Embed Size (px)
Citation preview

ARTICLEdoi:10.1016/j.ymthe.2005.05.016
Efficient Hepatic Delivery and Expression from aRecombinant Adeno-Associated Virus 8
Pseudotyped A1-Antitrypsin Vector
Thomas J. Conlon,1 Travis Cossette,1 Kirsten Erger,1 Young-Kook Choi,2 Tracy Clarke,3
Marda Scott-Jorgensen,4 Sihong Song,2 Martha Campbell-Thompson,3
James Crawford,3 and Terence R. Flotte1,*
1Department of Pediatrics and Powell Gene Therapy Center, University of Florida, Gainesville, FL 32608, USA2Department of Pharmaceutics; 3Department of Pathology,
Center for Immunology and Transplantation, and 4University of Florida Shands Cancer Center,
Stem Cell Biology Program, University of Florida, Gainesville, FL 32610, USA
*To whom correspondence and reprint requests should be addressed. E-mail: [email protected].
Available online 8 August 2005
MOLECULA
Copyright C
1525-0016/$
A1-Antitrypsin (AAT) deficiency is a single-gene disorder in which a mutation in the AAT (approvedsymbol SERPINA1) gene (PI*Z) leads to misfolding of the protein, loss of the protective antiproteaseeffect of AAT for the lungs, and a toxic effect on hepatocytes. Optimal therapy for AAT deficiencywill require a high percentage of hepatocyte transduction to be effective for liver and lung disease.Recently, rAAV genomes pseudotyped with capsids from serotypes 7 and 8 showed efficient hepatictransduction. We hypothesized that upon portal vein injection to target hepatocytes, serotype 8would better transduce target cells and therefore express hAAT in both a greater percentage of cellsand greater amounts. AAV2 and pseudotyped vectors for serotypes 1, 5, and 8 carrying the humanAAT transgene were injected at 1 � 1010 particle doses into C57Bl/6 mice. Circulating hAAT fromAAV2/8-injected animals showed a 2-log advantage over AAV2 and 3-log increase over AAV2/1 and5 for the 24-week study. Most significantly, up to 40% of total liver cells stained positive for thetransgene in AAV2/8 subjects while remaining primarily episomal. Therefore, pseudotyped AAV8provides a vehicle to infect a high percentage of hepatocytes stably and thereby express therapeuticmolecules to modify AAT PiZ transcripts.
R
Th
30
Key Words: adeno-associated virus, gene therapy, A1-antitrypsin deficiency, hepatocytes, viralvectors, liver, serotype, episome, hepatectomy
INTRODUCTION
a1-Antitrypsin (AAT) deficiency is a well-described single-gene defect for which muscle-directed recombinantadeno-associated virus (rAAV)-based gene therapy isalready in clinical trials [1]. One particular AAT mutation,PiZ homozygous deficiency, affects 1 in 6000–8000 birthsin the United States [2]. AAT is a 52-kDa protease inhibitorpredominantly expressed in the liver, from where it issecreted to act upon its major target organ, the lung.Within the lung, AAT helps prevent damage to theextracellular matrix and to cells by balancing the protea-se:antiprotease ratio. The primary substrate is neutrophilelastase (NE), which AAT binds to and inactivates. AAThas additional activity against cathepsin B and otherproteases. When serum concentrations of AAT fall below11 AM (b570 Ag/ml) [3,4], the protective effect is lost andthe uncontrolled activity of NE proceeds to destroy the
THERAPY Vol. 12, No. 5, November 2005
e American Society of Gene Therapy
.00
elastin fibers supporting the alveolar network, leading toloss of airway tethering and destruction of alveolar septae.Current clinical treatments involve the infusion ofpurified protein or plasma-derived blood products [5–7].These lengthy treatments can be performed weekly with asignificant protective effect and few side effects, but withconsiderable cost and discomfort to the patient. A subsetof patients with AAT deficiency also experiences liverdisease, which is due to accumulation of the PiZ mutantdue to impaired secretion within hepatocytes.
For clinical replacement, gene therapy for lung diseasein AAT deficiency can be viewed as a long-lastingalternative to protein replacement therapy, with the goalbeing maintenance of serum levels at or above the 11 AMthreshold. Delivery of the AAT gene (approved symbolSERPINA1) to any tissue capable of expressing andsecreting this protein would be expected to accomplish
867

ARTICLE doi:10.1016/j.ymthe.2005.05.016
this goal. Indeed, AAT expression and secretion havebeen demonstrated from a number of ectopic sites,including muscle [8,9], bronchial epithelium [10], andskin [11]. In contrast, any gene therapy with potential totarget both liver and lung disease must involve deliveryof therapeutic molecules to the hepatocytes themselves.Down-regulation of the mutant PiZ-AAT with ribozymesin vitro has been demonstrated by Zern et al. using anSV40-based vector [12], but any such approach willrequire a vector capable of transduction of a highpercentage of hepatocytes. Standard rAAV2 vectors, suchas those used in current clinical trials, appear to beincapable of stably transducing more than 5% of thehepatocyte population. Recombinant AAV2/8 pseudo-typed (rAAV8) vectors have recently been shown to bemore efficient for hepatocyte transduction than rAAV2vectors [13]. In the current study, we evaluated a rAAV2/8-AAT vector to assess its potential for stable and efficientdelivery of human AAT in the C57Bl/6 mouse. We alsoevaluated the percentage of hepatocytes transduced andthe molecular fate of rAAV2/8-delivered genomes.
