Embed Size (px)
Citation preview

ORIGINAL RESEARCH
Effect of N-salicyloyltryptamine (STP), a novel tryptamineanalogue, on parameters of cell viability, oxidative stress,and immunomodulation in RAW 264.7 macrophages
Juciano Gasparotto & Matheus Augusto de Bittencourt Pasquali & Nauana Somensi &Laura Milán Vasques & José Claudio Fonseca Moreira & Reinaldo Nobrega de Almeida &
Jose Maria Barbosa-Filho & Maria de Fátima Vanderlei de Souza &
Stanley Juan Chavez Gutierrez & Lucindo José Quintans Júnior & Daniel Pens Gelain
Received: 17 December 2012 /Accepted: 8 April 2013 /Published online: 21 April 2013# Springer Science+Business Media Dordrecht 2013
Abstract Immunomodulatory actions exerted by someclasses of tryptamines, such as benzoyltryptamine ana-logues, suggest these molecules as promising candidatesto develop new therapies to treat conditions associatedto acute and chronic pain and inflammation. N-salicyloyltryptamine (STP) was observed to act as ananticonvulsive agent and exert antinociceptive effects inmouse. In the present work, we performed a screeningof cytotoxic, cytoprotective, immunomodulatory, andredox properties of STP in RAW 264.7 macrophageschallenged with hydrogen peroxide and LPS. Our
results show that STP presents no cytotoxicity in therange of 0.001 to 1 μg/mL, but doses of 50 and100 μg/mL caused loss of cell viability (IC50=22.75 μg/mL). Similarly, STP at 0.001 to 1 μg/mL didnot cause oxidative stress to RAW 264.7 cells, althoughit did not prevent cell death induced by H2O2 0.5 mM.At 1 μg/mL, STP reversed some redox and inflammatoryparameters induced by LPS. These include thiol(sulfhydryl) oxidation, superoxide dismutase activation,and morphological changes associated to macrophageactivation. Besides, STP significantly inhibited LPS-induced TNF-α and IL-1β release, as well as CD40 andTNF-α protein upregulation. Signaling events induced byLPS, such as phosphorylation of ERK 1/2 and IκBα andp65 nuclear translocation (NF-kB activation) were alsoinhibited by STP. These data indicate that STP is able tomodulate inflammatory parameters at doses that do notinterfere in cell viability.
Keywords N-salicyloyltryptamine .
Inflammation . Cytokines . LPS . Oxidative stress .
RAW264.7
Introduction
Tryptamines are monoamine alkaloids widely distrib-uted in different groups of organisms, includingplants, animals, fungi, and prokaryotes (Jones 1982).
Cell Biol Toxicol (2013) 29:175–187DOI 10.1007/s10565-013-9245-2
J. Gasparotto :M. A. de Bittencourt Pasquali :N. Somensi : L. M. Vasques : J. C. F. Moreira :D. P. Gelain (*)Centro de Estudos em Estresse Oxidativo, Departamentode Bioquímica, Instituto de Ciências Básicas da Saúde,Universidade Federal do Rio Grande do Sul—Porto Alegre,Rua Ramiro Barcelos, 2600-anexo,CEP 90035-003 Porto Alegre, RS, Brazile-mail: [email protected]
R. N. de Almeida : J. M. Barbosa-Filho :M. de Fátima Vanderlei de Souza : S. J. C. GutierrezLaboratório de Tecnologia Farmaceutica, UniversidadeFederal da Paraiba, João Pessoa, Paraiba, Brazil
L. J. Q. JúniorDepartamento de Fisiologia, Universidade Federalde Sergipe, Sergipe, São Cristóvão, Brazil

These substances are characterized by the presence ofan indole ring structure, and take part in a variety ofstructural and signaling cellular processes. In mam-mals, endogenous tryptamines (e.g., serotonin andmelatonin) were first recognized by their neurotrans-mitter and neuromodulatory actions, which are in-volved in the control of brain metabolic homeostasis,circadian rhythm regulation, and modulation of excit-atory signaling (Deneris and Wyler 2012; Dieudonne2001). Nonetheless, some tryptamines endogenouslysynthesized by humans, such as melatonin, were laterobserved to exhibit significant antioxidant and anti-inflammatory activities, thus being considered impor-tant immunomodulators (Nosal et al. 2010). Melatoninexerts protective effects in septic and hemorrhagicshock (Sacco et al. 1998), reverses chronic and acuteinflammation (Missbach et al. 1996; Cuzzocrea et al.1997), and endotoxin-induced sickness behaviorslike adepsia and hyperthermia (Nava et al. 1997;Raghavendra et al. 1999). Besides, melatonin re-verses lipopolysacharide (LPS)-induced thermal andbehavioral hyperalgesia in mice (Raghavendra et al.2000).
Based on this wide range of biological effects,interest in the potential pharmacological effects oftryptamine derivatives has been raising. Synthetictriptans such as sumatriptan, naratriptan, and almotriptanare used as antimigraine agents due to neurogenic anti-inflammatory and vasoconstrictive properties (Jhee et al.2001; Evans et al. 2012). As melatonin and its metabo-lites N1-acetyl-N2-formyl-5-methoxykynuramine(AFMK) and N1-acetyl-5-methoxykynuramine (AMK)were observed to exert anti-inflammatory effects bypreventing cyclooxygenase-2 (COX-2) activation in-duced by LPS in macrophages (Mayo et al. 2005),tryptamine derivatives have also been investigated fortheir antinociceptive and anti-inflammatory properties,besides their neuroprotective/neurotoxic potential.
N-Salicyloyltryptamine (STP), a benzoyltryptamineanalogue, was demonstrated to act as an anticonvul-sive agent (Oliveira et al. 2001) exerting neuroprotectiveeffects by preventing the ability of neuronal cells togenerate action potentials (Araujo et al. 2003). STPwas also observed to exert an antinociceptive activity,which was suggested to be associated to the modulationof inflammatory pathways (Quintans et al. 2010). Theseworks raised the interest on the clinical potential of STP,which could be useful in therapies for conditions asso-ciated to pain and inflammation in the central nervous
system and also in other organs (Araujo et al. 2003;Oliveira et al. 2001; Quintans et al. 2010). However,there is no information on whether STP could modulatefunctional parameters on cells of the immune system,such as the expression/activation of inflammatorymarkers induced by LPS. Besides, studies on cytotoxic-ity and cell viability parameters of STP in different cellsare still scarce.
