Embed Size (px)
Citation preview

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/51411816
EffectofCopperExcessonH2O2AccumulationandPeroxidaseActivitiesinBeanRoots
ARTICLEinACTABIOLOGICAHUNGARICA·JUNE2008
ImpactFactor:0.59·DOI:10.1556/ABiol.59.2008.2.9·Source:PubMed
CITATIONS
11
READS
95
4AUTHORS,INCLUDING:
AnjaGeitmann
McGillUniversity
111PUBLICATIONS2,556CITATIONS
SEEPROFILE
EzzedineElFerjani
UniversityofCarthage,FacultyofSciencesof…
98PUBLICATIONS1,725CITATIONS
SEEPROFILE
Availablefrom:AnjaGeitmann
Retrievedon:05February2016

We studied oxidative stress and peroxidase activity resulting from application of excess copper in the
nutrient medium on the roots of young bean seedlings. The change in H2O2 content, lipid peroxidation
and oxidative enzymes activities were quantified and located. Excess of copper caused a loss of mem-
brane integrity and the formation of hydrogen peroxide (H2O2) as visualized in the transmission electron
microscopy and measured using spectrophotometry. H2O2 accumulated in the intercellular spaces and in
the cell wall. The production of H2O2 was accompanied by an increase in the activity of soluble and ionic
GPX (Guaicol peroxidase, EC 1.11.17), CAPX (Coniferyl alcohol peroxidase) and NADH oxidase.
Keywords: Bean – peroxidase – H2O2 – cell wall – copper
INTRODUCTION
In plant cells, an excess of heavy metal ions shift the balance of free radicals metab-
olism towards an accumulation of hydrogen peroxide (H2O2). In the presence of
redox-active transition metals such as Cu2+ and Fe2+, H2O2 can be converted to the
highly reactive molecules like .OH [13, 14, 31, 38].
These reactive molecules induce the activation of chain reactions leading to oxida-
tive stress.
Cu (copper) toxicity was previously believed to be mainly due to the loss of mem-
brane integrity and the formation of free radicals [36].
Transition metals like copper and iron catalyze the formation of .OH and H2O2
radicals from the non enzymatic chemical reaction via the Haber-Weiss and Fenton
reaction [14, 15].
Acta Biologica Hungarica 59 (2), pp. 233–245 (2008)
DOI: 10.1556/ABiol.59.2008.2.9
0236-5383/$ 20.00 © 2008 Akadémiai Kiadó, Budapest
EFFECT OF COPPER EXCESS
ON H2O2 ACCUMULATION
AND PEROXIDASE ACTIVITIES IN BEAN ROOTS
HOUDA BOUAZIZI,1 H. JOUILI,1 ANJA GEITMANN2 and E. EL FERJANI1*
1 Laboratoire de Bio-Physiologie Cellulaires, Faculté des Sciences de Bizerte, 7021 Zarzouna, Tunisia2 Laboratoire de Microscopie, Institut de recherche en biologie végétale 4101 rue Sherbrooke,
Montréal, Québec H1X 2B2, Canada
(Received: May 3, 2006; accepted: April 13, 2007)
* Corresponding author; e-mail: [email protected]

234 H. BOUAZIZI et al.
Acta Biologica Hungarica 59, 2008
H2O2 is generated via superoxide, during electron transport processes such as pho-
tosynthesis and mitochondrial respiration.
The enzymatic sources of H2O2 generated in response to specific stimuli are still
under discussion; various potential sources have been described in plants. These
include NADPH oxidase [16], cell wall peroxidases, oxalate oxidase, plast amine,
diamine and polyamine oxidase-type enzyme and flavin-containing oxidases [5,
6, 21].
Hydrogen peroxide generated by abiotic and biotic stress can act as a signalling
molecule that regulates plant development, stress adaptation and programmed cell
death [1].
In the cell, the over-production of H2O2 activates peroxide-detoxifying mecha-
nisms; these include scavenging the reactive oxygen by non enzymatic systems such
as ascorbate and α-tocopherol and the use of an enzymatic antioxidant system that
includes glutathion reductase (GR), superoxide dismutase (SOD), catalase (CAT),
guaiacol peroxidase (GPX) and ascorbate peroxidase (APX).
In this work, we tried to highlight the involvement of copper excess in the stimu-
lation of the production of hydrogen peroxide and we located the molecule cyto-
chemically using transmission electron microscopy. Also, the effect of copper excess
on growth, metal accumulation and peroxidase activities in bean roots were studied.
