Embed Size (px)
Citation preview

Efferent Connections of Septal Nuclei ofthe Domestic Chick (Gallus
domesticus): An Anterograde PathwayTracing Study with a Bearing on
Functional Circuits
CATHERINE M. MONTAGNESE, ANDREA D. SZEKELY, AGOTA ADAM,
AND ANDRAS CSILLAG*
Department of Anatomy, Semmelweis University of Medicine, Budapest, 1094, Hungary
ABSTRACTSmall iontophoretic injections of the anterograde tracer Phaseolus vulgaris leucoagglutinin
were placed in different subregions of the septum of domestic chicks. The main targets of septalprojections comprised the ipsi- and contralateral septal nuclei, including the nucleus of thediagonal band, basal ganglia, including the ventral paleostriatum, lobus parolfactorius, nucleusaccumbens, and olfactory tubercle, archistriatum, piriform cortex, and anterior neostriatum.Further diencephalic and mesencephalic septal projections were observed in the ipsilateralpreoptic region, hypothalamus (the main regions of afferentation comprising the lateral hypo-thalamic nuclei, ventromedial, paraventricular and periventricular nuclei, and the mammillaryregion), dorsal thalamus, medial habenular and subhabenular nuclei, midbrain central gray, andventral tegmental area. Contralateral projections were also encountered in the septal nuclei,ventral paleostriatum, periventricular and anteromedial hypothalamic nuclei, suprachiasmaticnucleus, and the lateral hypothalamic area. Avian septal efferents are largely similar to those ofmammals, the main differences being a relatively modest hippocampal projection arising mainlyfrom the nucleus of the diagonal band (as confirmed by a specific experiment with the retrogradepathway tracer True blue), the lack of interpeduncular projection, and a greater contingent ofamygdalar efferents arising from the lateral septum rather than the nucleus of the diagonalband. This pattern of connectivity is likely to reflect an important role of the avian septal nucleiin the coordination of limbic circuits and the integration of a wide variety of information sourcesmodulating the appropriate behavioral responses: attention and arousal level, memory forma-tion, hormonally mediated behaviors, and their affective components (such as ingestive, repro-ductive, and parental behaviors), social interaction, locomotor modulation, and circadian rhythm.J. Comp. Neurol. 469:437–456, 2004. © 2004 Wiley-Liss, Inc.
Indexing terms: avian brain; limbic system; Phaseolus vulgaris leucoagglutinin; Galliformes;
telencephalon; diencephalon
The septal nuclei are an important component of thelimbic system which has been extensively studied in mam-mals. However, little is known of the septal connectivity inbirds. The only known systematic description of the septalefferents in the pigeon by Krayniak and Siegel (1978) hasshown a basic organization similar to that of the mam-mals: the septal nuclei are strategically placed betweenthe hippocampus on the one hand and the basal forebrainand diencephalon on the other. They are connected to thehippocampal formation (mammals: Freund and Antal,1988; Risold and Swanson 1997b; pigeon: Krayniak andSiegel, 1978) and the diencephalon, the hypothalamusbeing a major recipient site (mammals: Meibach and Sie-gel, 1977a; Risold and Swanson 1997b; pigeon: Krayniak
and Siegel, 1978). The septum also has projections on themidbrain reticular formation (mammals: Meibach and
Grant sponsor: Fondation Franco-Hongroise pour la Jeunesse; Grantnumber: OTKA T 029613; Grant number: OTKA T 043462; Grant number:ETT 309/2000.
*Correspondence to: Andras Csillag, Department of Anatomy, Semmel-weis University of Medicine, Tuzolto u. 58, Budapest, 1094, Hungary.E-mail: [email protected]
Received 8 May 2003; Revised 9 September 2003; Accepted 15 October2003
DOI 10.1002/cne.11018Published online the week of January 5, 2004 in Wiley InterScience
(www.interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 469:437–456 (2004)
© 2004 WILEY-LISS, INC.

Siegel, 1977a; Risold and Swanson 1997b; pigeon: Krayn-iak and Siegel, 1978). In mammals, a major afferent sys-tem is represented by the hippocampal fibers, which ter-minate in the lateral septal area (SL; Meibach and Siegel,1977b; Risold and Swanson, 1997b). Direct afferents also
arise from hippocampal nonpyramidal neurons to the cellsof the medial septum (SM)-diagonal band of Broca (DBB)complex (Toth and Freund, 1992; Toth et al., 1993). Otherseptal afferents include fibers originating from the ento-rhinal cortex (Alonso and Kohler, 1984), hypothalamus
Abbreviations
AA archistriatum anteriusAc nucleus accumbensAid archistriatum intermedium pars dorsalisAiv archistriatum intermedium pars ventralisAL ansa lenticularisAm archistriatum medialeAM hypothalamic anterior nucleusAp archistriatum posteriusAP pretectal areaAVT ventral tegmental area (of Tsai)BCS brachium conjunctivum ascendensBSTl lateral part of the bed nucleus of the stria terminalisCA anterior commissureCb cerebellumCHCS corticohabenular and corticoseptal tractCO optic chiasmCP posterior commissureCPa pallial commissureCPi piriform cortexCT tectal commissureCTz trapezoid body (Papez)DBB diagonal band of BrocaDHA dorsal hypothalamic areaDIP thalamic posterior dorsointermediate nucleusDLAl thalamic anterior dorsolateral nucleus, pars lateralisDLAm thalamic anterior dorsolateral nucleus, pars medialisDLAmc thalamic anterior dorsolateral nucleus, pars magnocellu-
larisDLP thalamic posterior dorsolateral nucleusDMA thalamic anterior dorsomedial nucleusDMN hypothalamic dorsomedial nucleusDMP thalamic posterior dorsomedial nucleusDSD dorsal supraoptic decussationE ectostriatumFDB fasciculus diagonalis BrocaeFLM medial longitudinal fascicleFPL lateral forebrain bundleFRL mesencephalic lateral reticular formationFRM mesencephalic medial reticular formationGCt midbrain central grayGLdp lateral geniculate nucleus, pars dorsalis principalisGLv lateral geniculate nucleus, pars ventralisHL lateral habenular nucleusHM medial habenular nucleusHIP habenulointerpeduncular tractHP hippocampusHV hyperstriatum ventraleICo intercollicular nucleusICT thalamic intercalate nucleusIH hypothalamic inferior nucleusImc nucleus isthmi, pars magnocellularisIN infundibular nucleusIP interpeduncular nucleusIpc nucleus isthmi, pars parvocellularisLA thalamic anterior lateral nucleusLHy lateral hypothalamusLM nucleus lentiformis mesencephaliLoC locus coeruleusLPO lobus parolfactoriusME median eminenceMFB medial forebrain bundlemHy medial hypothalamusML lateral mammillary nucleusMLd dorsal lateral mesencephalic nucleusMM medial mammillary nucleusMPv nucleus mesencephalicus profundus, pars ventralisN neostriatumnCPa nucleus of the pallial commissure
nBOR nucleus of the basal optic rootNI neostriatum intermediumnI nucleus c of RendahlNIII oculomotor nervenPrV principal sensory nucleus of the trigeminal nervenST bed nucleus of the stria terminalisnTSM nucleus of the septo-mesencephalic tract (nucleus superfi-
cialis parvocellularis)OM occipito-mesencephalic tractOMd nucleus of the oculomotor nerve, pars dorsalisOMv nucleus of the oculomotor nerve, pars ventralisOv nucleus ovoidalisOVLT vascular organ of the lamina terminalisPA paleostriatum augmentatumPap nucleus papillioformisPha-L Phaseolus vulgaris leucoagglutininPHN hypothalamic periventricular nucleusPL lateral pontine nucleusPM medial pontine nucleusPMN premamillary nucleusPMI nucleus paramedianus internus thalamiPO preoptic regionPOA preoptic areaPOM medial preoptic nucleusPOP preoptic periventricular nucleusPP paleostriatum primitivumPPN pedunculopontine nucleusPT pretectal nucleusPTM medial pretectal nucleusPVN paraventricular nucleusPVT ventral paleostriatum (Kitt and Brauth)QF quintofrontal tractR rapheRI infundibular recessROT nucleus rotundusRPgc gigantocellular caudal pontine reticular nucleusRPO nucleus reticularis pontis oralisRSd dorsal superior reticular nucleusRSv ventral superior reticular nucleusSCd nucleus subcoeruleus dorsalisSCN suprachiasmatic nucleusSCv nucleus subcoeruleus ventralisSGP stratum griseum periventriculareSHL lateral subhabenular nucleusSHM medial subhabenular nucleusSL lateral septal nucleusSM medial septal nucleusSMN interstitial supramammillary nucleusSOe external supraoptic nucleusSOv ventral supraoptic nucleusSP subpretectal nucleusT nucleus triangularisTeO optic tectumTIO isthmooptic tractTn nucleus taeniaeTO olfactory tubercleTOV tract of the nucleus ovoidalisTPc/SN tegmental pedunculo-pontine nucleus pars compacta/
substantia nigraTrO optic tractTSM septomesencephalic tractTVM vestibulo-mesencephalic tractV ventricleVeM medial vestibular nucleusVLT thalamic ventrolateral nucleusVMN hypothalamic ventromedial nucleusVTA ventral tegmental area
438 C.M. MONTAGNESE ET AL.