During inflammation, a wide range of signaling path-ways and metabolic processes are activated in macro-phages, which are associated with changes in thetranscription of genes and the phosphorylation state ofseveral proteins. Among these processes, the activationof CD40 by interferon-γ (IFNγ) and CD40L inducesthe activation of nuclear factor-κB (NF-κB), which inturn activates TNF-α and TNF-α receptor expression(Foey et al. 2001; Scott et al. 2004). TNF-α is a pro-inflammatory cytokine and its expression and release bymacrophages is highly stimulated in response to a widerange of inflammatory stimuli, such as LPS (Wang et al.2012; Shih et al. 2012). In macrophages, NF-κB isnormally complexed with IκBα in an inactive form inthe cytoplasm; in response to TNF-α stimulation, IκBαundergoes phosphorylation and degradation, resultingin NF-κB nuclear translocation into the nucleus whereit activates CD40 transcription (Kang et al. 2009; Lin etal. 2012; Yang et al. 2012). NF-κB activation by pro-inflammatory cytokines also results in the upregulationof the receptor for advanced glycation endproducts(RAGE) (Sims et al. 2010). RAGE activation reinforcesthe inflammatory estate by increasing MAPK ERK1/2phosphorylation, resulting in the triggering of signalcascades that further enhance the expression of pro-inflammatory cytokines (Maczurek et al. 2008; Boppet al. 2008). The increase in the production of reactivespecies due to macrophage activation during inflamma-tion contributes to consolidate these changes, as thesesignaling pathways are sensitive to oxidative stress(Halliwell and Gutteridge 2007).
In order to better understand the properties of STPat cellular level in the immune system, we used themacrophage line RAW 264.7 to perform studies onparameters of cell viability and immunomodulation.These cells constitute a suitable model to investigateanti-inflammatory effects of novel compounds, asmacrophages are crucial mediators of the inflammato-ry response to infectious stimuli, such as LPS. Giventhe relationship of inflammatory processes with freeradicals and oxidative stress, we also investigated the
176 Cell Biol Toxicol (2013) 29:175–187

effect of STP in cells challenged with hydrogen per-oxide (H2O2).
Material and methods
Chemicals
Glycine, Griess reagent, sodium nitroprusside (SNP),hydrogen peroxide (H2O2), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), dimethyl sulf-oxide (DMSO), 2′,7′-dichlorohydrofluorescein diacetate(DCFH-DA), sulforhodamine (SRB), 5,5′-dithionitrobis2-nitrobenzoic acid (DTNB), and LPS were purchasedfrom Sigma Chemical Co. (St. Louis, MO, USA).Materials used in cell culture were acquired fromGibco®/Invitrogen (São Paulo, SP, Brazil) and from theRio de Janeiro Cell Bank (BCRJ, Rio de Janeiro, Brazil).Electrophoresis and immunoblot reagents and materialswere from Bio-Rad (Hercules, USA), GE HealthcareBrazilian Headquarter (São Paulo, Brazil), and Sigma.ELISA microplates were from Greiner Bio-One(Monroe, USA) and ELISATMB spectrophotometric de-tection kit was from BD Biosciences (San Diego, USA).CD40 rabbit polyclonal antibody, TNF-α rabbit polyclonalantibody, phosphorylated IκBα rabbit polyclonal antibody,IκB-α rabbit polyclonal antibody, β-actin rabbit polyclon-al antibody, phospho-p44/42 MAPK (ERK1/2) rabbitpolyclonal antibody, p44/42 MAPK (ERK1/2) mousemonoclonal antibody, and anti-rabbit immunoglobulinlinked to peroxidase were from Cell Signaling (Beverly,USA). Purified recombinant TNF-α protein and RAGErabbit polyclonal antibody were from Abcam (Cambridge,UK). Immunoblot chemiluminescence detection was car-ried out with the West Pico detection kit from Pierce(Rockford, USA). All other reagents used in this studywere of analytical or HPLC grade. STP was synthesizedby Laboratory of Pharmaceutical Technology (JoãoPessoa, Paraiba, Brazil; patent claimed, BR 200304393-A) (Quintans et al. 2010). STP was dissolved in DMSO(100 %) and serial dilutions were obtained from this stocksolution. Therefore, at the highest concentration of STP inthe assays, concentration of the vehicle DMSO corre-sponds to 0.1 %.
RAW 264.7 cell culture
Macrophages constitute one of the first-line defensecells in immune response. Thus, we used a well-suited
cell line for in vitro experiments on inflammatoryparameters. Exponentially growing macrophage-derived cell line RAW 264.7, obtained from the Riode Janeiro Cell Bank (BCRJ, Rio de Janeiro, Brazil),were maintained in Dulbecco’s Modified EagleMedium (DMEM) supplemented with 10 % heat-inactivated fetal bovine serum (FBS), 2 mM of gluta-mine, 0.28 μg/μL of gentamicin, and 250 μg ofamphotericin B, in a humidified atmosphere of 5 %of CO2 in air at 37 °C. Cell medium was replaced each2 days and the cells were sub-cultured once theyreached 90 % confluence. All treatments wereperformed when cells were 70–90 % confluence, inDMEM 1 % FBS.
MTT assay
Following treatments, RAW 264.7 metabolic/mitochondrial viability was assessed by the MTT as-say as previously described (Gelain and Moreira2008). This method is based on the ability of viablecells to reduce MTT and form a blue formazan prod-uct. MTT solution (sterile stock solution of 5 mg/mL)was added to the incubation medium in the wells at afinal concentration of 0.2 mg/mL. The cells were leftfor 45 min at 37 °C in a humidified 5 % CO2 atmo-sphere. The medium was then removed and plateswere shaken with DMSO for 30 min. The opticaldensity of each well was measured at 550 nm (test)and 690 nm (reference). H2O2 0.5 mM was used aspositive control for cell death, as this concentrationwas found close to the IC50 value for loss of cellviability in RAW 264.7 cells (data not shown). Datawere expressed as percentage of the formazan forma-tion in untreated cells (control).
SRB incorporation-based viability assay
This colorimetric assay estimates cell number bystaining total cellular protein with SRB (Skehan etal. 1990). Briefly, cells were fixed by layering 100 μlof ice-cold 40 % trichloroacetic acid on top of theculture medium and incubated at 4 °C for 1 h. Plateswere then washed five times with cold water. Theexcess water was then decanted and the plates left todry in air. SRB stain (50 μl; 0.4 % in 1 % acetic acid)was added to each well and left in contact with the cellsfor 30 min. The cells were then washed with 1 % aceticacid, rinsed four times until only the dye adhering to the
Cell Biol Toxicol (2013) 29:175–187 177

cells was left. The plates were then air-dried and 100 μlof 10 mM Tris base pH 10.5 were added to each well todilute the dye. The plates were gently shaken for 20 minon a gyratory shaker and the absorbance of each wellwas read at 492 nm. Cell survival was measured as thepercentage absorbance compared to the absorbance ofcontrol (non-treated cells).
