Embed Size (px)
Citation preview

Effect of Cell-Based Intercellular Delivery of TranscriptionFactor GATA4 on Ischemic Cardiomyopathy
Jing Bian, Zoran B. Popovic, Carlos Benejam, Matthew Kiedrowski,L. Leonardo Rodriguez, Marc S. Penn
Abstract—Recent loss-of-function studies highlight the importance of the transcription factor GATA4 in the myocardialresponse to injury in the adult heart. However, the potential effects of gain-in-function of GATA4 overexpression, andtranscription factors in general, is hindered by the fact that transcription factors are neither secreted nor taken up by cells.Chimeric proteins incorporating motifs of cell-penetrating proteins are secreted from cells and internalized bysurrounding cells. We engineered a chimeric protein consisting of GATA4 and the cell-penetrating protein VP22.Cardiac fibroblasts stably transfected with the GATA4:VP22, GFP:VP22, or green fluorescent protein (GFP) constructswere transplanted into Lewis rats 1 month after left anterior descending ligation. GATA4:VP22 expression in the borderzone was associated with increased cardiac myosin expression and cardiac myocyte size (30 �m versus 19 �m,P�0.01). Compared with the GFP:VP22 control group, 6 weeks after cardiac fibroblast transplantation (10 weeks aftermyocardial infarction), animals that received GATA4:VP22-expressing cardiac fibroblasts demonstrated increasedcardiac function and less negative remodeling. These data demonstrate a novel strategy for transcription factor deliveryto injured myocardium and indicate that the delivery of GATA4 locally to the infarct border zone induces multiple localeffects in the border zone cardiac myocytes resulting in beneficial ventricular remodeling and improved global leftventricular function. (Circ Res. 2007;100:1626-1633.)
Key Words: GATA4 � myocardial infarct � cell-based gene therapy � VP22 � intercellular delivery of protein
GATA4 is critical for the viability and hypertrophicresponse of cardiac myocytes following myocardial
injury.1,2 Consistent with these findings, GATA4 has beenshown to directly induce the expression of BCL2 in cardiacmyocytes in vitro.3 These findings suggest that the localoverexpression of GATA4 could lead to local cardiac hyper-trophy and improved cardiac myocyte survival. The goal ofthis study was to assess, for the first time, the effects of localGATA4 overexpression in the infarct border zone at a timeremote from myocardial infarction in a model of ischemiccardiomyopathy. To do so, because transcription factors areneither secreted nor internalized by surrounding cells, weused a novel method of local GATA4 delivery to the infarctborder zone.
We achieved local GATA4 delivery by combining cell-based gene therapy with a cell-penetrating protein (CPP).Cell-based gene therapy has been shown to be an effectivestrategy for stimulating angiogenesis and improving heartfunction.4–6 Moreover CPPs, by serving as vectors for thetransmembrane intercellular delivery of fused proteins, haveemerged as a tool to modulate biological activities.7–11 Car-diac fibroblasts were engineered in culture to overexpress achimeric protein encoding GATA4 and the CPP VP22. We
recently demonstrated the utility of this strategy to deliver thenonsecreted marker protein green fluorescent protein (GFP)to myocardial tissue.12 In this previous study, we demon-strated that VP22 chimeric proteins are delivered to �1 mm2
of myocardium in a direction radial to the needle track. In thepresent study, we quantified the effects of sustainedGATA4:VP22 release into the infarct border zone in a ratmodel of ischemic cardiomyopathy induced by left anteriordescending (LAD) ligation 1 month before cell delivery. Weaim to show that the cell-based sustained delivery of nonse-creted functional proteins offers a potential novel strategy tostudy the effects of nonsecreted proteins in adult hearts at atime remote from myocardial injury, as well as a potentiallyinteresting strategy for the optimization of cardiac functionfollowing myocardial injury.
Materials and MethodsPlasmid ConstructsGATA4:VP22 constructs were made by inserting GATA4 (gener-ously provided by Dr J. D. Molkentin at Cincinnati Children’sHospital Medical Center, Ohio) into C termini of VP22 in frame inthe plasmid pVP22/myc-His (Invitrogen, Carlsbad, Calif). Thecytomegalovirus promoter drove GATA4 expression in the vectorpVP22/GATA4. The anticipated molecular mass for fusion protein
Original received November 5, 2006; revision received April 10, 2007; accepted May 1, 2007.From the Departments of Cell Biology (J.B., M.K., M.S.P.), Biomedical Engineering (M.S.P.), and Cardiovascular Medicine (Z.B.P., C.B., L.L.R.,
M.S.P.), Cleveland Clinic Foundation; and Department of Chemical and Biomedical Engineering (J.B., M.S.P.), Cleveland State University, Ohio.Correspondence to Marc S. Penn, MD, PhD, Director, Bakken Heart-Brain Institute, Departments of Cardiovascular Medicine and Cell Biology, NE3,
9500 Euclid Ave, Cleveland, OH 44195. E-mail [email protected]© 2007 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000269778.75877.68
1626
Integrative Physiology
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from

GATA4:VP22 was �80 kDa. Plasmids of pVP22/GFP and GFPwere constructed and transfected into rat cardiac fibroblasts asdescribed previously.12
Luciferase GATA4 promoter reporter construct containingGATA4 binding site was made using the GATA motif (A/T)GATA(A/G).13 The insert containing (A/T) GATA (A/G) motifwith 3 repeats was pGL3 vector (Promega Corp, Madison, Wis) at
the XhoI and HindIII sites. The DNA sequence insert from 5� to 3�ligated into pGL3 was as depicted in Figure 1C.
Effect of GATA4:VP22 on Gene Expression inMesenchymal Stem CellsCell culture 6-well plates and inserts (BD Biosciences) were used totest the effect of intercellular delivery of GATA4 on mesenchymal
A
GATA4CMV VP22
B
GATA4 mycCMV VP22
GATA4 VP22
a b c d100100
anti-GATA4
GATA4:VP22
VP22
GATA4
anti-myc
75
50
37kDa
75
50
37kDa
CXhoI insert luciferaseHindIII
anti GATA4 anti myc
-actin
GATA4 binding sequence between two insert sites XhoI and HindIIIIn pGL3 vector: (XhoI) CTCGAGATCTGCCAGCCTGAGATA
e
ACAGGGCCAGCCTGAGATAACAGGGCCAGCCTGAGATAACAGGAAGCTT (HindIII)
D
**
ativ
e lu
cife
rase
acti
vity
1
2 *
Rel
a
HeLa cell stable transfectant
0
Null GATA4 GATA4:VP22
E
Figure 1. Intercellular delivery of GATA4:VP22 and its DNA binding activity. A,Plasmid construct of GATA4:VP22 wasconstructed with GATA4 at the C termi-nus of VP22 and a myc tag at the C ter-minus of GATA4. B, Western blot ofRCFs transfected with GATA4:VP22 (lanea) and GATA4 (lane b) plasmid constructsprobed with a monoclonal anti-GATA4antibody; RCFs transfected with GATA4:VP22 (lane c) and VP22 (lane d) plasmidconstructs probed with monoclonal anti-myc. The expected molecular masses ofGATA4, VP22, and GATA4:VP22 are 46,38, and 84 kDa, respectively. Westernblotting results of HeLa and RCFs trans-fected with a GATA4:VP22 plasmid con-struct probed with monoclonal anti-mycand a polyclonal anti-GATA4 antibody. C,A GATA4 responsive promoter constructthat was generated by placing a GATA4responsive element containing 5�-(A/T)GATA(A/G)-3� sites was inserted into apGL3–luciferase reporter plasmid. D,GATA4 promoter–luciferase constructwas transfected into HeLa cells that sta-bly expressed GATA4 or the GATA4:VP22 constructs. Luciferase activity wasquantified (24 hours after transfection)and is expressed as fold induction rela-tive to PGL3 without plasmid. Treatmentgroups were performed in triplicates perexperiment. The data are representativeof 6 separate experiments and representmeans�SD. *P�0.05. E, RepresentativeWestern blot of �-actin, troponin I, andNkx2.5 expression in MSCs grown in thepresence of cardiac fibroblasts thatexpress GATA4:VP22 or GFP:VP22 for 3and 7 days.
