells displayed mostly a ‘‘central memory

C. Macedo et al. / Transplant Immunology 14 (2005) 109–116 115
phenotype’’. The BMLF1 (lytic)-specific CD8+ T cells (Fig.
3A,B) in both patients and controls are mostly part of the
‘‘effector memory’’ subset [21]. When taken together, these
results support the idea that stable transplant recipients may
experience chronic higher endogenous EBV ‘‘latent’’-anti-
gen challenge, leading to ‘‘latent’’-specific clonal prolifer-
ation of potential ‘‘promptly’’ functional T cells (Fig. 3A,B).
Unfortunately, we were not able to quantify the viral load in
PBMC of the patients at the time of phenotypic analysis.
Previous reports suggest that transplant patients have an
increased number of circulating EBV-infected B cells in
their peripheral blood, and that these cells carry an elevated
EBV copy number per genome [24,25]. Babcock et al.
speculate that the increased EBV genome burden per B cell
is caused by a more frequent viral replication seen in
patients [25]. In stable SOTx patients, although circulating
EBV-infected B cells are resting memory B cells, they may
frequently convert into proliferating B cell blasts due to
impaired T cell immune-surveillance caused by persistent
immunosuppression. These B cell blasts that express latency
III phenotype, may therefore represent a constant source of
EBV ‘‘latent’’ antigens [25].
We next examined the frequency of IFN-g producing,
EBV-specific CD8+ T cells in the peripheral blood of stable
IS SOTx patients using the ELISPOT assay. Previous
reports, including our own published results, have deter-
mined that in healthy EBV+ controls approximately 10–
20% of circulating EBV-specific CD8+ T cells readily
respond to in vitro antigen-specific stimulation by produc-
ing IFN-g [19]. The frequency of IFN-g producing BMLF1-
specific CD8+ T cells was (on average) higher than the
frequencies of IFN-g producing EBNA3A and LMP2
specific CD8+ T cells (Fig. 4B). This is most likely due to
the higher frequency of BMLF1-specific CD8+ T cells,
which display an ‘‘effector memory’’ phenotype (CD45RO+/
CD62L� or CD45RA+/CD62L�) in peripheral blood
[11,19].
Our current results show that the frequency of CD8+ T
cells producing IFN-g in patients and controls was similar
when assessed by PMA and ionomycin stimulation,
supporting the notion that stable SOTx recipients were not
over-immunosuppressed. In contrast, the response to EBV-
specific stimulation induced lower frequencies of IFN-g
producing CD8+ T cells in patients, as compared with
healthy controls (Fig. 4B). This is intriguing, in view of an
increased proportion of ‘‘effector memory’’ cells among the
patients’ LMP2 and EBNA3A tetramer-specific CD8+ T
cells, as compared with that of healthy control donors.
Based on this phenotype, patients’ cells would be expected
to respond readily (i.e. IFN-g production) upon EBV-
specific stimulation [9,10]. We therefore speculate that the
antigen-specific functional (IFN-g) impairment of CD8+ T
cells in study patients may be related to the chronic
administration of IS drugs, and may be facilitated by the
development of antigen-specific suppressor/regulatory cells
[26]. The relationship between tetramer-specific and IFN-
gproducing antigen-specific CD8+ T cells in SOTx patients
has also been analyzed in the setting of alternate viral
infections [27]. Engstrand et al. showed that CMV-specific
CD8+ T cells are maintained in the peripheral blood of
SOTx patients, at levels comparable to those detected in
healthy subjects [27]. However, even though they express
an activated effector phenotype, they are functionally
impaired at the level of IFN-g production [27]. Of note, in
our study, the ratio of tetramer+ to ELISPOT+ cells further
revealed a significant decrease in IFN-g secretion specific
for the ‘‘latency’’ associated epitopes in stable patients (Fig.
5). These subtle alterations in the phenotype and function of
LMP2- and EBNA3A-specific memory CD8+ T cells in
patients may therefore, at least in part, explain the tendency
towards increased susceptibility of these individuals to
EBV-latency associated complications. Further studies are
needed to identify additional immune defects that may
render certain patients to be at greater risk than others to
develop EBV-related malignancies.
Acknowledgments
We thank Allison Logar for her outstanding assistance
with flow cytometric data analysis; Dr. Russell Salter
(University of Pittsburgh) for his gift of the T2 cell line.
This work was supported by grants from American Cancer
Society and American Heart Association. [DM]
References
[1] Rickinson A. Epstein-Barr virus. Virus Res 2002;82:109–13.
[2] Kuppers R. B cells under the influence: transformation of B cells by
Epstein-Barr Virus. Nat Rev Immunol 2003;3:801–12.
[3] Murray PG, Young LS. Epstein-Barr virus infection: basis of
malignancy and potential for therapy. Expert Rev Mol Med
2001:1–20.
[4] Thorley-Lawson DA. Epstein-Barr virus: exploiting the immune
system. Nat Rev Immunol 2001;1:75–82.
[5] Rickinson AB, Moss DJ. Human cytotoxic T lymphocyte responses to
Epstein Barr virus infection. Annu Rev Immunol 1997;15:405–13.
[6] Khanna R, Moss DJ, Burrows SR. Vaccine strategies against
Epstein-Barr virus-associated diseases: lessons from studies on
cytotoxic T-cell-mediated immune regulation. Immunol Rev 1999;
170:49–64.
[7] Nalesnik MA. Clinicopathologic characteristics of post-transplant
lymphoproliferative disorders. Recent Results Cancer Res
2002;159:9–18.
[8] Damania B. Oncogenic gamma-herpes viruses: comparison of viral
proteins involved in tumorigenesis. Nat Rev Microbiol 2004;8:
656–68.
[9] Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets
of memory T lymphocytes with distinct homing potentials and effector
functions. Nature 1999;401:708–12.
[10] Sallusto F, Lanzavecchia A. Exploring pathways of memory T cell
generation. J Clin Invest 2001;108:805–6.
[11] Hislop AD, Annels NE, Gudgeon NH, Leese AM, Rickinson AB.
Epitope-specific evolution of human CD8(+) T cell responses from
primary to persistent phases of Epstein-Barr virus infection. J Exp
Med 2002;195:893–905.