(Jakab and Leranth, 1993; Szeidelmann et al., 1995a,b),supramammillary nucleus (Vertes, 1992; Vertes and Mc-Kenna, 2000), tegmental nuclei (Szeidelmann et al.,1995b), ventral tegmental area (VTA; Lindvall and Ste-nevi, 1978; Jakab and Leranth, 1993), raphe (Kohler et al.,1982; Vertes, 1991; Acsady et al., 1996), and locus coer-uleus (LoC; Dahlstrom and Fuxe, 1964; Fuxe, 1965;McBride and Sutin, 1976; Lindvall and Stenevi, 1978). Asimilar organization of the afferent septal connections isevident in birds. The hippocampal formation denselyprojects to the septal area in the pigeon (Krayniak andSiegel, 1978; Casini et al., 1986) and the zebra finch(Szekely and Krebs, 1996; Szekely, 1999). Several hypo-thalamic nuclei send numerous afferents to both SM andSL (Berk and Butler, 1981; Berk and Finkelstein, 1983;Berk and Hawkin, 1985; Balthazart et al., 1994). Mesen-cephalic terminals in the septal nuclei arise from thetegmental pedunculo-pontine nucleus pars compacta, theavian substantia nigra (TPc/SN; Kitt and Brauth, 1986b),LoC, and the dorsal and ventral subcoeruleus nuclei (SCd,SCv; Kitt and Brauth, 1986a).
No detailed study has been published on the efferents oflateral and medial septal nuclei in the domestic chick.Unlike pigeons, young domestic chicks are precocial andnidifugous. Their brain is well developed at the time ofhatching and therefore comparable to the adult brain.This species is widely used in behavioral neuroanatomyand neuropharmacology, in particular learning and im-printing tasks, with a bearing on limbic circuits (Horn,1991; Rose, 1991). The relationship remains unclear be-tween the septum and the areas involved in the earlyadaptive learning of young domestic chicks, which includethe lobus parolfactorius (LPO), the limbic archistriatum,and possibly the dorsal thalamic zone, a putative equiva-lent of the mammalian thalamic intralaminar, midline,and mediodorsal nuclei (Veenman et al., 1995a, 1997;Montagnese et al., 2003). Using small injections of thesensitive anterograde pathway tracer Phaseolus vulgarisleucoagglutinin (Pha-L), the present study was under-taken to investigate the ascending and descending effer-ents specific to the septal subregions, which were ex-pected, on the basis of previous information, to beanatomically and functionally heterogeneous in the do-mestic chick.
MATERIALS AND METHODS
Experimental animals
Purchased from a local supplier, freshly hatched Hun-nia broiler chicks of either sex were housed in groups withwater and food ad libitum. The experiments were con-ducted in conformity with the laws and regulations con-trolling experiments and procedures in live animals, asdescribed in the Principles of Laboratory Animal Care(NIH Publication 85-23, revised 1985).
Anterograde tracer injection
One-week-old chicks (70–80 g body weight) were anes-thetized with 150 �l mixture of 2.5% ketamine (KobanyaiGyogyszerarugyar, Budapest, Hungary), 2% Rompun (xy-lazine; Haver, Shawnee, KS) in physiological saline. Thechick’s head was fixed in a Kopf (Tujunga, CA) stereotaxicframe and a burr hole was made in the skull with a dentaldrill. For injecting the anterograde pathway tracer Pha-L
(Vector, Burlingame, CA) we followed the method of Ger-fen and Sawchenko (1984). Reconstituted aliquots of thetracer were kept frozen until the day of the experiment. Aglass capillary was filled with a 2.5% solution of Pha-L (insaline) and lowered into the septum. Coordinates were asfollows: anteroposterior �2.5 mm from bregma, lateral�0.3–0.4 mm from bregma, and depth 4.5–4.6 mm fromdura (coordinates based on the chick brain atlas by Kuen-zel and Masson (1988) and modified according to our ex-perience from preliminary experiments).
For iontophoretic injection of tracer, direct current wasapplied for 20–30 minutes with an intensity of 5–7 �A, at7-second on/off cycles. The electrode was removed 5 min-utes after switching off the current. The scalp wound wasclosed with fast-setting tissue adhesive under antisepticconditions. Two weeks later the birds were deeply anes-thetized with an overdose of equithesin (0.85 g chloralhydrate, 0.21 g sodium pentobarbital, 8.6 ml propyleneglycol, 2.2 ml 100% ethanol, 6.7 ml distilled water, 0.25ml/100 g body weight) (Green, 1979) and perfused tran-scardially with 0.9% saline, followed by a mixture contain-ing 4% paraformaldehyde, 2.5% glutaraldehyde, and 0.2%picric acid in phosphate buffer (PB; 0.1 M, pH 7.3). Brainswere dissected out, postfixed for 2 hours in a similarfixative with no picric acid and left overnight in a 20%sucrose solution. Serial 75-�m-thick sections were cut on afreezing microtome (Zeiss Frigomobile) and every fifthsection was collected. They were washed several times inPB and Pha-L was visualized by immunocytochemistry.
Immunohistochemical procedure
Endogeneous peroxidase activity was blocked by a5-minute treatment with 1% H2O2. Sections were thenrinsed extensively with PB, followed by TBS (0.05 M Tris-HCl, pH 7.3, containing 0.9% NaCl). Nonspecific immuno-reactivity was suppressed by a 1-hour incubation in 20%normal goat serum (NGS) in TBS containing 0.5% TritonX-100 (TBS-Tx), followed by 10 minutes in 1% NGS inTBS-Tx. Sections were incubated for 48 hours with biotin-ylated anti-Phaseolus lectin antibody (Vector), diluted at1/200 in 1% NGS in TBS-Tx. After three washes in 1%NGS in TBS-Tx, sections were incubated for 3 hours inavidin-biotin-horseradish peroxidase complex (ABC Vec-tastain kit; Vector) diluted at 1/50 in 1% NGS in TBS.Three washes in TBS and two in Tris buffer (0.05 MTris-HCl, pH 8) followed thereafter. Peroxidase activitywas revealed by reacting the sections with nickel-intensified 3,3�-diaminobenzidine tetrahydrochloride(DAB, 15 mg; Sigma, St. Louis, MO; 250 mg ammoniumnickel sulfate in 25 ml Tris buffer). After a 5-minutepreincubation, H2O2 was added at a final dilution of0.01%. The reaction was visually monitored and stoppedwhen the injection site became visible, usually 4-5 min-utes later, by rinsing first with Tris buffer, then with PB.Sections were mounted on gelatin-coated glass slides, de-hydrated, and mounted in DPX.
Charting
The contours of each section were drawn at low magni-fication with the help of a microscope specimen projector.The sections were observed under high magnification us-ing a Zeiss Axioskop or Olympus Vanox microscope, andthe fibers were manually charted on the templates. Thebrain nuclei were identified with the help of a chick brainatlas (Kuenzel and Masson, 1988). Identification of the
439SEPTAL EFFERENTS OF DOMESTIC CHICK

hypothalamic centers was made according to Kuenzel andvan Tienhoven (1982). The avian brain nomenclature isnow in a process of profound revision and some of theterms used in the present article will likely change soon. Asummary of the discussion on the topic is currently avail-able at the Avian Brain Nomenclature Exchange web site(http://jarvis.neuro.duke.edu./nomen/index.html). Finaldrawings of brain charts were prepared from scans ofmanual records using a professional drawing software(Canvas, Deneba).
Retrograde tracer injection
For a selective study of septohippocampal pathwaysfour 1-week-old chicks were anesthetized and prepared forintracranial surgery as described previously. Using a Kopfmicroinjection unit mounted on a stereotaxic frame, 0.1 �lTrue Blue (2% in distilled water, Sigma) were pressure-injected in the hippocampal formation. Coordinates wereas follows: anteroposterior �2.5 mm from bregma, lateral�0.3 mm from bregma, and depth 0.8 mm from dura(coordinates based on the chick brain atlas by Kuenzel andMasson, 1988). Following a survival period of 8–10 days,the birds were deeply anesthetized, perfused with fixative,and sectioned as described above for the Pha-L procedure.The sections were mounted in a 1:1 mixture of glyceroland 0.1 M PB and viewed and photographed using anOlympus BX 51 fluorescent microscope equipped with anOlympus DP 50 digital microphotographic camera.
RESULTS
Nine injections were placed in the septum: a dorsal SMinjection at the level of CA (brain (br) 21) (Figs. 1A, 2), twoprecommissural SL injections, one being ventral (br12;Figs. 1B, 3), the other dorsal (brLPO3) and four postcom-missural injections, one in the ventral part of SM, verynear to the midline (br22; Figs. 1C, 4), two centered in thedorsalmost (apical) part of SM, also extending to dorsolat-eral SL (brS2 and brS6; Figs. 5B, 6), and three (br11, br38,and br40) in the border region between the septum andthe thalamic anterior dorsomedial nucleus (DMA), cover-ing both structures to a variable degree. The br11 injec-tion was located at the level of the caudal end of subseptalorgan, mainly covered the ventral SL, caudal to the nu-cleus of the pallial commissure (nCPa), and reached therostral limit of DMA, but no neurons were stained in thelatter area. The large and more caudal br38 injectioninvolved SM, SL, habenular nuclei, and rostral dorsalDMA. The br40 site was restricted to the caudal SL, whereit gives way to the corticohabenular and corticoseptaltracts (CHCS). This injection also included a very smallpart of the rostromedial DMA, near the ependyma. Twocontrol injections were placed in nCPa (br20) and thedorsal preoptic region (PO) at the level of CA (br71; Figs.1D, 7), respectively.
Septal projections
Telencephalic projections. Lateral and medial septalneurons were found to project to several septal areas.Local ipsilateral varicose fibers coursed caudalward inboth ipsilateral SM (Figs. 2D,E, 3B,C, 6B–E) and SL (Figs.4A, 6B–E), independent of the location of the injectionsite. They branched into a loose and extended terminalplexus with varicosities and terminal boutons. A modestcontralateral projection was also evident. Fibers in the
contralateral SM originated from both ventral, pre/postcommissural SM and SL, and apical septum (Fig. 6E).Only the ventral SM projected to contralateral SL. Rarefibers in ipsilateral nCPa were present only in br22 (in-jection site in SM near nCPa). Fibers in contralateralnCPa were found only in br11 (septothalamic junctioninjection). The nucleus of the diagonal band of Broca(FDB) received some efferents from both SM (Fig. 2B) andSL (those arising from the apical septum being particu-larly heavy; Fig. 6A) with the exception of caudal SL.Pha-L-labeled fibers were seen coursing in TSM (Figs. 2B,6A), medial forebrain bundle. Some fibers from SL left theseptal area through CHCS.
Septal projections to the basal ganglia were restricted toPVT (Figs. 2A–C, 3A, 6A,B) and LPO. In PVT, ipsilateraland contralateral fibers, which originated from both SMand SL often coursed along the septomesencephalic tract(TSM; Fig. 8A) and sometimes in an uncharted area be-tween TSM and the lateral forebrain bundle (FPL), whichcould belong to PVT (Fig. 8B). They rarely branched.These fibers were heavily beaded and bore terminal bou-tons (Fig. 8B). Rare contralateral PVT fibers arose frompre- and postcommissural dorsal and ventral SL. A smallipsilateral projection to the rostral LPO originated fromventral SL, although, in two cases (br38, br12), a dorsalthalamic contribution could not be ruled out either.
A very sparse projection in the ipsilateral nucleus ac-cumbens (Ac) and olfactory tubercle (TO) arose in SL andSM. It has to be noted that the term Ac used on the chartsand in the text refers to both the lateral part of the bednucleus of the stria terminalis (BSTl) and Ac proper (seealso Discussion). The caudal part of the ipsilateral arch-istriatum intermedium pars ventralis (Aiv) and the ven-tral posterior archistriatum (Ap) received a small SL pro-jection. A modest projection to the ipsilateralarchistriatum intermedium also arise from the apical sep-tum (Fig. 6E). Other telencephalic projections originateexclusively from SL. Some ipsilateral fibers projected tothe nucleus taeniae (Tn; Figs. 3C, 8C). Fibers werescarcely seen in the ipsilateral piriform cortex (CPi) and inthe ipsilateral neostriatum (N; Fig. 3E). The taenial fiberswere very thin and heavily varicose (Fig. 8C). Sparseipsilateral fibers in anterior N and rare contralateral fi-bers in N arose from dorsal SL. A modest but distinct fiberinput arising from the apical SM/dorsal SL was observedin the V-shaped area of hippocampus (Figs. 5A, 6E,F). Inthe same brain, we also observed a terminal field of pro-jections lateral to the nucleus of the pallial commissure(nCPa; Fig. 6C) .
Diencephalic projections. The bulk of septal projec-tions was directed to the ipsilateral hypothalamus (Figs.2C–H, 3A–H, 6B–F). Fibers were found in the anterior PO(Figs. 2D, 3B, 6C), medial preoptic nucleus (POM, Fig.6A), hypothalamic periventricular nucleus (PHN; Figs.2D–G, 3C–F, 4B, 6D, 9D), hypothalamic anterior nucleus(AM; Figs. 2C,D, 3A, 9F), preoptic periventricular nucleus(POP; Fig. 2C), lateral hypothalamus (LHy; Figs. 2C–F,3C,D, 4A, 6C,E, 9A,B), medial hypothalamic area (mHy;Fig. 9C), the paraventricular nucleus (PVN; Figs. 2D,E,3B–D), suprachiasmatic nucleus (SCN; Fig. 3A) (the termSCN refers to the medial hypothalamic retinorecipientnucleus in the updated nomenclature; see Discussion),hypothalamic dorsal area (DHA; Figs. 2E–G, 3C,D, 6E),hypothalamic inferior nucleus (IH; Figs. 2H, 3F) and lat-eral mammillary nucleus (ML). Except in the case of
440 C.M. MONTAGNESE ET AL.