Bian et al Cell-Based Delivery of GATA4 1627
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from

stem cells (MSCs) in vitro. MSCs isolated from the bone marrowof Lewis rat14 (5000 cells) were placed in one well of the cellculture plates. Rat cardiac fibroblasts (5000 cells) stably express-ing GFP, GFP:VP22, or GATA4:VP22 were added to the cellculture insert cup. The bottom layer of the cup consisted of aporous polyethylene terephthalate membrane with 1-�m pores.Analyses of these cells are detailed in the online data supplementat http://circresahajournals.org.
Myocardial Infarction and Cell DeliveryThe Animal Research Committee approved all animal protocols, andall animals were housed in the Association for the Assessment andAccreditation of Laboratory Animal Care animal facility of theCleveland Clinic. Myocardial infarction and cell delivery wereperformed as described previously5,6 and as detailed in the onlinedata supplement.
EchocardiographyWe performed echocardiography as detailed previously15,16 and asdescribed in the online data supplement.
Quantification of Cardiac Myosin Expressionin MyocardiumTo quantify cardiac myosin expression in the infarct border zone andareas remote from myocardial infarction, immunofluorescence wasperformed by incubation with monoclonal cardiac myosin heavy chain(Chemicon) with secondary donkey anti-mouse Alexa fluor 594(Invitrogen). Fluorescence images were obtained with a Leica TCS SP2AOBS confocal laser scanning system (Leica Co, Wetzlar, Germany).
Means of fluorescence intensity (FI) of cardiac myosin in themyocardium were measured from 4 chosen fields per animal, eitherin the periinfarct zone (FIp) or remote areas (FIr) using ImagePro Plussoftware at a magnification of �63. Relative fluorescence intensity(R) was obtained by dividing mean of fluorescence intensity inperiinfarct zone by the one in remote areas per animal.
Statistical AnalysisAll data are presented as means�SD. Comparisons between groupsfor continuous variables were made by Student t test or varianceanalysis followed by post hoc Tukey’s honestly significantly differ-ent test for multiple comparisons, where appropriate. For segmentalradial strain data, we used repeated measures analysis of variance toanalyze the change of segmental radial strains during a 6-weektreatment period, followed by analysis of contrasts. Results wereconsidered statistically significant if P�0.05. For animal survivalanalysis Kaplan–Meier curves were constructed to assess survivalrates during follow up. Log rank statistics was used to comparesurvival of GATA4 treatment group with the survival of 2 mergedcontrol groups. The control groups were merged to eliminate theneed for multiple comparisons.
ResultsGATA4:VP22 Is Delivered Intercellularlyand Binds in the NucleusA plasmid encoding GATA4:VP22 was constructed thatincluded a myc tag in the C terminus of the fusion proteins(Figure 1A). We transfected this construct into HeLa cellsand rat cardiac fibroblasts (RCFs). Western blot using anti-bodies against myc or GATA4 epitopes showed that thechimeric protein was expressed with the predicted molecularmass in RCFs with the GATA4:VP22 construct being 84 kDa(Figure 1B).
To verify that the chimeric GATA4 protein retained itsability to activate GATA4 responsive promoters, we used aluciferase reporter assay with a GATA4 responsive promoterin HeLa cells. A GATA4 binding sequence containing
5�-(A/T)GATA(A/G)-3� was inserted into the pGL3–lucif-erase construct (Figure 1C). The reporter construct wastransiently transfected into wild-type HeLa cells or HeLacells stably expressing empty vector, GATA4, orGATA4:VP22. The results of luciferase assay demonstratedthat GATA4:VP22 chimeric protein maintained the ability toactivate a GATA4 responsive promoter (Figure 1D).
To further determine whether the chimeric GATA4 proteinwas active, we evaluated the effects of coculturing MSCs inthe presence of GFP-, GFP:VP22-, and GATA4:VP22-expressing cardiac fibroblasts. In these experiments, weseparately cultured the MSCs and the cardiac fibroblasts byculturing the MSCs on the cell culture well and growing thecardiac fibroblasts on a well insert. In this configuration, theMSCs were exposed to proteins secreted by the cardiacfibroblasts. Western blot analysis of the MSCs after 3 and 7days after culture demonstrated that the MSCs had increasedexpression of the GATA4 inducible gene troponin I. Theupregulation of troponin I in the presence of GATA4:VP22-expressing, but not GFP:VP22-expressing, cardiac fibroblastsis consistent with the GATA4:VP22 construct having nativeGATA4 activity. Interestingly, we also observed an increasein the cardiac transcription factor Nkx2.5 as well as thestructural protein �-actin.
Intercellular Delivery of GATA4 Improved LeftVentricular Function of Infarcted HeartsWe investigated the effects of intercellular delivery ofGATA4 1 month after myocardial infarction on left ventric-ular (LV) remodeling and function. LV function and contrac-tility were evaluated by detailed echocardiography. RCFsstably expressing GATA4:VP22 or control groups of RCFsstably expressing GFP:VP22 or GFP were directly injectedinto the periinfarct zones of Lewis rats 1 month after LADligation in 5 divided doses around the infarct zone.5,6
Baseline echocardiography was performed within 2 daysbefore cell transplantation (4 weeks after LAD ligation) and4 and 6 weeks after cell transplantation (total, 10 weeks afterLAD ligation). Four weeks after cell transplantation, thepercentage change in LV fractional shortening compared withbaseline was significantly increased (37% increase) in theGATA4:VP22 group compared with a decline in function inthe GFP:VP22 (�11%, P�0.013) and the GFP (�9%,P�0.027) control groups (Figure 2A). Six weeks after celltransplantation, the improvement in fractional shortening wasmaintained in hearts that received GATA4:VP22 (54% in-crease) compared with the GFP:VP22 (�15%, P�0.036) andGFP (�27%, P�0.007) control groups (Figure 2A).
Systolic and diastolic thickness of the anterior and inferiorwalls and shortening fraction at baseline as well as 4 and 6weeks after cell transplantation are presented in Tables 1 and2, respectively. The data in Table 1 demonstrate that betweenbaseline and 4 and 6 weeks, there was a trend toward greaterpathological hypertrophy of the inferior wall in those animalsthat received control cardiac fibroblasts compared with thosethat received GATA4:VP22-expressing cardiac fibroblasts.Despite the increase in wall thickness, or perhaps because ofthe pathological nature of the wall thickening in the nonin-farcted myocardium, the wall thickening of the inferior wall
1628 Circulation Research June 8, 2007
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from
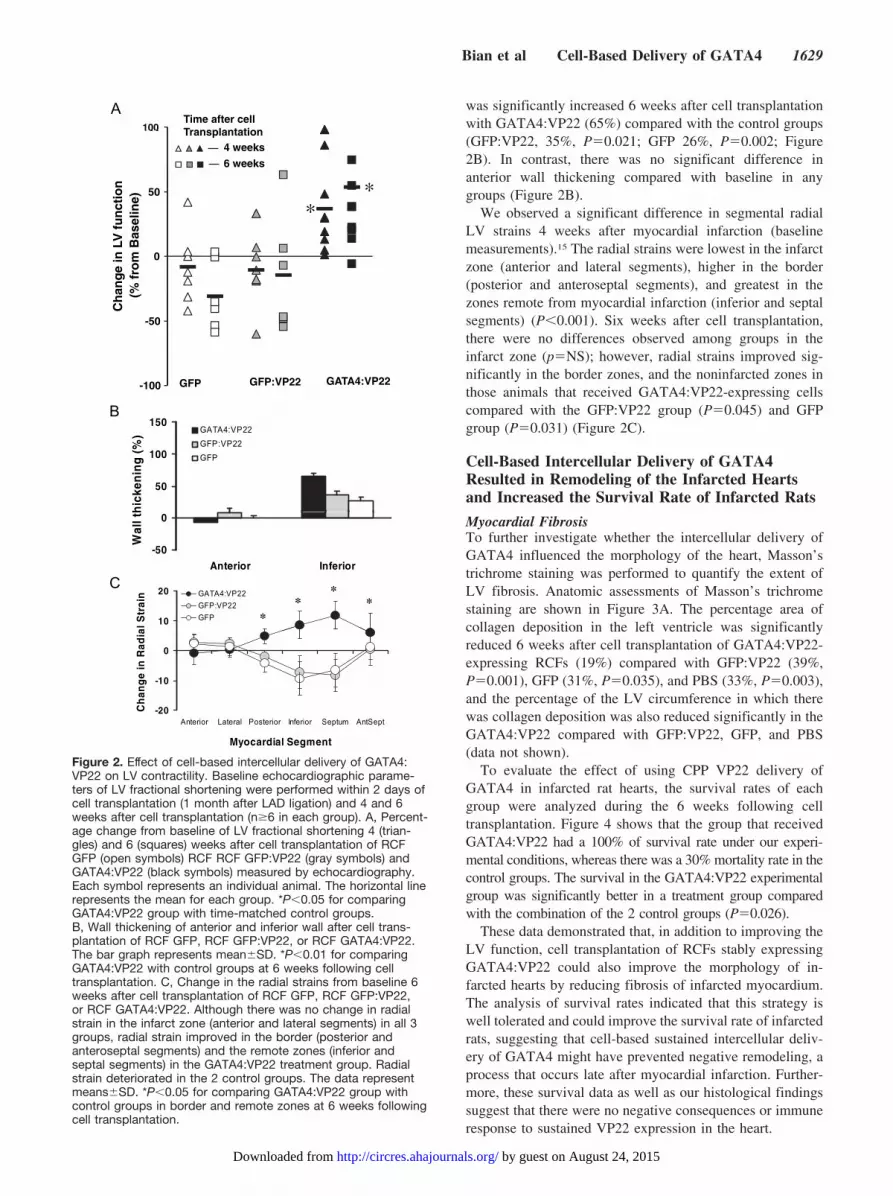
was significantly increased 6 weeks after cell transplantationwith GATA4:VP22 (65%) compared with the control groups(GFP:VP22, 35%, P�0.021; GFP 26%, P�0.002; Figure2B). In contrast, there was no significant difference inanterior wall thickening compared with baseline in anygroups (Figure 2B).