RESULTS
In Vivo Liver Transduction Comparison of AAVSerotypesTo continue to enhance transduction of liver hepatocytesfor increased transgene expression, we evaluated the
FIG. 1. Transduction of hepatocytes by AAV serotype 8 produces high levels of a
capsid sequences for AAV serotypes 1, 2, 5, and 8 were cotransfected with pCBAT
titered and injected into the portal vein of C57Bl/6 mice to target the liver with eq
8, 12, 16, 20, and 24 weeks postinjection, serum samples were analyzed for tran
pattern of heightened expression by 1 week can be detected for each serotype a
weeks after being injected through the portal vein with 1 � 1011 particles, animal
DNA was extracted. Purified DNA was assayed by quantitative RT-PCR for AAV c
genomes per microgram of genomic DNA standard. The mean F standard devi
868
newly discovered AAV8 serotype with three rAAV sero-types used in our lab and by others. We packaged thehuman a1-antitrypsin complementary DNA sequencedriven by the highly active cytomegalovirus (CMV)enhancer/chicken h-actin promoter (CB) plasmid intovirions by cotransfection with plasmids containing thecapsid sequences for AAV serotypes 1, 2, 5, and 8. All ITRswere from serotype 2 to control for possible differences ingenome manipulations once uncoated. We introduced allfour vectors into C57Bl/6 mice via direct injection intothe portal vein with 9.6 � 1010 particles per animal. Weevaluated infection efficiency by analyzing serum levelsof the AAT transgene by ELISA for up to 24 weeks. Peaklevels for each vector were obtained within 1 to 3 weekspostinjection and remained stable throughout the courseof the study (Fig. 1A). We saw a greater than 100-foldenhancement of hAAT by AAV2/8 over AAV2/2 and a1000-fold advantage over AAV2/1 and 5.
To confirm AAV2/8 further as a more efficient liver-mediated gene therapy vector, we excised liver tissue at24 weeks and subjected genomic DNA to real-time PCR.Primer probes specific for the CB promoter were used in areaction with 1 Ag of genomic DNA. Copy number percell correlated with hAAT transgene expression in termsof AAV2/8 outperforming vectors based on AAV serotypes1, 2, and 5 (Fig. 1B). In two cases, though, animalsinjected with the AAV2/8 vector had genome counts percell that were lower than the range for those receiving
1-antitrypsin and more genome copies. (A) Packaging plasmids encoding the
into 293 cells to generate complete virions. The array of viruses generated wa
ual doses of 9.6 � 1010 vector particles per animal. At time points of 0, 1, 3, 5
sgene (AAT) expression, and means at each time point are graphed. A simila
nd stable transgene levels are detected throughout the study. (B) Twenty-fou
s underwent a partial hepatectomy. Liver biopsies were collected and genomic
opies using the pCBAT plasmid as the standard and using the 166,000 mouse
ation of copies per cell for each animal is plotted for comparison.
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therap
s
,
r
r
y

ARTICLEdoi:10.1016/j.ymthe.2005.05.016
AAV2, but were still secreting hAAT levels highenough to contribute to the groupTs log increaseaverage. The AAT levels from these two subjects mayresult from transduction and secretion from pancreaticcells, where positive staining was observed (data notshown). Also, a maximum concentration of circulatinghAAT in the murine serum may have been reached.Therefore, the full potential of using AAV2/8 in theliver may not be seen. Overall, the higher AAT proteinlevels reflect the number of vector genomes per cellwithin the liver.
Histology and Immunohistochemistry for Humana1-antitrypsin in the Murine LiverAdministration of AAV vectors did not result in morpho-logical changes as assessed on H&E-stained paraffinsections for all mice. Occasional focal to multifocalmononuclear infiltrates were observed in PBS- and vec-tor-treated animals and were considered normal foranimals from this colony.
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therapy
We subsequently performed immunostaining ofmouse liver sections for the human AAT protein todetermine the proportion of hepatocytes infected. Weconfirmed specificity of the primary antibody for humanAAT using normal human liver compared to normalmouse liver (data not shown). Additionally, visualizationof the tissue allows for the determination of cell typestransduced and intensity of AAT expression. As shown inFig. 2A, we observed no staining for human AAT in a PBS-injected animal. In comparison, the AAV2 (Fig. 2C) andAAV2/1, 5, and 8 (Figs. 2B, 2D, and 2E, respectively)pseudotype-treated animals consistently show positivestaining. Semiquantitative evaluation of human AATimmunoreactivity revealed a positive signal for eachserotype for nearly all criteria in contrast to no stainingin the PBS-injected controls. Observed trends includedscores of b2Q and above for only type 2- and 8-transducedtissue in the categories of panlobular blush, which canresult from the AAT secreted being endocytosed byneighboring cells and midzonal and periportal staining.
FIG. 2. Immunostaining for hAAT in the murine liver.
All sections were incubated with a rabbit anti-human
AAT antibody as the primary antibody. (A) Liver
section of a PBS-injected mouse to serve as a
negative control reference showing no positive
staining for hAAT. (B) AAV2/1-CBAT, (C) AAV2-
CBAT, (D) AAV5-CBAT, and (E) AAV8-CBAT liver
sections from mice administered with 1 � 1013
vector particles via the portal vein. Staining of livers
from each serotype reveals an overall panlobular
blush, midzonal, periportal, and perivenous detec-
tion of hAAT. Central veins (CV) and portal veins (PV)
are noted. (F) 40� magnification of outlined portion
in (E) demonstrating staining around the central
vein. Tissues were harvested at 24 weeks post-
injection during a partial hepatectomy procedure.