MATERIALS AND METHODS
Plant material and growth conditions
Bean seeds (Phaseolus vulgaris) were germinated on glass plates covered with moist
filter paper in darkness at 25 °C for 3 days. The seedlings were grown in a controlled
chamber at 25 °C ± 2 °C and 65% RH, with a photoperiod of 16 h and a light inten-
sity of 150 µmol/m2/s. Fourteen-day old seedlings were treated for 3 days by addi-
tion of 50 and 75 µM of CuSO4 to the nutrient medium, as previously described by
Mazhoudi et al. [20].
Total copper content and cell wall fractions content
Total copper content
Dried root material was ground to powder and wet-digested in 65% nitric acid (1 ml
per 0.1 g of dry matter). The digested material was resuspended in distilled water.
The Cu content of the roots was determined using an atomic absorption spectropho-
tometer (Perkin Elmer-model 2380, C.R.G.R.).

Copper content in the cell wall fractions
Excised roots were incubated in 1% (v/v) Triton X-100. This solution was periodi-
cally renewed during 3 weeks, which was sufficient for the lysis of the cellular con-
tent. The resulting cell walls were dried and treated according to the same protocol
described previously for the determination of the copper content in soluble fraction.
Quantification of H2O2
Hydrogen peroxide levels were determined according to Sergiev et al. [30]. Root tis-
sues (500 mg) were homogenized in an ice bath with 5 ml 0.1% (w/v) TCA. The
homogenate was centrifuged at 12,000 × g for 15 min and 0.5 ml of the supernatant
was added to 0.5 ml 10 mM potassium phosphate buffer (pH 7.0) and 1 ml 1M KI.
The absorbance of the supernatant was read at 390 nm and the content of H2O2
was extrapolated from a standard curve.
Malondialdehyde determination
The level of lipid peroxidation was evaluated by the thiobarbituric acid (TBA) reac-
tion. Fresh tissue was homogenized in 0.5% (w/v) TBA prepared in 30% (w/v) TCA
as described by Baccouche et al. [2]. The homogenate was incubated at 95 °C in a
water bath for 30 min and then cooled in an ice bath. After centrifugation at 10,000×g
for 10 min, the supernatant was measured at 532 nm and corrected by substracting
the non-specific absorbance at 600 nm. MDA concentration was calculated using an
extinction coefficient of 155 mM–1 cm–1.
Peroxidase extraction
Plant material was extracted in 50 mM K-Phosphate buffer (pH 7.0) containing 5
mM sodium ascorbate and 0.2 mM EDTA. The homogenate was centrifuged at
13,000 × g for 15 min. The resulting supernatant was considered as soluble enzymat-
ic fraction.
The pellet was rinsed several times with extraction buffer, stirred for 1hour in 1 M
NaCl (1.5 ml g–1 FW) in 50 mM K-phosphate buffer and centrifuged at 13,000 × g
for 15min; the supernatant was then desalted and concentrated. The resultant prepa-
ration was considered as ionically bound enzymatic fraction. All operations were
performed at 4 °C.
Soluble and ionic protein concentrations were determined according to Bradford
[8] using bovine serum albumin as a standard.
Oxidative stress in bean roots 235
Acta Biologica Hungarica 59, 2008

236 H. BOUAZIZI et al.
Acta Biologica Hungarica 59, 2008
Enzyme assays
Guaiacol peroxidase activities in both soluble and cell wall fractions were deter-
mined following the increase in absorbance at 470 nm by adding the enzymatic
preparation to 2 ml of 9 mM guaiacol and 10 mM hydrogen peroxide in 50 mM K-
phosphate buffer (pH 7.0) [10].
Coniferyl alcohol peroxidase activities in both soluble and ionically cell wall were
measured by monitoring the decrease in absorbance at 260 nm [25]. NADH-oxidase
activity was measured by monitoring the decrease in absorbance at 340 nm accord-
ing to Ishida et al. [16].
Transmission electron microscopy (TEM)
In situ localization of H2O2
Hydrogen peroxide production was assessed cytochemically via determination of
cerium perhydroxide formation after the reaction of CeCl3 with endogenous H2O2
[26] Fresh roots were cut into slices (2 mm2) and were incubated for 1 h in 5 mM
CeCl3 in 50 mM 3-(N-morpholino) propanesulphonic acid (MOPS) pH 7.2. After
incubation, sections were fixed in 2 % formaldehyde, 2.5% glutaraldehyde in 0.05 M
phosphate buffer (pH 7.2) (freshly prepared). Samples were washed twice in the
same buffer, dehydrated in a graded aceton series (25, 50, 70, 90, 95 and 100%).