We observed a significant difference in segmental radialLV strains 4 weeks after myocardial infarction (baselinemeasurements).15 The radial strains were lowest in the infarctzone (anterior and lateral segments), higher in the border(posterior and anteroseptal segments), and greatest in thezones remote from myocardial infarction (inferior and septalsegments) (P�0.001). Six weeks after cell transplantation,there were no differences observed among groups in theinfarct zone (p�NS); however, radial strains improved sig-nificantly in the border zones, and the noninfarcted zones inthose animals that received GATA4:VP22-expressing cellscompared with the GFP:VP22 group (P�0.045) and GFPgroup (P�0.031) (Figure 2C).
Cell-Based Intercellular Delivery of GATA4Resulted in Remodeling of the Infarcted Heartsand Increased the Survival Rate of Infarcted Rats
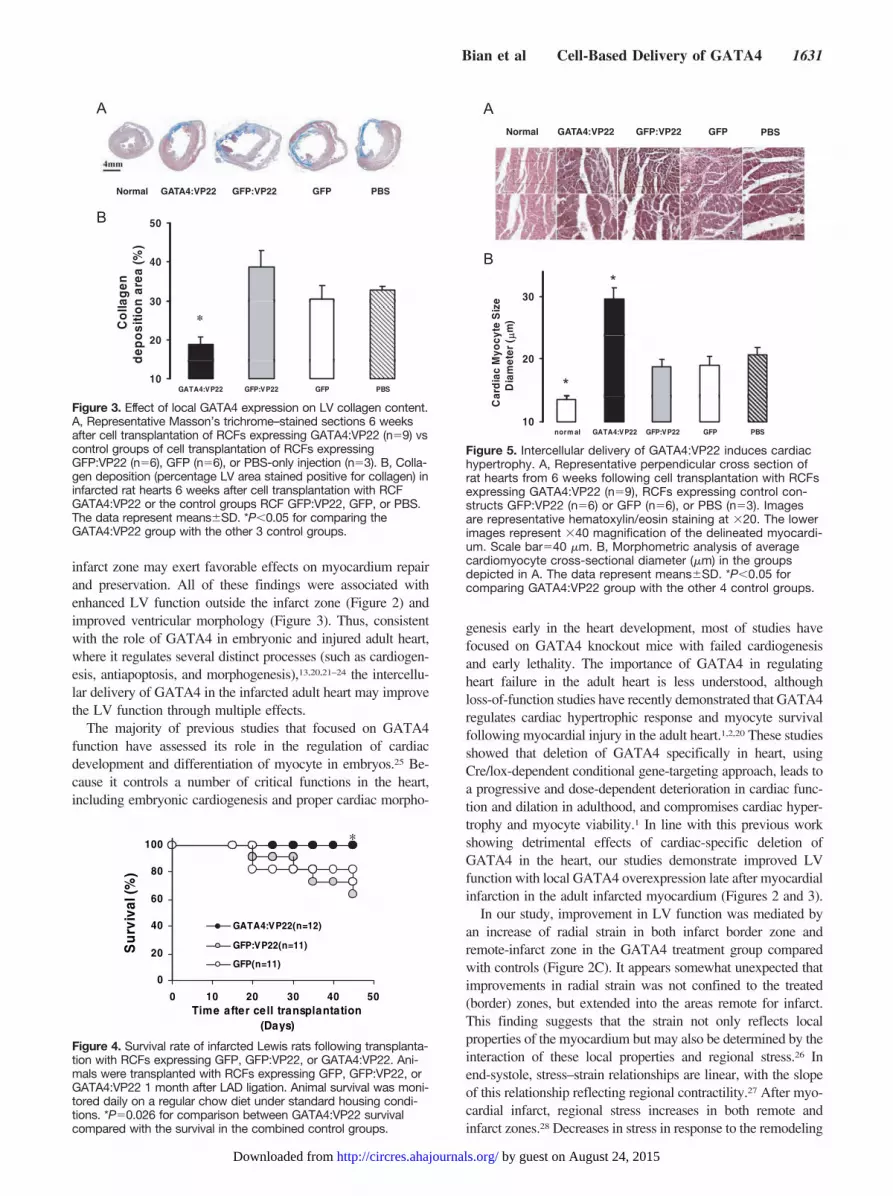
Myocardial FibrosisTo further investigate whether the intercellular delivery ofGATA4 influenced the morphology of the heart, Masson’strichrome staining was performed to quantify the extent ofLV fibrosis. Anatomic assessments of Masson’s trichromestaining are shown in Figure 3A. The percentage area ofcollagen deposition in the left ventricle was significantlyreduced 6 weeks after cell transplantation of GATA4:VP22-expressing RCFs (19%) compared with GFP:VP22 (39%,P�0.001), GFP (31%, P�0.035), and PBS (33%, P�0.003),and the percentage of the LV circumference in which therewas collagen deposition was also reduced significantly in theGATA4:VP22 compared with GFP:VP22, GFP, and PBS(data not shown).
To evaluate the effect of using CPP VP22 delivery ofGATA4 in infarcted rat hearts, the survival rates of eachgroup were analyzed during the 6 weeks following celltransplantation. Figure 4 shows that the group that receivedGATA4:VP22 had a 100% of survival rate under our experi-mental conditions, whereas there was a 30% mortality rate in thecontrol groups. The survival in the GATA4:VP22 experimentalgroup was significantly better in a treatment group comparedwith the combination of the 2 control groups (P�0.026).
These data demonstrated that, in addition to improving theLV function, cell transplantation of RCFs stably expressingGATA4:VP22 could also improve the morphology of in-farcted hearts by reducing fibrosis of infarcted myocardium.The analysis of survival rates indicated that this strategy iswell tolerated and could improve the survival rate of infarctedrats, suggesting that cell-based sustained intercellular deliv-ery of GATA4 might have prevented negative remodeling, aprocess that occurs late after myocardial infarction. Further-more, these survival data as well as our histological findingssuggest that there were no negative consequences or immuneresponse to sustained VP22 expression in the heart.