Original magnifications: A–E, 20�; F, 40�.
869

ARTICLE doi:10.1016/j.ymthe.2005.05.016
Only AAV2/8-injected liver tissue resulted in scores of b3Qor above. Decile scores, an indicator of the overallpercentage of AAT-positive cells with intensity over theblush level, were at least in the 0–10% range for everymouse scored in all pseudotype groups. Most impres-sively, only the AAV8 tissue reached decile scores up to40%. Of note, additional positive cell types and patternsseen in this group include bile duct staining in seven ofeight animals reviewed and intense nuclear and granular,diffuse cytoplasmic staining in AAV8-treated animals.The trends observed herein parallel those seen in genomecopy number and circulating AAT levels with AAV2immunoreactivity exceeding the AAV2/1 and 2/5 groupsand AAV2/8 outperforming all groups in cellular inten-sity and positive spread over the tissue.
Persistence of AAV2/8 Genomes in the Liver Is Similarto That of AAV2 GenomesTo determine the preferred molecular state of AAV2/8 intransduced liver cells, we performed a partial hepatec-tomy at 24 weeks in the previous group of animals. Weobserved the effects on transgene expression and molec-ular persistence after the partial hepatectomy by trans-gene expression, copies per liver cell, and Southern blotto determine molecular state. As shown in Fig. 3A, hAATlevels expressed from the various AAV vectors wereplotted in relationship to pre-partial hepatectomy levels.
FIG. 3. Decreased levels of hAAT and AAV genome copies after partial hepatect
animals in Fig. 2 underwent a partial hepatectomy that could remove up to 85%
until week 8, when the animals were sacrificed. AAT transgene means for each ser
obtained at week 24. A minor decrease at week 1 reflects the 2-week half-life of A
propensity toward integration compared to other serotypes. (B) This graph rep
surgery removing three lobes of the liver. Each bar representing the mean and sta
obtained the same way before surgery 8 weeks prior. Again, a trend toward copy r
copies for AAV8.
870
As expected from previous studies [17], AAV2 animalsrecorded a drop to 10 to 40% of pretreatment levels aftera washout period of circulating AAT (compare week 1 toweek 3). Likewise, cohorts injected with AAV2/1 and 2/8saw an even greater decrease in hAAT to below 10% ofweek 24 values. Unexpectedly, AAV2/5 showed a decid-edly different pattern with hAAT levels rising as liverregeneration occurred. Also, it is important to note thatalthough administration was directed to the liverthrough an intravascular injection, one cannot rule outthe possibility that the total AAT level determined inserum sampling is not in part due to expression fromadditionally infected organs and may be reflected in post-partial hepatectomy AAT level differences between pseu-dotype groups (Fig. 3A). For instance, AAV5 has beenshown to have a greater affinity for pancreatic cells [21]and the lung epithelial cells [22]. Although AAV2/5 isshown, this study specifically addressed serotype 8 inhepatocytes. The reduction in serum AAT data wassupported by real-time PCR of liver genomic DNA uponsacrifice at 8 weeks post-partial hepatectomy. A directcorrelation (r2 = 0.9809) between the percentage decreasein hAAT (Fig. 3A) expression levels and percentagereduction in copy number is observed (Fig. 3B) beforeand after surgery. Similar to Fig. 3A, pseudotypes 1, 2, and8 all exhibited a considerable loss of vector copies per cellafter partial hepatectomy.
omy for serotypes 1, 2, and 8. (A) At 24 weeks after portal vein injection, the
of the total liver weight. Starting 1 week after surgery serum sampling resumed
otype cohort determined by ELISA are plotted as a percentage of the AAT level
AT in circulation. The increase in levels for the serotype 5 group may reflect a
resents the AAV copy number as detected by real-time PCR before and afte
ndard deviation of a serotype cohort is a percentage of the total copy numbe
egeneration within the proliferating liver is seen as well as the expected drop in
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therap
s
r
r
y

IG. 4. Southern blot for genomic persistence of AAV serotypes in the liver.
otal cellular genomic DNA was extracted from frozen liver tissue. Forty
icrograms of genomic DNA was digested with either HindIII (one-cutter
ithin the vector) or ApaI (two-cutter within the vector) overnight and
parated on agarose gel. (A) Schematic of the restriction sites and expected
and sizes and annealing location of the hAAT probe. (B) After blotting to
itrocellulose, a 32P-labeled probe to the hAAT sequence was hybridized.
ndogenous bands not vector related, head-to-tail (H-T) concatemers, and the
ouble-digested internal fragments are noted.
ARTICLEdoi:10.1016/j.ymthe.2005.05.016
Both sets of data suggest AAV2/8 and AAV2/1 persist in anonintegrated state similar to that of AAV2. One caninfer episomal persistence because, with integration ofthe vector genome, liver regeneration would cause addi-tional copies to be replicated during cell division.Unexpectedly, in both sets of experiments, pseudotypedAAV5-injected animals produced very different resultsfrom those anticipated. By 8 weeks after partial hepatec-tomy, AAT levels in these mice had returned to presur-gical levels. In addition, quantification of total AAVgenome copies before and after surgery resulted in nearlyequivalent copy numbers at both time points. At least90% of the copies present before removal of the liver werethere at sacrifice, suggesting that some greater degree ofvector genomes were integrated and replicated alongsideliver regeneration. Support for this conclusion is theobservation that the AAV2/5 group had an initial drop inAAT levels at week 1 after partial hepatectomy, thusindicating that productively transduced cells wereremoved along with the AAV genomes driving AATexpression. The wild-type AAV2/5 genome tends to differfrom other known serotypes on several levels. Thesedifferences include an efficient transcription initiationstart site within the ITR; all Rep-coding transcriptspolyadenylate within the intron and adenovirus co-infection does not enhance Cap protein expression [23].Although significant, these differences and others notmentioned cannot explain the results presented in thisstudy since all vectors were pseudotyped with AAV2 ITRsand genome sequences were identical. Therefore, thecapsid proteins were the only difference and could resultin a cellular processing that is different from that of theother three. Furthermore, the use of an endosomalpathway could potentially bmarkQ the DNA that leads togreater integration efficiency once in the cell. We canalso try to explain this by the novelty of the partialhepatectomy procedure.
