Embed Size (px)
Citation preview

sapsoeSaptmP
Available online at www.sciencedirect.com
Theriogenology 76 (2011) 342–350
0d
Effects of antioxidants and duration of pre-freezing equilibrationon frozen-thawed ram semen
D.R. Câmaraa,b, S.V. Silvab, F.C. Almeidab, J.F. Nunesc, M.M.P. Guerrab,*a Department of Veterinary Medicine, Federal University of Alagoas; Fazenda São Luiz, s/n. Zona Rural de Viçosa. Viçosa-AL. Brazil. Post-
Graduate Student, Northeast Biotechnology Networkb Laboratory of Andrology, Department of Veterinary Medicine, Federal Rural University of Pernambuco; R. Dom Manoel de Medeiros, s/n,
Dois Irmãos, Recife-PE, Brazil CEP 52171-900c Laboratory of Sheep and Goat Semen Technology, Ceará State University; Av. Paranjana, 1700 - Campus do Itaperi, Fortaleza-CE, Brazil
Received 25 August 2010; received in revised form 5 January 2011; accepted 6 January 2011
Abstract
The objective was to evaluate the effects of various antioxidants and duration of pre-freezing equilibration on cryopreservationof ram semen. Semen samples from four rams were pooled, diluted with Tris-egg yolk extender without antioxidants (control),or supplemented with reduced glutathione (GSH: 0.5, 1.0, and 2.0 mM), superoxide dismutase (SOD: 5, 10, and 20 U/mL), orcatalase (CAT: 5, 10, and 20 U/mL), and cryopreserved, immediately after thermal equilibrium was reached at 5 °C (0 h), or 12or 24 h after equilibration. Total antioxidant capacity was determined in the in natura extenders and after addition of semenamples for various durations of processing (fresh/dilute, throughout refrigeration, and post-thaw). Plasma membrane (PI-CFDA),crosome integrity (FITC-PNA), and mitochondrial membrane potential (JC-1) were determined in fresh/diluted, refrigerated, andost-thaw samples. Post-thaw sperm motility was assessed with a computerized analysis system (CASA). There were noignificant differences in acrosome damage or mitochondrial membrane potential after refrigeration and freeze-thaw, regardlessf antioxidant addition. Sperm plasma membrane integrity was worse (P � 0.05) with cryopreservation immediately afterquilibration (average 20.1 � 8.3; mean � SD) than after 12 h of equilibration (average 42.5 � 10.9); however, the addition ofOD and CAT (10 and 20 U/mL) resulted in no significant difference between post-equilibration intervals of 0 and 12 h. Totalntioxidant activity was not different (P � 0.05) among treatments after sperm addition or throughout the refrigeration andost-thaw. In conclusion, adding GSH, SOD or CAT did not increase the total antioxidant capacity of semen, nor did it enhancehe quality of the post-thaw sperm. However, maintenance of ram semen at 5 °C for 12 h prior to cryopreservation reducedembrane damage of frozen-thawed sperm.ublished by Elsevier Inc.
Keywords: Antioxidant; Equilibration time; Semen cryopreservation; Viability
www.theriojournal.com
1. IntroductionIn 1970, Lightfoot and Salamon [1] demonstrated
that the number of sperm recovered from the uterus ofan artificially inseminated sheep was lower after the useof frozen than fresh semen, regardless of sperm con-
* Corresponding author. Tel.: �055 81 3320 6412; fax: � 055 813320 6057.
E-mail address: [email protected] (M.M.P. Guerra).
093-691X/$ – see front matter Published by Elsevier Inc.oi:10.1016/j.theriogenology.2011.02.013
centration. Cryopreservation causes extensive chemicaland physical damages to sperm membranes, which areattributed to alterations in the transition from the lipidphase, increases in lipid peroxidation of the membraneinduced by reactive oxygen species (ROS), and me-chanical stress on cell membranes due to osmotic stressand temperature changes [2,3].
It is well documented that ROS, formed by the
univalent reduction of oxygen [4,5], are responsible for
ac
(tt
2
tEw((afismFcrOaCs°°sreab
2a
smaccwee
2p
fay
343D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
maintenance of physiological processes in sperm, e.g.,capacitation and the acrosome reaction [6,7]. However,large quantities of ROS decreased the viability of cryo-preserved sperm, giving rise to lipid peroxidation dueto the large amount of polyunsaturated fatty acids pres-ent in the sperm plasma membrane [8]. Approaches toincrease the percentage of viable post-thaw cells haveincluded various extenders [9], refrigeration times[10,11], dilution rates [12], glycerol concentrations[13], and equilibration times [14–16].
Equilibration time is the interval between addition ofglycerol and freezing sperm; it is expected to be ben-eficial, as it allows a better osmotic balance after sperminteract with a cryoprotectant. Concurrently, it also hasrelevance for semen collection and processing, due totime needed prior to cryopreservation [17]. Studiescarried out since the late 1960s [18] and more recently[19] reported the beneficial effects of equilibration be-fore cryopreserving bovine sperm. Although equilibra-tion of ram semen for 24 h did not improve fertility[15], the value of an intermediate equilibration interval(12 h) for cryopreserving ram sperm has apparently notbeen reported. The objective of the present study was toassess the effects of various antioxidant substances andpre-freezing equilibration intervals on the quality ofram sperm during cryopreservation.
2. Materials and methods
2.1. Chemicals
The antioxidants (GSH, SOD and CAT) and otherchemicals used in this study were obtained from SigmaAldrich Chemicals (St. Louis, MO, USA).
2.2. Rams and semen collection
Four mixed-breed Santa Inês rams, 1 to 2 y old andraised in a confinement system with natural light(8.0314 S, 34.5252 W), were used. They had ad libitumaccess to hay and good quality water, and were supple-mented with 500 g/head/d of concentrate. The ramswere previously deemed breeding sound and were inroutine semen collection (artificial vagina and a teaserfemale) thrice weekly. Four semen samples were col-lected per ram (total of 16 samples); samples from allfour rams were pooled to eliminate individual differ-ences [20]. Minimum requirements set to freeze ejac-ulates pooled were volume �3.0 mL, motility �80%,and a minimum of 3 � 109 sperm/mL. Motility wasssessed subjectively (�400) with phase-contrast mi-
roscopy (Olympus, Tokyo, Japan). Semen was diluted t1:200) in formol citrate solution, and sperm concen-ration was calculated using a Neubauer hemocytome-er chamber [11], with bright-field microscopy (�400).
.3. Extenders and semen processing
Tris-egg yolk extender was used as the base ex-ender medium, with 6% glycerol and 10% egg yolk.ach pool was split into 10 equal aliquots and dilutedith base extender containing reduced glutathione
GSH: 0.5, 1.0, and 2.0 mM), superoxide dismutaseSOD: 5, 10, and 20 U/mL) and catalase (CAT: 5, 10nd 20 U/mL), and no antioxidants (control), with anal concentration of 100 � 106 sperm/mL. Extendedemen samples were then manually packed into 0.25L mini straws (IMV
®
Technologies, L’Aigle, Cedex,rance) at room temperature, and were immediatelyooled (average of � 0.2 °C/min) to a thermal equilib-ium of 5 °C, which occurred in approximately 90 min.ne-third of the doses of each treatment was placed inprogrammable freezer (TK-3000, TK Tecnologia emonservação Ltda., Uberaba, MG, Brazil), previously
tabilized at 5 °C. Semen was cooled from 5 to �120C at a rate of 12.5 °C/min. Once straws reached �120C, they were plunged directly into liquid nitrogen andtored until subsequent evaluation. The procedure wasepeated with remaining samples after 12 and 24 h ofquilibration. After a minimum of 15 d of frozen stor-ge, straws were thawed at 37 °C for 30 s in a waterath and evaluated.