Specimens were gradually embedded in Spurr’s resin.
Thin sections of embedded tissues were obtained on a Leica Ultracut microtome,
contrasted by uranyl acetate in ethanol, mounted on uncoated copper grids and
observed using a transmission electron microscope (JEOL-JEM-100 S) operated at
80 kV.
Statistical analysis
The results presented are the values ± standard error obtained from at least six repli-
cates, except for MET (n = 3). Significant differences between treated and control
plants are determined using the ANOVA test (P < 0.05).
RESULTS
Effect of copper excess on seedling growth and protein content
In order to determine the effect of copper excess on the morphological aspect, we
tried to highlight the effect of Cu excess on the production of fresh and dry biomass
and on the longitudinal growth.
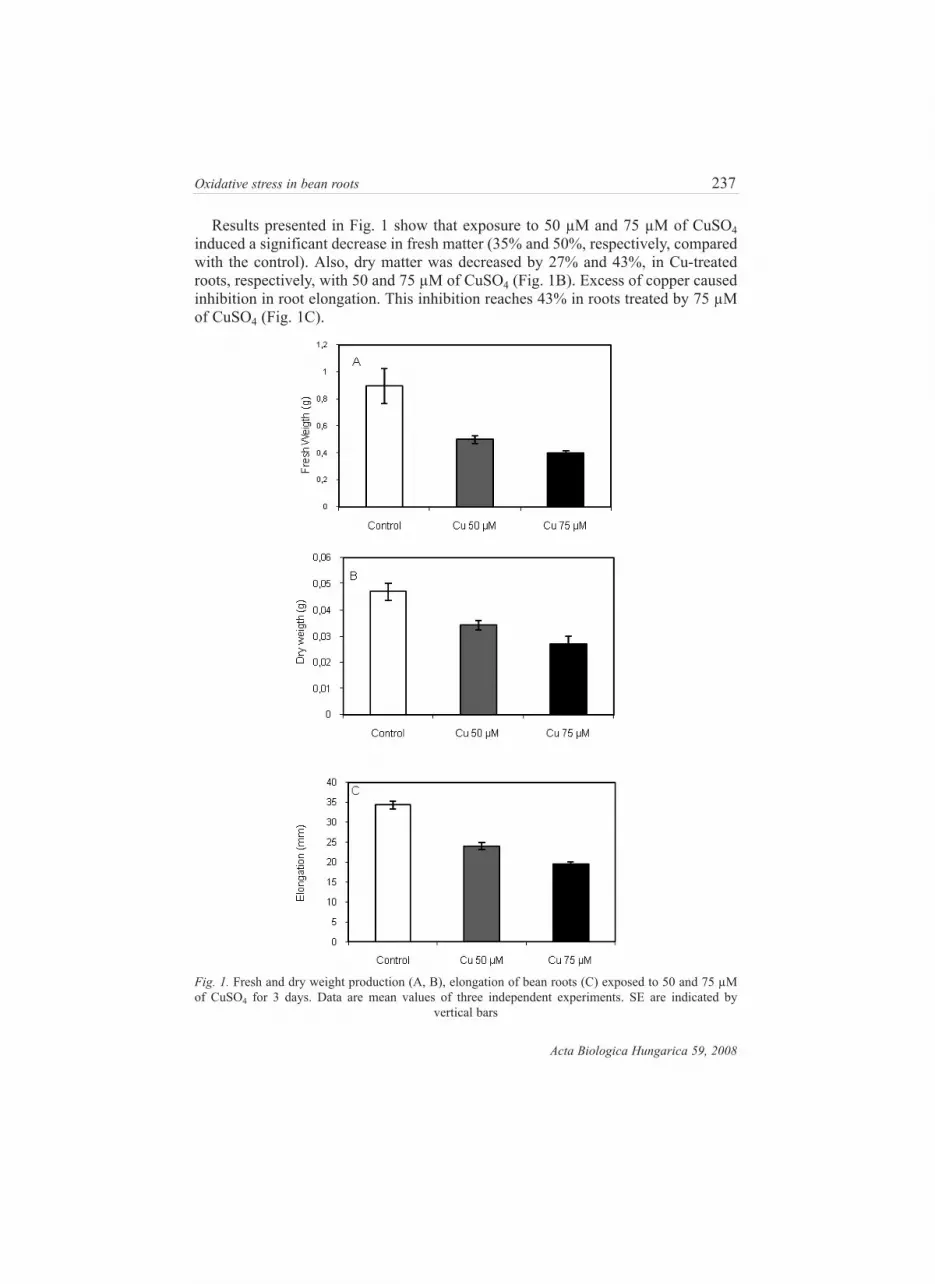
Results presented in Fig. 1 show that exposure to 50 µM and 75 µM of CuSO4
induced a significant decrease in fresh matter (35% and 50%, respectively, compared
with the control). Also, dry matter was decreased by 27% and 43%, in Cu-treated
roots, respectively, with 50 and 75 µM of CuSO4 (Fig. 1B). Excess of copper caused
inhibition in root elongation. This inhibition reaches 43% in roots treated by 75 µM
of CuSO4 (Fig. 1C).