Molecular Fate of the Various AAV Serotypes bySouthern BlotAnother method of comparing the molecular fate of AAVserotypes whether episomal or integrated is by analysis ofgenomic DNA. To perform this, we purified total livergenomic DNA from the above-described experiment anddigested the resulting DNA with an enzyme that eithercuts once within the CBAT vector or cuts twice (Fig. 4A).We separated the digested DNA by agarose gel andblotted it to a membrane. A hAAT 32P-labeled probeallowed for the identification and size determination ofthe restricted DNA (Fig. 4B). As previously observed andreported [24], AAV2 primarily exists long term in anextrachromosomal form, linear or circular, evidenced bythe one-cutter digest bands running at a lower molecularweight than if the majority of the vector were integrated,for which a higher molecular weight band would appear(lane 11). In addition, it can be concluded that alter-
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therapy
F
T
m
w
se
b
n
E
d
native serotypes 1, 5, and 8 persist in the same manner(lanes 2, 5, 7, 9, and 13). Furthermore, these data supportthe correlation of serum hAAT levels with cellularinfection. After digestion with an enzyme that cuts twicewithin the vector, all AAV genome copies, whetherintegrated or episomal, will result in a 2.28-kb bandrepresenting the total cellular copies (lanes 3, 4, 6, 8, 10,and 12). An overabundance of genomes present in AAV2/8-treated animals is clearly shown in signal strengthcompared to AAV1, 2, and 5 pseudotyped vectors.
Dose Response of AAV2/8 in VivoWhile developing a viral vector for a secreted proteindisorder such as AAT deficiency, the minimum effectivedose may be sought for preventing toxic levels of thetransgene and reducing production costs passed down tothe patient. Therefore, we next conducted a doseescalation study with AAV8 to determine the scalabilityof this vector. We injected cohorts of C57Bl/6 mice withincreasing doses of our AAV2/8/CBAT vector (1.65 � 109,1.65 � 1010, and 1.65 � 1011 vector genomes/mouse) intothe portal vein and measured the reporter proteinexpression by ELISA (Fig. 5). A 15-fold increase in trans-gene expression could be seen for each 10-fold increase ofvector administered. This measure was maintained forthe 24-week study. At the highest dose, maximum
871
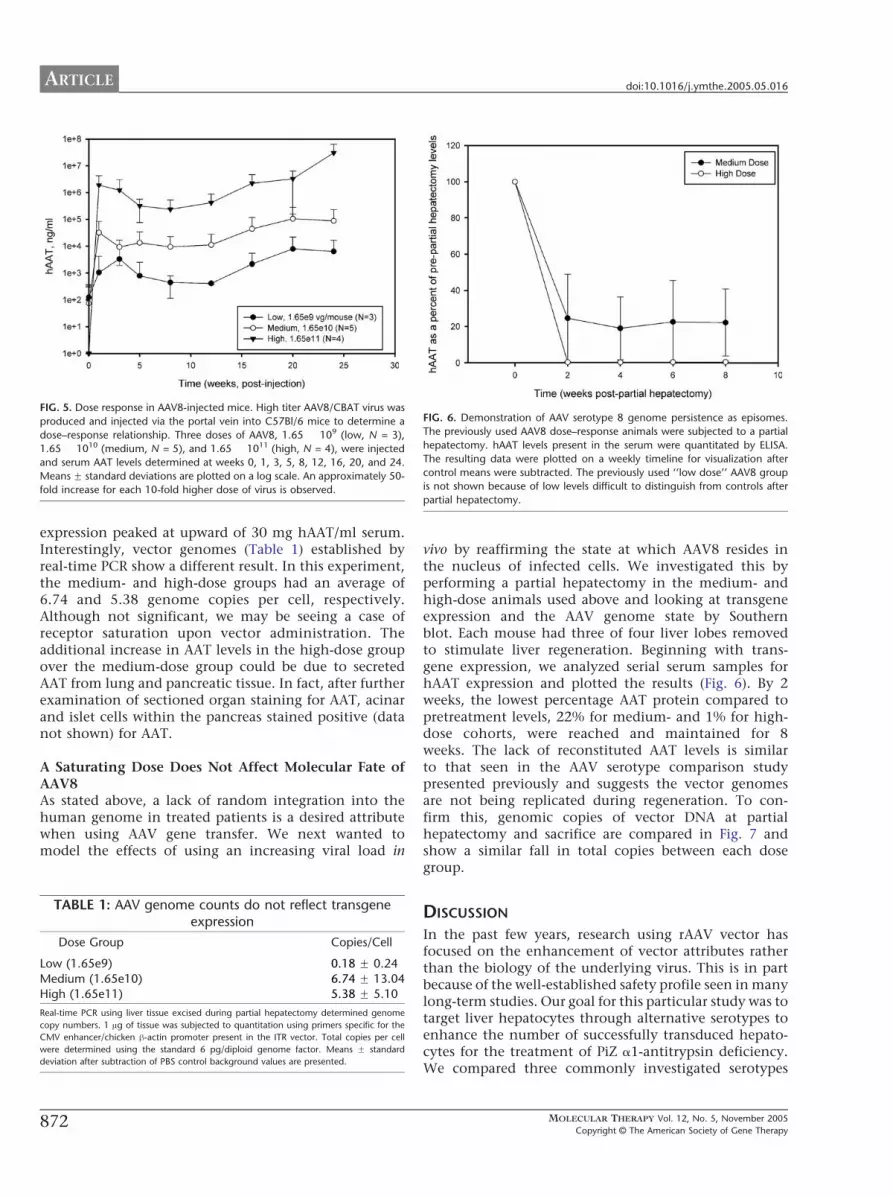
FIG. 5. Dose response in AAV8-injected mice. High titer AAV8/CBAT virus wa
produced and injected via the portal vein into C57Bl/6 mice to determine a
dose–response relationship. Three doses of AAV8, 1.65 � 109 (low, N = 3)
1.65 � 1010 (medium, N = 5), and 1.65 � 1011 (high, N = 4), were injected
and serum AAT levels determined at weeks 0, 1, 3, 5, 8, 12, 16, 20, and 24
Means F standard deviations are plotted on a log scale. An approximately 50
fold increase for each 10-fold higher dose of virus is observed.