.4. Subjective assessment of motility of fresh/dilutednd refrigerated sperm
The percentage of motile cells was subjectively as-essed by a visual estimation using a phase contrasticroscope (�400; Olympus, Tokyo, Japan) immedi-
tely after dilution of the semen in the Tris-egg yolkontrol and Tris-egg yolk, supplemented with variousoncentrations of antioxidants (fresh/diluted). Motilityas assessed when the samples had reached thermal
quilibrium at 5 °C (0 h), and after 12 and 24 h ofquilibration.
.5. Assessment of sperm viability using fluorescentrobes
Sperm viability analyses were performed in the pre-reezing periods (fresh/diluted, 0, 12, and 24 h) andfter thawing in the Tris-egg yolk control and Tris-eggolk, supplemented with various concentrations of an-
ioxidants.
ii
cmflG5wd
ah(
2
c5sgSSpmesaCl
sa7a
344 D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
2.5.1. Membrane integrity
The integrity of the sperm membrane was deter-mined using a combination of propidium iodide (PI)and carboxyfluorescein diacetate (CFDA), as describedby Harrison and Vickers [21] and modified by Coleto etal [22]. Aliquots (50 �L) of each sample were dilutedn 150 �L of Tris containing 20 �L of PI (0.5 mg/mLn PBS) and 5 �L of CFDA (0.46 mg/mL in DMSO).
Using DBP 485/20 nm excitation and DBP 580-630 nmemission filters, 200 cells from each sample were ex-amined under an epifluorescence microscope (�400;Carl Zeiss, Göttingen, Germany). Green fluorescencewas interpreted as an intact membrane, whereas redindicated a damaged membrane.
2.5.2. Acrosomal integrity
For detection of acrosomal integrity, sperm werestained with fluorescein isothiocyanate conjugated topeanut agglutinin (FITC-PNA), as described [23]. Ali-quots (5 �L) of semen from each treatment were placedon microscope slides and air dried. The slides had 20�L of FITC-PNA working solution (100 �g/mL)spread over them, and were incubated at 4 °C in amoisture chamber for 15 to 20 min (in the dark). Theslides were then immersed in PBS at 4 °C twice anddried naturally in the absence of light. At the time ofevaluation, 5 �L of solution containing 4.5 mL ofglycerol, 0.5 mL of PBS and 5.0 mg of phenylenedi-amine was placed on the slide. Then the slide wascovered with a slip cover and subjected to epifluores-cence analysis (�1000; Carl Zeiss, Göttingen, Ger-many) using BP 450–490 nm excitation and LP 515nm emission filters. Among the 200 cells examined,sperm were classified as having an intact acrosome(iAC) when the acrosome region was stained fluores-cent green, and as having a reacted acrosome when thefluorescent green was absent from the head region, orwhen it was present in the equatorial region of thesperm head.
2.5.3. Mitochondrial membrane potential
Aliquots (50 �L) of semen from each sample werediluted in 150 �L of Tris containing 5 �L of lipophilicationic JC-1 (0.15 mM in DMSO), incubated for 10in, fixed with glutaraldehyde, and subjected to epi-uorescence microscopy (�400; Carl Zeiss, Göttingen,ermany) using BP 450–490 nm excitation and LP15 nm emission filters. From each sample, 200 cellsere examined and classified as having high mitochon-
rial membrane potential (H�m) when emitting or-nge fluorescence in the region of the midpiece, and asaving low mitochondrial membrane potentialL�m) when emitting green fluorescence.
.6. Computer Assisted Sperm Analysis (CASA)
The CASA system consisted of an optical phase-ontrast microscopy system (Nikon™ H5505, Eclipse0i, Japan) with stroboscopic illumination, a warmingtage (37 °C), video camera (Basler Vision Tecnolo-ie™ A312FC, Ahrensburg, Germany), and a PC withperm Class Analyzer (SCA™) software (Microptics,.L., Version 3.2.0, Barcelona, Spain). Motility endoints were assessed from two thawed straws/treat-ent/equilibration time, previously pooled and homog-
nized in a 1.0 mL microcentrifuge tube, diluted inodium citrate solution at 2.94% (v/v), and incubated inwater bath at 37 °C for 5 min. A pre-warmed Maklerhamber® (Sefi Medical Instrument, Haifa, Israel) was
oaded with 5 �L of diluted sample; at least four non-consecutive, randomly selected microscopic fields persample were scanned, recording at least 400 motilesperm. Events not related to sperm were removed, andimage sequences were saved and analyzed later. Thefollowing end points were analyzed: total motility(TM), progressive motility (PM), path velocity (VAP),progressive velocity (VSL), curvilinear velocity(VCL), amplitude of lateral head displacement (ALH),beat frequency of the tail (BCF), straightness (STR),and linearity (LIN). Motility end points were measuredwith the following settings: temperature 37 °C; framesacquired, 25; frame rate, 25 s; minimal contrast, 75;frame number, 25 per field; sperm velocity that can beanalyzed, 0 to 180 �m/s; and threshold STR, 75%.
2.7. Total antioxidant capacity
Total antioxidant capacity (TAC) was determinedthrough spectrophotometry of extenders in natura(without the addition of semen), as well as fresh/dilutedsemen samples, after 0, 12, and 24 h of equilibration at5 °C, and after thawing. The content of two straws/treatment/equilibration time in refrigerated and thawedsamples was pooled and well mixed in a 1.0 mL mi-crocentrifuge tube before analysis. For this, the sampleswere subjected to centrifugation at 10,000 � g for 15min. Then 5 �L of the supernatant of each semenample was placed in each well of the reading plate,long with 10 �L of myoglobin working solution and5 �L of ABTS working solution, and were incubatedt room temperature for 5 min. Then 50 �L of stop
solution was added. All readings were performed
within 30 min after finalizing the preparation.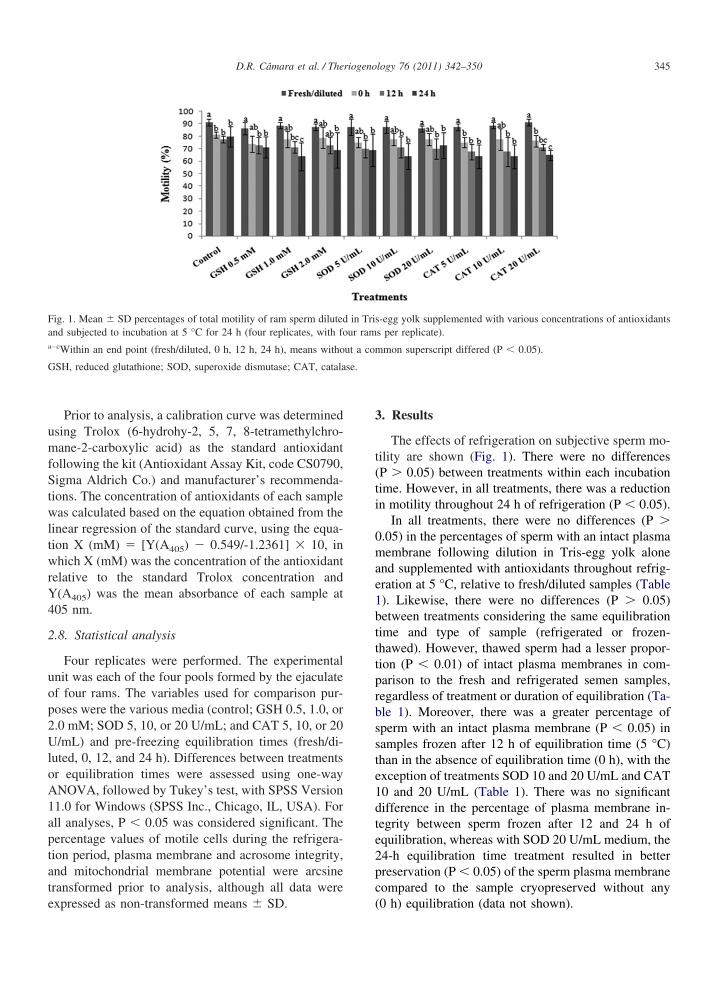
c
se.