smallest injection site (br40), ipsilateral fibers were alsopresent in the external and ventral supraoptic nuclei(SOe, SOv; Fig. 2B), hypothalamic ventromedial nucleus
(VMN; Figs. 2E, 3E), hypothalamic dorsomedial nucleus(DMN; Figs. 2H, 3F, 6F), premammillary nucleus (PMN),infundibular nucleus (IN), and median eminence (ME;
Fig. 1. Micrographs of ionto-phoresis sites of Phaseolus lectinin the medial septal nucleus atcommissural level (A, brain 21),ventral lateral septal nucleus atprecommissural level (B, brain12), medial septal nucleus at post-commissural level (C, brain 22)and in the dorsal preoptic region(D, brain 71). The correspondingpatterns of fiber distribution arerepresented in Figures 2–5, re-spectively. Note that in D the an-terior commissure (CA), whichcrosses the injection site, remainsdevoid of staining. Scale bar � 100�m.
441SEPTAL EFFERENTS OF DOMESTIC CHICK
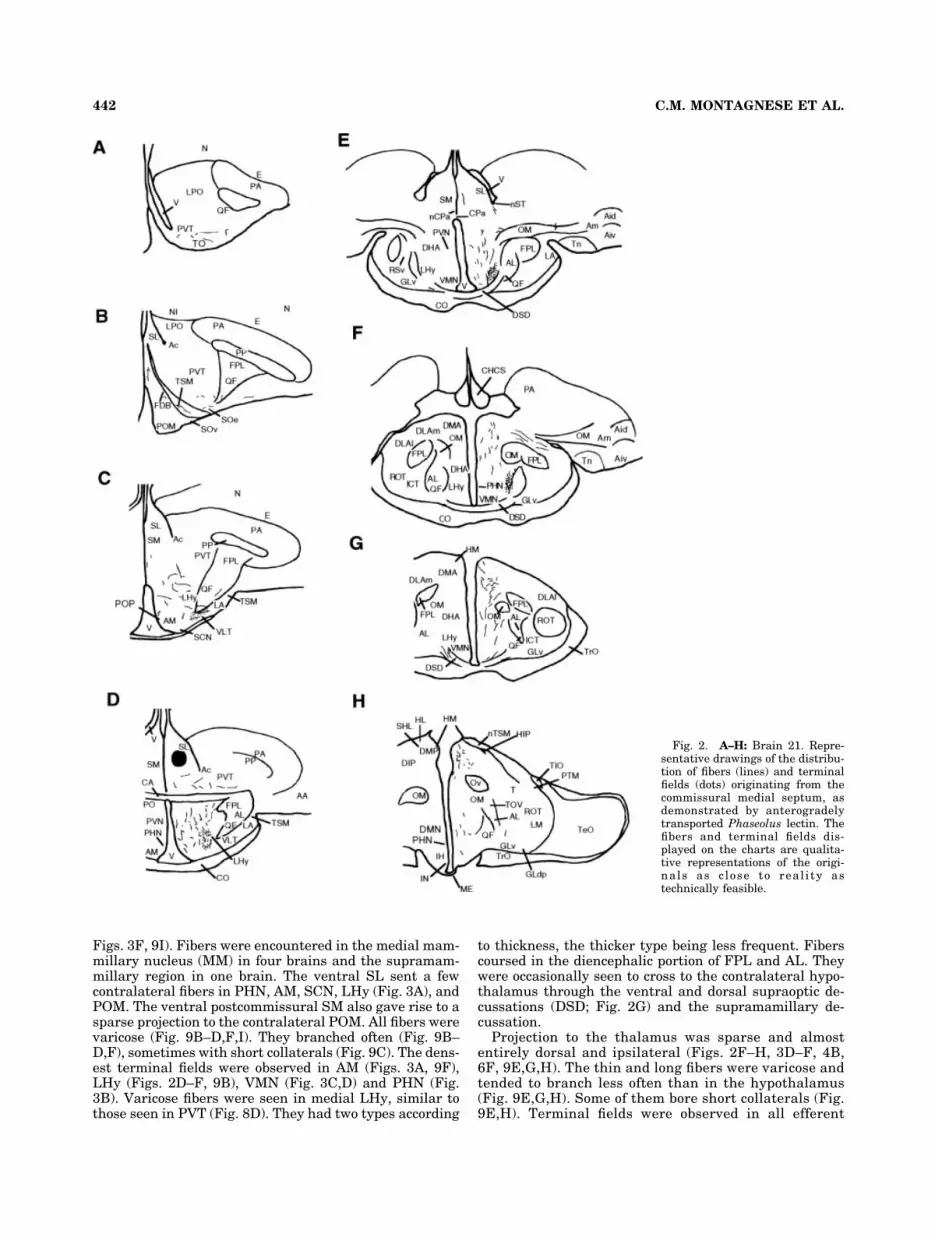
Figs. 3F, 9I). Fibers were encountered in the medial mam-millary nucleus (MM) in four brains and the supramam-millary region in one brain. The ventral SL sent a fewcontralateral fibers in PHN, AM, SCN, LHy (Fig. 3A), andPOM. The ventral postcommissural SM also gave rise to asparse projection to the contralateral POM. All fibers werevaricose (Fig. 9B–D,F,I). They branched often (Fig. 9B–D,F), sometimes with short collaterals (Fig. 9C). The dens-est terminal fields were observed in AM (Figs. 3A, 9F),LHy (Figs. 2D–F, 9B), VMN (Fig. 3C,D) and PHN (Fig.3B). Varicose fibers were seen in medial LHy, similar tothose seen in PVT (Fig. 8D). They had two types according
to thickness, the thicker type being less frequent. Fiberscoursed in the diencephalic portion of FPL and AL. Theywere occasionally seen to cross to the contralateral hypo-thalamus through the ventral and dorsal supraoptic de-cussations (DSD; Fig. 2G) and the supramamillary de-cussation.
Projection to the thalamus was sparse and almostentirely dorsal and ipsilateral (Figs. 2F–H, 3D–F, 4B,6F, 9E,G,H). The thin and long fibers were varicose andtended to branch less often than in the hypothalamus(Fig. 9E,G,H). Some of them bore short collaterals (Fig.9E,H). Terminal fields were observed in all efferent
Fig. 2. A–H: Brain 21. Repre-sentative drawings of the distribu-tion of fibers (lines) and terminalfields (dots) originating from thecommissural medial septum, asdemonstrated by anterogradelytransported Phaseolus lectin. Thefibers and terminal fields dis-played on the charts are qualita-tive representations of the origi-nals as close to real i ty astechnically feasible.
442 C.M. MONTAGNESE ET AL.

target areas. Both SM and SL projected to DMA (Figs.2F,G, 3D,E, 4A, 9E,G), fibers from SM being more nu-merous (compare Fig. 9E,G). Both nuclei sent afferentsto the thalamic posterior dorsomedial (DMP; Figs. 2H,3F, 6F), and dorsointermediate (DIP; Fig. 2H) nuclei,the ansa lenticularis (AL) and its nucleus and, moresparsely, to the nucleus paramedianus internus andthalamic ventrolateral nucleus (VLT, Fig. 2C). Fibers inthe anterior dorsolateral nucleus pars medialis andlateralis (DLAm and DLAl; Fig. 2F) originated fromrostral and mid-level SL and SM. A small area of SL
situated near CA sent some fibers through FPL toDLAm. These fibers put out short collaterals insideFPL. Contralateral fibers were very rare. The dorsaland ventral precommissural SL projected on the con-tralateral VLT and DMP, respectively. ContralateralDMA fibers seemed to come from the ventral postcom-missural SL and SM (Fig. 4B).
Both septal nuclei sparsely projected to the medial ha-benular (HM) and medial subhabenular (SHM) nuclei.Fibers in the lateral habenular nucleus (HL) were seen inbr11 and br38 only.
Fig. 3. A–F: Brain 12. Repre-sentative drawings of the distribu-tion of fibers (lines) and terminalfields (dots) originating from theprecommissural lateral septum, asdemonstrated by anterogradelytransported Phaseolus lectin. Onlythe main differences with Figure 2are represented.
443SEPTAL EFFERENTS OF DOMESTIC CHICK

Mesencephalic and brainstem projections. Rareand ipsilateral fibers projected to the midbrain centralgray (GCt) and the ventral tegmental area of Tsai (AVT,Fig. 6H). In brLPO3 (dorsal precommissural SL; Fig 8E),fibers were varicose and branching, with short collaterals.They were exclusively found in the rostral GCt. In br21(dorsal SM), fibers distributed along the entire rostrocau-dal extent of GCt. In the caudal GCt, fibers were very thin,rarely branching, and extending laterally over a long dis-tance. The fibers projecting to AVT originated from dorsalSM (br21), apical septum (Fig. 6H) and ventral postcom-missural SL (br11). The projection to AVT may have comefrom a restricted portion of SM, since in br11 the caudalinjection site, albeit centered essentially on the ventralSL, also included a small area of SM and DMA (diffusionof tracer). A further important contingent of projectionswas seen in the area including and surrounding the nu-cleus mesencephalicus profundus pars ventralis (MPv),recently defined as the pedunculopontine nucleus (PPN)(Medina and Reiner, 1994) arising from the dorsalmost(apical) septum (Fig. 6G).
Projections of the nucleus of the pallialcommissure and the preoptic region
Phaseolus injection in nCPa resulted in efferent fibersand axon terminals confined to the septal nuclei (SL, SM,FDB, CHCS), and rostral hypothalamus (PO, POM, AM,SOv). A few fibers also coursed in TSM.
Injection in PO was placed at the level of CA (Figs. 1D,7). The Pha-L immunolabeling was most prominent ven-tral to the commissure (Fig. 1D). The commissure wasdevoid of immunoreactivity, splitting the injection siteinto two visible deposits, confirming that Pha-L was nottaken up by passing myelinated fibers. In the telenceph-alon, the fibers were all ipsilateral and situated in SM andSL (Fig. 7C,D), PVT (Fig. 7A,B), LPO (Fig. 7A), TO, andAc. In PVT, two types of fibers were encountered: somewere thin and long, others were thick and short (Fig. 8D).
Fig. 4. A,B: Brain 22. Two representative drawings of the distri-bution of fibers (lines) and terminal fields (dots) originating from thepostcommissural medial septum, as demonstrated by anterogradelytransported Phaseolus lectin. Only the main differences with Figure 2are represented.
Fig. 5. A,B: Brain S6. Hippocampal afferent labeled by antero-gradely transported Phaseolus lectin injected in dorsalmost (apicalseptum). The varicose fiber (arrows) leaves the fiber tract (asterisk).Scale bar � 10 �m. Micrograph of the injection site of Phaseolus lectinin the apical septum. Scale bar � 100 �m.
444 C.M. MONTAGNESE ET AL.