Oxidative stress in bean roots 237
Acta Biologica Hungarica 59, 2008
Fig. 1. Fresh and dry weight production (A, B), elongation of bean roots (C) exposed to 50 and 75 µM
of CuSO4 for 3 days. Data are mean values of three independent experiments. SE are indicated by
vertical bars

238 H. BOUAZIZI et al.
Acta Biologica Hungarica 59, 2008
Soluble protein content was reduced by 70% and 73% in roots, treated with 50 µM
and 75 µM of CuSO4, respectively (Fig. 2A). The cell wall bound proteins were
increased (31% compared with the control) in roots treated with 50 µM CuSO4 and
were decreased (27% compared with the control) in roots treated with 75 µM CuSO4
(Fig. 2B).
Fig. 2. Protein content: Soluble and Ionic fraction (A and B), total copper content (black columns), cop-
per content in the cell wall fraction (white columns) (C), H2O2 level (D), MDA accumulation (E) in bean
roots exposed to 50 and 75 µM of CuSO4 for 3 days. Data are mean values of three independent exper-
iments. SE are indicated by vertical bars

Copper content
When hydroponically grown beans were exposed to 50 and 75 µM of CuSO4, cop-
per was highly accumulated in root tissues.
Figure 2 C shows that the quantity of accumulated copper in the roots treated by
50µM of CuSO4 is sixty times that of the control. In the presence of 75 µM of CuSO4,
the quantity of accumulated copper exceeds 100 times that of the control. The analy-
sis of the parietal fraction of copper in the treated seedlings shows that this fraction
presents 40% of the total copper accumulated on the level of the roots treated by 50
µM of CuSO4. This fraction reaches 50% of total copper accumulated in 75 µM Cu-
treated roots.
H2O2 determination
Hydrogen peroxide levels were measured in the roots of bean seedlings treated with
50 µM and 75 µM of CuSO4. Figure (2D) shows that the copper excess causes a sig-
nificant production of hydrogen peroxide in the treated roots. We also noted that the