345D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
Prior to analysis, a calibration curve was determinedusing Trolox (6-hydrohy-2, 5, 7, 8-tetramethylchro-mane-2-carboxylic acid) as the standard antioxidantfollowing the kit (Antioxidant Assay Kit, code CS0790,Sigma Aldrich Co.) and manufacturer’s recommenda-tions. The concentration of antioxidants of each samplewas calculated based on the equation obtained from thelinear regression of the standard curve, using the equa-tion X (mM) [Y(A405) � 0.549/-1.2361] � 10, inwhich X (mM) was the concentration of the antioxidantrelative to the standard Trolox concentration andY(A405) was the mean absorbance of each sample at405 nm.
2.8. Statistical analysis
Four replicates were performed. The experimentalunit was each of the four pools formed by the ejaculateof four rams. The variables used for comparison pur-poses were the various media (control; GSH 0.5, 1.0, or2.0 mM; SOD 5, 10, or 20 U/mL; and CAT 5, 10, or 20U/mL) and pre-freezing equilibration times (fresh/di-luted, 0, 12, and 24 h). Differences between treatmentsor equilibration times were assessed using one-wayANOVA, followed by Tukey’s test, with SPSS Version11.0 for Windows (SPSS Inc., Chicago, IL, USA). Forall analyses, P � 0.05 was considered significant. Thepercentage values of motile cells during the refrigera-tion period, plasma membrane and acrosome integrity,and mitochondrial membrane potential were arcsinetransformed prior to analysis, although all data were
Fig. 1. Mean � SD percentages of total motility of ram sperm dilutedand subjected to incubation at 5 °C for 24 h (four replicates, with foa–cWithin an end point (fresh/diluted, 0 h, 12 h, 24 h), means witho
GSH, reduced glutathione; SOD, superoxide dismutase; CAT, catala
expressed as non-transformed means � SD. (
3. Results
The effects of refrigeration on subjective sperm mo-tility are shown (Fig. 1). There were no differences(P � 0.05) between treatments within each incubationtime. However, in all treatments, there was a reductionin motility throughout 24 h of refrigeration (P � 0.05).
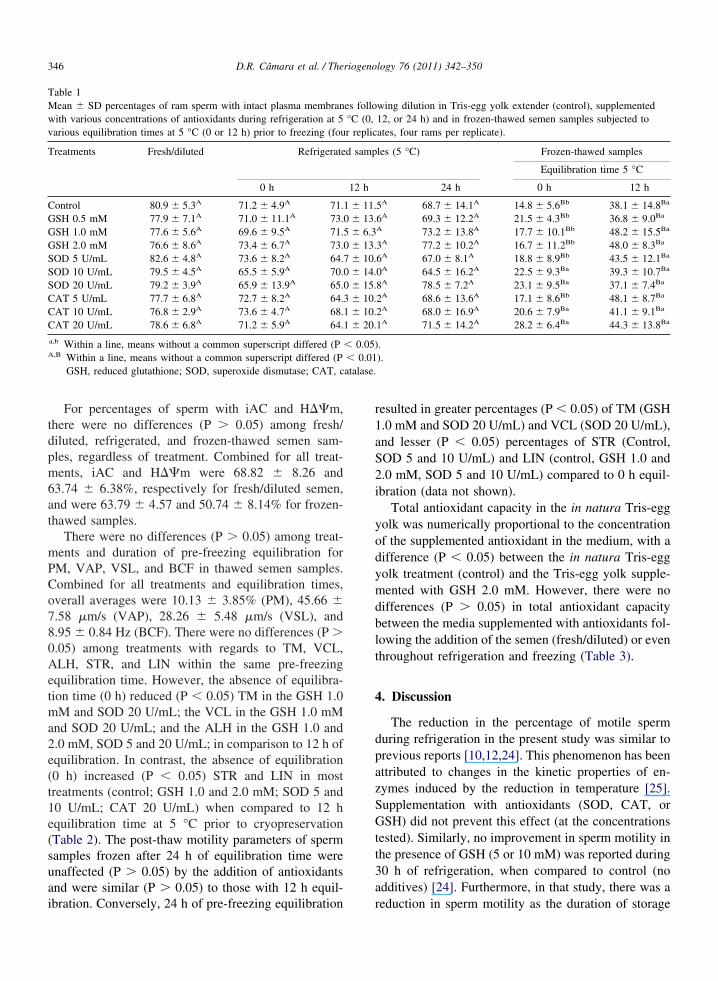
In all treatments, there were no differences (P �0.05) in the percentages of sperm with an intact plasmamembrane following dilution in Tris-egg yolk aloneand supplemented with antioxidants throughout refrig-eration at 5 °C, relative to fresh/diluted samples (Table1). Likewise, there were no differences (P � 0.05)between treatments considering the same equilibrationtime and type of sample (refrigerated or frozen-thawed). However, thawed sperm had a lesser propor-tion (P � 0.01) of intact plasma membranes in com-parison to the fresh and refrigerated semen samples,regardless of treatment or duration of equilibration (Ta-ble 1). Moreover, there was a greater percentage ofsperm with an intact plasma membrane (P � 0.05) insamples frozen after 12 h of equilibration time (5 °C)than in the absence of equilibration time (0 h), with theexception of treatments SOD 10 and 20 U/mL and CAT10 and 20 U/mL (Table 1). There was no significantdifference in the percentage of plasma membrane in-tegrity between sperm frozen after 12 and 24 h ofequilibration, whereas with SOD 20 U/mL medium, the24-h equilibration time treatment resulted in betterpreservation (P � 0.05) of the sperm plasma membraneompared to the sample cryopreserved without any
s-egg yolk supplemented with various concentrations of antioxidantss per replicate).
mon superscript differed (P � 0.05).
in Triur ram
ut a com
0 h) equilibration (data not shown).
80Aetma2e(t1e(suai
r1aS2i
yodymdblt
4
dpazSGtt3a
atalase.
346 D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
For percentages of sperm with iAC and H�m,there were no differences (P � 0.05) among fresh/diluted, refrigerated, and frozen-thawed semen sam-ples, regardless of treatment. Combined for all treat-ments, iAC and H�m were 68.82 � 8.26 and63.74 � 6.38%, respectively for fresh/diluted semen,and were 63.79 � 4.57 and 50.74 � 8.14% for frozen-thawed samples.