Superoxide dismutase activity
The activity of superoxide dismutase (SOD, EC1.15.1.1) was measured by quantifying the inhibitionof superoxide-dependent adrenaline auto-oxidation.Superoxide generation was initiated by addition ofadrenaline 2 mM, and adrenochrome formation wasmonitored at 480 nm for 5 min (32 °C) in a spectro-photometer. Results are expressed as SOD units permilligram of protein (Misra and Fridovich 1972).
Protein sulfhydryl quantification
Oxidative alterations were used to measure the levelof total reduced sulfhydryl (SH) content in samples(Ellman 1959). Briefly, for total SH content mea-surement, a 50 μg sample aliquot was diluted inPBS and 10 mM DTNB, and read in a spectropho-tometer at 412 nm after 40 min incubation in roomtemperature.
Intracellular reactive species production(DCFH-DA assay)
Intracellular reactive species production was deter-mined by the DCFH-DA-based real-time assay usingintact living cells, as previously described (Gelain etal. 2008). Briefly, 2×104 RAW 264.7 cells were seed-ed in 96-well plates and 100 μM DCFH-DA dissolvedin medium containing 1 % FBS was added to eachwell and incubated for 2 h to allow cellular incorpo-ration. Medium was then discarded and cells weresubjected to treatments; DCF fluorescence was readat 37 °C during 1 h with 10 min intervals in a fluores-cence plate reader (Spectra Max M2, MolecularDevices, USA) with an emission wavelength set at535 nm and an excitation wavelength set at 485 nm.The percentage increase in each well was calculatedaccording to the formula [(Ft2−Ft0)/Ft0×100], whereFt2=fluorescence at time 1 h and Ft0=fluorescence attime 0 min. Hydrogen peroxide (1 mM) was used as
positive control for intracellular reactive speciesproduction.
Nitric oxide (NO) production
NO production was estimated by quantification ofnitrite in the cell culture incubation medium by themethod of Griess, as previously described (de Souza etal. 2006). In brief, cells were treated with the differentconcentrations of STP and co-treatments for 24 h andthen the medium was collected and centrifuged at750×g to precipitate potential cell debris. Supernatantwas collected and incubated 1:1 with Griess reactantfor 30 min as described by the manufacturer. Sampleswere read at 540 nm and nitrite produced was calcu-lated using a SNP-based standard curve.
Western blot
To perform immunoblot experiments, RAW 264.7cells were lysed in 1X radioimmunoprecipitation as-say buffer (20 mM Tris–HCl pH 7.5, 150 mM NaCl,1 mM Na2 EDTA, 1 mM EGTA, 1 % NP-40, 1 %sodium deoxycholate, 2.5 mM sodium pyrophosphate,1 mM b-glycerophosphate, 1 mM Na3VO4, and1 μg/mL leupeptin), and centrifuged (10 min at14,000×g). Supernatant was removed for proteinquantification by Bradford method. ConcentratedLaemmli sample buffer (62.5 mM Tris–HCl, pH 6.8,1 % (w/v) SDS, 10 % (v/v) glycerol) was added andequal amounts of cell protein (30 μg/well) were frac-tionated by SDS-PAGE and then electro-blotted ontonitrocellulose membranes. Protein loading and electro-blotting efficiency were verified through Ponceau Sstaining, and the membrane was blocked in Tween-Tris buffered saline (TTBS, 100 mM Tris–HCl,pH 7.5, containing 0.9 % NaCl, and 0.1 % Tween-20) containing 5 % albumin. Membranes were incu-bated overnight at 4 °C with each antibody separatelyin TTBS, at different working dilutions as suggestedby the manufacturers, and then washed with TTBS.Anti-rabbit IgG peroxidase-linked secondary antibodywas incubated with the membranes for additional 1 h(1:5,000 dilution range), washed again and the immu-noreactivity was detected by enhanced chemilumines-cence using a kit Super Signal West Pico Chemiluminescence kit (Pierce). The membrane was ex-posed to the Western blot imaging system ChemiDOC–Bio-Rad and densitometric analysis was
178 Cell Biol Toxicol (2013) 29:175–187

performed with ImageJ software. Blots were devel-oped to be linear in the range used for densitometry.All results were expressed as a relative ratio to the β-actin or lamin B internal control or their respectivetotal protein.
Cellular fractionation
To obtain the cytoplasmic and nuclear contents thecells were washed with cold phosphate-buffered salineand suspended in 0.4 mL hypotonic lysis buffer(10 mmol/L HEPES pH 7.9, 1.5 mM MgCl2, 10 mMKCl, 0.5 mM phenylmethylsulfonyl fluoride, 0.5 mMdithiothreitol plus protease inhibitor cocktail). After15 min cells were lysed with 12.5 μL 10 % NonidetP-40. The homogenate was centrifuged (13,000×g,30 s), and supernatants containing the cytoplasmicextracts (fraction 1) were stored at −80 °C. Thenuclear pellet was resuspended in 100 μL ice-cold hy-pertonic extraction buffer (10 mM HEPES pH 7.9,0.42 M NaCl, 1.5 mM MgCl2, 10 mM KCl, 0.5 mMphenylmethylsulfonyl fluoride, 1 mM dithiothreitol plusprotease inhibitors). After 40 min with vortex each10 min, the homogenate was centrifuged (13,000×g,10 min, 4 °C), and supernatants containing nuclear pro-teins were collected (de Bittencourt Pasquali et al.2013).
IL-1β and TNF-α release (ELISA)
TNF-α was quantified by indirect ELISA. A 96-wellELISA plate was coated with 200 μl of culture medi-um and purified recombinant TNF-α protein (Abcam—Cambridge, UK) diluted in 50 mM carbonate buff-er, pH 9.0, for standard curve calculation. After 24 hincubation, plates were washed three times withTween-Tris buffered saline (TTBS, 100 mM Tris–HCl, pH 7.5, containing 0.9 % NaCl, and 0.1 %Tween-20). Subsequently, 200 μl of anti-TNF-α(1:10,000) were added and incubation was carriedfor 24 h at 4 °C. The plate was washed three timeswith TTBS and incubated with 200 μl of a rabbit IgGperoxidase-linked secondary antibody (1:10,000) for2 h. After washing the plate three times with TTBS,200 μl of substrate solution (TMB spectrophotometricELISA detection kit) were added to each well andincubated for 15 min. The reaction was terminatedwith 50 μL/well of 12 M sulfuric acid stopping re-agent and the plate read at 450 nm in a microplate
reader. IL-1β was detected by Quantikine® ELISAmouse IL-1β/IL-1 F2 Immunoassay Kit followingthe manufacturer instructions.