TABLE 1: AAV genome counts do not reflect transgeneexpression
Dose Group Copies/Cell
Low (1.65e9) 0.18 F 0.24
Medium (1.65e10) 6.74 F 13.04High (1.65e11) 5.38 F 5.10
Real-time PCR using liver tissue excised during partial hepatectomy determined genom
copy numbers. 1 Ag of tissue was subjected to quantitation using primers specific for th
CMV enhancer/chicken h-actin promoter present in the ITR vector. Total copies per ce
were determined using the standard 6 pg/diploid genome factor. Means F standard
deviation after subtraction of PBS control background values are presented.
FIG. 6. Demonstration of AAV serotype 8 genome persistence as episomes
The previously used AAV8 dose–response animals were subjected to a partia
hepatectomy. hAAT levels present in the serum were quantitated by ELISA
The resulting data were plotted on a weekly timeline for visualization afte
control means were subtracted. The previously used ‘‘low dose’’ AAV8 group
is not shown because of low levels difficult to distinguish from controls afte
partial hepatectomy.
ARTICLE doi:10.1016/j.ymthe.2005.05.016
872
s
,
.
-
expression peaked at upward of 30 mg hAAT/ml serum.Interestingly, vector genomes (Table 1) established byreal-time PCR show a different result. In this experiment,the medium- and high-dose groups had an average of6.74 and 5.38 genome copies per cell, respectively.Although not significant, we may be seeing a case ofreceptor saturation upon vector administration. Theadditional increase in AAT levels in the high-dose groupover the medium-dose group could be due to secretedAAT from lung and pancreatic tissue. In fact, after furtherexamination of sectioned organ staining for AAT, acinarand islet cells within the pancreas stained positive (datanot shown) for AAT.
A Saturating Dose Does Not Affect Molecular Fate ofAAV8As stated above, a lack of random integration into thehuman genome in treated patients is a desired attributewhen using AAV gene transfer. We next wanted tomodel the effects of using an increasing viral load in
e
e
ll
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therap
.
l
.
r
r
vivo by reaffirming the state at which AAV8 resides inthe nucleus of infected cells. We investigated this byperforming a partial hepatectomy in the medium- andhigh-dose animals used above and looking at transgeneexpression and the AAV genome state by Southernblot. Each mouse had three of four liver lobes removedto stimulate liver regeneration. Beginning with trans-gene expression, we analyzed serial serum samples forhAAT expression and plotted the results (Fig. 6). By 2weeks, the lowest percentage AAT protein compared topretreatment levels, 22% for medium- and 1% for high-dose cohorts, were reached and maintained for 8weeks. The lack of reconstituted AAT levels is similarto that seen in the AAV serotype comparison studypresented previously and suggests the vector genomesare not being replicated during regeneration. To con-firm this, genomic copies of vector DNA at partialhepatectomy and sacrifice are compared in Fig. 7 andshow a similar fall in total copies between each dosegroup.
DISCUSSION
In the past few years, research using rAAV vector hasfocused on the enhancement of vector attributes ratherthan the biology of the underlying virus. This is in partbecause of the well-established safety profile seen in manylong-term studies. Our goal for this particular study was totarget liver hepatocytes through alternative serotypes toenhance the number of successfully transduced hepato-cytes for the treatment of PiZ a1-antitrypsin deficiency.We compared three commonly investigated serotypes
y

FIG. 7. Total AAV copy number decreases after partial hepatectomy in mice
administered three doses of AAV2/8. This graph represents the AAV copy
number as detected by real-time PCR before and after surgery removing three
lobes of the liver. Each bar represents the mean and standard deviation of a
serotype cohort as a percentage of the total copy number obtained the same
way before surgery 8 weeks prior. A comparable reduction in total copy is seen
with each dose group.