Both types branched and terminated in PVT (Fig. 7A,B).Their termination field extended to the junction betweenthe septum and PVT. In LPO, the fibers were thin andbranched off before terminating, in the rostral ventrome-dial and central parts of the nucleus (Fig. 7A). Fibers wererare in TO, SM, and SL.
Most diencephalic efferent fibers left the injection sitemediocaudally, forming a small tract toward the medial
hypothalamus (Fig. 7B–I). The lateral hypothalamus re-ceived a more modest innervation (Fig. 7C–I). Thin andvaricose fibers and terminals were encountered in POM(Fig. 7B), AM (Fig. 7C), POP (Fig. 7A), PHN (Fig. 7C–G),SOe (Fig. 7A), PVN (Fig. 7C–E), LHy (Fig. 7B–G), DHA(Fig. 7E–F), VMN, DMN (Fig. 7G), IH (Fig. 7G,H), IN (Fig.7H,I), PMN (Fig. 7H), MM (Fig. 7H,I), ML (Fig. 7H),posterior nucleus of AL and nucleus c of Rendahl (nI; Fig.
Fig. 6. A–H: Brain S6. Repre-sentative drawings of the distribu-tion of fibers originating from thedorsalmost (apical) septum, asdemonstrated by anterogradelytransported Phaseolus lectin.
445SEPTAL EFFERENTS OF DOMESTIC CHICK

7H). Contralateral fibers were found in PVN, IN (Fig. 7H),PMN (Fig. 7H), and ML. Thalamic fibers were rare andrestricted to the dorsal areas, i.e., DMA (Fig. 7F), DMP(Fig. 7G), DIP (Fig. 7G), and the dorsal superior reticularnucleus (RSd). These fibers were long and terminated inthe dorsal thalamus after a few branchings. Fibers alsoprojected to the ipsilateral habenular nuclei (Fig. 7F–H).
Ipsilateral mesencephalic and brainstem projectionsand terminal fields were seen in the pretectal area (AP;Fig. 7I), medial spiriform nucleus, optic tectum (TeO; Fig.7J), stratum griseum periventriculare (SGP; Fig. 7I), in-tercollicular nucleus (ICo; Fig. 7J–K), GCt (Fig. 7K), AVT,MPv (PPN), TPc/SN (Fig. 7K), LoC (Fig. 7K), SCd (Fig.7L),SCv (Fig. 7M), both lateral and medial mesencephalic
Fig. 7. A–M: Brain 71. Repre-sentative drawings of the distribu-tion of fibers (lines) and terminalfields (dots) originating from thedorsal anterior preoptic region, asdemonstrated by anterogradelytransported Phaseolus lectin.
446 C.M. MONTAGNESE ET AL.

reticular formation (FRL; Fig. 7J; FRM), nucleus of thetectal commissure (CT), nucleus of the lateral lemniscus,pontine reticular formation, in particular the gigantocel-lular caudal pontine reticular nucleus (RPgc; Fig. 7M) andpontine reticular nucleus, trapezoid body (CTz; Fig. 7L),and descending vestibular nucleus. Fibers were alsopresent in the pretecto-subpretectal tract and vestibulo-mesencephalic tract (TVM; Fig. 7K). Rare contralateralfibers were encountered in the nucleus of the basal opticroot, GCt, CT nucleus, nucleus of the lateral lemniscus,and the parvocellular reticular nucleus. Abundant thinfibers branched extensively in TPc/SN before terminating(Fig. 7K). Short and thick fibers were distributed in the
entire GCt, whereas long and thin axons branched in looseplexuses in caudal GCt, laterally entering ICo (Fig. 7J,K).
Septohippocampal connections as revealedby retrograde pathway tracing
Of the four injections attempted, we rejected two caseswhere most of the tracer was dissipated in the lateral ven-tricle. Both of the remaining two birds had their tracerdeposits in the hippocampus, in particular the V-shapedarea, one of them being precisely centered there (Fig. 10A),whereas, in the other specimen, the True Blue injection alsocovered parts of the Wulst. Due to the well-known afferentprojections of the latter, only those components of retro-
Figure 7 (Continued)
447SEPTAL EFFERENTS OF DOMESTIC CHICK

gradely filled neurons were taken into account, which couldbe clearly attributed to the common hippocampal target.Retrogradely labeled cells were found sporadically in SM butthe vast majority of these were located in the nucleus ofdiagonal band (Fig. 10B,C). Although a comprehensive de-
scription of connections was not attempted in this part of thestudy (we focused our attention on the septohippocampalpathways) we also observed labeled neurons in the olfactorybulb, ventral paleostriatum, preoptic area, subhabenular re-gion, and the central gray.
Fig. 8. Telencephalic afferents labeled by anterogradely trans-ported Phaseolus lectin injected in the septum. A: Fibers from SL,running in the ventral part of the ventral paleostriatum (PVT) alongthe septomesencephalic tract (TSM). The varicosities (arrowhead)suggest that these fibers may be forming synapses en passant. B: Fi-bers running between the lateral forebrain bundle (FPL) and theseptomesencephalic tract (TSM), in an uncharted area, presumablybelonging to PVT. Originating in SM, they possibly establish synaptic
contacts with terminal boutons (arrowhead) and varicosities (openarrowhead). C: Varicose (arrowhead) fibers in the ventral nucleustaeniae (Tn), originating from SL. D: Two types of fibers from thedorsal preoptic region are visible in PVT: some are thick and varicose(arrow), others thin and nonvaricose (arrowhead). E: Fibers from SLare branching (arrows) in the midbrain central gray (GCt). Scalebars � 10 �m.
448 C.M. MONTAGNESE ET AL.

DISCUSSION
We report here the connectivity of the septal areas of thechick, which may play a pivotal role as an interface be-tween the limbic system and other parts of the brain. Thepresent findings confirm and extend the observationsmade in pigeons for the septal nuclei (Krayniak and Sie-gel, 1978). Discrepancies between our work and those ofprevious authors may arise from several sources. First,the bird species used in both studies belong to two differ-ent orders (Galliformes and Columbiformes; Krayniak andSiegel, 1978). Huber and Crosby (1929) observed differ-ences in the size of the septum of parakeet, dove, sparrow,duck, and chick. Septum volume in food-storing parids islarger than in nonstoring parids (Shiflett et al., 2002).Differences in the distribution of neurotensin and sub-stance P immunoreactivity have been observed, respec-tively, between pigeons and chicks (Atoji et al., 1996), andfood-storing and nonstoring birds (Gould et al., 2001).Differences in the density of alpha- and beta-adrenoceptors have also been reported between pigeonsand chicks (Fernandez-Lopez et al., 1997).
The use of different tracing methods may be a secondsource of discrepancy. Krayniak and Siegel (1978) studiedthe septal connections by the retrogradely transportedhorseradish peroxidase and the anterograde tracer triti-ated leucine, which does not distinguish between axons ofpassage and axon terminals. We preferred iontophoreticinjections of the kidney bean lectin Phaseolus vulgarisleucoagglutinin detected by the immunoperoxidase tech-nique. This technique is rather sensitive and reveals fur-ther details of axonal projections (for further discussion,see Smith, 1992). In addition, transneuronal transport ofPha-L has not been reported, unlike in the case of, e.g.,horseradish peroxidase conjugated wheat germ agglutinin(Spatz, 1989). The size of our injections was generallysmaller than that used in previous studies. This allows theconnectivity of the examined nuclei to be described with abetter spatial resolution. On the other hand, the rarestprojections may be missed, since a restricted number ofneurons will capture the marker, leading to false-negativeresults. However, an absence of marker in a given struc-ture cannot have been caused by an insufficient time fortransport, since in one case labeling was observed as far asthe beginning of the spinal cord.
A third source of variation may come from developmen-tal variations and growth rearrangements, as observed inthe thalamic projections of the young zebra finch (Johnsonand Bottjer, 1992), or in the catecholaminergic innerva-tion of the nucleus accumbens and caudate putamen inmammals (see Smeets and Gonzalez, 2000). This, how-ever, is unlikely, since chicks are precocial birds and allmajor connections are likely to be established at hatching.
Comparison with earlier studies
Before discussing our results, it should be noted that weused a different terminology from that of Krayniak andSiegel (1978). Their “dorsal septum” corresponded to theseptal nuclei proper, i.e., SM and SL. We focused ourattention on the efferents of these two nuclei. Our “ventralseptum” corresponds to the ventral half of Krayniak andSiegel’s “dorsal septum.” Their “ventral septum” essen-tially corresponds to the nucleus of the diagonal band(FDB), which we did not study. Concerning the term “ac-cumbens,” the site previously defined in the chick atlas
(Kuenzel and Masson, 1988) as Ac is now considered asBSTl, whereas the Ac proper is coextensive with the me-dioventral LPO (Veenman et al., 1995b; Mezey and Csil-lag, 2002). Lacking specific staining (e.g., DARPP-32) inthe present study, such a distinction would have beenimpractical. Therefore, the cover term Ac was used todesignate Ac-BSTl collectively. In the old atlas of thechick, the term SCN covered the sites which are nowknown as the medial hypothalamic retinorecipient nu-cleus and the more caudally situated lateral hypothalamicretinorecipient nucleus, medial to Glv (Shimizu et al.,1994; Wallman et al., 1994). Our term SCN refers to themedial hypothalamic retinorecipient nucleus.
The major septal projections were similar to those ob-served in the pigeon by Krayniak and Siegel (1978), thatis, to the diencephalon (especially the lateral and periven-tricular hypothalamus, and dorsomedial thalamus), andto the midbrain tegmentum. Nevertheless, we showed ev-idence that additional areas also received afferents fromSM and/or SL. First, all septal nuclei were ipsilaterallyand contralaterally connected to each other. This fact wasconfirmed by the nCPa projection pattern, which was sim-ilar to that of SM and SL, but more restricted. This recip-rocal connection may also involve FDB, another septalnucleus, since we observed fibers terminating in that area,and Krayniak and Siegel (1978) noted ascending fibersfrom FDB coursing through SM.
A second novel finding is the presence of a few fibersprojecting to a number of ipsilateral and (rarely) con-tralateral telencephalic areas (PVT, LPO, Ac, TO, CPi,Aiv, Ap, Tn, N, and caudal N). Although some of theseregions received input from both SL and SM (e.g., PVT,Ac, Tn, FDB, TO), careful analysis suggested that SM andSL may possess specific sets of telencephalic efferents.Thus, CPi and Ap afferents seemed to arise exclusivelyfrom SL, whereas nCPa afferents arose from SM. In pi-geons, a very small ipsilateral projection to the hippocam-pus arises from sporadic neurons located mainly in theventral caudal SM (Benowitz and Karten, 1976; Casini etal., 1986; Atoji et al., 2002). Krayniak and Siegel (1978)did not see this connection. In our experiments very fewfibers from dorsal SL reach the hippocampal formation.These discrepancies might be explained by the small num-ber of efferents, the different tracers used, or a speciesdifference.
A third difference concerned the distribution of fiberswithin the diencephalon. We identified fibers ipsilaterallyin the supraoptic nuclei, PO, and anterior hypothalamus,i.e., more rostrally than reported by Krayniak and Siegel(1978). Hitherto unreported projections were detected inmedial hypothalamic nuclei (VMN, PHN), ME, and habe-nular nuclei, and in the contralateral diencephalon. Theyappeared similar to those observed by Krayniak and Sie-gel (1978) to arise from FDB, suggesting the existence of aconvergent innervation from both septal areas. A conver-gent septal projection to ME arises from SL and FDBnuclei in the dove (Knapp and Silver, 1995). Finally, un-like Krayniak and Siegel (1978), we did not find fibers inthe thalamic superior reticular nucleus. The distributionof fibers arising from SL or SM did not conspicuously differwithin either the hypothalamus or the thalamus.
A fourth discrepancy occurred in the midbrain. We iden-tified a small projection on AVT and MPv (PPN) but nonein the reticular formation or the perirubral region, unlikeKrayniak and Siegel (1978).
449SEPTAL EFFERENTS OF DOMESTIC CHICK