Data normalization and statistical analysis
Protein content in samples was quantified for datanormalization according to Lowry and Bradford-based methods. Statistical analysis was performedwith GraphPad 5.0 software. One- or two-wayANOVA were applied, according the number of vari-ables to be considered (see figure legends for details)and differences were considered to be significant whenp was at least lesser than 0.05. Tukey’s post hoc testwas applied following one-way ANOVA. All datapresented here are the result of three or more indepen-dent experiments.
Results
We first evaluated the effect of varying concentrations ofSTP onRAW264.7 cell viability by performingMTTandSRB-based assays. During a 24-h period of incubation,STP at 0.001 to 1 μg/mL did not affect RAW 264.7 cellviability; however, concentrations of 50 and 100 μg/mLsignificantly decreased both MTT reduction (Fig. 1a) andSRB incorporation (Fig. 1b). Concentrations above100 μg/mL produced extensive loss of viability (100 %)and were not evaluated in further experiments (notshown). These results suggest that STP concentrationsequal or below 1 μg/mL are safe for macrophages, as cellviability was not compromised. To identify thetoxic limit of STP on RAW 264.7 cells, we searchedfor IC50 values in the range between 1 and 50 μg/mLand we observed similar values in both MTT reduction(IC50=22.72 μg/mL) and SRB incorporation (IC50=22.75 μg/mL) assays (Fig. 1c and d). Contrast-phasemicroscopy (Fig. 1e) showed that STP 1 μg/mL did notinduce significant alterations in the number and mor-phology of RAW 264.7 cells.
To further characterize the effect of STP on macro-phage viability, we evaluated oxidative stress-relatedparameters in RAW 264.7 cells. We observed thatSTP, at concentrations from 0.001 to 1 μg/mL, didnot modify basal reactive species (RS) production bythe real-time DCFH-DA oxidation assay in RAW264.7 macrophages; however, at 50 μg/mL, intracel-lular RS production was significantly increased in
Cell Biol Toxicol (2013) 29:175–187 179

both 1 and 8 h of incubation (Fig. 2a and b). At100 μg/mL, most cells were detached, probably dueto extensive cytotoxicity (see Fig. 1a, b) so DCFHoxidation-based fluorescence was not detectable (notshown). We also verified the effect of STP on RSproduction in cells treated with H2O2, and we ob-served that co-incubation of STP with H2O2 co-incubation caused a modest decrease in DCFH oxida-tion at 0.05 and 1 μg/mL, but at 50 μg/mL STP furtherincreased RS production by H2O2 (Fig. 2c and d).H2O2 at 0.5 mM was chosen since this concentrationwas observed to cause a significant impairment on cellviability but did not cause extensive cell death (datanot shown). This result suggests that STP, in doses up1 μg/mL, is neither antioxidant nor pro-oxidant to
RAW 264.7 cells, and that cytotoxicity by STP at50 μg/mL or higher concentrations are probably asso-ciated to oxidative stress. To evaluate if STP at 0.05and 1 μg/mL was able to protect RAW 264.7 cellsagainst H2O2-induced cytotoxicity, we performedMTT- and SRB-based assays but no effect was ob-served using H2O2 0.5 mM (Fig. 2e and f). This resultsuggests that STP at 0.05 and 1 μg/mL has no effect(positive or negative) on oxidative stress-induced celldeath in RAW 264.7 cells
We next evaluated the effect of STP on redox andinflammatory parameters affected by macrophage acti-vation by LPS 1 μg/mL. Nitric oxide is a well-knowninflammatory mediator and vasodilator and may induceoxidative stress in pathological conditions (Abramson
Fig. 1 Parameters of viabili-ty in RAW 264.7 cells incu-bated with differentconcentrations of STP. Cellswere incubated for 24 h withSTP from 0.001 to 100 μg/mL in the incubation mediumas described in “material andmethods” section, and viabil-ity was assessed by analyzingMTT reduction (a) and SRBincorporation (b). To deter-mine the IC50 values for STP,concentrations between 1 and50 μg/mL were also testedfor MTT reduction (c) andSRB incorporation (d) as-says. Contrast-phase micros-copy analysis (e) showed noapparent modifications in thenumber and morphology ofcells treated with STP 1 μg/mL for 24 h (×100 magnifi-cation). Bars represent mean± SEM of replicate samplesfrom three independent ex-periments (n=16 in each ex-periment). One-way ANOVAwith Tukey’s post hoc testwas applied. Asterisks denotesignificance level in relationto control (***p<0.001)
180 Cell Biol Toxicol (2013) 29:175–187

2008). We therefore quantified the levels of nitrite, aproduct from NO decomposition, in the incubation me-dium of RAW 264.7 cells incubated for 24 h with STP.We observed that STP (1 μg/mL) had no significanteffect on nitrite accumulation (Fig. 3a). Besides,STP did not change the increase in nitrite productionby LPS. We also assessed the levels of sulfhydrylcontent which are modulated in macrophages due toincreased thiol oxidation during the oxidative stresscaused by LPS activation. In this case, STP reversedthe decrease in sulfhydryl induced by LPS (Fig. 3b),suggesting a protective effect. We also observed thatSTP inhibited the effect of LPS on the activity ofthe antioxidant enzyme SOD (Fig. 3c). Besides,evaluation by contrast-phase microscopy shows thatSTP decreases the number of cells displaying mor-phologic characteristics of activated macrophages in-duced by LPS (Fig. 3e). These results, altogether,indicate that STP affects general parameters of mac-rophage activation induced by LPS.
During infection, macrophages respond to LPS by stim-ulating TNF-α secretion, triggering pro-inflammatory
cascades (Kenneth et al. 2012). LPS also stimulates thesecretion of IL-1β, which plays a central role in immuneand inflammatory responses. We evaluated the effect ofSTP onLPS-induced secretion of the cytokines TNF-α andIL-1β by quantitative ELISA of cell culture medium. At1 μg/mL, STP caused a 30.74 % decrease in LPS-inducedTNF-α secretion by RAW 264.7 cells, suggesting a signif-icant anti-inflammatory effect (Fig. 4a). Besides, the releaseof IL-1β as reduced to control levels (Fig. 4b). Theseresults indicate that STP possesses an anti-inflammatoryeffect at the level of TNF-α and IL-1β release bymacrophages.