level of accumulated H2O2 increases with the concentration of metal in the nutrient
medium. H2O2 production was not detectable in control roots.
MDA determination
The level of malondialdehyde (MDA) was increased significantly in treated roots
(Fig. 2E). This increase reaches 100% in 75 µM Cu-treated roots and 32% in roots
treated by 50 µM of CuSO4. Copper induced a lipoperoxidation process in roots.
Effect of copper excess on peroxidase activities
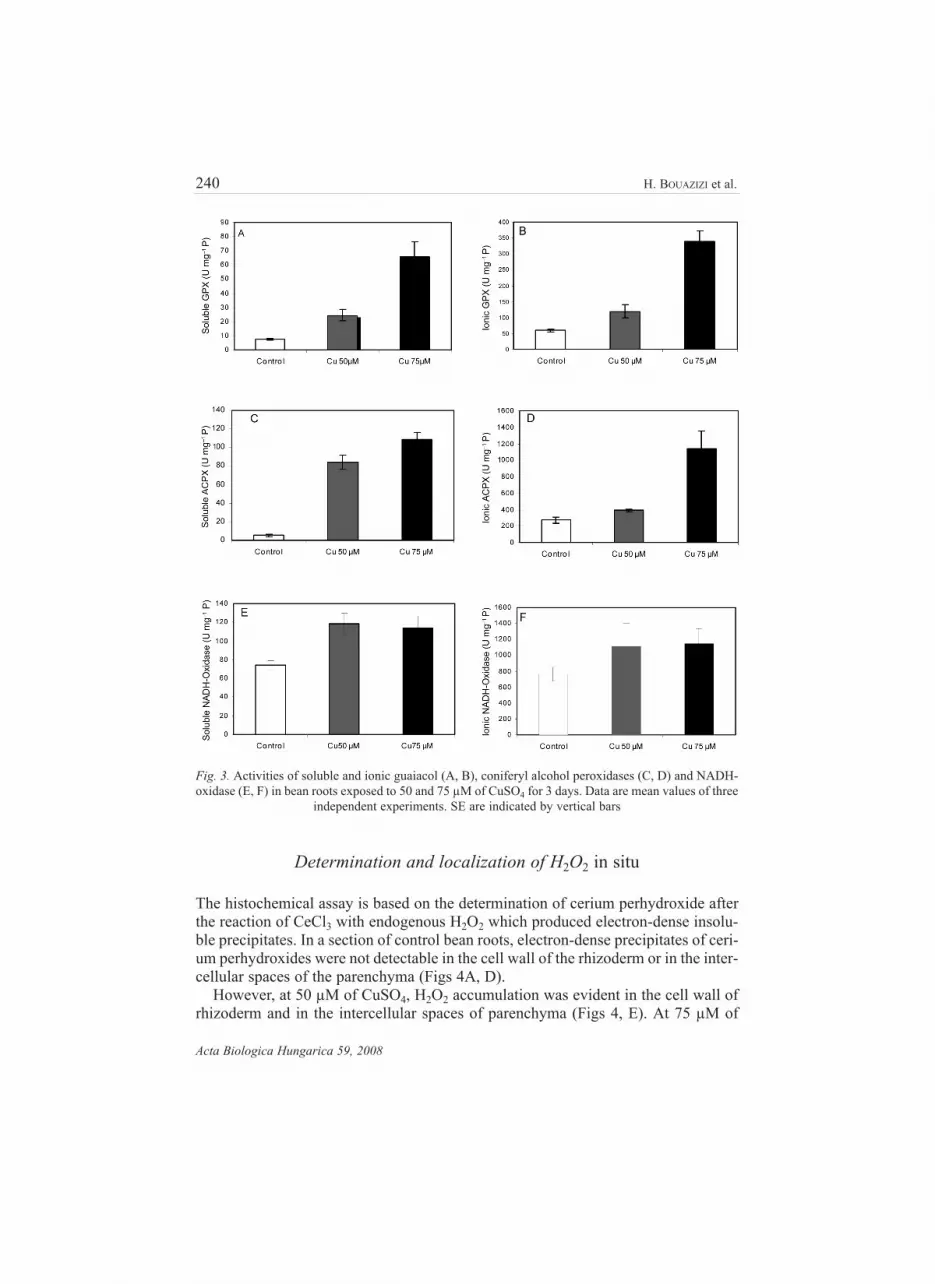
Copper-treatment significantly enhanced the peroxidase activities in soluble and ion-
ically cell wall-bound fractions (Fig. 3). The data show a notable stimulation of sol-
uble and ionic guaiacol peroxidase activities. This stimulation reached 236% and
797% in the soluble fraction after treatment with 50 and 75 µM of CuSO4, respec-
tively (Figs 3A, B). Similarly, we noted a significant increase in soluble and ionic
alcohol peroxidase activities (Figs 3C, D).
The soluble NADH-oxidase activity was modulated by copper excess, when a sig-
nificant increase was detected in roots treated with 50 and 75 µM of CuSO4, this
stimulation is estimated to be 57% and 54%, respectively, compared with the control.
Also, we noted a stimulation in ionic NADH-oxidase activity. This stimulation is
estimated at 46.5% and 50%, respectively, compared to the control (Figs 3E, F).
Oxidative stress in bean roots 239
Acta Biologica Hungarica 59, 2008
240 H. BOUAZIZI et al.
Acta Biologica Hungarica 59, 2008
Determination and localization of H2O2 in situ
The histochemical assay is based on the determination of cerium perhydroxide after
the reaction of CeCl3 with endogenous H2O2 which produced electron-dense insolu-
ble precipitates. In a section of control bean roots, electron-dense precipitates of ceri-
um perhydroxides were not detectable in the cell wall of the rhizoderm or in the inter-
cellular spaces of the parenchyma (Figs 4A, D).