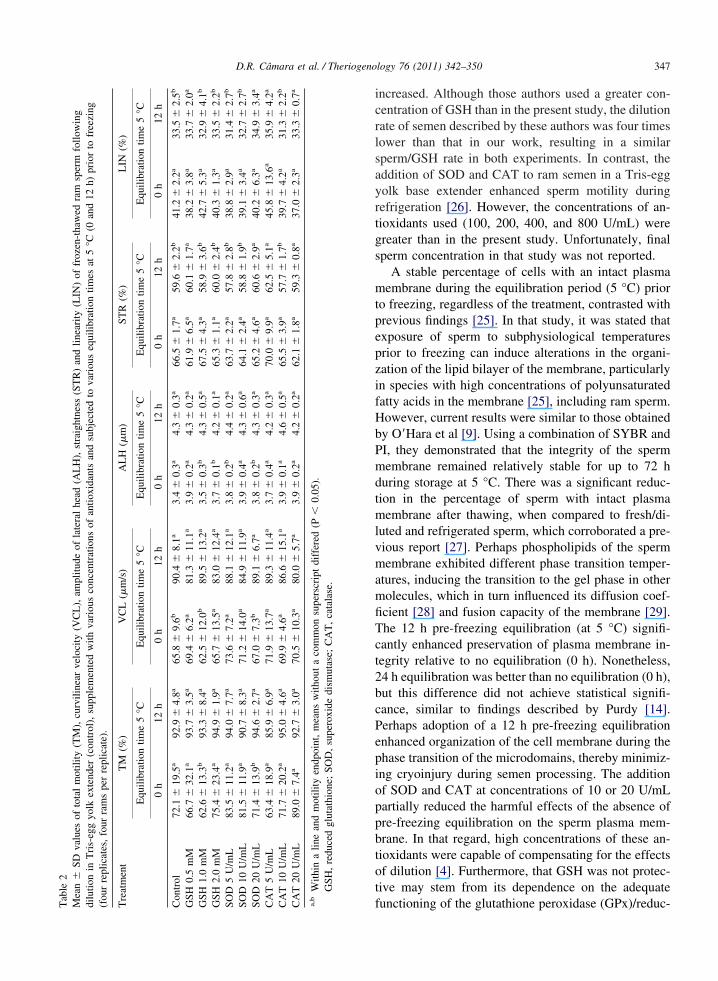
There were no differences (P � 0.05) among treat-ments and duration of pre-freezing equilibration forPM, VAP, VSL, and BCF in thawed semen samples.Combined for all treatments and equilibration times,overall averages were 10.13 � 3.85% (PM), 45.66 �7.58 �m/s (VAP), 28.26 � 5.48 �m/s (VSL), and.95 � 0.84 Hz (BCF). There were no differences (P �.05) among treatments with regards to TM, VCL,LH, STR, and LIN within the same pre-freezing
quilibration time. However, the absence of equilibra-ion time (0 h) reduced (P � 0.05) TM in the GSH 1.0M and SOD 20 U/mL; the VCL in the GSH 1.0 mM
nd SOD 20 U/mL; and the ALH in the GSH 1.0 and.0 mM, SOD 5 and 20 U/mL; in comparison to 12 h ofquilibration. In contrast, the absence of equilibration0 h) increased (P � 0.05) STR and LIN in mostreatments (control; GSH 1.0 and 2.0 mM; SOD 5 and0 U/mL; CAT 20 U/mL) when compared to 12 hquilibration time at 5 °C prior to cryopreservationTable 2). The post-thaw motility parameters of spermamples frozen after 24 h of equilibration time werenaffected (P � 0.05) by the addition of antioxidantsnd were similar (P � 0.05) to those with 12 h equil-
Table 1Mean � SD percentages of ram sperm with intact plasma membranwith various concentrations of antioxidants during refrigeration at 5various equilibration times at 5 °C (0 or 12 h) prior to freezing (fou
Treatments Fresh/diluted Refrigerate
0 h
Control 80.9 � 5.3A 71.2 � 4.9A 71.GSH 0.5 mM 77.9 � 7.1A 71.0 � 11.1A 73.GSH 1.0 mM 77.6 � 5.6A 69.6 � 9.5A 71.GSH 2.0 mM 76.6 � 8.6A 73.4 � 6.7A 73.SOD 5 U/mL 82.6 � 4.8A 73.6 � 8.2A 64.SOD 10 U/mL 79.5 � 4.5A 65.5 � 5.9A 70.SOD 20 U/mL 79.2 � 3.9A 65.9 � 13.9A 65.CAT 5 U/mL 77.7 � 6.8A 72.7 � 8.2A 64.CAT 10 U/mL 76.8 � 2.9A 73.6 � 4.7A 68.CAT 20 U/mL 78.6 � 6.8A 71.2 � 5.9A 64.
a,b Within a line, means without a common superscript differed (PA,B Within a line, means without a common superscript differed (P
GSH, reduced glutathione; SOD, superoxide dismutase; CAT, c
bration. Conversely, 24 h of pre-freezing equilibration r
esulted in greater percentages (P � 0.05) of TM (GSH.0 mM and SOD 20 U/mL) and VCL (SOD 20 U/mL),nd lesser (P � 0.05) percentages of STR (Control,OD 5 and 10 U/mL) and LIN (control, GSH 1.0 and.0 mM, SOD 5 and 10 U/mL) compared to 0 h equil-bration (data not shown).
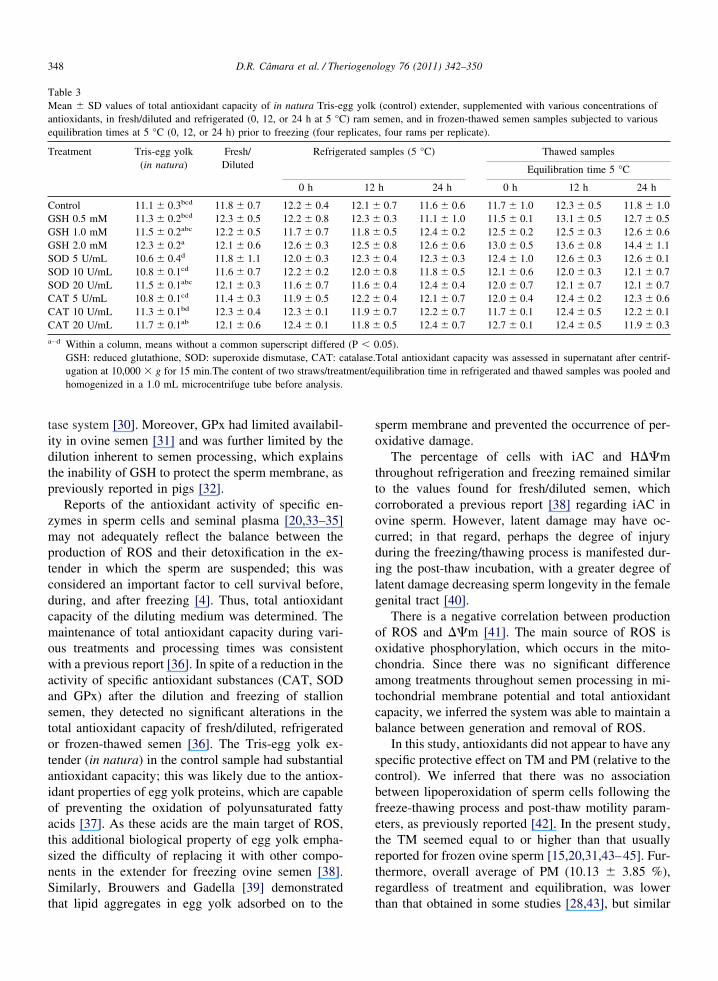
Total antioxidant capacity in the in natura Tris-eggolk was numerically proportional to the concentrationf the supplemented antioxidant in the medium, with aifference (P � 0.05) between the in natura Tris-eggolk treatment (control) and the Tris-egg yolk supple-ented with GSH 2.0 mM. However, there were no
ifferences (P � 0.05) in total antioxidant capacityetween the media supplemented with antioxidants fol-owing the addition of the semen (fresh/diluted) or evenhroughout refrigeration and freezing (Table 3).