To further investigate the effect of STP on param-eters related to cell survival and inflammation, weassessed how STP modulated the expression ofCD40, TNF-α, and RAGE. The total immunocontentof these proteins in cell extracts was assessed byWestern blot. LPS is a potent inducer of CD40 expres-sion in RAW 264.7 cells, and we observed that STPinhibited the effect of LPS during a 24-h period of co-incubation (Fig. 5a). Total levels of TNF-α were alsodecreased by STP in LPS-treated cells (Fig. 5b), which
Fig. 2 Modulation of reactive species production and H2O2-induced cytotoxicity by STP. RAW 264.7 cells were pre-incubated with DCFH-DA for 1 h and then medium containingdifferent concentrations of STP (0.001 to 50 μg/mL) was added.The rate of fluorescence production was evaluated at 1 h (a) and8 h (b) of incubation. The effect of STP in DCFH oxidation inthe presence of H2O2 0.5 mM at 1 h (c) and 8 h (d) of incubationwas also evaluated. To analyze the effect of non-cytotoxicconcentrations of STP on cells submitted to oxidative stress,
RAW 264.7 cells were co-incubated for 24 h with STP at either0.05 or 1 μg/mL in the presence of H2O2 0.5 mM and cellviability was determined by MTT reduction (e) and SRB incor-poration (f) assays. One-way ANOVA with Tukey’s post hoctest was applied. Asterisks denote significance level in relationto control (*p<0.05, **p<0.01, and ***p<0.001). Number signdifferent from ‘H2O2’ group (p<0.05). Vehicle is DMSO 0.1 %.Bars represent mean ± SEM of replicate samples from threeindependent experiments (n=16 in each experiment)
Cell Biol Toxicol (2013) 29:175–187 181

indicate that STP also inhibits the cellular productionof this cytokine. RAGE expression may be enhancedby inflammatory mediators or accumulation of RAGEligands, and RAGE activation is an important compo-nent of chronic pro-inflammatory signaling (Bopp etal. 2008). As observed in Fig. 5c, STP (1 μg/mL) hadno effect on the increase in RAGE induced by LPS.
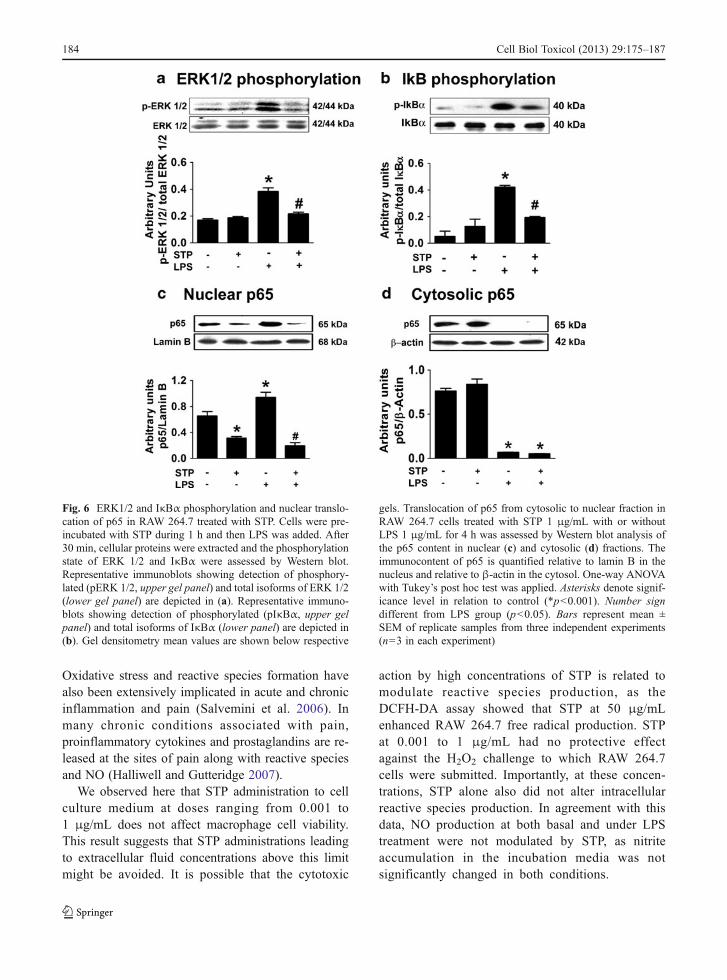
The MAPK ERK 1/2 are main mediators of thesignal cascades triggered by LPS during inflammation.In response to LPS and other pro-inflammatory stim-uli, ERK 1/2 are rapidly activated by extracellularreceptor-dependent phosphorylation and provokedownstream activation of several components of in-flammation signal cascades by phosphorylation(Rehman et al. 2012). STP inhibited the phosphoryla-tion of ERK 1/2 induced by LPS observed in 30 minof incubation (Fig. 6a). In later periods (18–24 h),however, the increase in ERK 1/2 phosphorylation inLPS-treated cells co-incubated with STP is restored,probably due to secondary effects related to pathways
not directly associated with LPS-activated receptors ordue to STP metabolization/cell internalization (notshown).
In RAW 264.7 cells, NF-κB is complexed with IκBαin an inactive form in the cytoplasm. The phosphoryla-tion of IκBα by IKK leads to its dissociation from p65(the active form of NF-κB) and targets IκBα toproteasomic degradation, allowing translocation ofNF-κB into the nucleus and activation of gene transcrip-tion associated with inflammation (Kang et al. 2009).We analyzed the phosphorylation of IκB-α by Westernblot and observed that STP was able to inhibit theincrease in phosphorylated IκB-α levels induced byLPS in 30 min of incubation (Fig. 6b). This resultsuggests that STP is able to reverse a necessary step inthe pathway leading to NF-κB activation. To assess ifthe effect of STP on IκB-α phosphorylation was indeedrelated to p65 nuclear translocation, we isolated thecytosolic and nuclear fractions after 4-h treatments andevaluated the immunocontent of p65 by Western blot in
Fig. 3 Redox parameters and morphological changes affected byLPS. RAW 264.7 cells were co-incubated for 24 h with STP (1 μg/mL) and LPS (1 μg/mL). NO production was assessed by quantifi-cation of nitrite accumulation in the incubationmediumby themethodofGriess (a). Total reduced thiol content (b) and SODactivity (c)wereassessed as described in “Material and methods” section. Contrast-phase microscopy (e, ×100 magnification) was applied to visualize
morphological changes characteristic of macrophage activation(black arrows indicate cells presenting altered morphology).One-way ANOVA with Tukey’s post hoc test was applied in (a),(b), (c), and (d). Asterisks denote significance level in relation tocontrol (*p<0.05, **p<0.01). Number sign different from LPSgroup (p<0.05). Bars represent mean ± SEM of replicate samplesfrom three independent experiments (n=8 in each experiment)
182 Cell Biol Toxicol (2013) 29:175–187

both fractions. STP inhibited the increase of p65 in thenucleus induced by LPS (Fig. 6c), indicating itprevented NF-κB activation. In the cytosolic fraction,
however, STP did not restore the immunocontent of p65(Fig. 6d). It is possible that this effect may be the resultof increased p65 proteasomic degradation or othermechanism related to increased protein turnover.