However, at 50 µM of CuSO4, H2O2 accumulation was evident in the cell wall of
rhizoderm and in the intercellular spaces of parenchyma (Figs 4, E). At 75 µM of
Fig. 3. Activities of soluble and ionic guaiacol (A, B), coniferyl alcohol peroxidases (C, D) and NADH-
oxidase (E, F) in bean roots exposed to 50 and 75 µM of CuSO4 for 3 days. Data are mean values of three
independent experiments. SE are indicated by vertical bars

Oxidative stress in bean roots 241
Acta Biologica Hungarica 59, 2008
Fig. 4. Cytochemical localization of Cu-induced H2O2 accumulation in cell wall, rhizoderme (A, B, C)
(×15,000) and intercellular spaces of parenchyma (D, E, F) (×15,000) in bean roots , exposed to 50 and
75 µM of CuSO4 for up to 3 days

242 H. BOUAZIZI et al.
Acta Biologica Hungarica 59, 2008
CuSO4, the electron-dense precipates were clearly visible in the apoplastic spaces of
rhizoderm, in the cell wall and in the apoplastic spaces of parenchyma (Figs 4C, F).
The size of electron-dense precipitates increased with the increase in Cu levels.
DISCUSSION
We examined the physiological responses of bean roots to copper excess, in particu-
lar the effect on peroxidase activities in soluble and cell wall bound fractions. The
effect of Cu2+ ions on soluble and ionic protein contents and H2O2 levels was also
studied. We studied the modulation of peroxidase response under stress application
and we investigated the response at ultrastructural level by visualizing the accumu-
lation and intracellular distribution of H2O2 by transmission electron microscopy
(TEM).
Our results confirm that excess of copper is a potent growth inhibitor. The pres-
ence of 50 µM or 75 µM of CuSO4 in the nutrient medium inhibited the production
of fresh and dry matter and reduced root elongation. The inhibition of growth can be
explained by the activation of peroxidases. These enzymes are involved in the degra-
dation of the endogenous indole-3-acetic acid [12] and in the lignification process
and cell wall cross linking [9].
Accordingly, our results showed the inhibition of growth in bean roots accompa-
nied by an accumulation of copper in both soluble and cell wall fractions. The data
show that copper level increased when Cu amounts increased in nutrient solution.
Also, we noted that the cell wall was the location of the highest Cu accumulation.
Almost half of absorbed copper was fixed to the cell wall. O’Neill et al. [23] showed
that the insoluble copper in cell wall must be bound to a polymeric structural com-
ponent. They suggested that one possible class of binding sites in the primary cell
wall is that comprising acidic polysaccharides, particularly pectins. But pectins do
not bind all the Cu2+ ions [22, 28, 37]. Copper can also be fixed on neutral polysac-
charides, like xyloglucan [17, 27]. Some authors suggested that Cu2+ can bind to cell
wall glycoprotein rich in His and/or Cys residues [31] and affinity for extensins has
been proposed [3, 7].
Fry et al. [11] suggested that the fixation of Cu2+ to the cell wall is likely to act as
a site-specific pro-oxidant, leading to the cleavage of polysaccharide chains. The
accumulation of copper in the cell wall and in the soluble fraction of roots induced
oxidative stress as is evident from increased levels of malondialdehyde (MDA) and
hydrogen peroxide (H2O2). MDA, a cellular indicator of lipid peroxidation was used
to assess oxidative stress. MDA and H2O2 accumulation increased with higher cop-
per concentration in the nutrient medium. The overproduction of H2O2 and accumu-
lation of MDA constitutes two of the detectable metabolic responses to the abiotic
stress especially to heavy metals stress.
Consistent with our results, some authors have reported that copper induced per-
oxidation of polyunsaturated fatty acids in membrane lipids and the generation of

malondialdehyde [19]. Lipid peroxidation process in plants can be initiated by
enzymes such as an á-dioxygenase, peroxidases or lipoxygenases [21].
The release of hydrogen peroxide radicals during abiotic stress is a characteristic
of oxidative stress. The H2O2 produced can play the role of a local signal leading to
cellular death. It can, in the same way, be diffused through the cells causing a change
in the redox status of the cellular membranes and their polarity; this would stimulate
the antioxidant mechanisms of defense [34].
The cytochemical localization of cerium perhydroxide precipitates in Cu-treated
bean roots demonstrated that H2O2 accumulation only occurred in the apoplastic
spaces and it coincided with the increase of the activity of soluble and ionic NADH-
oxidase and coniferyl alcohol peroxidase. The increase of these enzyme activities can
be explained by activation of lignification process in response to cupric stress [9, 18,
19]. In fact, NADH-oxidase could be the primary site of H2O2 generation [4, 26].
The stimulation of soluble and ionic guaiacol and coniferyl alcohol peroxidase
activities can be explained by the role of these enzymes in the elimination of hydro-
gen peroxide. Since these enzymes were reported to reduce H2O2 using phenolic
compounds as electron donors [2, 18, 24, 29]. The hydrogen peroxide would consti-
tute a signal of transduction. It was suggested that the increase in the content of this
radical stimulates the oxidative polymerization of certain organic defense molecules
in the cell wall which form a mechanical barrier towards the stress inducing agent
[33].