. Discussion
The reduction in the percentage of motile spermuring refrigeration in the present study was similar torevious reports [10,12,24]. This phenomenon has beenttributed to changes in the kinetic properties of en-ymes induced by the reduction in temperature [25].upplementation with antioxidants (SOD, CAT, orSH) did not prevent this effect (at the concentrations
ested). Similarly, no improvement in sperm motility inhe presence of GSH (5 or 10 mM) was reported during0 h of refrigeration, when compared to control (nodditives) [24]. Furthermore, in that study, there was a
wing dilution in Tris-egg yolk extender (control), supplemented12, or 24 h) and in frozen-thawed semen samples subjected toates, four rams per replicate).
les (5 °C) Frozen-thawed samples
Equilibration time 5 °C
24 h 0 h 12 h
5A 68.7 � 14.1A 14.8 � 5.6Bb 38.1 � 14.8Ba
6A 69.3 � 12.2A 21.5 � 4.3Bb 36.8 � 9.0Ba
A 73.2 � 13.8A 17.7 � 10.1Bb 48.2 � 15.5Ba
3A 77.2 � 10.2A 16.7 � 11.2Bb 48.0 � 8.3Ba
6A 67.0 � 8.1A 18.8 � 8.9Bb 43.5 � 12.1Ba
0A 64.5 � 16.2A 22.5 � 9.3Ba 39.3 � 10.7Ba
8A 78.5 � 7.2A 23.1 � 9.5Ba 37.1 � 7.4Ba
2A 68.6 � 13.6A 17.1 � 8.6Bb 48.1 � 8.7Ba
2A 68.0 � 16.9A 20.6 � 7.9Ba 41.1 � 9.1Ba
1A 71.5 � 14.2A 28.2 � 6.4Ba 44.3 � 13.8Ba
.).
es follo°C (0,r replic
d samp
12 h
1 � 11.0 � 13.5 � 6.30 � 13.7 � 10.0 � 14.0 � 15.3 � 10.1 � 10.1 � 20.
� 0.05)� 0.01
eduction in sperm motility as the duration of storage
Tct2bc
le2
an�
SDva
lues
ofto
tal
mot
ility
(TM
),cu
rvili
near
velo
city
(VC
L),
ampl
itude
ofla
tera
lhe
ad(A
LH
),st
raig
htne
ss(S
TR
)an
dlin
eari
ty(L
IN)
offr
ozen
-tha
wed
ram
sper
mfo
llow
ing
tion
inT
ris-
egg
yolk
exte
nder
(con
trol
),su
pple
men
ted
with
vari
ous
conc
entr
atio
nsof
antio
xida
nts
and
subj
ecte
dto
vari
ous
equi
libra
tion
times
at5
°C(0
and
12h)
prio
rto
free
zing
urre
plic
ates
,fo
urra
ms
per
repl
icat
e).
atm
ent
TM
(%)
VC
L(�
m/s
)A
LH
(�m
)ST
R(%
)L
IN(%
)
Equ
ilibr
atio
ntim
e5
°CE
quili
brat
ion
time
5°C
Equ
ilibr
atio
ntim
e5
°CE
quili
brat
ion
time
5°C
Equ
ilibr
atio
ntim
e5
°C
0h
12h
0h
12h
0h
12h
0h
12h
0h
12h
ntro
l72
.1�
19.5
a92
.9�
4.8a
65.8
�9.
6b90
.4�
8.1a
3.4
�0.
3a4.
3�
0.3a
66.5
�1.
7a59
.6�
2.2b
41.2
�2.
2a33
.5�
2.5b
H0.
5m
M66
.7�
32.1
a93
.7�
3.5a
69.4
�6.
2a81
.3�
11.1
a3.
9�
0.2a
4.3
�0.
2a61
.9�
6.5a
60.1
�1.
7a38
.2�
3.8a
33.7
�2.
0a
H1.
0m
M62
.6�
13.3
b93
.3�
8.4a
62.5
�12
.0b
89.5
�13
.2a
3.5
�0.
3b4.
3�
0.5a
67.5
�4.
3a58
.9�
3.6b
42.7
�5.
3a32
.9�
4.1b
H2.
0m
M75
.4�
23.4
a94
.9�
1.9a
65.7
�13
.5a
83.0
�12
.4a
3.7
�0.
1b4.
2�
0.1a
65.3
�1.
1a60
.0�
2.4b
40.3
�1.
3a33
.5�
2.2b
D5
U/m
L83
.5�
11.2
a94
.0�
7.7a
73.6
�7.
2a88
.1�
12.1
a3.
8�
0.2b
4.4
�0.
2a63
.7�
2.2a
57.8
�2.
8b38
.8�
2.9a
31.4
�2.
7b
D10
U/m
L81
.5�
11.9
a90
.7�
8.3a
71.2
�14
.0a
84.9
�11
.9a
3.9
�0.
4a4.
3�
0.6a
64.1
�2.
4a58
.8�
1.9b
39.1
�3.
4a32
.7�
2.7b
D20
U/m
L71
.4�
13.9
b94
.6�
2.7a
67.0
�7.
3b89
.1�
6.7a
3.8
�0.
2b4.
3�
0.3a
65.2
�4.
6a60
.6�
2.9a
40.2
�6.
3a34
.9�
3.4a
T5
U/m
L63
.4�
18.9
a85
.9�
6.9a
71.9
�13
.7a
89.3
�11
.4a
3.7
�0.
4a4.
2�
0.3a
70.0
�9.
9a62
.5�
5.1a
45.8
�13
.6a
35.9
�4.
2a
T10
U/m
L71
.7�
20.2
a95
.0�
4.6a
69.9
�4.
6a86
.6�
15.1
a3.
9�
0.1a
4.6
�0.
5a65
.5�
3.9a
57.7
�1.
7b39
.7�
4.2a
31.3
�2.
2b
T20
U/m
L89
.0�
7.4a
92.7
�3.
0a70
.5�
10.3
a80
.0�
5.7a
3.9
�0.
2a4.
2�
0.2a
62.1
�1.
8a59
.3�
0.8a
37.0
�2.
3a33
.3�
0.7a
With
ina
line
and
mot
ility
endp
oint
,m
eans
with
out
aco
mm
onsu
pers
crip
tdi
ffer
ed(P
�0.
05).
GSH
,re
duce
dgl
utat
hion
e;SO
D,
supe
roxi
dedi
smut
ase;
CA
T,
cata
lase
.