100
150
(%) GATA4:VP22
GFP:VP22
B
50
100
hic
ken
ing
GFP
-50
0
Anterior Inferior
Wal
l th
10
20
al S
trai
n GATA4:VP22
GFP:VP22
GFP **
**
C
-10
0
ng
e in
Rad
ia
Myocardial Segment
-20Anterior Lateral Posterior Inferior Septum AntSept
Ch
an
y g
100Time after cellTransplantation
A
50
100ct
ion
n
e)Transplantation
4 weeks
6 weeks
**
0
ang
e in
LV
fu
n%
fro
m B
asel
i *
-50
Ch
a (%
-100 GFP GFP:VP22 GATA4:VP22
Figure 2. Effect of cell-based intercellular delivery of GATA4:VP22 on LV contractility. Baseline echocardiographic parame-ters of LV fractional shortening were performed within 2 days ofcell transplantation (1 month after LAD ligation) and 4 and 6weeks after cell transplantation (n�6 in each group). A, Percent-age change from baseline of LV fractional shortening 4 (trian-gles) and 6 (squares) weeks after cell transplantation of RCFGFP (open symbols) RCF RCF GFP:VP22 (gray symbols) andGATA4:VP22 (black symbols) measured by echocardiography.Each symbol represents an individual animal. The horizontal linerepresents the mean for each group. *P�0.05 for comparingGATA4:VP22 group with time-matched control groups.B, Wall thickening of anterior and inferior wall after cell trans-plantation of RCF GFP, RCF GFP:VP22, or RCF GATA4:VP22.The bar graph represents mean�SD. *P�0.01 for comparingGATA4:VP22 with control groups at 6 weeks following celltransplantation. C, Change in the radial strains from baseline 6weeks after cell transplantation of RCF GFP, RCF GFP:VP22,or RCF GATA4:VP22. Although there was no change in radialstrain in the infarct zone (anterior and lateral segments) in all 3groups, radial strain improved in the border (posterior andanteroseptal segments) and the remote zones (inferior andseptal segments) in the GATA4:VP22 treatment group. Radialstrain deteriorated in the 2 control groups. The data representmeans�SD. *P�0.05 for comparing GATA4:VP22 group withcontrol groups in border and remote zones at 6 weeks followingcell transplantation.
Bian et al Cell-Based Delivery of GATA4 1629
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from
Cardiac Myocyte HypertrophyGATA4 is known to be involved in cardiac hypertrophy, whichcan protect the heart by reducing wall stress and the subsequentsparing of energy and oxygen consumption,17 thereby compen-sating for decreased cardiac function following myocardialinfarction1 or pressure overload.2 Therefore we examined theextent of cardiac hypertrophy by measuring myocyte cross-sectional areas in the infarct border zone and areas remote fromthe infarct zone 6 weeks following cell transplantation withGATA4:VP22-expressing RCFs or the control cell groups.
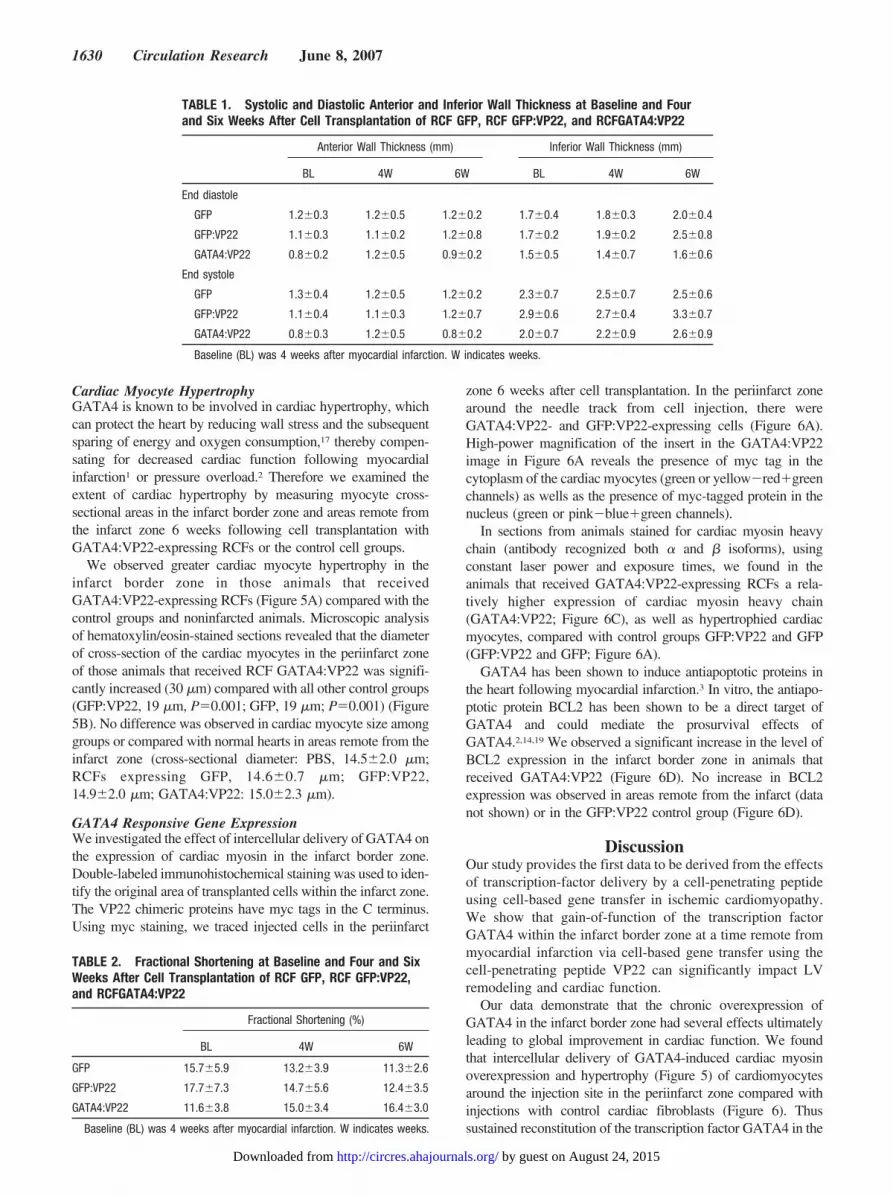
We observed greater cardiac myocyte hypertrophy in theinfarct border zone in those animals that receivedGATA4:VP22-expressing RCFs (Figure 5A) compared with thecontrol groups and noninfarcted animals. Microscopic analysisof hematoxylin/eosin-stained sections revealed that the diameterof cross-section of the cardiac myocytes in the periinfarct zoneof those animals that received RCF GATA4:VP22 was signifi-cantly increased (30 �m) compared with all other control groups(GFP:VP22, 19 �m, P�0.001; GFP, 19 �m; P�0.001) (Figure5B). No difference was observed in cardiac myocyte size amonggroups or compared with normal hearts in areas remote from theinfarct zone (cross-sectional diameter: PBS, 14.5�2.0 �m;RCFs expressing GFP, 14.6�0.7 �m; GFP:VP22,14.9�2.0 �m; GATA4:VP22: 15.0�2.3 �m).
GATA4 Responsive Gene ExpressionWe investigated the effect of intercellular delivery of GATA4 onthe expression of cardiac myosin in the infarct border zone.Double-labeled immunohistochemical staining was used to iden-tify the original area of transplanted cells within the infarct zone.The VP22 chimeric proteins have myc tags in the C terminus.Using myc staining, we traced injected cells in the periinfarct
zone 6 weeks after cell transplantation. In the periinfarct zonearound the needle track from cell injection, there wereGATA4:VP22- and GFP:VP22-expressing cells (Figure 6A).High-power magnification of the insert in the GATA4:VP22image in Figure 6A reveals the presence of myc tag in thecytoplasm of the cardiac myocytes (green or yellow�red�greenchannels) as wells as the presence of myc-tagged protein in thenucleus (green or pink�blue�green channels).
In sections from animals stained for cardiac myosin heavychain (antibody recognized both � and � isoforms), usingconstant laser power and exposure times, we found in theanimals that received GATA4:VP22-expressing RCFs a rela-tively higher expression of cardiac myosin heavy chain(GATA4:VP22; Figure 6C), as well as hypertrophied cardiacmyocytes, compared with control groups GFP:VP22 and GFP(GFP:VP22 and GFP; Figure 6A).
GATA4 has been shown to induce antiapoptotic proteins inthe heart following myocardial infarction.3 In vitro, the antiapo-ptotic protein BCL2 has been shown to be a direct target ofGATA4 and could mediate the prosurvival effects ofGATA4.2,14,19 We observed a significant increase in the level ofBCL2 expression in the infarct border zone in animals thatreceived GATA4:VP22 (Figure 6D). No increase in BCL2expression was observed in areas remote from the infarct (datanot shown) or in the GFP:VP22 control group (Figure 6D).