ARTICLEdoi:10.1016/j.ymthe.2005.05.016
with the newly discovered AAV8 serotype isolated fromrhesus monkeys [13]. After portal vein introduction intoC57Bl/6 mice, serum AAT levels in the AAV8 vector-treated animals were significantly higher than for allother serotypes tested, up to 100-fold greater than for theAAV2 vectors. Similar results have been documented byother investigators [13]. The performance of AAV8 wasalso evaluated on the molecular level with real-time PCRfor vector genomes and Southern blotting. Based on thefinding of 166,000 double-stranded DNA genomes permicrogram of isolated genomic DNA, an average of 4.3genomes were present per cell in AAV8-infected livers, incontrast to 0.54 AAV genomes per cell for AAV2 tissue.Both sets of data confirm AAV8 as a better alternative topreviously used serotypes for human AAT secretion fromthe liver. Finally, immunohistochemistry for AAT in liversections of the injected mice showed up to 40% of thecell population was intensely positive for AAV8-treatedanimals. The difference may reflect the rate of uncoatingof the capsids once internalized [25]. In addition, theAAV8 capsid proteins may target internalized virions toan alternative endosomal pathway that leads to anincrease in stable genomes. Of note, in this studyAAV2/5 pseudotyped vectors behaved differently from1, 2, and 8 pseudotyped vectors. After partial hepatec-tomy, regeneration of vector genomes was observed bycopy number and transgene levels as the liver massincreased in size over time. Unfortunately, these resultscould not be confirmed by Southern blot analysis due tothe low intensity of high-molecular-weight bands in
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therapy
which integrated genomes would appear and would berepresented along with endogenous sequences bound tothe probe. Therefore, if AAV2/5 did have a greaternumber of integrants the degree could not be deter-mined. These confounding AAV5 observations havebeen made for the first time and require further inves-tigation. Using AAV2/8 pseudotyped vectors in conjunc-tion with the equal or greater expression potential of theliver-specific albumin promoter [26] may provide for theoptimal combination for the down-regulation of AAT-PiZ. Through these data, we have made considerableprogress toward developing a rAAV viral vector therapycapable of infecting a large enough population of cellswithin the liver, with the goal of down-regulatingmutant transcripts in human AAT-deficiency patientsby expressing therapeutic molecules.
Of importance when selecting a virus or serotype touse in gene therapy applications is the potential forintegration and disruption of tumor suppressor and genesessential to the normal functioning and maintenance ofthe human body. In one retroviral gene therapy trial inparticular, 2 of 10 treated patients acquired a leukemiacondition resulting from the vector integration withinthe LMO2 oncogene and activation (reviewed in [27,28]).This result was confirmed through sequencing of vector–chromosome junctions and the identification of clonalexpansion of the disrupted genome. Ongoing clinicaltrials using AAV2-based vectors have shown them to besafe for up to 4 years [29]. Chances of vector integrationmay be modeled in mouse studies with the use ofSouthern blotting. Our data presented here do not showthe presence of migrating high-molecular-weightgenomic DNA when total genomic DNA is digested withan enzyme that cuts once within the vector, indicatingvery low chances of integration. We cannot rule out thepossibility of a very limited number of integratedgenomes (b1 copy/cell) in the present study. Never-theless, it will be critical to follow human patientsthrough liver biopsy to address this issue fully.
Three major conclusions can be drawn from these dataregarding the future of AAV as a gene vector for treatinggenetic disorders. First of all, the higher potency ofAAV2/8 vectors investigated here can mean a saferapplication of the virus. When a lower dose of the virusis required to achieve the same therapeutic effect, theamount of antigen introduced and subsequent immuno-genic potential of the vector can be reduced, as well assecondary site infections when introduced into thecirculation. Secondary benefits of this attribute ofAAV2/8 may be a safer drug and an increased potentialfor multiple injections without rejection if needed.Finally, given current production limits and costs accom-panying the use of AAV at GLP and GMP standards for aclinical application, the ability to correct secreted andmonogenic disorders with lower doses could considerablyexpand the utility of the system.
873

ARTICLE doi:10.1016/j.ymthe.2005.05.016
METHODS
Producing and purifying rAAV vectors. The University of Florida Powell
Gene Therapy Center produced rAAV serotype 1, 2, 5, and 8 vectors for
these studies. The techniques for AAV2 used included heparan-affinity
column chromatography as previously reported [14]. Virus production
included use of the helper/packaging plasmid pDG, which supplies all the
necessary helper functions as well as rep and cap in trans. Vector plasmid
(622.5 Ag) pCB-AAT was cotransfected by calcium phosphate precipitation
with 1867 Ag of pDG into one cell factory of ~70–95% confluent 293 cells.
Recombinant AAV was prepared by iodixanol centrifugation and hand-
packed heparan column purification. Physical titer (genome number) was
determined by dot blot. The infectious titer and extent of wild-type AAV2
contamination were determined by infectious center assay [15]. Spin
concentrators were used to desalt. The packaging protocols for AAV
pseudotypes are described in [16]. The AAV vectors produced were titered
by dot blot assay and results in particles per milliliter were AAV2/1, 1.75 �1013; AAV2/2, 6.16 � 1012; AAV2/5, 7.47 � 1013; and AAV2/8, 4.14 � 1012.
Portal vein injection and partial hepatectomy. Eight-week-old female
C57Bl/6 mice were purchased from the University of Florida Pathology
Mouse Colony and handled as approved by the University of Florida
Institutional Animal Care and Use Committee. Mice were randomly
assigned to treatment group. For portal vein injections, all animals were
anesthetized with 3% isoflurane. A ventral midline abdominal incision
was made into the peritoneal cavity, and the portal vein was exposed.