347D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
increased. Although those authors used a greater con-centration of GSH than in the present study, the dilutionrate of semen described by these authors was four timeslower than that in our work, resulting in a similarsperm/GSH rate in both experiments. In contrast, theaddition of SOD and CAT to ram semen in a Tris-eggyolk base extender enhanced sperm motility duringrefrigeration [26]. However, the concentrations of an-tioxidants used (100, 200, 400, and 800 U/mL) weregreater than in the present study. Unfortunately, finalsperm concentration in that study was not reported.
A stable percentage of cells with an intact plasmamembrane during the equilibration period (5 °C) priorto freezing, regardless of the treatment, contrasted withprevious findings [25]. In that study, it was stated thatexposure of sperm to subphysiological temperaturesprior to freezing can induce alterations in the organi-zation of the lipid bilayer of the membrane, particularlyin species with high concentrations of polyunsaturatedfatty acids in the membrane [25], including ram sperm.However, current results were similar to those obtainedby O=Hara et al [9]. Using a combination of SYBR andPI, they demonstrated that the integrity of the spermmembrane remained relatively stable for up to 72 hduring storage at 5 °C. There was a significant reduc-tion in the percentage of sperm with intact plasmamembrane after thawing, when compared to fresh/di-luted and refrigerated sperm, which corroborated a pre-vious report [27]. Perhaps phospholipids of the spermmembrane exhibited different phase transition temper-atures, inducing the transition to the gel phase in othermolecules, which in turn influenced its diffusion coef-ficient [28] and fusion capacity of the membrane [29].
he 12 h pre-freezing equilibration (at 5 °C) signifi-antly enhanced preservation of plasma membrane in-egrity relative to no equilibration (0 h). Nonetheless,4 h equilibration was better than no equilibration (0 h),ut this difference did not achieve statistical signifi-ance, similar to findings described by Purdy [14].
Perhaps adoption of a 12 h pre-freezing equilibrationenhanced organization of the cell membrane during thephase transition of the microdomains, thereby minimiz-ing cryoinjury during semen processing. The additionof SOD and CAT at concentrations of 10 or 20 U/mLpartially reduced the harmful effects of the absence ofpre-freezing equilibration on the sperm plasma mem-brane. In that regard, high concentrations of these an-tioxidants were capable of compensating for the effectsof dilution [4]. Furthermore, that GSH was not protec-tive may stem from its dependence on the adequate
functioning of the glutathione peroxidase (GPx)/reduc-TabM
edi
lu(f
o
Tre
Co
GS
GS
GS
SO SO SO CA
CA
CA
a,b
aastotaioatsnS
348 D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
tase system [30]. Moreover, GPx had limited availabil-ity in ovine semen [31] and was further limited by thedilution inherent to semen processing, which explainsthe inability of GSH to protect the sperm membrane, aspreviously reported in pigs [32].
Reports of the antioxidant activity of specific en-zymes in sperm cells and seminal plasma [20,33–35]may not adequately reflect the balance between theproduction of ROS and their detoxification in the ex-tender in which the sperm are suspended; this wasconsidered an important factor to cell survival before,during, and after freezing [4]. Thus, total antioxidantcapacity of the diluting medium was determined. Themaintenance of total antioxidant capacity during vari-ous treatments and processing times was consistentwith a previous report [36]. In spite of a reduction in thectivity of specific antioxidant substances (CAT, SODnd GPx) after the dilution and freezing of stallionemen, they detected no significant alterations in theotal antioxidant capacity of fresh/diluted, refrigeratedr frozen-thawed semen [36]. The Tris-egg yolk ex-ender (in natura) in the control sample had substantialntioxidant capacity; this was likely due to the antiox-dant properties of egg yolk proteins, which are capablef preventing the oxidation of polyunsaturated fattycids [37]. As these acids are the main target of ROS,his additional biological property of egg yolk empha-ized the difficulty of replacing it with other compo-ents in the extender for freezing ovine semen [38].imilarly, Brouwers and Gadella [39] demonstrated
Table 3Mean � SD values of total antioxidant capacity of in natura Tris-egantioxidants, in fresh/diluted and refrigerated (0, 12, or 24 h at 5 °Cequilibration times at 5 °C (0, 12, or 24 h) prior to freezing (four re
Treatment Tris-egg yolk(in natura)
Fresh/Diluted
Refrige
0 h
Control 11.1 � 0.3bcd 11.8 � 0.7 12.2 � 0.4GSH 0.5 mM 11.3 � 0.2bcd 12.3 � 0.5 12.2 � 0.8GSH 1.0 mM 11.5 � 0.2abc 12.2 � 0.5 11.7 � 0.7GSH 2.0 mM 12.3 � 0.2a 12.1 � 0.6 12.6 � 0.3SOD 5 U/mL 10.6 � 0.4d 11.8 � 1.1 12.0 � 0.3SOD 10 U/mL 10.8 � 0.1cd 11.6 � 0.7 12.2 � 0.2SOD 20 U/mL 11.5 � 0.1abc 12.1 � 0.3 11.6 � 0.7CAT 5 U/mL 10.8 � 0.1cd 11.4 � 0.3 11.9 � 0.5CAT 10 U/mL 11.3 � 0.1bd 12.3 � 0.4 12.3 � 0.1CAT 20 U/mL 11.7 � 0.1ab 12.1 � 0.6 12.4 � 0.1
a–d Within a column, means without a common superscript differedGSH: reduced glutathione, SOD: superoxide dismutase, CAT: cugation at 10,000 � g for 15 min.The content of two straws/treahomogenized in a 1.0 mL microcentrifuge tube before analysis.
that lipid aggregates in egg yolk adsorbed on to the
sperm membrane and prevented the occurrence of per-oxidative damage.
The percentage of cells with iAC and H�mthroughout refrigeration and freezing remained similarto the values found for fresh/diluted semen, whichcorroborated a previous report [38] regarding iAC inovine sperm. However, latent damage may have oc-curred; in that regard, perhaps the degree of injuryduring the freezing/thawing process is manifested dur-ing the post-thaw incubation, with a greater degree oflatent damage decreasing sperm longevity in the femalegenital tract [40].
There is a negative correlation between productionof ROS and �m [41]. The main source of ROS isoxidative phosphorylation, which occurs in the mito-chondria. Since there was no significant differenceamong treatments throughout semen processing in mi-tochondrial membrane potential and total antioxidantcapacity, we inferred the system was able to maintain abalance between generation and removal of ROS.