DiscussionOur study provides the first data to be derived from the effectsof transcription-factor delivery by a cell-penetrating peptideusing cell-based gene transfer in ischemic cardiomyopathy.We show that gain-of-function of the transcription factorGATA4 within the infarct border zone at a time remote frommyocardial infarction via cell-based gene transfer using thecell-penetrating peptide VP22 can significantly impact LVremodeling and cardiac function.
Our data demonstrate that the chronic overexpression ofGATA4 in the infarct border zone had several effects ultimatelyleading to global improvement in cardiac function. We foundthat intercellular delivery of GATA4-induced cardiac myosinoverexpression and hypertrophy (Figure 5) of cardiomyocytesaround the injection site in the periinfarct zone compared withinjections with control cardiac fibroblasts (Figure 6). Thussustained reconstitution of the transcription factor GATA4 in the
TABLE 1. Systolic and Diastolic Anterior and Inferior Wall Thickness at Baseline and Fourand Six Weeks After Cell Transplantation of RCF GFP, RCF GFP:VP22, and RCFGATA4:VP22
Anterior Wall Thickness (mm) Inferior Wall Thickness (mm)
BL 4W 6W BL 4W 6W
End diastole
GFP 1.2�0.3 1.2�0.5 1.2�0.2 1.7�0.4 1.8�0.3 2.0�0.4
GFP:VP22 1.1�0.3 1.1�0.2 1.2�0.8 1.7�0.2 1.9�0.2 2.5�0.8
GATA4:VP22 0.8�0.2 1.2�0.5 0.9�0.2 1.5�0.5 1.4�0.7 1.6�0.6
End systole
GFP 1.3�0.4 1.2�0.5 1.2�0.2 2.3�0.7 2.5�0.7 2.5�0.6
GFP:VP22 1.1�0.4 1.1�0.3 1.2�0.7 2.9�0.6 2.7�0.4 3.3�0.7
GATA4:VP22 0.8�0.3 1.2�0.5 0.8�0.2 2.0�0.7 2.2�0.9 2.6�0.9
Baseline (BL) was 4 weeks after myocardial infarction. W indicates weeks.
TABLE 2. Fractional Shortening at Baseline and Four and SixWeeks After Cell Transplantation of RCF GFP, RCF GFP:VP22,and RCFGATA4:VP22
Fractional Shortening (%)
BL 4W 6W
GFP 15.7�5.9 13.2�3.9 11.3�2.6
GFP:VP22 17.7�7.3 14.7�5.6 12.4�3.5
GATA4:VP22 11.6�3.8 15.0�3.4 16.4�3.0
Baseline (BL) was 4 weeks after myocardial infarction. W indicates weeks.
1630 Circulation Research June 8, 2007
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from
infarct zone may exert favorable effects on myocardium repairand preservation. All of these findings were associated withenhanced LV function outside the infarct zone (Figure 2) andimproved ventricular morphology (Figure 3). Thus, consistentwith the role of GATA4 in embryonic and injured adult heart,where it regulates several distinct processes (such as cardiogen-esis, antiapoptosis, and morphogenesis),13,20,21–24 the intercellu-lar delivery of GATA4 in the infarcted adult heart may improvethe LV function through multiple effects.
The majority of previous studies that focused on GATA4function have assessed its role in the regulation of cardiacdevelopment and differentiation of myocyte in embryos.25 Be-cause it controls a number of critical functions in the heart,including embryonic cardiogenesis and proper cardiac morpho-
genesis early in the heart development, most of studies havefocused on GATA4 knockout mice with failed cardiogenesisand early lethality. The importance of GATA4 in regulatingheart failure in the adult heart is less understood, althoughloss-of-function studies have recently demonstrated that GATA4regulates cardiac hypertrophic response and myocyte survivalfollowing myocardial injury in the adult heart.1,2,20 These studiesshowed that deletion of GATA4 specifically in heart, usingCre/lox-dependent conditional gene-targeting approach, leads toa progressive and dose-dependent deterioration in cardiac func-tion and dilation in adulthood, and compromises cardiac hyper-trophy and myocyte viability.1 In line with this previous workshowing detrimental effects of cardiac-specific deletion ofGATA4 in the heart, our studies demonstrate improved LVfunction with local GATA4 overexpression late after myocardialinfarction in the adult infarcted myocardium (Figures 2 and 3).
In our study, improvement in LV function was mediated byan increase of radial strain in both infarct border zone andremote-infarct zone in the GATA4 treatment group comparedwith controls (Figure 2C). It appears somewhat unexpected thatimprovements in radial strain was not confined to the treated(border) zones, but extended into the areas remote for infarct.This finding suggests that the strain not only reflects localproperties of the myocardium but may also be determined by theinteraction of these local properties and regional stress.26 Inend-systole, stress–strain relationships are linear, with the slopeof this relationship reflecting regional contractility.27 After myo-cardial infarct, regional stress increases in both remote andinfarct zones.28 Decreases in stress in response to the remodeling
A
Normal GATA4:VP22 GFP:VP22 GFP PBS
B 50
30
40
agen
n a
rea
(%)
20
30
Co
llad
epo
sitio
n
*
10GATA4:VP22 GFP:VP22 GFP PBS
d
Figure 3. Effect of local GATA4 expression on LV collagen content.A, Representative Masson’s trichrome–stained sections 6 weeksafter cell transplantation of RCFs expressing GATA4:VP22 (n�9) vscontrol groups of cell transplantation of RCFs expressingGFP:VP22 (n�6), GFP (n�6), or PBS-only injection (n�3). B, Colla-gen deposition (percentage LV area stained positive for collagen) ininfarcted rat hearts 6 weeks after cell transplantation with RCFGATA4:VP22 or the control groups RCF GFP:VP22, GFP, or PBS.The data represent means�SD. *P�0.05 for comparing theGATA4:VP22 group with the other 3 control groups.
100 *
60
80
val (
%)
20
40
Su
rviv
GATA4:VP22(n=12)
GFP:VP22(n=11)
GFP(n=11)
0
0 10 20 30 40 50Time after cell transplantation
(Days)(Days)
Figure 4. Survival rate of infarcted Lewis rats following transplanta-tion with RCFs expressing GFP, GFP:VP22, or GATA4:VP22. Ani-mals were transplanted with RCFs expressing GFP, GFP:VP22, orGATA4:VP22 1 month after LAD ligation. Animal survival was moni-tored daily on a regular chow diet under standard housing condi-tions. *P�0.026 for comparison between GATA4:VP22 survivalcompared with the survival in the combined control groups.
Normal GATA4 VP22 GFP VP22 GFP PBS
ANormal GATA4:VP22 GFP:VP22 GFP PBS
B
30
yte
Siz
em
)
*
20
ard
iac
Myo
cyD
iam
eter
(
*
10norm al GATA4:VP22 GFP:VP22 GFP PBS
Ca
Figure 5. Intercellular delivery of GATA4:VP22 induces cardiachypertrophy. A, Representative perpendicular cross section ofrat hearts from 6 weeks following cell transplantation with RCFsexpressing GATA4:VP22 (n�9), RCFs expressing control con-structs GFP:VP22 (n�6) or GFP (n�6), or PBS (n�3). Imagesare representative hematoxylin/eosin staining at �20. The lowerimages represent �40 magnification of the delineated myocardi-um. Scale bar�40 �m. B, Morphometric analysis of averagecardiomyocyte cross-sectional diameter (�m) in the groupsdepicted in A. The data represent means�SD. *P�0.05 forcomparing GATA4:VP22 group with the other 4 control groups.