Discussion
Tryptamines represent a wide group of biomoleculesand synthetic derivatives with a broad spectrum ofpharmacological actions. We evaluated here the effectof N-salicyloyltryptamine, a new anticonvulsant com-pound previously reported to modulate voltage-dependent Na+, Ca2+, and K+ channels and exertantinociceptive activity (Araujo et al. 2003; Quintanset al. 2010). Seizures and pain involve a multitude ofmolecular processes at cellular and systemic levels;inflammatory and nociceptive pathways involve sev-eral common mechanisms, and inflammatory markersare greatly enhanced after epileptic episodes (Silveiraet al. 2012). For this reason, we studied the modula-tion of some cellular parameters of inflammation bySTP.
Currently, many tryptamines are used as powerfulanti-migraine agents, such as naratriptan, sumatriptan,and almotriptan (Yu 2008). The development of cen-tral analgesic drugs which act by non-opioid mecha-nisms has been one of the noticeable research fields inthe past few years. STP was developed as an anticon-vulsant agent; nonetheless, most antiepileptic drugscurrently used in therapy of epilepsy exhibit analgesiceffect and are used in the treatment of chronic neuro-pathic pain (trigeminal pain) (Silveira et al. 2012).
Fig. 4 Effect of STP on cytokine release in LPS-stimulated cells.a RAW 264.7 cells were incubated with LPS 1 μg/mL in thepresence of STP at 1 μg/mL for 24 h and TNF-α (a) and IL-1β(b) release to the incubation medium were assessed by ELISA asdescribed in “Material and methods”. Bars represent mean ± SEMof replicate samples from three independent experiments (n=8 ineach experiment). One-way ANOVA with Tukey’s post hoc testwas applied. Asterisks different from control (p<0.001); numbersign different from LPS group (p<0.05)
Fig. 5 Effect of STP on CD40, TNF-α, and RAGEimmunocontent. RAW 264.7 cells were co-incubated for 24 hwith LPS (1 μg/mL) and STP (1 μg/mL). Cell lysates (30 μgproteins) were subjected to SDS-PAGE and immunoblot analy-sis was performed with anti-CD40 using β-actin as a loadingcontrol (a). Representative immunoblot and respective mean ±
SEM values are depicted. The same analysis was applied toverify TNF-α (b) and (c) RAGE immunocontent. One-wayANOVA with Tukey’s post hoc test was applied. Asterisksdifferent from control (p<0.0001). Number sign different fromLPS group (p<0.05). Experiments were repeated three times (n=3in each experiment)
Cell Biol Toxicol (2013) 29:175–187 183
Oxidative stress and reactive species formation havealso been extensively implicated in acute and chronicinflammation and pain (Salvemini et al. 2006). Inmany chronic conditions associated with pain,proinflammatory cytokines and prostaglandins are re-leased at the sites of pain along with reactive speciesand NO (Halliwell and Gutteridge 2007).
We observed here that STP administration to cellculture medium at doses ranging from 0.001 to1 μg/mL does not affect macrophage cell viability.This result suggests that STP administrations leadingto extracellular fluid concentrations above this limitmight be avoided. It is possible that the cytotoxic
action by high concentrations of STP is related tomodulate reactive species production, as theDCFH-DA assay showed that STP at 50 μg/mLenhanced RAW 264.7 free radical production. STPat 0.001 to 1 μg/mL had no protective effectagainst the H2O2 challenge to which RAW 264.7cells were submitted. Importantly, at these concen-trations, STP alone also did not alter intracellularreactive species production. In agreement with thisdata, NO production at both basal and under LPStreatment were not modulated by STP, as nitriteaccumulation in the incubation media was notsignificantly changed in both conditions.
Fig. 6 ERK1/2 and IκBα phosphorylation and nuclear translo-cation of p65 in RAW 264.7 treated with STP. Cells were pre-incubated with STP during 1 h and then LPS was added. After30 min, cellular proteins were extracted and the phosphorylationstate of ERK 1/2 and IκBα were assessed by Western blot.Representative immunoblots showing detection of phosphory-lated (pERK 1/2, upper gel panel) and total isoforms of ERK 1/2(lower gel panel) are depicted in (a). Representative immuno-blots showing detection of phosphorylated (pIκBα, upper gelpanel) and total isoforms of IκBα (lower panel) are depicted in(b). Gel densitometry mean values are shown below respective
gels. Translocation of p65 from cytosolic to nuclear fraction inRAW 264.7 cells treated with STP 1 μg/mL with or withoutLPS 1 μg/mL for 4 h was assessed by Western blot analysis ofthe p65 content in nuclear (c) and cytosolic (d) fractions. Theimmunocontent of p65 is quantified relative to lamin B in thenucleus and relative to β-actin in the cytosol. One-way ANOVAwith Tukey’s post hoc test was applied. Asterisks denote signif-icance level in relation to control (*p<0.001). Number signdifferent from LPS group (p<0.05). Bars represent mean ±SEM of replicate samples from three independent experiments(n=3 in each experiment)
184 Cell Biol Toxicol (2013) 29:175–187

However, we observed that STP reversed someredox-associated modifications induced by LPS inmacrophages. The decrease in free sulfhydryl groupsis related to the increased thiol oxidation resultingfrom macrophage activation by LPS. This effect wasreversed by co-incubation with STP 1 μg/mL. In thesame context, the increase in the activity of SOD,which is observed in cells treated with LPS and in-dicates an increase in the cellular production of super-oxide radicals, was also significantly inhibited by STP.Altogether, these results suggest that although STPdoes not significantly alter parameters related to freeradical generation and oxidative stress in basal condi-tions and when cells are challenged with a pro-oxidantinsult (H2O2), STP is able to reverse some of the redoxparameters altered by LPS stimulation in RAW 264.7macrophages.
TNF-α release by macrophages is a well-knownparameter of pro-inflammatory activation. TNF-αbinds to TNF receptor 1 (TNFR1, also known asp55; TNFRSF1a), present on most cell types, and toTNFR2 (also known as p75; TNFRSF1b), expressedmainly by immune system cells (Vendramini-Costaand Carvalho 2012). In response to acute or chronicinfection, TNF-α is released by activated macro-phages and triggers pro-inflammatory cascades in oth-er cells, enhancing reactive species production and
cytokine expression and release. TNF-α acts synergis-tically with other cytokines such as IFNγ and IL-17,propagating the pro-inflammatory state (Vendramini-Costa and Carvalho 2012). We observed here that STPsignificantly inhibited part of TNF-α release to theextracellular environment caused by LPS stimulation,indicating a clear anti-inflammatory effect at cellularlevel. Besides, LPS-induced IL-1β release wascompletely inhibited by STP. The release of IL-1βby macrophages elicits inflammation and pain-relatedresponses in different cell types. The expression ofcyclooxygenase-2 is induced by IL-1β in CNS cells,which contributes to inflammatory pain hypersensitiv-ity (Neeb et al. 2011). Thus, it is possible that theantinociceptive effect by STP observed previously(Quintans et al. 2010) may be related to IL-1βinhibition.