In this study, antioxidants did not appear to have anyspecific protective effect on TM and PM (relative to thecontrol). We inferred that there was no associationbetween lipoperoxidation of sperm cells following thefreeze-thawing process and post-thaw motility param-eters, as previously reported [42]. In the present study,the TM seemed equal to or higher than that usuallyreported for frozen ovine sperm [15,20,31,43–45]. Fur-thermore, overall average of PM (10.13 � 3.85 %),regardless of treatment and equilibration, was lower
(control) extender, supplemented with various concentrations ofemen, and in frozen-thawed semen samples subjected to various, four rams per replicate).
mples (5 °C) Thawed samples
Equilibration time 5 °C
h 24 h 0 h 12 h 24 h
0.7 11.6 � 0.6 11.7 � 1.0 12.3 � 0.5 11.8 � 1.00.3 11.1 � 1.0 11.5 � 0.1 13.1 � 0.5 12.7 � 0.50.5 12.4 � 0.2 12.5 � 0.2 12.5 � 0.3 12.6 � 0.60.8 12.6 � 0.6 13.0 � 0.5 13.6 � 0.8 14.4 � 1.10.4 12.3 � 0.3 12.4 � 1.0 12.6 � 0.3 12.6 � 0.10.8 11.8 � 0.5 12.1 � 0.6 12.0 � 0.3 12.1 � 0.70.4 12.4 � 0.4 12.0 � 0.7 12.1 � 0.7 12.1 � 0.70.4 12.1 � 0.7 12.0 � 0.4 12.4 � 0.2 12.3 � 0.60.7 12.2 � 0.7 11.7 � 0.1 12.4 � 0.5 12.2 � 0.10.5 12.4 � 0.7 12.7 � 0.1 12.4 � 0.5 11.9 � 0.3
.05).Total antioxidant capacity was assessed in supernatant after centrif-quilibration time in refrigerated and thawed samples was pooled and
g yolk) ram splicates
rated sa
12
12.1 �12.3 �11.8 �12.5 �12.3 �12.0 �11.6 �12.2 �11.9 �11.8 �
(P � 0atalase.tment/e
than that obtained in some studies [28,43], but similar

it
mUdrsderS
A
fdN(
R
[
[
[
[
[
[
[
349D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
to others [13,44,46]. For unknown reasons, additionalmotility end points assessed in thawed semen samples(VAP, VSL, VCL, ALH, BCF, STR and LIN) seemedlower than those previously reported [15,28,43,44].However, despite the numerous advantages of CASA[47], comparing sperm kinematics between studies isdifficult, since CASA settings, sperm concentrationsand diluents used can affect these end points [48,49],and furthermore, CASA settings are frequently notspecified.
It was not possible to identify any factor that couldexplain the reduction in TM in GSH 1.0 mM and SOD20 U/mL; in VCL in the control, GSH 1.0 mM andSOD 20 U/mL treatments; and in ALH in the GSH 1.0and 2.0 mM and SOD 5 and 20 U/mL treatmentssubjected to 0 h of equilibration in comparison to 12 hpre-freezing equilibration. Perhaps this variation wasrelated to subpopulations of sperm with different anti-oxidant enzyme distribution patterns and the proportionof each cell subtype may be altered by the processing[50], which was reflected in the metabolic and func-tional characteristics of the sample. The greater LIN inthe sperm in the absence of equilibration relative to12 h equilibration in the control and samples supple-mented with GSH (1.0 and 2.0 mM), SOD (5 and 10U/mL) and CAT (10 U/mL) may be indicative of lowerin vivo fertility rates. Verstegen et al. [47] stated that anncrease in LIN values may be related to reduced fer-ilization capacity.
In summary, the addition of GSH (0.5, 1.0, or 2.0M), SOD (5, 10, or 20 U/mL) and CAT (5, 10, or 20/mL) did not significantly influence the total antioxi-ant capacity of the Tris-egg yolk extender throughoutefrigeration and freezing. However, maintaining ovineemen at 5 °C for 12 h prior to cryopreservation re-uced cell membrane damage. Moreover, the harmfulffects of cryopreserving semen once equilibration waseached were partially overcome by the addition ofOD (10 or 20 U/mL) and CAT (10 or 20 U/mL).
cknowledgements
The authors are grateful to the following Brazilianostering agencies: Coordenação de Aperfeiçoamentoe Pessoal de Nível Superior (CAPES) and Conselhoacional de Desenvolvimento Científico e Tecnológico
CNPq) for their financial support.
eferences
[1] Lightfoot RJ, Salamon S. Fertility of ram spermatozoa frozen by
the pellet method: I—Transport and viability of spermatozoawithin the genital tract of the ewe. J Reprod Fertil 1970;22:385–98.
[2] Alvarez JG, Storey BT. Evidence for increased lipid peroxida-tive damage and loss of superoxide dismutase activity as amodel of sublethal cryodamage to human sperm during cryo-preservation. J Androl 1992;13:232–41.
[3] Meyers SA. Spermatozoal response to osmotic stress. AnimReprod Sci 2005;89:57–64.
[4] Bilodeau JF, Chatterjee S, Sirard MA, Gagnon C. Level ofantioxidant defenses are decreased in bovine spermatozoa aftera cycle of freezing and thawing. Mol Reprod Dev 2000;55:282–8.
[5] Misra HP, Fridovich I. The univalent reduction of oxygen byreduced flavins and quinones. J Biol Chem 1972;247:188–92.
[6] Aitken RJ. Free radicals, lipidic peroxidation and sperm func-tion. Reprod Fertil Dev 1995;7:659–68.
[7] O’Flaherty CM, Beorlegui NB, Beconi MT. Reactive oxygenspecies requirements for bovine sperm capacitation and acro-some reaction. Theriogenology 1999;52:289–301.
[8] Taylor CT. Antioxidants and reactive oxygen species in humanfertility. Environ Toxicol Pharmacol 2001;10:189–98.
[9] O’Hara L, Hanrahan JP, Richardson L, Donovan A, Fair S,Evans ACO, Lonergan P. Effect of storage duration, storagetemperature, and diluent on the viability and fertility of freshram sperm. Theriogenology 2010;73:541–9.
10] Paulenz H, Söderquist L, Pérez-Pé R, Berg KA. Effect ofdifferent extenders and storage temperatures on sperm viabilityof liquid ram semen. Theriogenology 2002;57:823–36.
11] Menchaca A, Pinczak A, Queirolo D. Storage of ram semen at5 °C: effects of preservation period and timed artificial insem-ination on pregnancy rate in ewes. Anim Reprod 2005;2:195–8.
12] Kasimanickam R, Kasimanickam V, Pelzer KD, Dascanio JJ.Effect of breed and sperm concentration on the changes instructural, functional and motility parameters of ram-lamb sper-matozoa during storage at 4 °C. Anim Reprod Sci 2007;101:60–73.
13] El-Alamy MA, Foote RH. Freezability of spermatozoa fromFinn and Dorset rams in multiple semen extenders. Anim Re-prod Sci 2001;65:245–54.
14] Purdy PH. The post-thaw quality of ram sperm held for 0 to 48 hat 5 °C prior to cryopreservation. Anim Reprod Sci 2006;93:114–23.
15] Purdy PH, Mocé E, Stobart R, Murdoch WJ, Moss GE, LarsonB, Ramsey S, Graham JK, Blackburn HD. The fertility of ramsperm held for 24 h at 5 degrees C prior to cryopreservation.Anim Reprod Sci 2010;118:231–5.
16] Herold FC, Haas K, Colenbrander B, Gerber D. Comparison ofequilibration times when freezing epididymal sperm from Af-rican buffalo (Syncerus caffer) using Triladyl™ and An-dromed®. Theriogenology 2006;66:1123–30.
[17] Foote RH, Kaproth MT. Large batch freezing of bull semen:effect of time of freezing and fructose on fertility. J Dairy Sci2002;85:453–6.
[18] Sullivan JJ, Mixner JP. Effects of method of egg yolk additionand of glycerol equilibration time upon post-thawing motitlityand metabolic activity of frozen Bull semen. J Dairy Sci 1963;46:463–7.
[19] Leite TG, Vale-Filho VR, Arruda RP, Andrade AFC, EmerickLL, Zaffalon FG, Martins JAM, Andrade VJ. Effects of ex-tender and equilibration time on post-thaw motility and mem-brane integrity of cryopreserved Gyr Bull semen evaluated by
CASA and flow cytometry. Anim Reprod Sci 2010;120:31–8.