Bian et al Cell-Based Delivery of GATA4 1631
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from

of fibrotic myocardium improves regional fractional shortening(a measure related to radial strain) of remote regions.29 There-fore, a regional improvement of myocardial properties in anysegment can lead to better global contraction, smaller end-sys-tolic volume, and therefore reduction of stresses and improve-ment in strains26 in all LV regions. The only regions in whichimprovement will not occur, as seen in our study, are regionswith complete or near-complete fibrosis, with the strains beingvirtually nonexistent because of high-tensile properties of fi-brous tissue and lack of contractile tissue.28
Moreover, this improved LV function was associated withhigher survival rate in the GATA4 treatment group (Figure 4).With that said, we and others have demonstrated the potential forcell based therapies to alter the arrhythmogenic potential of themyocardium.30,31 The cardiac myocyte hypertrophy induced byGATA4:VP22, combined with the heterogeneity of cardiacmyocytes size induced by local GATA4 delivery, could theoret-ically increase arrhythmogenic risk of the remodeled ventricle.Future studies will be required to determine whether localalterations in cardiac myocyte gene expression alters the arrhyth-mogenic potential of the infarct border zone.
As we know, the limited mitotic capacity of adult cardiomyo-cyte restricts the repair of the ischemic myocardium, leading toreplacement by fibrotic tissue, which disrupts proper contractilefunction and results in decreased cardiac performance. Cell-based intercellular delivery of GATA4 might prevent the postin-farct remodeling and death rate by direct protection throughantiapoptotic mechanisms (Figure 6C) and by preserving ven-tricular morphology and reducing fibronecrosis (Figure 3) andby inducing cardiac myosin overexpression in cardiomyocytes(Figure 6A and 6B) and myocardial hypertrophy in the infarctborder zone (Figure 5). Because there was no functional im-provement in the infarct zone, GATA4 might only function inthe survival and remodeling of spared, but not deadmyocardium.
Cell-penetrating peptides have been previously used to delivera vast range of different biologically active proteins in differenttissues.9,32 Our data provide the first evidence using cell-penetrating peptide to modulate remodeling in ischemic cardio-myopathy. As compared with other delivery vectors, such aselectroporation and microinjection that are impractical to use invivo or liposome encapsulation and receptor-mediated endocy-tosis that are limited by the lack of targeting and low yield ofdelivery, cell-based delivery of peptide-based chimeric proteinsis, to date, the only method that succeeds in delivering a cargo
AGATA4:VP22 GFP:VP22 GFP
myc
+DA
PI
m
B
C
Figure 6. GATA4 responsive gene expression. A, Immunofluo-rescent staining of myc tag (fluorescein isothiocyanate [FITC],green), cardiac myosin (CM) heavy chain (Alexa fluor 594, red),and cellular nuclei (2�,6�-diamidino-2-phenylindole [DAPI], blue)were performed on the tissue sections 6 weeks after transplan-tation (10 weeks after LAD ligation) with RCF GATA4:VP22 orcontrol groups RCF GFP:VP22 and RCF GFP. Images wereobtained from periinfarct zones in areas of cell injection. DAPIand myc merged images are presented at the top, and DAPI,myc, and cardiac myosin merged images are in the bottom
panels. Scale bar�40 �m. B, High-resolution image of inset inGATA4:VP22 image in A. Yellow is the combination of greenand red channels (myc tag in presence of cardiac myosin); pinkis the combination of blue and green channels (myc tag innucleus). C, Quantification of cardiac myosin expression in car-diac myocytes in the infarct border zone following transplanta-tion with RCF GATA4:VP22 or control groups RCF GFP:VP22and RCF GFP. The data represent means�SD. *P�0.05 forcomparing GATA4:VP22 group with time-matched controlgroups. D, Representative confocal microscopy images follow-ing BCL2 staining (FITC, green), cardiac myosin heavy chain(Alexa fluor 594, red), and cellular nuclei (DAPI, blue) of tissuesections 6 weeks following transplantation with RCF GATA4:VP22 or control group RCF GFP:VP22 or normal noninfarctedmyocardial tissue. Merged images are in the top panels. Thebottom panels are BCL2 staining alone. Scale bar�40 �m.
1632 Circulation Research June 8, 2007
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from

without disturbing the plasma membrane and is applicable invivo.7 From the analysis of survival rate at 6 weeks beyond thetime an antibody response could have been mounted (Figure 4),we provide evidence that cell-based intercellular delivery couldprovide sustained release of functional proteins in the injuredheart without causing toxic side effects.
In conclusion, we implemented a new carrier system thatcombines cell transplantation with cell-penetrating peptideVP22 to induce sustained local intercellular delivery offunctional transcription factors such as GATA4 to the injuredheart. This sustained delivery of GATA4 resulted in multiplechanges in the myocardium, including alterations in cardiacfibrosis and local hypertrophy of cardiac myocytes, with thefinal result of improved cardiac remodeling and function.
Sources of FundingThis work was supported by NIH grant HL74400, the ShalomFoundation, the Wilson Foundation, and the State of Ohio.
DisclosuresM.S.P. and M.K. are named as coinventors on a pending patent filedby the Cleveland Clinic Foundation that relates to the use of cellpenetrating peptides for modulation of LV remodeling.
References1. Oka T, Maillet M, Watt AJ, Schwartz RJ, Aronow BJ, Duncan SA,
Molkentin JD. Cardiac-specific deletion of Gata4 reveals its requirement forhypertrophy, compensation, and myocyte viability. Circ Res. 2006;98:837–845.
2. Bisping E, Ikeda S, Kong SW, Tarnavski O, Bodyak N, McMullen JR,Rajagopal S, Son JK, Ma Q, Springer Z, Kang PM, Izumo S, Pu WT.Gata4 is required for maintenance of postnatal cardiac function andprotection from pressure overload-induced heart failure. Proc Natl AcadSci U S A. 2006;103:14471–14476.
3. Kobayashi S, Lackey T, Huang Y, Bisping E, Pu WT, Boxer LM, LiangQ. Transcription factor gata4 regulates cardiac BCL2 gene expression invitro and in vivo. FASEB J. 2006;20:800–802.
4. Yau TM, Fung K, Weisel RD, Fujii T, Mickle DA, Li RK. Enhancedmyocardial angiogenesis by gene transfer with transplanted cells.Circulation. 2001;104(suppl I):I-218 –I-222.
5. Askari A, Unzek S, Goldman CK, Kiedrowski M, Forudi F, Ellis SG,Thomas JD, Topol EJ, DiCorleto PE, Penn MS. Cellular, but not directadenoviral delivery of VEGF results in improved LV function and neo-vascularization in dilated ischemic cardiomyopathy. J Am Coll Cardiol.2004;43:1908–1914.
6. Askari A, Unzek S, Popovic ZB, Goldman CK, Forudi F, Kiedrowski M,Rovner A, Ellis SG, Thomas JD, DiCorleto PE, Topol EJ, Penn MS.Effect of stromal-cell-derived factor-1 on stem cell homing and tissueregeneration in ischemic cardiomyopathy. Lancet. 2003;362:697–703.
7. Lundberg P, Langel U. A brief introduction to cell-penetrating peptides.J Mol Recognit. 2003;16:227–233.
8. Maiolo JR, Ferrer M, Ottinger EA. Effects of cargo molecules on thecellular uptake of arginine-rich cell-penetrating peptides. BiochimBiophys Acta. 2005;1712:161–172.
9. Green KL, Southgate TD, Mulryan K, Fairbairn LJ, Stern PL, Gaston K.Diffusible VP22–E2 protein kills bystander cells and offers a route forcervical cancer gene therapy. Hum Gene Ther. 2006;17:147–157.
10. Lee KC, Hamstra DA, Bullarayasamudram S, Bhojani MS, Moffat BA,Dornfeld KJ, Ross BD, Rehemtulla A. Fusion of the HSV-1 tegumentprotein vp22 to cytosine deaminase confers enhanced bystander effectand increased therapeutic benefit. Gene Ther. 2006;13:127–137.
11. Suzuki K, Murtuza B, Smolenski RT, Sammut IA, Suzuki N, Kaneda Y,Yacoub MH. Cell transplantation for the treatment of acute myocardialinfarction using vascular endothelial growth factor-expressing skeletalmyoblasts. Circulation. 2001;104(suppl I):I-207–I-212.
12. Bian J, Kiedrowski M, Mal N, Forudi F, Penn MS. Engineered celltherapy for sustained local myocardial delivery of nonsecreted proteins.Cell Transplant. 2006;15:67–74.
13. Molkentin JD. The zinc finger-containing transcription factors GATA-4,-5, and -6. Ubiquitously expressed regulators of tissue-specific geneexpression. J Biol Chem. 2000;275:38949–38952.