In conclusion, the present work has shown that the presence of excess Cu causes
oxidative stress in bean roots and subsequent increase in antioxidant responses due
to increased production of H2O2 and MDA. Accordingly, it was observed that excess
Cu in roots led to lipoperoxidation and production of H2O2 inducing changes in the
activities of soluble and ionic GPX, ACPX and NADH oxidase.
REFERENCES
1. Apel, K., Hirt, H. (2004) Reactive oxygen species: metabolism, oxidative stress, and signal traduc-
tion. Plant Biol. 55, 373–399.
2. Baccouche, S., Chaoui, A., El Ferjani, E. (1998) Nickel induced oxidative damage and antioxidant
responses in Zea mays shoots. Plant Physiol. Biochem. 36, 689–694.
3. Biggs, K. J., Fry, S. C. (1990) Solubilization of covalently bound extensin from capsicum cell walls.
Plant Physiol. 92, 197–204.
4. Bolwell, G. P. (1999) Role of active oxygen species and nitric oxide in plant defence response. Curr.
Opinion Plant Biol. 2, 287–294.
5. Bolwell, G. P., Wojatzek, P. (1997) Mechanisms for the generation of reactive oxygen species in plant
defence – a abroad perspective. Physiological and Molecular Plant Pathology. 51, 347–366.
6. Bolwell, G. P., Bindschedler, L. V., Blee, K. A., Butt, V. S., Davies, D. R., Gardner, S. L., Gerrish,
C., Minibayeva, F. (2002) The apoplastic oxidative brust in response to biotic stress in plants: a three-
component system, Journal of Experimental Botany 53, 1367–1376.
7. Bown, D. P., Bolwell, G. P., Gatehouse, J. A. ( 1993) Characterization of potato (Solanum tuberosum
L.) extensine- a novel extensin –like cDNA from dormant tubers. Gene 134, 229–233.
8. Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of
protein utilizing the principal of protein-dye binding. Anal. Biochem. 72, 248–258.
Oxidative stress in bean roots 243
Acta Biologica Hungarica 59, 2008

244 H. BOUAZIZI et al.
Acta Biologica Hungarica 59, 2008
9. Chen, E. L., Chen, Y. A., Chen, L. M., Liu, Z. H. (2002) Effect of copper on peroxidase activity and
lignin content in Raphanus sativus. Plant Physiol. Biochem. 40, 439– 444.
10. Fielding, J. L., Hall, J. L. (1978) A biochemical and cytochemical study of peroxidase activity in
roots of Pisum sativum. J. Exp. Bot. 29, 979–986.
11. Fry, S. C., Miller, J. G., Dumville, J. C. (2002) A proposed role for copper ions cell wall loosening.
Plant and Soil 247, 57–67.
12. Gazaryon, I. G., Chubar, T. A., Mareeva, E. A., Lagrimini, L. M., Van Huystee, R. B., Thorneleley,
R. N. F. (1999) Aerobic oxidation of indole-3-acetic acid oxidation by peanut peroxidase.
Phytochemistry 51, 175–186.
13. Gupta, M., Cuypers, A. J., Vanngrosveld, Clijsters, H. (1990) Copper affects the enzymes of the
ascorbate-glutathione cycle and its related metabolites in the roots of Phaseolus vulgaris. Plant phys-
iol. 64, 418–423.
14. Halliwell, B., Gutteridge, J. M. C. (1984) Oxygen toxicity, oxygen radicals, transition metals and dis-
ease. Biochem. J. 219, 1–14.
15. Halliwell, B., Gutteridge, J. M. C. (1990) Role of free radicals and catalytic metalions in human dis-
ease; an overview. Meth.Enzymol. 186, 1–85.
16. Ishida, A., Ookubo., Ono, K. (1987) Formation of hydrogen peroxide by NAD(P)H oxidation with
isolated cell wall-associated peroxidase from cultured liverwort cells, Marchantia polymorpha L.
Plant Cell Physiol. 28, 723–726.
17. Jones, J. K. N., Stoodley, R. J. (1965) Fractionation using copper complexes, in: Methods in
Carbohydrate Chemistry, Vol 5. (Eds). RL Whistler, Academic Press, New York, pp 36–38.