[
[
[
350 D.R. Câmara et al. / Theriogenology 76 (2011) 342–350
[20] Bucak MN, Atessahim A, Yüce A. Effect of anti-oxidants andoxidative stress parameters on ram semen after the freezing-thawing process. Small Rum Res 2008;75:128–34.
[21] Harrison RAP, Vickers SE. Use of fluorescent probes to assessmembrane integrity in mammalian spermatozoa. J Reprod Fertil1990;88:343–52.
[22] Coleto ZF, Guerra MMP, Batista AM. Avaliação do sêmencongelado de caprinos com drogas fluorescentes. Rev Bras MedVet 2002;24:101–4.
[23] Roth TL, Weiss RB, Buff JL, Bush LM, Wildt DE, Bush M.Heterologous in vitro fertilization and sperm capacitation in anendangered African antelope, the Scimitar-Horned Oryx (Oryxdammah). Biol Reprod 1998;58:475–82.
[24] Bucak MN, Tekin N. Protective effect of taurine, glutathioneand trehalose on the liquid storage of ram semen. Small RumRes 2007;73:103–8.
[25] Holt WV. Fundamental aspects of sperm cryobiology: the im-portance of species and individual differences. Theriogenology2000;53:47–58.
[26] Maxwell WMC, Stojanov T. Liquid storage of ram semen in theabsence or presence of some antioxidants. Reprod Fertil Dev1996;8:1013–20.
[27] Wolf CA, James PS, Mackie AR, Ladha S, Jones R. Regional-ized lipid diffusion in the plasma membrane of mammalianspermatozoa. Biol Reprod 1998;59:1506–14.
[28] Maia MS, Bicudo SD, Azevedo HC, Sicherle CC, Sousa DB,Rodello L. Motility and viability of ram semen cryopreserved ina Tris-egg yolk extender supplemented with anti-oxidants.Small Rum Res 2009;85:85–90.
[29] Holt WV, North RD. Thermotropic phase transitions in theplasma membrane of ram spermatozoa. J Reprod Fertil 1986;78:447–57.
[30] Baumber J, Ball BA, Gravance CG, Medina V, Davies-MorelMCG. The effect of reactive oxygen species on equine spermmotility, viability, acrosomal integrity, mitochondrial mem-brane potential, and membrane lipid peroxidation. J Androl2000;21:895–902.
[31] Gadea J, Sellés E, Marco MA, Coy P, Matás C, Romar R, RuizS. Decrease in glutathione content in boar sperm after cryo-preservation – Effect of the addition of reduced glutathione tothe freezing and thawing extender. Theriogenology 2004;62:690–701.
[32] Whitaker BD, Carle B, Mukai T, Simpson A, Vu L, Knight JW.Effect of exogenous glutathione supplementation on motility,viability, and DNA integrity of frozen-thawed boar semen.Anim Reprod 2008;5:127–31.
[33] Martí JI, Martí E, Cebrián-Peréz JA, Muiño-Blanco T. Survivalrate and antioxidant enzyme activity of ram spermatozoa afterdilution with different extenders or selection by an dextranswim-up procedure. Theriogenology 2003;60:1025–37.
[34] Kasimanickam R, Pelzer KD, Kasimanickam V, Swecker WS,Thatcher CD. Association of classical semen parameters, spermDNA fragmentation index, lipid peroxidation and antioxidantenzymatic activity of semen in ram-lambs. Theriogenology2006;65:1407–21.
[35] Martí E, Mara L, Martí JI, Muiño-Blanco T, Cebrián-Peréz JA.Seasonal variations in antioxidant enzyme activity in ram sem-
inal plasma. Theriogenology 2007;67:1446–54.[36] Bustamante Filho IC, Pederlozzi CD, Sgaravatti AM, GregoryRM, Dutra Filho CS, Jobim MIM, Mattos RC. Skim Milk-eggbased semen extender compensates for non-enzymatic antioxi-dant activity loss during equine semen cryopreservation. AnimReprod 2009;6:392–9.
[37] Sakanaba S, Tachibana Y, Ishihara N, Juneja LR. Antioxidantactivity of egg-yolk protein hydrolysates in a linoleic acid ox-idation system. Food Chem 2004;86:99–103.
[38] Valente SS, Pereira RM, Baptista MC, Marques CC, VasquesMI, Silva Pereira MVC, Horta AEM, Barbas JP. In vitro and invivo fertility of ram semen cryopreserved in different extenders.Anim Reprod Sci 2010;117:74–7.
[39] Brouwers JFHM, Gadella BM. In situ detection and localizationof lipid peroxidation in individual bovine sperm cells. FreeRadic Biol Medic 2003;35:1382–91.
[40] Bag S, Joshi A, Naqvi SMK, Mittal JP. Effect of post-thawincubation on sperm kinematics and acrosomal integrity of ramspermatozoa cryopreserved in medium-sized French straws.Theriogenology 2004;62:415–24.
[41] Wang XW, Sharma RK, Gupta A, George V, Thomas AJ, JrFalcone T, Agarwal A. Alterations in mitochondria membranepotential and oxidative stress in infertile men: a prospectiveobservational study. Fertil Steril 2003;80(Suppl. 2):844–50.
[42] Maia MS, Bicudo SD, Sicherle CC, Rodello L, Gallego ICS.Lipid peroxidation and generation of hydrogen peroxide infrozen-thawed ram semen cryopreserved in extender with anti-oxidants. Small Rum Res 2010:122;118–23.
[43] Anel L, Paz P, Álvarez, M, Chamorro CA, Boixo JC, Manso A,González M, Kaabi M, Anel E. Field and in vitro assay of threemethods for freezing ram semen. Theriogenology 2003;60:1293–308.
[44] Moustacas VS, Zaffalon FG, Lagares MA, Loaiza-EccheverriAM, Varago FC, Neves MM, Heneine LGD, Arruda RP, HenryM. Natural, but not lyophilized, low density lypoproteins werean acceptable alternative to egg yolk for cryopreservation ofram semen. Theriogenology 2010;75:300–7.
[45] Nur R, Zik B, Ustuner B, Sagirkaya H, Ozguden CG. Effects ofdifferent cryoprotective agents on ram sperm morphology andDNAintegrity. Theriogenology 2010;73:1267–75.
[46] Sánchez-Partida LG, Windsor DP, Eppleston J, Setchell BP,Maxwell WMC. Fertility and its relationship to motility char-acteristics of spermatozoa in ewes after cervical, transcervical,and intrauterine insemination with frozen-thawed ram semen. JAndrol 1999;20:280–8.
[47] Mocé E, Graham JK. In vitro evaluation of sperm quality. AnimReprod Sci 2008;105:104–18.
48] Verstegen J, Iguer-Ouada M, Onclin K. Computer assisted se-men analyzers in andrology research and veterinary practice.Theriogenology 2002;57:149–79.
49] Mortimer ST, Maxwell WMC. Effect of medium on the kine-matics of frozen-thawed ram spermatozoa. Reproduction 2004;127:285–91.
50] Martí E, Martí JI, Muiño-Blanco T, Cebrián-Pérez JA. Effect ofthe cryopreservation process on the activity and immunolocal-ization of antioxidant enzymes in ram spermatozoa. J Androl
2008;29:459–67.




