AAV vectors or PBS (100 Al) were administered into the portal vein using a
30-gauge needle (BD Pharmagen). Hemostasis was achieved by applica-
tion of a small piece of sterile swab directly onto the portal vein. Surgeries
were performed on a thermoregulated operating board designed to
maintain a temperature of 378C and routinely lasted 10 min. For
monitoring hAAT expression, serum samples were obtained via the tail
vein at weekly intervals (0, 1, 2, 4, 6, 8, 12, 16, and 24).
At 24 weeks post-AAV administration, a partial hepatectomy was
performed by excision of three lobes as previously described [17].
Although not adversely effecting the animalTs normal behavior and organ
system function, this procedure results in removal of up to 85% liver mass
and initiates liver regeneration with nearly total liver regeneration by 4
weeks. Resected tissue was divided into samples for genomic DNA
isolation and Southern/Q-PCR and histopathology.
Histopathology and immunohistochemistry. At 24 weeks post-AAV
administration, mice were euthanized and blood samples harvested for
serum. The liver and pancreas were completely removed. Representative
samples were snap frozen for subsequent genomic DNA isolation or fixed in
10% neutral-buffered formalin for histology and immunohistochemistry.
Formalin-fixed paraffin-embedded tissue sections (4 Am) were sequen-
tially deparaffinized, rehydrated, and blocked for endogenous peroxidase
activity. Following antigen retrieval in Target Retrieval solution (Dako-
Cytomation, Carpinteria, CA, USA), slides were serum blocked and
incubated with either rabbit anti-human AAT (1:100; Research Diagnostic
Institute, Flanders, NJ, USA) or normal rabbit immunoglobulin as a
negative control. Antibody binding was detected using the EnVision+
HRP kit (DakoCytomation) and DAB+ (DakoCytomation). Slides were
counterstained using hematoxylin (Vector Labs, Burlingame, CA, USA)
and mounted. Representative digital images were captured using a Zeiss
Axioskop equipped with an Axiocam camera. Camera exposure settings
were constant for all images.
Semiquantitative evaluation of human AAT immunoreactivity was
performed by an independent observer blinded to treatment groups.
Scoring was performed for six parameters (distribution of panlobular
staining (diffuse versus regional), individual cell intensity (perivenous,
periportal, pericentral), decile for numbers of intensely positive cells, and
nuclear staining). Scores ranged from 0 to 3 (none, mild, moderate,
intense).
ELISA for human a1-antitrypsin. Culture media and mouse serum levels
of AAT were analyzed by ELISA as previously described with minor
modifications [18]. Serum was diluted in PBS–0.5% Tween 20. The
874
substrate was TMB peroxidase (KPL) and the stopping buffer used was
H3PO4 (Fisher). Data are means F standard deviation.
Genomic DNA isolation and Southern blot. Total liver genomic DNA was
isolated from 300–400 mg of tissue by a previously published protocol
[18]. Following digestion of tissue with proteinase K, restriction digests
were performed with 40 Ag DNA with 100 units of either ApaI or HindIII
(NEB) overnight at 378C. The next day, 10 units of additional enzyme was
added and incubated for 2 h. Genomic DNA (40 Ag/lane) was electro-
phoresed in a 1% agarose gel and transferred to membrane for subsequent
Southern analysis as described [18].
Genomic DNA extraction and quantitative PCR. Isolation of genomic
DNA was performed with the Qiagen DNAeasy Tissue Kit and DNA
concentrations (1:25) were determined. One microgram of DNA was used
in all quantitative PCRs according to a previously used protocol in our lab
[19] and reaction conditions followed those recommended by the
manufacturer to include 50 cycles of 948C for 40 s, 378C for 2 min,
558C for 4 min, and 688C for 30 s. Primer pairs were designed for the CMV
enhancer/chicken h-actin promoter as described [20] and a standard curve
was established using the pCBAT plasmid.
ACKNOWLEDGMENTS
The authors acknowledge Tina Yanchis and Kristi Vale-Cruz, Molecular
Pathology Core, for technical assistance. This work was supported by grants
from the NIDDK (DK58327) and NHLBI (HL59412, HL69877) and the Alpha
One Foundation.
RECEIVED FOR PUBLICATION APRIL 11, 2005; REVISED MAY 10, 2005;
ACCEPTED MAY 24, 2005.
REFERENCES1. Flotte, T. R., et al. (2004). Phase I trial of intramuscular injection of a recombinant adeno-
associated virus alpha 1-antitrypsin (rAAV2-CB-hAAT) gene vector to AAT-deficient
adults. Hum. Gene Ther. 15: 93 – 128.
2. Perlmutter, D. H. (1993). Liver disease associated with alpha 1-antitrypsin deficiency.
Prog. Liver Dis. 11: 139 – 165.
3. Brantly, M. L., et al. (1991). Use of a highly purified alpha 1-antitrypsin standard to
establish ranges for the common normal and deficient alpha 1-antitrypsin phenotypes.
Chest 100: 703 – 708.
4. Ogushi, F., et al. (1988). Evaluation of the S-type of alpha-1-antitrypsin as an in vivo
and in vitro inhibitor of neutrophil elastase. Am. Rev. Respir. Dis. 137: 364 – 370.
5. Wewers, M. D., et al. (1987). Replacement therapy for alpha 1-antitrypsin deficiency
associated with emphysema. N. Engl. J. Med. 316: 1055 – 1062.