C. Macedo et al. / Transplant Immunology 14 (2005) 109–116116
[12] Catalina MD, Sullivan JL, Brody RM, Luzuriaga K. Phenotypic and
functional heterogeneity of EBV epitope-specific CD8+T cells. J
Immunol 2002;168:4184–91.
[13] Tussey L, Speller S, Gallimore A, Vessey R. Functionally distinct
CD8+ memory T cell subsets in persistent EBV infection are
differentiated by migratory receptor expression. Eur J Immunol
2000;30:1823–9.
[14] Faint JM, Annels NE, Curnow SJ, Shields P, Pilling D, Hislop AD,
et al. Memory T cells constitute a subset of the human
CD8+CD45RA+ pool with distinct phenotypic and migratory
characteristics. J Immunol 2001;167:212–20.
[15] Falco DA, Nepomuceno RR, Krams SM, Lee PP, Davis MM,
Salvatierra O, et al. Identification of Epstein-Barr virus-specific
CD8+ T lymphocytes in the circulation of pediatric transplant
recipients. Transplantation 2002;74:501–10.
[16] Smets F, Latinne D, Bazin H, Reding R, Otte JB, Buts JP, et al. Ratio
between Epstein-Barr viral load and anti-Epstein-Barr virus specific T-
cell response as a predictive marker of posttransplant lymphoprolifer-
ative disease. Transplantation 2002;73(10):1603–10.
[17] Baudouin V, Dehee A, Pedron-Grossetete B, Ansart-Pirenne H,
Haddad E, Maisin A, et al. Relationship between CD8+ T-cell
phenotype and function, Epstein-Barr virus load, and clinical outcome
in pediatric renal transplant recipients: a prospective study. Trans-
plantation 2004;77(11):1706–13.
[18] Boyum A, Lovhaug D, Tresland L, Nordlie EM. Separation of
leucocytes: improved cell purity by fine adjustments of gradient
medium density and osmolarity. Scand J Immunol 1991;34:697–712.
[19] Popescu I, Macedo C, Zeevi A, Nellis J, Patterson KR, Logar A, et al.
Ex vivo priming of naı̈ve T cells into EBV-specific Th1/Tc1 effector
cells by mature autologous DC loaded with apoptotic/necrotic LCL.
Am J Transplant 2003;3:1369–77.
[20] Hoffmann TK, Donnenberg VS, Friebe-Hoffmann U, Meyer EM,
Rinaldo CR, DeLeo AB, et al. Competition of peptide-MHC class I
tetrameric complexes with anti-CD3 provides evidence for specific-
ity of peptide binding to the TCR complex. Cytometry 2000;41:
321–8.
[21] Hislop AD, Gudgeon NH, Callan MF, Fazou C, Hasegawa H, Salmon
M, et al. EBV-specific CD8+ T cell memory: relationship between
specificity, cell phenotype, and immediate effector function. J Immunol
2001;167:2019–29.
[22] Siekierka JJ, Sigal NH. FK-506 and Cyclosporin A: immunosuppres-
sive mechanism of action and beyond. Curr Opin Immunol 1992;4:
548–52.
[23] Goulding NJ, Guyre PM. Glucocorticoids, lipocortins and the immune
response. Curr Opin Immunol 1993;5:108–13.
[24] Rose C, Green M, Webber S, Kingsley L, Day R, Watkins S, et al.
Detection of Epstein-Barr virus genomes in peripheral blood B cells
from solid-organ transplant recipients by fluorescence in situ hybrid-
ization. J Clin Microbiol 2002;40:2533–44.
[25] Babcock GJ, Decker LL, Freeman RB, Thorley-Lawson DA. Epstein-
Barr virus-infected resting memory B cells, not proliferating lympho-
blasts, accumulate in the peripheral blood of immunosuppressed
patients. J Exp Med 1999;190(4):567–76.
[26] Li Y, Koshiba T, Yoshizawa A, Yonekawa Y, Masuda K, Ito A, et al.
Analyses of peripheral blood mononuclear cells in operational
tolerance after pediatric living donor liver transplantation. Am J
Transplant 2004;4(12):2118–25.
[27] Engstrand M, Lidehall AK, Totterman TH, Herrman B, Eriksson BM,
Korsgren O. Cellular responses to cytomegalovirus in immunosup-
pressed patients: circulating CD8+ T cells recognizing CMVpp65 are
present but display functional impairment. Clin Exp Immunol 2003;
132(1):96–104.

![[Vaccinations for the traveling adult solid organ transplant recipient (excluding hematopoietic stem cell transplant recipients)]](https://img.pdfslide.net/doc/110x75/633690ebb5f91cb18a0be1e3/vaccinations-for-the-traveling-adult-solid-organ-transplant-recipient-excluding.jpg)



