Figure 9

Comparison with the mammalian septum
The mammalian and avian septa appear homologous ondevelopmental grounds. Both genes dlx-2 and nkx-2.1,which are characteristic for the subpallium, are expressedin the mouse and chick septa as well as in the mouseamygdala and the equivalent part of the chick archistria-tum. Similarly, the pallial marker genes Pax-6, tbr-1, andemx-1 are expressed in a dorsal part of the avian septumas in some part of the mouse septum (Puelles et al., 1999).
In mammals, the projection of SL is topographicallyorganized along a dorsoventral and a rostrocaudal gradi-ent. The telencephalic efferents invade ventral pallidalregions (SM/DBB, substantia innominata, as well as SLitself), nucleus accumbens, and bed nucleus of the striaterminalis (Risold and Swanson, 1997b). Some efferentsenter SM (Leranth et al., 1992). There is also a smallprojection to the hippocampus, anterior amygdaloid area,amygdalar nuclei, and rare fibers in the olfactory pedun-cle, and dorsal taenia tecta (Risold and Swanson, 1997b).Fibers from SL invade massively the preoptic and anteriorhypothalamic areas and also project to the periventricu-lar, lateral and posterior hypothalamus (Meibach and Sie-gel, 1977a; Krayniak et al., 1980; Wayner et al., 1983;Chiba and Murata, 1985; Shibata, 1989; Varoqueaux andPoulain, 1994; Thompson and Swanson, 1998). The con-nection between the ventral SL and periventricular zoneis reciprocal. A small to moderate projection reaches themidline thalamus and ventral regions of the thalamus(Meibach and Siegel, 1977a; Risold and Swanson, 1997b).A few axons project to the lateral habenula (Meibach andSiegel, 1977a; Risold and Swanson, 1997b). Caudally, SLfibers terminate in the midbrain, in particular, in theventral tegmental area (VTA) and the periaqueductal grayand raphe nuclei (Risold and Swanson, 1997b).
Both SM and DBB project to the hippocampal formation(Meibach and Siegel, 1977a; Amaral and Cowan, 1980;Krayniak et al., 1980; Woolf et al., 1984; Gaykema et al.,1990). Some additional projections to the cortex, includingthe entorhinal cortex, arise from SM (Alonso and Kohler,1984; Woolf et al., 1984; Gaykema et al., 1990). Efferents
from DBB reach the amygdala (Meibach and Siegel.,1977a; Krayniak et al., 1980; Ottersen, 1980; Russchen,1982; Woolf et al., 1984; Leranth et al., 1999). Dience-phalic projections of DBB and SM include the medialpreoptic area (POA), periventricular and posterior hypo-thalamus (Meibach and Siegel, 1977a; Krayniak et al.,1980; Wayner et al., 1983; Wouterlood et al., 1988), mam-millary and supramammillary regions (Borhegyi andFreund, 1998; Shibata, 1989; Leranth et al., 1999), antero-medial and paratenial thalamus (Meibach and Siegel,1977; Parent et al., 1988), and habenular nuclei (Herken-ham and Nauta, 1977; Parent et al., 1981). Efferents fromDBB also terminate in VTA (Meibach and Siegel, 1977a;Krayniak et al., 1980). The dorsal tegmental nucleus andthe interpeduncular nucleus receive afferents from theseptum and DBB (Krayniak et al., 1980; Hamill and Ja-cobowitz, 1984; Liu et al., 1984; Vertes and Fass, 1988).
When compared to mammals, avian septal areas aregenerally less well defined, but some parallelisms could
Fig. 10. Brain S8 micrograph showing the tracer deposit of Trueblue in the V-shaped area of the hippocampus (HP). Low-power mi-crograph of retrogradely labeled cells located in FDB. High-powermicrograph of a retrogradely labeled cell located in FDB. Scale bars inA,B �100 �m; in C � 20 �m.
Fig. 9. Diencephalic afferents labeled by anterogradely trans-ported Phaseolus lectin injected in the septum. Directional markings:m. medial; d. dorsal. A: Low-power micrograph of the hypothalamusfrom a bird in which Phaseolus lectin was injected in SL. An enlargedview of the framed area is presented in B. B: Enlargement of theframed area in A, showing the dense network of anterogradely labeledfibers in LHy. These fibers are branching (arrow) and establish syn-apses en passant (arrowhead). C: A fiber from SM is branching (ar-rows) in the lateral part of the medial hypothalamus (mHy). Note thethickness of fibers and the presence of terminal boutons (arrowhead).D: Varicose fiber from SL, coursing and branching (arrow) in thehypothalamic periventricular nucleus (PHN). v third ventricle. E: Fi-bers from SL, coursing and branching (arrow) in the DMA. Note thepresence of terminal boutons (arrowhead). F: Fibers from SM in thehypothalamic anterior nucleus (AM). The fibers are branching (arrow)and display synapses en passant (arrowhead). G: Fibers from SMcoursing and branching (arrow) in the DMA. They probably establishsynapses en passant at the level of varicosities (arrowhead). H: Fibersfrom SM passing in the lateral forebrain bundle (FPL) towards themagnocellular part of the thalamic anterior dorsolateral nucleus(DLAmc). The fibers bear varicosities (open arrowhead) and terminalboutons (arrowhead). I: Varicose (arrowhead) fiber from SL in themedian eminence (ME). v third ventricle. Scale bars in A � 100 �m;in B–I � 10 �m.
451SEPTAL EFFERENTS OF DOMESTIC CHICK

nevertheless be established. The distribution of efferentsfrom the avian septal nuclei (present data; Krayniak andSiegel, 1978) appeared fairly similar to their mammaliancounterparts. In both vertebrate classes, 1) septal nucleiare interconnected, SL projecting on SM and FDB (Krayn-iak and Siegel, 1978, present data) or its mammaliancounterpart DBB (Krayniak et al., 1980; Leranth et al.,1992); 2) septal projections on LHy and PO are extensive(mammals: Meibach and Siegel, 1977a; Krayniak et al.,1980; Wayner et al., 1983; Chiba and Murata, 1985; Risoldand Swanson, 1997b; birds: Krayniak and Siegel, 1978;Balthazart et al., 1994; present data). In birds, the effer-ents from SL tend to project more medially in the hypo-thalamus than the ones from SM. However, it is prema-ture to establish a firm topographical pattern, similar tothat observed by Risold and Swanson (1997b) in rats. 3) Inboth mammals and birds, SM projects to the supramam-millary regions (mammals: Borhegyi and Freund, 1998;Leranth et al., 1999; birds: present data). 4) Compared tothose from SL, the projections from SM reaching the mid-line thalamus, hypothalamus, and habenular nuclei aremodest (mammals: Meibach and Siegel, 1977a; Herken-ham and Nauta, 1977; Krayniak et al., 1980; Parent et al.,1981, 1988; birds: Krayniak and Siegel, 1978; presentdata). 5) Some septal projections reach the midbrain cen-tral gray and ventral tegmental area (Risold and Swan-son, 1997b; present data).
Nevertheless, avian and mammalian septal efferentsare not entirely similar in their organization. In birds, SMand SL projection to the hippocampal formation is ratherweak (Benowitz and Karten, 1976; Casini et al., 1986;Atoji et al., 2002; present data), FDB constituting the bulkof septal efferents to that area (Benowitz and Karten,1976; Krayniak and Siegel, 1978; Atoji et al., 2002). Con-versely, in mammals, SM projects massively on the den-tate gyrus and CA3 field of the hippocampal formation(Meibach and Siegel, 1977a; Krayniak et al., 1980; Amaraland Kurz, 1985; Gaykema et al., 1990). It has been pro-posed on the basis of its location, connectivity, and immu-noreactivity that the substance P-positive field of theparahippocampus is the avian counterpart of the mamma-lian entorhinal cortex (Erichsen et al., 1991; Szekely,1999). Neither Krayniak and Siegel (1978) nor our resultsshowed an input from SM to the area parahippocampalis.Such projection arises from FDB (Krayniak and Siegel,1978). In the chick, projection to the limbic archistriatum,which is the avian equivalent of the mammalian pallialamygdala, arises from SL, whereas in mammals the mainseptal afferents to the amygdala arise from DBB, theinput from SL being modest (Meibach and Siegel, 1977a;Krayniak et al., 1980; Ottersen, 1980; Russchen, 1982;Woolf et al., 1984; Leranth et al., 1999). In mammals,Gaykema et al. (1990) identified a projection from DBB tothe olfactory nuclei. This projection may be equivalent tothe observed avian septal projection on TO and perhapsalso to CPi. In mammals, SM and DBB send efferents todifferent subdivisions of the interpeduncular nucleus (Ha-mill and Jacobowitz, 1984; Vertes and Fass, 1988). Nosuch connections were observed in the present study.However, the habenular nuclei (the main source of inter-peduncular input) did receive a sparse projection fromboth septal nuclei.
Functional considerations
A number of avian studies have shown the importanceof the septal-hypothalamic axis in behavioral and endo-crine functions.
A direct fiber connection exists between the septum andthe adrenocorticotropic area of the hypothalamus (presentdata; Krayniak and Siegel, 1978), which is situated in theposterior medial and LHy (Bouille and Bayle, 1973a,b).The septal areas directly inhibit the pituitary-adrenocortical axis (Bouille and Bayle, 1975). In geese andcocks, lesions of the septal area induced hyperphagia(Robinzon et al., 1978) and a feedback mechanism hasbeen established between LHy and the septum (Felix andRoesch, 1984). The modulation of ingestive behavior bythe septal areas may be particularly important in migra-tory birds, possibly through the humoral control of meta-bolic activity (body weight regulation), as in mammals(King and Nance, 1986). A putative neurotransmitter in-volved in the modulation of ingestive behavior could beneuropeptide Y, as it is present in septal areas and itincreases food intake in white crowned sparrow (Richard-son et al., 1995).
The involvement of the septum in avian reproduction isexemplified by the presence of neurons synthesizinggonadotropin-releasing hormone (Panzica et al., 2001), sex-ually dimorphic distribution of vasotocin-immunoreactive fi-bers in SL (Jurkevich et al., 1996; Panzica et al., 2001), densepopulations of estrogen-receptors (Panzica et al., 2001), andseptal projection to POA (present data; Krayniak and Siegel,1978). Vasotocin is involved in oviposition, aggressive, andsinging component of the male reproductive behavior (Good-son, 1998a,b; Goodson and Adkins-Regan, 1999).
Septal lesions facilitate male courtship behavior andvocalization in ring doves (Streptopelia risoria; Cooperand Erickson, 1976). In black-capped chickadees, the food-storing behavior is sensitive to the social organizationconcomitant with the seasonal variation of septal volume(Shiflett et al., 2002). The increase in septal volume ismaximal when the storing behavior is at its peak of activ-ity. The role of the septum in intraspecific aggressiondepends on the social organization of the studied species(Goodson et al., 1999). The modulation of this behavior ismediated by septal vasotocin and vasointestinal peptide(Goodson, 1998a,b; Goodson and Adkins-Regan, 1999).The role of avian septum in the defensive behavior is lesswell established than in mammals (Kask et al., 2001).Cohen and Goff (1978) have shown that, in pigeon, theextensive destruction of septal areas has no effect on heartrate conditioning. A projection from the archistriatum tothe septal nuclei has been demonstrated in the pigeon(Zeier and Karten, 1971) and the chick (Davies et al.,1997). Conversely, part of the limbic (amygdala equiva-lent) archistriatum receives afferent septal fibers (presentstudy). Within that frame, the direct septal connectionwith the thalamic nucleus DMA, reported in the chick,may be particularly important (Montagnese et al., 2003),since part of this nucleus could be the equivalent of themammalian thalamic mediodorsal nucleus. In mammals,this nucleus receives septal afferents (Hreib et al., 1988)and is connected with the prefrontal cortex and LHy, twomajor sites related to the control of aggression (Brutus etal., 1984).
Septal areas may be directly or indirectly involved inlocomotor modulation through their connections with the
452 C.M. MONTAGNESE ET AL.