14. Schenk S, Mal N, Finan A, Zhang M, Kiedrowski M, Popovic Z, McCarthyPM, Penn MS. Monocyte chemotactic protein-3 is a myocardial mesen-chymal stem cell homing factor. Stem Cells. 2007;25:245–251.
15. Popovic ZB, Benejam C, Bian J, Mal N, Drinko J, Lee K, Forudi F, ReegR, Greenberg NL, Thomas JD, Penn MS. Speckle tracking echocardiog-raphy correctly identifies segmental left ventricular dysfunction inducedby scarring in a rat model of myocardial infarction. Am J Physiol HeartCirc Physiol. In press.
16. Cho GY, Chan J, Leano R, Strudwick M, Marwick TH. Comparison oftwo-dimensional speckle and tissue velocity based strain and validation withharmonic phase magnetic resonance imaging. Am J Cardiol. 2006;97:1661–1666.
17. Grossman W, Jones D, McLaurin LP. Wall stress and patterns of hyper-trophy in the human left ventricle. J Clin Invest. 1975;56:56–64.
18. Rota M, Boni A, Urbanek K, Padin-Iruegas ME, Kajstura TJ, Fiore G,Kubo H, Sonnenblick EH, Musso E, Houser SR, Leri A, Sussman MA,Anversa P. Nuclear targeting of Akt enhances ventricular function andmyocyte contractility. Circ Res. 2005;97:1332–1341.
19. Misao J, Hayakawa Y, Ohno M, Kato S, Fujiwara T, Fujiwara H.Expression of bcl-2 protein, an inhibitor of apoptosis, and Bax, an accel-erator of apoptosis, in ventricular myocytes of human hearts with myo-cardial infarction. Circulation. 1996;94:1506–1512.
20. Liang Q, De Windt LJ, Witt SA, Kimball TR, Markham BE, MolkentinJD. The transcription factors GATA4 and GATA6 regulate cardiomyo-cyte hypertrophy in vitro and in vivo. J Biol Chem. 2001;276:30245–30253.
21. Grepin C, Nemer G, Nemer M. Enhanced cardiogenesis in embryonicstem cells overexpressing the GATA-4 transcription factor. Development.1997;124:2387–2395.
22. Suzuki YJ, Nagase H, Day RM, Das DK. GATA-4 regulation of myo-cardial survival in the preconditioned heart. J Mol Cell Cardiol. 2004;37:1195–1203.
23. Watt AJ, Battle MA, Li J, Duncan SA. GATA4 is essential for formationof the proepicardium and regulates cardiogenesis. Proc Natl Acad SciU S A. 2004;101:12573–12578.
24. Garg V, Kathiriya IS, Barnes R, Schluterman MK, King IN, Butler CA,Rothrock CR, Eapen RS, Hirayama-Yamada K, Joo K, Matsuoka R,Cohen JC, Srivastava D. GATA4 mutations cause human congenital heartdefects and reveal an interaction with TBX5. Nature. 2003;424:443–447.
25. Molkentin JD, Lin Q, Duncan SA, Olson EN. Requirement of the tran-scription factor GATA4 for heart tube formation and ventral morpho-genesis. Genes Dev. 1997;11:1061–1072.
26. Delhaas T, Arts T, Bovendeerd PH, Prinzen FW, Reneman RS. Subepi-cardial fiber strain and stress as related to left ventricular pressure andvolume. Am J Physiol. 1993;264:H1548–H1559.
27. Kaseda S, Tomoike H, Ogata I, Nakamura M. End-systolic pressure-volume, pressure-length, and stress-strain relations in canine hearts. Am JPhysiol. 1985;249:H648–H654.
28. Rohde LE, Aikawa M, Cheng GC, Sukhova G, Solomon SD, Libby P,Pfeffer J, Pfeffer MA, Lee RT. Echocardiography-derived left ventricularend-systolic regional wall stress and matrix remodeling after experi-mental myocardial infarction. J Am Coll Cardiol. 1999;33:835–842.
29. DiDonato M, Toso A, Dor V, Sabatier M, Menicanti L, Fantini F,Buckberg G. Mechanical synchrony: role of surgical ventricular resto-ration in correcting LV dyssynchrony during chamber rebuilding. HeartFail Rev. 2004;9:307–315.
30. Mills WR, Mal N, Kiedrowski MJ, Unger R, Forudi F, Popovic ZB, PennMS, Laurita KR. Stem cell therapy enhances electrical viability in myo-cardial infarction. J Mol Cell Cardiol. 2007;42:304–314.
31. Deglurkar I, Mal N, Mills WR, Popovic ZB, McCarthy P, Blackstone EH,Laurita KR, Penn MS. Mechanical and electrical effects of cell-basedgene therapy for ischemic cardiomyopathy are independent. Hum GeneTher. 2006;17:1144–1151.
32. Dietz GP, Kilic E, Bahr M. Inhibition of neuronal apoptosis in vitro andin vivo using TAT-mediated protein transduction. Mol Cell Neurosci.2002;21:29–37.
Bian et al Cell-Based Delivery of GATA4 1633
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from

Online Supplement Bian et al. Cell based delivery of GATA4
Materials and Methods:
Transfection and luciferase assays
The insert containing DNA sequence (A/T) GATA (A/T) motif in pGL3 luciferase reporter as
described above was transfected into HeLa, HeLa stably expressing the GATA4 expression construct
and HeLa cells stably expressing the GATA4:VP22 construct. Empty pGL3 vector containing no insert
was transfected into the three corresponding cell lines was used as a negative control. In these
experiments promoter activity was determined by the ratio of luciferase activity from GATA4 binding
sequence promoter driven luciferase activity to empty vector for each of the cell lines studied. Similar
levels of cell transfection across groups was confirmed by co-transfection with a plasmid encoding
GFP. Cells were transfected overnight and the percentage of GFP positive cells was determined prior
to assaying for luciferase activity using the Luciferase Assay Kit (Promega, Madison, MI, USA),
which quantifies firefly luciferase.
Cell culture and western blot analysis
Rat cardiac fibroblasts were harvested from rat heart and cultured in vitro,1, 2 and plasmids used for
transfection were prepared using FuGENE6 (Roche, Indianapolis, IN, USA) and a population of stably
rat cardiac fibroblasts were selected by culturing cells in the presence of G418.2 Western blot analysis
was performed 2 and probed with anti-myc monoclonal antibody (Santa Cruz, CA, USA) and anti-
GATA4 polyclonal antibody (Santa Cruz).
Effect of GATA4:VP22 on gene expression in mesenchymal stem cells
Cell-culture 6-well plates and inserts (BD Biosciences) were used to test the effect of intercellular
delivery of GATA4 on meschenchymal stem cells (MSC) in vitro. Mesenchymal stem cells isolated
from the bone marrow of Lewis rat 3 (5,000 cells) were placed in one well of the cell-culture plate. Rat
cardiac fibroblasts (5,000 cells) stably expressing GFP, GFP:VP22 or GATA4:VP22 were added to the
cell-culture insert cup. The bottom layer of the cup consisted of a porous PET membrane with 1 µm

Online Supplement Bian et al. Cell based delivery of GATA4
2
pores. This configuration allowed diffusion of proteins secreted by the rat cardiac fibroblasts into the
cell culture well containing the mesenhymal stem cells. After 3 and 7 days of culture, the mesenchymal
stem cells from the bottom well were harvested and analyzed by western blot. Western blot analysis of
MSCs was performed using monoclonal anti-alpha sacromeric actin (Sigma, MI, USA), rabbit
polyclonal anti-Troponin I (Santa Cruz) and goat polyclonal anti-Nkx 2.5 (Santa Cruz). Mouse
monoclonal anti-β actin (Sigma) was used as loading control. Secondary antibodies were abeled with
peroxidase and the gels were developed using ECL and chemiluminescence.
Myocardial infarction and cell delivery
The Animal Research Committee approved all animal protocols, and all animals were housed in
the Association for the Assessment and Accreditation of Laboratory Animal Care animal facility of the
Cleveland Clinic. Myocardial infarction was induced by ligation of the left anterior descending
coronary artery in male Lewis rats weighing 250-300 gram.1, 4 The presence of infarct was assessed by
echocardiography immediately after chest closure. Infarcted animals were randomized into four
treatment groups RCF-GFP:VP22, RCF-GFP, RCF-GATA4:VP22 and PBS.