NF-κB activation by LPS, TNF-α, and other pro-inflammatory mediators involve the phosphorylationand subsequent degradation of IκBα, which iscomplexed with the transcriptional subunit (p65) ofNF-κB (Bowie and O’Neill 2000). Multiple kinasesignaling triggered by TNF-α receptor activation leadsto IκBα phosphorylation, resulting in transcriptionalactivation of several pro-inflammatory genes, such asIL-6, IL-1β, and TNF-α itself. Another pro-inflammatory effect of LPS is CD40 up-regulation,which is hindered by co-treatment with STP. Our re-sults confirm that anti-inflammatory properties of STPextend to intracellular signaling, as IκBα phosphory-lation and downstream effects of NF-kB activation(TNF-α and IL-1β release) induced by LPS wereinhibited by co-treatment with STP (Fig. 7).
The functional expression of CD40 by macro-phages confers upon them the ability to play a signif-icant role in the outcome of adaptive immuneresponses, suggesting that macrophages are excellenttargets in immunological therapies. For example,macrophage-targeted blockade of CD40 signalingcould provide benefits in ongoing autoimmune inflam-matory disease (Suttles and Stout 2009). The inhibition ofLPS-induced suggests that this compound is able to re-vert, at least in part, some of the pro-inflammatory effectscaused by LPS in macrophages. Anti-inflammatory ef-fects by endogenous or synthetic tryptamines are notuncommon. Melatonin, produced by many vertebrates,was observed to prevent inflammatory stimuli by LPS inmacrophages (Mayo et al. 2005). In the same work,however, melatonin induced a strong antioxidant effect
Fig. 7 A schematic diagram proposing the action of STP in signalpathways affected by LPS. LPS triggers ERK1/2and IκB phos-phorylation. Activation of ERK 1/2 and IκB resulted in activationof the transcription factor NF-kB. Activated NF-kB (p65) migratesto the cell nucleus and results in TNF-α and IL-1β release
Cell Biol Toxicol (2013) 29:175–187 185

in macrophages and caused apoptosis. Here, at the dosesfound to decrease TNF-α production without affectingcell viability, STP had no apparent effects on cellular freeradical production. STP was suggested to exert itsantinociceptive action through blocking of voltage-gatedsodium channels and a mechanism not involving theopioid system, for these reasons being indicated to treatpain disorders (Quintans et al. 2010).
Acknowledgments This work was funded by grants from theBrazilian governmental agencies FAPERGS (PqG 12/0998-0,ARD 11/1893-7, PRONEX 1000274) CAPES (PROCAD 066/2007), CNPq, IBN-Net (#01.06.0842-00), INCT-EN, andPROPESQ-UFRGS.
References
Abramson SB. Nitric oxide in inflammation and pain associatedwith osteoarthritis. Arthritis Res Ther. 2008;10 Suppl 2:S2.
Araujo DA et al. N-salicyloyltryptamine, a new anticonvulsantdrug, acts on voltage-dependent Na+, Ca2+, and K+ ionchannels. Br J Pharmacol. 2003;140:1331–9.
Bopp C, Bierhaus A, Hofer S, Bouchon A, Nawroth PP, MartinE, et al. Bench-to-bedside review: the inflammation-perpetuating pattern-recognition receptor RAGE as a ther-apeutic target in sepsis. Crit Care. 2008;12:201.
Bowie A, O’Neill LA. Oxidative stress and nuclear factor-kappaB activation: a reassessment of the evidence inthe light of recent discoveries. Biochem Pharmacol.2000;59:13–23.
Cuzzocrea S, Zingarelli B, Gilad E, Hake P, Salzman AL, SzaboC. Protective effect of melatonin in carrageenan-inducedmodels of local inflammation: relationship to its inhibitoryeffect on nitric oxide production and its peroxynitrite scav-enging activity. J Pineal Res. 1997;23:106–16.
DeBittencourt PasqualiMA et al. VitaminA (retinol) downregulatesthe receptor for advanced glycation endproducts (RAGE)by oxidant-dependent activation of p38 MAPK and NF-kBin human lung cancer A549 cells. Cell Signal.2013;25:939–54.
de Souza LF, Gelain DP, Jardim FR, Ribeiro GR, Zim M,Bernard EA. Extracellular inosine participates in tumornecrosis factor-alpha induced nitric oxide production incultured Sertoli cells. Mol Cell Biochem. 2006;281:123–8.
Deneris ES, Wyler SC. Serotonergic transcriptional networksand potential importance to mental health. Nat Neurosci.2012;15:519–27.
Dieudonne S. Serotonergic neuromodulation in the cerebellarcortex: cellular, synaptic, and molecular basis. Neuroscien-tist. 2001;7:207–19.
Ellman GL. Tissue sulfhydryl groups. Arch Biochem Biophys.1959;82:70–7.
Evans MS, Cheng X, Jeffry JA, Disney KE, Premkumar LS.Sumatriptan inhibits TRPV1 channels in trigeminal neu-rons. Headache. 2012;52:773–84.
Foey AD, Feldmann M, Brennan FM. CD40 ligation inducesmacrophage IL-10 and TNF-alpha production: differentialuse of the PI3K and p42/44 MAPK-pathways. Cytokine.2001;16:131–42.
Gelain DP, De Bittencourt Pasquali MA, Caregnato FF, Zanotto-Filho A, Moreira JC. Retinol up-regulates the receptorfor advanced glycation endproducts (RAGE) by in-creasing intracellular reactive species. Toxicol In Vitro.2008;22:1123–7.
Gelain DP, Moreira JC. Evidence of increased reactive speciesformation by retinol, but not retinoic acid, in PC12 cells.Toxicol In Vitro. 2008;22:553–8.
Halliwell B, Gutteridge JMC. Free radicals in biology andmedicine. Oxford: Oxford University Press; 2007.
Jhee SS, Shiovitz T, Crawford AW, Cutler NR. Pharmacokineticsand pharmacodynamics of the triptan antimigraine agents:a comparative review. Clin Pharmacokinet. 2001;40:189–205.
Jones RS. Tryptamine: a neuromodulator or neurotransmitter inmammalian brain? Prog Neurobiol. 1982;19:117–39.