lateral hypothalamic zone, which in mammals is involvedin arousal and the initiation of goal-oriented behaviors(see Risold and Swanson, 1997b). In zebra finch, the an-terior hypothalamic/preoptic area has direct projection tothe midbrain vocal motor structure, the nucleus intercol-licularis (Berk and Butler, 1981; Briganti et al., 1996). Thedorsomedial thalamus, which receives a modest inputfrom the septal areas (present data), is also connected tothe basal ganglia (Kitt and Brauth, 1982; Wild, 1987;Brauth et al., 1994; Medina and Reiner, 1997; Montagneseet al., 2003). The septo-thalamo-basal ganglia circuitmight play a rather important role in courtship vocaliza-tion in song birds. In the zebra finch, a portion of thelateral septum projects to the thalamic DMP, an afferentof the vocal forebrain circuitry (Foster et al., 1997; Suthersand Margoliash, 2002) connected to area X (motor part ofvocalization; Brainard and Doupe, 2000).
Another important functional loop exists between theseptal area and the mesolimbic centers. A projection fromAVT and TPc/SN to SM and SL has been observed in thepigeon (Kitt and Brauth, 1986b). The septal nuclei of thepigeon also contain a dense plexus of catecholaminergicfibers (Wynne and Gunturkun, 1995; Durstewitz et al.,1999; Smeets and Gonzalez, 2001). In birds, septal nucleicould modulate the activity of basal ganglia indirectly,through AVT, or directly, via connections with PVT (ven-tral pallidum), as shown in the present report.
Septal areas might serve as an important relay of thesuprachiasmatic nucleus (SCN), mediating between theseasonal cyclicity of some behaviors and the related phys-iological changes. The presence of extraocular photorecep-tors has been demonstrated in the avian SL (cerebrospinalfluid contacting neurons of lateral septal organ, Silver etal., 1988; Saldanha et al., 1994), which sends afferents toSCN (birds: present data; mammals: Moga and Moore,1997). The septal influence may be directly exerted bysending photic clues to SCN and/or indirectly by modulat-ing the activity of SCN and the other areas connected toSCN.
The existence of a theta rhythm, similar to that of cats,has been shown by Siegel et al. (2000) in pigeons. Inmammals, the theta rhythm, which is characterized byhigh amplitude 4–8 Hz oscillations in the hippocampusduring locomotion, exploration, and other voluntary be-haviors (see Kahana et al., 2001) is generated in SM/DBB.Despite the lack of direct evidence, we may assume thatthe neural loop between the hippocampal formation andseptal areas is responsible for the generation of thetarhythm in birds. In both classes, interconnected septalareas receive a strong input from the hippocampal forma-tion (Risold and Swanson, 1997b; Szekely, 1999; Atoji etal., 2002), whereas a relatively weaker but significantprojection from FDB (and, to a lesser degree, from otherseptal subdivisions) to the hippocampal formation wasdemonstrated, resembling the SM/DBB projection inmammals (Krayniak and Siegel, 1978; Atoji et al., 2002;present study).
In birds the hippocampal formation is also involved inthe memory associated with foraging (Shiflett et al., 2002)and homing (Gagliardo et al., 1999) behaviors. As theintegrity of SM/DBB is critical for the acquisition of novelinformation, and the reinforcement and consolidation ofmemory (Leutgeb and Mizumori, 1999; Wu et al., 2000;Kahana et al., 2001), this may be the case also in birds.
In mammals the septal formation project to the mesoac-cumbens circuit both at the level of Ac and VTA. Themesoaccumbens dopaminergic limbic system is known tofacilitate the exploratory-seeking arousal and the flexibleapproach responses and learning in an unfamiliar situa-tion, by evaluating the importance of the environmentalstimuli (Ikemoto and Panksepp, 1999). In birds, a similarcircuit might exist; we demonstrated a septal projection toAc as well as AVT, which are also connected to each other(Mezey and Csillag, 2002). As the septal formation is ableto modulate a wide variety of behaviors through its exten-sive connection with limbic and nonlimbic systems, it mayassist the mesolimbic system in the evaluation of novelsituations and learning.
Nucleus of the pallial commissure andpreoptic area
Panzica et al. (2001) suggested that the nucleus of thepallial commissure in quail is equivalent to the mamma-lian median preoptic nucleus. Our injection in nCPa wasvery small and resulted in a restricted pattern of projec-tion which does not allow us to comment on this issue.
The injection site in PO was located ventral to therostral CA, in an uncharted and ill-defined complex area,where POA and the anterior hypothalamic area come intocontact. It partly included nuclei B (a small round cellmass immediately rostral to CA), C (neurons encirclingand adhering to CA margin), and E (a broad cell mass,caudoventral to CA) of Yamauchi and Yasuda (1981). Thenucleus E may correspond to the anterior ventral area andpart of the medial preoptic nucleus of Huber and Crosby(1929). At the level of CA, POM is shifted dorsally to thearea ventral to CA, and, caudally, it partially overlaps AM(Yamauchi and Yasuda, 1981). The pattern of projectionobtained from this single injection was essentially similarto that arising from the medial preoptic region of the quail(Balthazart et al., 1994) and pigeon (Berk and Butler,1981). Although most fibers came from the PO region,some of the apparently PO-related fibers may have arisenfrom the border area between PO and the ventral septum.However, there were clear differences between the projec-tion patterns of PO and septal areas. A number of telen-cephalic regions (ipsilateral hyperstriatum, archistria-tum, N and CPi, as well as the contralateral SM, SL, andPVT) received fibers only from the septum. Basal gangliaafferents were more abundant after PO injection. Al-though both the septum and PO project on the medialdiencephalon, PO does not project on SCN and the ventralthalamus, but it does on RSd and RSv. Contralateralprojections from PO (but not the septum) are widespread.In the hypothalamus, preoptic fibers mostly invade themedial region, whereas they are less dense laterally. Theopposite pattern of distribution was observed followingseptal injections. In the thalamus, the afferents tend tospread more laterally after PO injection. Finally, POabundantly projects on numerous mesencephalic andbrainstem centers, in contrast to septal areas.
The overlap between the projections from septal nucleiand PO may subserve a dual control over the areas receiv-ing afferents from both centers. In both rodents (Smithand Flynn, 1980; Ferris et al., 1994) and birds (Barfield,1971), the anterior hypothalamic and preoptic areas areimportant sites for the regulation of aggressive and ago-nistic behaviors, which are also under septal control (seediscussion above). Since the septal nuclei also project on
453SEPTAL EFFERENTS OF DOMESTIC CHICK