Four weeks after infarction, the hearts were exposed by thoracotomy1, 2, 4 and 150 µl PBS without
cells (PBS) or RCF expressing GFP:VP22, GATA4:VP22 or GFP (1×106 cells) were injected through
a 30G 1/2 needle (PrecisionGlide Needle, BD Bioscience, Franklin Lakes, NJ) around the border zone
of the left ventricular (LV) infarcted areas in 5 divided doses.1, 2, 4
Echocardiography
We performed echocardiography after LAD ligation, and immediately before and 4 and 6 weeks
after cell transplantation. Echocardiography was performed in conscious rats sedated by ketamine (80
mg/kg body weight, i.p.) by using GE Vivid 7 machine (GE Medical, Milwaukee, WI, USA) equipped
with a 14-MHZ linear transducer. M-mode and 2-dimensional cross-sectional echocardiography data
were acquired at a papillary muscle level and digitally stored for further analysis. 5

Online Supplement Bian et al. Cell based delivery of GATA4
3
From M-mode tracings, anatomical parameters in diastole and systole were obtained. Shortening
fraction (SF) was derived from LV cross-sectional area in 2D short axis view: SF= [(LVEDD-
LVESD)/LVEDD]×100 where LVEDD and LVESD correspond to LV end-diastolic diameter and end-
systolic diameter.
The wall thickening (WT) for anterior and inferior wall was derived from the view in which M-
mode cursor was positioned perpendicular to the LV anterior and inferior walls. The wall thickening
was calculated as WT%=(WTHs-WTHd)/WTHd×100%, where WTH stands for the actual thickness of
the ventricular wall (mm), while d and s stand for end diastole and end systole.
To assess radial strains, we analyzed 2-dimensional high-frame rate (frame rate range 92-123 fps)
echocardiography data using Echopac PC workstation (GE Medical, Milwaukee, Ws). This recently
validated5, 6 software tracks the speckle inherent to a standard two dimensional echocardiography
image to obtain radial strain in six left ventricular (LV) segments. Data from at least two cardiac cycles
were then averaged. All echocardiograms were performed by a sonographer blinded to the treatment.
Similarly an investigator blinded to the identity of each animal performed all analyses.
Histological analysis and Immunohistochemistry
Rats were euthanized (KCl 2 mEq IV to arrest the heart in diastole) under deep anesthesia. The
hearts were excised and with histochoice and cut into 3 transverse slices. The middle slice was
embedded in paraffin and processed for hematoxylin-eosine and Masson’s trichrome staining and the
others were embedded in paraffin and used for immunofluorescence.
Collagen deposition was evaluated from sections (5-µm thickness) of the paraffin-embedded tissue
stained with Masson Trichrome stain as described previously by observers blinded to the identity of
individual samples.7 Images of tissue sections were acquired using an ArtixScan 4000tf (Microtek,
Carson, CA, USA) slide scanner with PathScan Enabler (Meyer Instruments, Houston, TX) at a
resolution of 4000 dpi. ImagePro Plus (Media Cybernetics, Silver Spring, MD, USA) was used to

Online Supplement Bian et al. Cell based delivery of GATA4
4
quantify percentage of myocardial area that had collagen deposition. Percentage length of collagen
deposition (%) was expressed as percentage of total LV cavity circumference occupied by collagen .
Myocyte cross-sectional dimensions in the peri-infarct zone and areas remote from the infarct zone
were measured from hematoxylin and eosin-stained slides. Images were obtained by a blinded observer
using Leica DMR upright microscope (Leica Co., Heidelberg, Germany) with cooled CCD camera (Q-
imaging Retiga Ex, British Columbia, Canada). Morphometric analysis of H&E-stained tissues was
performed using ImagePro Plus software at magnification of 200x. Cross-sectional diameters of five
randomly chosen cardiac myocytes were measured in each of eight randomly chosen fields per animal
by two observers blinded to the identity of the animal from which the images were obtained.8
In order to trace the injected cells in the peri-infarct zone, immunofluorescence was performed on
histochoice-fixed 5-µm-thick sections with antigen retrieval followed by incubation with the specified
antibodies. Since the VP22 chimeric proteins have myc tags in the C-terminus, the tissue sections
were first incubated with FITC labeled goat polyclonal anti-myc (Abcam Inc., Cambridge, MA) and
then incubated with monoclonal cardiac myosin heavy chain (Chemicon Internation Corp.) with Alexa
fluor 594 labeled secondary donkey anti-mouse (Invitrogen). In order to test for BCL2 expression
induced by intercellular delivery of GATA4, tissue sections were stained with antibody of BCL2
(Santa Cruz) with FITC labeled secondary antibody, and then double stained with α−cardiac myosin
heavy chain (Chemicon) with Alexa fluor 594 labeled secondary antibody (Invitrogen). Eight digital
images were obtained by Leica TCS SP2 AOBS confocal laser scanning system (Leica Co., Wetzlar,
Germany) using ultraviolet (351/364 nm), 488 nm argon and krypton/argon (568 nm).

Online Supplement Bian et al. Cell based delivery of GATA4
5
Reference List
(1) Askari A, Unzek S, Popovic ZB, Goldman CK, Forudi F, Kiedrowski M, Rovner A, Ellis SG, Thomas JD, DiCorleto PE, Topol EJ, Penn MS. Effect of stromal-cell-derived factor-1 on stem cell homing and tissue regeneration in ischemic cardiomyopathy. Lancet. 2003;362:697-703.
(2) Bian J, kiedrowski M, Mal N, Forudi F, Penn MS. Engineered cell therapy for sustained local myocardial delivery of nonsecreted proteins. Cell Transplant. 2006;15:67-74.
(3) Schenk S, Mal N, Finan A, Zhang M, kiedrowski M, Popovic Z, McCarthy PM, Penn MS. Monocyte chemotactic protein-3 is a myocardial mesenchymal stem cell homing factor. Stem Cells. 2007;25:245-51.
(4) Askari A, Unzek S, Goldman CK, Kiedrowski M, Forudi F, Ellis SG, Thomas JD, Topol EJ, DiCorleto PE, Penn MS. Cellular, but not direct adenoviral delivery of VEGF results in improved LV function and neovascularization in dilated ischemic cardiomyopathy. J Am Coll Cardiol. 2004;43:1908-14.
(5) Popovic ZB, Benejam C, Bian J, Mal N, drinko J, Lee K, Forudi F, Reeg R, Greenberg NL, Thomas JD, Penn MS. Speckle Tracking Echocardiography Correctly Identifies Segmental Left Ventricular Dysfunction Induced by Scarring in a Rat Model of Myocardial Infarction. Am J Physiol Heart Circ Physiol. 2007. In Press.
(6) Cho GY, Chan J, Leano R, Strudwick M, Marwick TH. Comparison of two-dimensional speckle and tissue velocity based strain and validation with harmonic phase magnetic resonance imaging. Am J Cardiol. 2006;97:1661-6.
(7) Liang Q, De Windt LJ, Witt SA, Kimball TR, Markham BE, Molkentin JD. The transcription factors GATA4 and GATA6 regulate cardiomyocyte hypertrophy in vitro and in vivo. J Biol Chem. 2001;276:30245-53.
(8) Rota M, Boni A, Urbanek K, Padin-Iruegas ME, Kajstura TJ, Fiore G, Kubo H, Sonnenblick EH, Musso E, Houser SR, Leri A, Sussman MA, Anversa P. Nuclear targeting of Akt enhances ventricular function and myocyte contractility. Circ Res. 2005;97:1332-41.

and Marc S. PennJing Bian, Zoran B. Popovic, Carlos Benejam, Matthew Kiedrowski, L. Leonardo Rodriguez
CardiomyopathyEffect of Cell-Based Intercellular Delivery of Transcription Factor GATA4 on Ischemic
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000269778.75877.682007;100:1626-1633; originally published online May 10, 2007;Circ Res.
http://circres.ahajournals.org/content/100/11/1626World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2007/05/10/01.RES.0000269778.75877.68.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on August 24, 2015http://circres.ahajournals.org/Downloaded from





