18. Jouili, H., El Ferjani, E. (2003) Changes in antioxidant and lignifying enzyme activities in sunflower
roots (Helianthus annuus L.) stressed with copper excess, C. R. Biologies. 326, 639–644.
19. Jouili, H., El Ferjani, E. (2004) Effect of copper excess on superoxide dismutase, catalase and per-
oxidase activities in sunflower seedlings (Helianthus annuus L.). Acta physiologiae Plantarum 26,
29–35.
20. Mazhoudi, S., Chaoui, A., Gorbal, M. H., El Ferjani, E. (1997) Response of antioxidant enzymes to
excess copper in tomato (Lycopersion esculentum, Mill.). Plant Sci. 127, 182–186.
21. Mithöfer, A., Schulze, B., Boland, W. (2004) Biotic and heavy metal stress response in plants: evi-
dence for common signals, FEBS Letters. 566, 1–5.
22. Nikdel, S., Mackellar, D. G., Rezaaiyan, R. (1991) Analysis of mineral-content and amount of chelat-
ed minerals in citrus juice by inductively coupled plasma emission-spectroscopy. J. Agric. Food
Chem. 39, 1775–1775.
23. O’Neill, M., Albersheim, P., Darvill, A. G. (1990) The pectic polysaccharides of primary cell walls.
In: Ed. P. M. Dey. Methods in Plant Biochemistry. 2. Academic Press London, pp. 415–441.
24. Pandolfini, T., Gabbrielli R., Comparini, C. (1992) Nickel toxicity and peroxidase activity in
seedlings of Triticum aestivum L. Plant, Cell Environm. 15, 719–725.
25. Pedereńo, M. A., Barcelo, A. R., Sabatter, F., Munoz, R. (1989) Control by pH of cell wall peroxi-
dase activity involved in lignification. Plant Cell Physiol. 30, 237–241.
26. Ranieri, A., Castagna, A., Pacini, J., Baldan, B., Mensuali, S. Soldatin, G. F. (2003) Early production
and scavenging of hydrogen peroxide in the apoplast of sunflower plants exposed to ozone. Journal
of Experimental Botany 54, 2529–2540.
27. Rao, P. S. (1959) Tamarind. In Industrial Gums. Ed. R. L. Whistler. Academic Press New York, pp.
461–504.
28. Schlemmer, U. (1989) Studies of the binding of copper, zinc and calcium to pectin, alginate, car-
rageenan and gum in HCO3– CO2 buffer. Food Chem. 32, 223–234.
29. Schützendübel, A., Nikolova, I. P., Rudolf, C., Polle, A. (2002) Cadmium and H2O2-induced oxida-
tive stress in Populus×canescens roots. Plant Physiol. Biochem. 40, 577–584.
30. Sergiev, I., Alexieva, V., Karanov, E. (1997) Effect of spermine, atrazine and combination between
them on some endogenous protective systems and stress markers in plants. Compt. Rend. Acad. Bulg.
Sci. 51, 121–124.

31. Sommer-Knudsen, J., Bacic, A. (1997) A micro-scale method for determining relative metal-binding
affinities of proteins. Mol. Biotechnol. 8, 215–218.
32. Van Assche, F., Clijsters, H. (1990) Effects of metals on enzyme activity in plants. Plant Cell
Environ. 13, 195–206.
33. Velikova, V., Tsonev, T., Yordanov, T. (1999) Light- and CO2-responses of photosynthesis and
chlorophyll fluorescence characteristics in bean plants after simulated acid rain. Physiol. Plant. 107,
77–83.
34. Velikova, V., Yordanov, I., Edreva, Q. (2000) Oxidative and some antioxidant systems in acid rain-
treated bean plants. Protection role of exogenous polyamines, Plant Science 151, 59–66.
35. Vranová, E., Inzé, D., Van Breusegem, F. (2002) Signal transduction during oxidative stress, J. Exp.
Botany 53, 1227–1236.
36. Wang, H., Shan, X. B. Wen., Zhang, S., Wang, Z. (2004) Responses of antioxidative enzymes to
accumulation of copper in a copper hyperaccumulator of Commoelina communis, Archive of
Environmental Contamination and Toxicology 47, 185–192.
37. Wucherpfennig, K. (1992) Removing iron and copper ions out of beverages complexing agents.
Deut. Lebensm.-Rundschau. 88, 313–317.
38. Yruela, I. (2005). Copper in plants. Braz. J. Plant Physiol. 17, 145–156.
Oxidative stress in bean roots 245
Acta Biologica Hungarica 59, 2008






