PO, they could directly and indirectly control limbic cir-cuits, such as the mesolimbic loop. The pedunculopontinearea and AVT receive afferents from both septal and POcenters (present data; Fahrbach et al., 1986; Swanson etal., 1987). The same areas project back to both septum andPO (birds: Kitt and Brauth, 1986b; mammals: Becksteadet al., 1979). In mammals, the integrity of PO/VTA circuitappears to be of prime importance for the normal expres-sion of, e.g., maternal behavior (Numan and Smith, 1984).
ACKNOWLEDGMENTS
The authors thank Professor Jozsef Kiss for criticalreading of the article and helpful comments, and Dr. Las-zlo Negyessy for valuable advice in retrograde pathwaytracing. We thank photographer J. Kiss for preparing thephotomicrographs and G. Zachar for help in artwork ed-iting.
LITERATURE CITED
Acsady L, Arabadzisz D, Katona I, Freund TF. 1996. Topographic distri-bution of dorsal and median raphe neurons with hippocampal, septaland dual projection. Acta Biol Hung 47:9–19.
Alonso A, Kohler C. 1984. A study of the reciprocal connections between theseptum and the entorhinal area using anterograde and retrogradeaxonal transport methods in the rat brain. J Comp Neurol 225:327–343.
Amaral DG, Cowan WM. 1980. Subcortical afferents to the hippocampalformation in the monkey. J Comp Neurol 189:573–591.
Amaral DG, Kurz J. 1985. An analysis of the origins of the cholinergic andnoncholinergic septal projections to the hippocampal formation of therat. J Comp Neurol 240:37–59.
Atoji Y, Shibata N, Yamamoto Y, Suzuki Y. 1996. Distribution ofneurotensin-containing neurons in the central nervous system of thepigeon and the chicken. J Comp Neurol 375:187–211.
Atoji Y, Wild JM, Yamamoto Y, Suzuki Y. 2002. Intratelencephalic con-nections of the hippocampus in pigeons (Columba livia). J Comp Neurol447:177–199.
Balthazart J, Dupiereux V, Aste N, Viglietti-Panzica C, Barrese M, Pan-zica GC. 1994. Afferent and efferent connections of the sexually dimor-phic medial preoptic nucleus of the male quail revealed by in vitrotransport of DiI. Cell Tissue Res 276:455–475.
Barfield RJ. 1971. Activation of sexual and aggressive behavior by andro-gen implanted into the male ring dove brain. Endocrinology 89:1470–1476.
Beckstead RM, Domesick VB, Nauta WJ. 1979. Efferent connections of thesubstantia nigra and ventral tegmental area in the rat. Brain Res175:191–217.
Benowitz LI, Karten HJ. 1976. The tractus infundibuli and other afferentsto the parahippocampal region in the pigeon. Brain Res 102:174–180.
Berk ML, Butler AB. 1981. Efferent projections of the medial preopticnucleus and medial hypothalamus in the pigeon. J Comp Neurol 203:379–399.
Berk ML, Finkelstein JA. 1983. Long descending projections of the hypo-thalamus in the pigeon, Columba livia. J Comp Neurol 220:127–136.
Berk ML, Hawkin RF. 1985. Ascending projections of the mammilllaryregion in the pigeon: emphasis on telencephalic connections. J CompNeurol 239:330–340.
Borhegyi Z, Freund TF. 1998. Dual projection from the medial septum tothe supramammillary nucleus in the rat. Brain Res Bull 46:453–459
Bouille C, Bayle JD. 1973a. Experimental studies on the adrenocortico-tropic area in the pigeon hypothalamus. Neuroendocrinology 11:73–91.
Bouille C, Bayle JD. 1973b. Effect of hypothalamic stimulation onpituitary-adrenocortical activity in conscious unrestrained pigeons.Neuroendocrinology 12:284–294.
Bouille C, Bayle JD. 1975. Influence of septal nuclei on basal pituitary-adrenocortical function in birds. Neuroendocrinology 18:281–289.
Brainard MS, Doupe AJ. 2000. Interruption of a basal ganglia-forebraincircuit prevents plasticity of learned vocalisations. Nature 404:762–766.
Brauth SE, Heaton JT, Durand SE, Liang W, Hall WS. 1994. Functionalanatomy of forebrain auditory pathways in the budgerigar (Melopsit-tacus undulatus). Brain Behav Evol 44:210–233.
Briganti F, Beani L, Panzica GC. 1996. Connections of the dorsomedialpart of the nucleus intercollicularis in male non-songbird, the greypartridge: a tract-tracing study. Neurosci Lett 221:61–65.
Brutus M, Shaikh MB, Siegel HE, Siegel A. 1984. An analysis of themechanisms underlying septal area control of hypothalamically elic-ited aggression in the cat. Brain Res 310:235–248.
Casini G, Bingman VP, Bagnoli P. 1986. Connections of the pigeon dorso-medial forebrain studied with WGA-HRP and 3H-proline. J CompNeurol 245:454–470.
Chiba T, Murata Y. 1985. Afferent and efferent connections of the medialpreoptic area in the rat: a WGA-HRP study. Brain Res Bull 14:261–272.
Cohen DH, Goff DM. 1978. Effect of avian basal forebrain lesions, includ-ing septum, on heart rate conditioning. Brain Res Bull 3:111–318.
Cooper RL, Erickson CJ. 1976. Effects of septal lesions on the courtshipbehavior of male ring doves (Streptopelia risoria). Horm Behav 7:441–450.
Dahlstrom A, Fuxe K. 1964. Evidence for the existence of monoamineneurons in the central nervous system. I. Demonstration of mono-amines in cell bodies of brainstem neurons. Acta Physiol Scand Suppl62:1–55.
Davies DC, Csillag A, Szekely AD, Kabai P. 1997 Efferent connections ofthe domestic chick archistriatum: a Phaseolus lectin anterograde trac-ing study. J Comp Neurol 389:679–693.
Durstewitz D, Kroner S, Gunturkun O. 1999. The dopaminergic innerva-tion of the avian telencephalon. Prog Neurobiol 59:161–195.
Erichsen JT, Bingman VP, Krebs JR. 1991. The distribution of neuropep-tides in the dorsomedial telencephalon of the pigeon (Columba livia): abasis for regional subdivisions. J Comp Neurol 314:478–492.
Fahrbach SE, Morrell JI, Pfaff DW. 1986. Identification of medial preopticneurons that concentrate estradiol and project to the midbrain in therat. J Comp Neurol 247:364–382.
Felix B, Roesch T. 1984. Lateral hypothalamic relations with medial hy-pothalamus and limbic areas in geese. Brain Res Bull 13:509–517.
Fernandez-Lopez A, Revilla V, Candelas MA, Gonzalez-Gil J, Diaz A, PazosA. 1997. A comparative study of alpha2- and beta-adrenoceptor distri-bution in pigeon and chick brain. Eur J Neurosci 9:871–883
Ferris CF, Delville Y, Irvin RW, Potegal M. 1994. Septo-hypothalamicorganization of a stereotyped behavior controlled by vasopressin ingolden hamsters. Physiol Behav 55:755–759.
Foster EF, Mehta RP, Bottjer SW. 1997. Axonal connections of the medialmagnocellular nucleus of the anterior neostriatum in zebra finches.J Comp Neurol 382:364–381.
Freund TF, Antal M. 1988. GABA-containing neurons in the septum con-trol of inhibitory interneurons in the hippocampus. Nature 336:170–173.
Fuxe K. 1965. Evidence for the existence of monoamine neurons in thecentral nervous system. IV. Distribution of monoamine nerve terminalsin the central nervous system. Acta Physiol Scand 64:39–85.
Gagliardo A, Ioale P, Bingman VP. 1999. Homing in pigeons: the role of thehippocampal formation in the representation of landmarks used fornavigation. J Neurosci 19:311–315.
Gaykema RP, Luiten PG, Nyakas C, Traber J. 1990. Cortical projectionpatterns of the medial septum-diagonal band complex. J Comp Neurol293:103–124.
Gerfen CR, Sawchenko PE. 1984. An anterograde neuroanatomical tracingmethod that shows the detailed morphology of neurons, their axons andterminals: immunohistochemical localization of an axonally trans-ported plant lectin, Phaseolus vulgaris leucoagglutinin (PHA-L). BrainRes 290:219–238.
Goodson JL. 1998a. Vasotocin and vasoactive intestinal polypeptide mod-ulate aggression in a territorial songbird, the violet-eared waxbill (Es-trildidae: Uraeginthus granatina). Gen Comp Endocrinol 111: 233–244.
Goodson JL. 1998b. Territorial aggression and dawn song are modulatedby septal vasotocin and vasoactive intestinal polypeptide in male fieldsparrows (Spizella pusilla). Horm Behav 34:67–77.
Goodson JL, Adkins-Regan E. 1999. Effect of intraseptal vasotocin andvasoactive intestinal polypeptide infusions on courtship and aggressionin the male zebra finch (Taeniopygia guttata). J Neuroendocr 11:19–25.
Goodson JL, Eibach R, Sakata J, Adkins-Regan E. 1999. Effect of septallesions on male song and aggression in the colonial zebra finch (Tae-
454 C.M. MONTAGNESE ET AL.