Kang EH, Gebru E, Kim MH, Cheng H, Park SC. EstA protein,a novel virulence factor of Streptococcus pneumoniae, in-duces nitric oxide and pro-inflammatory cytokine produc-tion in RAW 264.7 macrophages through NF-kappaB/MAPK. Microb Pathog. 2009;47:196–201.
Kenneth NS, Mudie S, Naron S, Rocha S. TFR1 interacts withthe IKK complex and is involved in IKK-NF-kappaB sig-nalling. Biochem J. 2013;449:275–84.
Lin QQ, Yan CF, Lin R, Zhang JY, Wang WR, Yang LN, et al.SIRT1 regulates TNF-alpha-induced expression of CD40in 3 T3-L1 adipocytes via NF-kappaB pathway. Cytokine.2012;60:447–55.
Maczurek A, ShanmugamK,Munch G. Inflammation and the redox-sensitive AGE-RAGE pathway as a therapeutic target inAlzheimer’s disease. Ann N YAcad Sci. 2008;1126:147–51.
Mayo JC, Sainz RM, Tan DX, Hardeland R, Leon J, RodriguezC, et al. Anti-inflammatory actions of melatonin and itsmetabolites, N1-acetyl-N2-formyl-5-methoxykynuramine(AFMK) and N1-acetyl-5-methoxykynuramine (AMK), inmacrophages. J Neuroimmunol. 2005;165:139–49.
Misra HP, Fridovich I. The role of superoxide anion in theautoxidation of epinephrine and a simple assay for super-oxide dismutase. J Biol Chem. 1972;247:3170–5.
Missbach M, Jagher B, Sigg I, Nayeri S, Carlberg C,Wiesenberg I. Thiazolidine diones, specific ligands of thenuclear receptor retinoid Z receptor/retinoid acid receptor-related orphan receptor alpha with potent antiarthritic ac-tivity. J Biol Chem. 1996;271:13515–22.
Nava F, Calapai G, Facciola G, Cuzzocrea S, Giuliani G, deSarro A, et al. Melatonin effects on inhibition of thirst andfever induced by lipopolysaccharide in rat. Eur JPharmacol. 1997;331:267–74.
Neeb L, Hellen P, Boehnke C, Hoffmann J, Schuh-Hofer S,Dirnagl U, et al. IL-1beta stimulates COX-2 dependentPGE(2) synthesis and CGRP release in rat trigeminalganglia cells. PLoS One. 2011;6:e17360.
Nosal R, Perecko T, Jancinova V, Drabikova K, Harmatha J,Svitekova K. Suppression of oxidative burst in humanneutrophils with the naturally occurring serotonin deriva-tive isomer from Leuzea carthamoides. Neuro EndocrinolLett. 2010;31 Suppl 2:69–72.
186 Cell Biol Toxicol (2013) 29:175–187

Oliveira FA, de Almeida RN, Sousa MF, Barbosa-Filho JM,Diniz SA, de Medeiros IA. Anticonvulsant properties ofN-salicyloyltryptamine in mice. Pharmacol BiochemBehav. 2001;68:199–202.
Quintans LJ et al. Bioassay-guided evaluation of antinociceptive effectof N-salicyloyltryptamine: a behavioral and electrophysiologicalapproach. J Biomed Biotechnol. 2010;2010:230745.
Raghavendra V, Agrewala JN, Kulkarni SK. Role of centrallyadministered melatonin and inhibitors of COX and NOS inLPS-induced hyperthermia and adipsia. ProstaglandinsLeukot Essent Fatty Acids. 1999;60:249–53.
Raghavendra V, Agrewala JN, Kulkarni SK. Melatonin reversalof lipopolysacharides-induced thermal and behavioralhyperalgesia in mice. Eur J Pharmacol. 2000;395:15–21.
Rehman MU, Yoshihisa Y, Miyamoto Y, Shimizu T. The anti-inflammatory effects of platinum nanoparticles on thelipopolysaccharide-induced inflammatory response in RAW264.7 macrophages. Inflamm Res. 2012;61:1177–85.
Sacco S, Aquilini L, Ghezzi P, Pinza M, Guglielmotti A. Mech-anism of the inhibitory effect of melatonin on tumor ne-crosis factor production in vivo and in vitro. Eur JPharmacol. 1998;343:249–55.
Salvemini D, Doyle TM, Cuzzocrea S. Superoxide, peroxynitriteand oxidative/nitrative stress in inflammation. Biochem SocTrans. 2006;34:965–70.
Scott MJ, Hoth JJ, Stagner MK, Gardner SA, Peyton JC,Cheadle WG. CD40–CD154 interactions between macro-phages and natural killer cells during sepsis are critical formacrophage activation and are not interferon gamma de-pendent. Clin Exp Immunol. 2004;137:469–77.
Shih MF, Chen LY, Tsai PJ, Cherng JY. In vitro and in vivotherapeutics of beta-thujaplicin on LPS-induced inflamma-tion in macrophages and septic shock in mice. Int JImmunopathol Pharmacol. 2012;25:39–48.
Silveira G, de Oliveira AC, Teixeira AL. Insights into inflam-mation and epilepsy from the basic and clinical sciences. JClin Neurosci. 2012;19:1071–5.
Sims GP, Rowe DC, Rietdijk ST, Herbst R, Coyle AJ. HMGB1and RAGE in inflammation and cancer. Annu RevImmunol. 2010;28:367–88.
Skehan P, Storeng R, Scudiero D, Monks A, McMahon J,Vistica D, et al. New colorimetric cytotoxicity assay foranticancer-drug screening. J Natl Cancer Inst. 1990;82:1107–12.
Suttles J, Stout RD. Macrophage CD40 signaling: a pivotalregulator of disease protection and pathogenesis. SeminImmunol. 2009;21:257–64.
Vendramini-Costa DB, Carvalho JE. Molecular Link Mecha-nisms between Inflammation and Cancer. Curr PharmDes. 2012;18:3831–52.
Wang Z, Jiang W, Zhang Z, Qian M, Du B. Nitidine chlorideinhibits LPS-induced inflammatory cytokines productionvia MAPK and NF-kappaB pathway in RAW 264.7 cells.J Ethnopharmacol. 2012;144:145–50.
Yang L, Zhang J, Yan C, Zhou J, Lin R, Lin Q, et al. SIRT1regulates CD40 expression induced by TNF-alpha via NF-kB pathway in endothelial cells. Cell Physiol Biochem.2012;30:1287–98.
Yu AM. Indolealkylamines: biotransformations and potentialdrug–drug interactions. AAPS J. 2008;10:242–53.
Cell Biol Toxicol (2013) 29:175–187 187





