6. Wewers, M. D., Casolaro, M. A., and Crystal, R. G. (1987). Comparison of alpha-1-
antitrypsin levels and antineutrophil elastase capacity of blood and lung in a patient
with the alpha-1-antitrypsin phenotype null(null before and during alpha-1-antitrypsin
augmentation therapy. Am. Rev. Respir. Dis. 135: 539 – 543.
7. Casolaro, M. A., et al. (1987). Augmentation of lung antineutrophil elastase capacity
with recombinant human alpha-1-antitrypsin. J. Appl. Physiol. 63: 2015 – 2023.
8. Levy, M. Y., Barron, L. G., Meyer, K. B., and Szoka, F. C., Jr., (1996). Characterization of
plasmid DNA transfer into mouse skeletal muscle: evaluation of uptake mechanism,
expression and secretion of gene products into blood. Gene Ther. 3: 201 – 211.
9. Qui, P., Zeigelhoffer, P., Sun, J., and Yang, N. S. (1996). Gene gun delivery of mRNA in
situ results in efficient gene transgene expression and genetic immunization. Gene Ther.
3: 262 – 268.
10. Rosenfeld, M. A., et al. (1991). Adenovirus-mediated transfer of a recombinant alpha
1-antitrypsin gene to the lung epithelium in vivo. Science 252: 431 – 434.
11. Setoguchi, Y., Jaffe, H. A., Danel, C., and Crystal, R. G. (1994). Ex vivo and in vivo gene
transfer to the skin using replication-deficient recombinant adenovirus vectors. J. Invest.
Dermatol. 102: 415 – 421.
12. Zern, M. A., et al. (1999). A novel SV40-based vector successfully transduces and
expresses an alpha 1-antitrypsin ribozyme in a human hepatoma-derived cell line. Gene
Ther. 6: 114 – 120.
13. Gao, G. P., et al. (2002). Novel adeno-associated viruses from rhesus monkeys as
vectors for human gene therapy. Proc. Natl. Acad. Sci. USA 99: 11854 – 11859.
14. Conway, J., et al. (1999). High-titer recombinant adeno-associated virus production
utilizing a recombinant herpes simplex virus type I vector expressing AAV-2 Rep and
Cap. Gene Ther. 6: 986 – 993.
15. Walz, C. M., et al. (1998). Detection of infectious adeno-associated virus particles in
human cervical biopsies. Virology 247: 97 – 105.
16. Zolotukhin, S., et al. (2002). Production and purification of serotype 1, 2, and 5
recombinant adeno-associated viral vectors. Methods 28: 158 – 167.
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therapy

ARTICLEdoi:10.1016/j.ymthe.2005.05.016
17. Song, S., et al. (2004). DNA-dependent PK inhibits adeno-associated virus DNA
integration. Proc. Natl. Acad. Sci. USA 101: 2112 – 2116.
18. Song, S., et al. (2001). Stable therapeutic serum levels of human alpha-1 antitrypsin
(AAT) after portal vein injection of recombinant adeno-associated virus (rAAV) vectors.
Gene Ther. 8: 1299 – 1306.
19. Song, S., et al. (2002). Intramuscular administration of recombinant adeno-associated
virus 2 a-1 antitrypsin (rAAV-SERPINA1) vectors in a nonhuman primate model: safety
and immunologic aspects. Mol. Ther. 6: 329 – 335.
20. Donsante, A., et al. (2001). Observed incidence of tumorigenesis in long-term rodent
studies of rAAV vectors. Gene Ther. 8: 1343 – 1346.
21. Flotte, T., et al. (2001). Efficient ex vivo transduction of pancreatic islet cells with
recombinant adeno-associated virus vectors. Diabetes 50: 515 – 520.
22. Zabner, J., et al. (2000). Adeno-associated virus type 5 (AAV5) but not AAV2 binds
to the apical surfaces of airway epithelia and facilitates gene transfer. J. Virol. 74:
3852 – 3858.
23. Qiu, J., Nayak, R., Tullis, G. E., and Pintel, D. J. (2002). Characterization of the
transcription profile of adeno-associated virus type 5 reveals a number of unique
MOLECULAR THERAPY Vol. 12, No. 5, November 2005
Copyright C The American Society of Gene Therapy
features compared to previously characterized adeno-associated viruses. J. Virol. 76:
12435 – 12447.
24. Song, S., Laipis, P. J., Berns, K. I., and Flotte, T. R. (2001). Effect of DNA-dependent
protein kinase on the molecular fate of the rAAV2 genome in skeletal muscle. Proc. Natl.
Acad. Sci. USA 98: 4084 – 4088.
25. Thomas, C. E., Storm, T. A., Huang, Z., and Kay, M. A. (2004). Rapid uncoating of
vector genomes is the key to efficient liver transduction with pseudotyped adeno-
associated virus vectors. J. Virol. 78: 3110 – 3122.
26. Xiao, W., et al. (1998). Adeno-associated virus as a vector for liver-directed gene
therapy. J. Virol. 72: 10222 – 10226.
27. Kang, E. M., and Tisdale, J. F. (2004). The leukemogenic risk of integrating retroviral
vectors in hematopoietic stem cell gene therapy applications. Curr. Hematol. Rep. 3:
274 – 281.
28. Relph, K., Harrington, K., and Pandha, H. (2004). Recent developments and current
status of gene therapy using viral vectors in the United Kingdom. BMJ 329: 839 – 842.
29. Manno, C. S., et al. (2003). AAV-mediated factor IX gene transfer to skeletal muscle in
patients with severe hemophilia B. Blood 101: 2963 – 2972.
875





