niopygia guttata) and the territorial field sparrow (Spizella pusilla).Behav Brain Res 98:167–180.
Gould KL, Newman SW, Tricomi EM, DeVoogdt TJ. 2001. The distributionof substance P and neuropeptide Y in four songbird species: a compar-ison of food-storing and non-storing birds. Brain Res 918:80–95.
Green CJ. 1979. Animal anesthesia. London: Laboratory Animal.Hamill GS, Jacobowitz DM. 1984. A study of afferent projections to the rat
interpeduncular nucleus. Brain Res Bull 13:527–539.Herkenham M, Nauta WJ. 1977. Afferent connections of the habenular
nuclei in the rat. A horseradish peroxidase study, with a note on thefiber-of-passage problem. J Comp Neurol 173:123–146.
Horn G. 1991. Imprinting and recognition memory; a review of neuralmechanisms. In: Andrew RJ, editor. Neural and behavioural plasticity:the use of the domestic chick as a model. Oxford: Oxford UniversityPress. p 219–261.
Hreib KK, Rosene DL, Moss MB. 1988. Basal forebrain efferents to themedial dorsal thalamic nucleus in the rhesus monkey. J Comp Neurol277:365–390.
Huber GC, Crosby EC. 1929. The nuclei and fiber paths of the aviandiencephalon, with consideration of telencephalic and certain mesen-cephalic centers and connections. J Comp Neurol 48:1–225.
Ikemoto S, Panksepp J. 1999. The role of nucleus accumbens dopamine inmotivated behavior: a unifying interpretation with special reference toreward-seeking. Brain Res Rev 31:6–41.
Jakab RL, Leranth C. 1993. Presence of somatostatin or neurotensin inlateral septal dopaminergic axon terminals of distinct hypothalamicand midbrain origins: convergence on the somatospiny neurons. ExpBrain Res 92:420–430.
Johnson F, Bottjer SW. 1992. Growth and regression of thalamic efferentsin the song control system of male zebra finches. J Comp Neurol326:442–450.
Jurkevich A, Barth SW, Aste N, Panzica GC, Grossmann R. 1996. Intra-cerebral sex differences in the vasotocin system in birds: possibleimplication in behavioral and autonomic functions. Horm Behav 30:673–681.
Kahana MJ, Seeling D, Madsen JR. 2001. Theta returns. Curr Opin Neu-robiol 11:739–744.
Kask A, Nguyen HP, Pabst R, Von Horsten S. 2001. Neuropeptide Y Y1receptor-mediated anxiolysis in the dorsocaudal lateral septum: func-tional antagonism of corticotropin-releasing hormone-induced anxiety.Neuroscience 104:799–806.
King TR, Nance DM. 1986. Neuroestrogenic control of feeding behavior andbody weight in rats with kainic lesions of the lateral septal area.Physiol Behav 37:475–481.
Kitt CA, Brauth SE. 1982. A paleostriatal-thalamic-telencephalic path inpigeons. Neuroscience 7:2735–2751.
Kitt CA, Brauth SE. 1986a. Telencephalic projections from midbrain andisthmal cell groups in the pigeon. I. Locus coeruleus and subcoeruleus.J Comp Neurol 247:69–91.
Kitt CA, Brauth SE. 1986b. Telencephalic projections from midbrain andisthmal cell groups in the pigeon. II. The nigral complex. J CompNeurol 247:92–110.
Knapp R, Silver R. 1995. Location of neurons projecting to the hypophysialstalk — median eminence in ring doves (Streptopelia roseogrisea). CellTissue Res 280:77–86
Kohler C, Chan-Palay V, Steinbusch H. 1982. The distribution and originof serotonin-containing fiber in the septal area: a combined immuno-histochemical and fluorescent retrograde tracing study in the rat.J Comp Neurol 209:91–111.
Krayniak PF, Siegel A. 1978. Efferent connections of the septal area in thepigeon. Brain Behav Evol 15:389–404.
Krayniak PF, Weiner S, Siegel A. 1980. An analysis of the efferent con-nections of the septal area in the cat. Brain Res 189:15–29.
Kuenzel WJ, Masson M. 1988. A stereotaxic atlas of the brain of the chick(Gallus domesticus). Baltimore: Johns Hopkins University Press.
Kuenzel WJ, van Tienhoven A. 1982. Nomenclature and location of avianhypothalamic nuclei and associated circumventricular organs. J CompNeurol 206:293–313.
Leranth C, Deller T, Buzsaki G. 1992. Intraseptal connections redefined:lack of a lateral septum to medial septum path. Brain Res 583:1–11.
Leutgeb S, Mizumori SJY. 1999. Excitotoxic septal lesions result in spatialmemory deficits and altered flexibility of hippocampal single-unit rep-resentations. J Neurosci 19:6661–6672.
Liu R, Chang L, Wickern G. 1984. The dorsal tegmental nucleus: anaxoplasmic transport study. Brain Res 310:123–132.
McBride RL, Sutin J. 1976. Projections of the locus coeruleus and adjacentpontine tegmentum in the cat. J Comp Neurol 165:265–284.
Medina L, Reiner A. 1994. Distribution of choline acetyltransferase immu-noreactivity in the pigeon brain. J Comp Neurol 342:497–537.
Medina L, Reiner A. 1997. The efferent projections of the dorsal andventral pallidal parts of the pigeon basal ganglia, studied with biotin-ylated dextran amine. Neuroscience 81:773–802.
Meibach RC, Siegel A. 1977a. Efferent connections of the septal area in therat: an analysis utilizing retrograde and anterograde transport meth-ods. Brain Res 119:1–20.
Meibach RC, Siegel A. 1977b. Efferent connections of the hippocampalformation in the rat. Brain Res 124:197–224
Mezey S, Csillag A. 2002. Selective striatal connections of midbrain dopa-minergic nuclei in the chick (Gallus domesticus). Cell Tissue Res 308:35–46.
Montagnese CM, Mezey SE, Csillag A. 2003. Efferent connections of thedorsomedial thalamic nuclei of the domestic chick (Gallus domesticus).J Comp Neurol 459:301–326.
Numan M, Smith HG. 1984. Maternal behavior in rats: evidence for theinvolvement of preoptic projections to the ventral tegmental area.Behav Neurosci 98:712–727.
Ottersen OP. 1980. Afferent connections to the amygdaloid complex of therat and cat. II. Afferents from the hypothalamus and the basal telen-cephalon. J Comp Neurol 194:267–289.
Panzica GC, Aste N, Castagna C, Viglietti-Panzica C, Balthazart J. 2001.Steroid-induced plasticity in the sexually dimorphic vasotocinergic in-nervation of the avian brain: behavioral implications. Brain Res Rev37:178–200.
Parent A, Gravel S, Boucher R. 1981. The origin of forebrain afferents tothe habenula in rat, cat and monkey. Brain Res Bull 6:23–38.
Parent A, Pare D, Smith Y, Steriade M. 1988. Basal forebrain cholinergicand noncholinergic projections to the thalamus and brainstem in catsand monkeys. J Comp Neurol 277:281–301.
Puelles L, Kuwana E, Puelles E, Rubenstein JL. 1999. Comparison of themammalian and avian telencephalon from the perspective of geneexpression data. Eur J Morphol 37:139–150.
Richardson RD, Boswell T, Raffety BD, Seeley RJ, Wingfield JC, WoodsSC. 1995. NPY increases food intake in white-crowned sparrows: effectin short and long photoperiods. Am J Physiol Regul Integr CompPhysiol 268: R1448–R1422.
Risold PY, Swanson LW. 1997. Connections of the rat lateral septal com-plex. Brain Res Rev 24:115–195.
Robinzon B, Snapir N, Perek M. 1978. Hyperphagia without obesity inseptal-lesioned cocks. Physiol Behav 20:1–6.
Rose SP. 1991. How chicks make memories: the cellular cascade from c-fosto dendritic remodelling. Trends Neurosci 14:390–397.
Russchen FT. 1982. Amygdalopetal projections in the cat. II. Subcorticalafferent connections. A study with retrograde tracing techniques.J Comp Neurol 207:157–176.
Saldanha CJ, Leak RK, Silver R. 1994. Detection and transduction ofdaylength in birds. Psychoneuroendocrinology 19:641–656.
Shibata H. 1989. Descending projections to the mammillary nuclei in therat, as studied by retrograde and anterograde transport of the wheatgerm agglutinin-horseradish peroxidase. J Comp Neurol 285:436–452.
Shiflett MW, Gould KL, Smulders TV, DeVoogd TJ. 2002. Septum andfood-storing behavior are related in parids. J Neurobiol 51:215–222.
Shimizu T, Cox K, Karten HJ, Britto LRG. 1994. Cholera toxin mapping ofretinal projections in pigeons (Columba livia), with emphasis on reti-nohypothalamic connections. Vis Neurosci 11:441–446.
Siegel JJ, Nitz D, Bingman VP. 2000. Hippocampal theta rhythm inawake, freely moving homing pigeons. Hippocampus 10:627–631.
Silver R, Witkovsky P, Horvath P, Alones V, Barnstable CJ, Lehman MN.1988. Coexpression of opsin- and VIP-like-immunoreactivity in CSF-contacting neurons of the avian brain. Cell Tissue Res 253:189–198.
Smeets WJAJ, Gonzalez A. 2000. Catecholamine systems in the brain ofvertebrates: new perspectives through a comparative approach. BrainRes Rev 33:308–379.
Smith Y. 1992. Anterograde tracing with PHA-L and biocytin at the elec-tron microscopic level. In: Bolam JP, editor. Experimental neuroanat-omy. A practical approach. Rickwood D, Hames BD, series editors. Thepractical approach series. Oxford: IRL Press. p 61–80.
455SEPTAL EFFERENTS OF DOMESTIC CHICK

Smith DA, Flynn JP. 1980. Afferent projections to affective attack sites incat hypothalamus. Brain Res 194:41–51.
Spatz WB. 1989. Differences in transneuronal transport of horseradishperoxidase conjugated wheat germ agglutinin in the visual system:marmoset monkey and guinea pig compared. J Hirnforsch 30:375–384.
Suthers RA, Margoliash D. 2002. Motor control of birdsong. Curr OpinNeurobiol 12:684–690.
Swanson LW, Mogenson CG, Simerly RB, Wu M. 1987. Anatomical andelectrophysiological evidence for a projection from the medial preopticarea to the “mesencephalic and subthalamic locomotor regions” in therat. Brain Res 405:108–122.
Szeidelmann Z, Shanabrough M, Leranth C. 1995a. Hypothalamic Leu-enkephalin-immunoreactive fibers terminate on calbindin-containingsomatospiny cells in the lateral septal area of the rat. J Comp Neurol358:573–583.
Szeidelmann Z, Jakab RL, Shanabrough M, Leranth C. 1995b. Extrinsicand intrinsic substance P innervation of the rat lateral septal areacalbindin cells. Neuroscience 69:1205–1221.
Szekely AD. 1999. The avian hippocampal formation: subdivisions andconnectivity. Behav Brain Res 98:219–225.
Szekely AD, Krebs JR. 1996. Efferent connectivity of the hippocampalformation of the zebra finch (Taenopygia guttata): an anterogradepathway tracing study using Phaseolus vulgaris leucoagglutinin.J Comp Neurol 368:198–214.
Thompson RH, Swanson LW. 1998. Organization of inputs to the dorso-medial nucleus of the hypothalamus: a reexamination with fluorogoldand PHAL in the rat. Brain Res Rev 27:89–118.
Toth K, Freund TF. 1992. Calbindin D28k-containing nonpyramidal cellsin the rat hippocampus: their immunoreactivity for GABA and projec-tion to the medial septum. Neuroscience 49:793–805.
Toth K, Borhegyi Z, Freund TF. 1993. Postsynaptic targets of GABAergichippocampal neurons in the medial septum-diagonal band of brocacomplex. J Neurosci 13:3712–3724.
Varoqueaux F, Poulain P. 1994. Lateral septal projections onto tubero-infundibular neurons in the hypothalamus of the guinea pig. CellTissue Res 278:217–225.
Veenman CL, Karle EJ, Anderson KD, Reiner A. 1995a. Thalamostriatalprojection neurons in birds utilize LANT6 and neurotensin: a light andelectron microscopic double-labeling study. J Chem Neuroanat 9:1–16.
Veenman CL, Wild JM, Reiner A. 1995b. Organization of the avian “cor-ticostriatal” projection system: a retrograde and anterograde pathwaytracing study in pigeons. J Comp Neurol 354:87–126.
Veenman CL, Medina L, Reiner A. 1997. Avian homologues of mammalian
intralaminar, mediodorsal and midline thalamic nuclei: immunohisto-chemical and hodological evidence. Brain Behav Evol 49:78–98.
Vertes RP. 1991. A PHA-L analysis of ascending projections of the dorsalraphe nucleus in the rat. J Comp Neurol 313:643–668.
Vertes RP. 1992. PHA-L analysis of projections from the supramammillarynucleus in the rat. J Comp Neurol 326:595–622.
Vertes RP, Fass B. 1988. Projections between the interpeduncular nucleusand basal forebrain in the rat as demonstrated by the anterograde andretrograde transport of WGA-HRP. Exp Brain Res 73:23–31.
Vertes RP, McKenna JT. 2000. Collateral projections from the supramam-millary nucleus to the medial septum and hippocampus. Synapse 38:281–293.
Wallman J, Saldanha CJ, Silver R. 1994. A putative suprachiasmaticnucleus of birds responds to visual motion. J Comp Physiol (A) 174:297–304.
Watson JT, Adkins-Regan E, Whiting P, Lindstrom JM, Podleski TR. 1988.Autoradiographic localization of nicotinic acetylcholine receptors in thebrain of zebra finch (Poephila guttata). J Comp Neurol 274:255–264.
Wayner MJ, Barone FC, Scharoun SL, Guevara-Aguilar R, Aguilar-Baturoni HU. 1983. Limbic connections to the lateral preoptic area: ahorseradish peroxidase study in the rat. Neurosci Biobehav Rev 7:375–384.
Wild JM. 1987. Thalamic projections to the paleostriatum and neostriatumin the pigeon (Columba livia). Neuroscience 20:305–327.
Woolf NJ, Eckenstein F, Butcher LL. 1984. Cholinergic systems in the ratbrain. I. projections to the limbic telencephalon. Brain Res Bull 13:751–784.
Wouterlood FG, Gaykema RP, Steinbusch HW, Watanabe T, Wada H.1988. The connections between septum-diagonal band complex andhistaminergic neurons in the posterior hypothalamus of the rat. An-terograde tracing with Phaseolus vulgaris-leucoagglutinin combinedwith immunocytochemistry of histidine decarboxylase. Neuroscience26:827–845.
Wu M, Shanabrough M, Leranth Cs, Alreja M. 2000. Cholinergic excitationof septohippocampal GABA but not cholinergic neurons: implicationsfor learning and memory. J Neurosci 20:3900–3908.
Wynne B, Gunturkun O. 1995. Dopaminergic innervation of the telenceph-alon of the pigeon (Columba livia): a study with antibodies againsttyrosine hydroxylase and dopamine. J Comp Neurol 357:446–464.
Yamauchi K, Yasuda M. 1981. Several cell masses around the commissuraanterior in the chicken. J Hirnforsch. 22:189–194.
Zeier H, Karten HJ. 1971. The archistriatum of the pigeon: organization ofafferent and efferent connections. Brain Res 31:313–326.
456 C.M. MONTAGNESE ET AL.





























