Embed Size (px)
Citation preview

Behavioural Brain Research 196 (2009) 37–43
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
Research report
Electroconvulsive shock alters the rat overt rhythms of motor activityand temperature without altering the circadian pacemaker
Montserrat Anglès-Pujolràsa, Antoni Díez-Nogueraa, Virginia Soriab, Mikel Urretavizcayab,c,José Manuel Menchónb,c, Trinitat Cambrasa,∗
a Departament de Fisiologia, Facultat de Farmàcia, Universitat de Barcelona, Av Joan XXIII s/n, 08028 Barcelona, Catalonia, Spainb Mood Disorders Clinical and Research Unit, CIBER-SAM, Psychiatry Department, Bellvitge University Hospital, Feixa Llarga s/n, L’Hospitalet de Llobregat,Barcelona 08907, Catalonia, Spainc Department of Clinical Sciences, Bellvitge Campus, Barcelona University, Barcelona, Catalonia, Spain
a r t i c l e i n f o
Article history:Received 22 May 2008Received in revised form 8 July 2008Accepted 10 July 2008Available online 25 July 2008
Keywords:Electroconvulsive therapyCircadian rhythmMotor activityTemperatureSuprachiasmatic nucleusElectroconvulsive shock
a b s t r a c t
The hypothetical relationship between circadian rhythms alterations and depression has prompted stud-ies that examine the resultant effects of various antidepressants. Electroconvulsive therapy (ECT) exertssignificant antidepressant effects that have been modelled in the laboratory via the use of electrocon-vulsive shock (ECS) in rats. However, data on the effects of ECT or ECS vis-à-vis the circadian rhythmsremain scarce. Thus, we report here the effects of acute and chronic ECS administration on the tempera-ture and motor activity circadian rhythms of rats. The motor activity and core body temperature of ratswere continuously recorded to determine the circadian rhythms. We carried out three experiments. Inthe first, we analyzed the effects of acute ECS on both the phase and period when applied at differenttimes of the subjective day. In the second and third experiments ECS was nearly daily applied to ratsfor 3 weeks: respectively, under dim red light, which allows a robust free-running circadian rhythm; andunder light–dark cycles of 22 h (T22), a setting that implies dissociation in the circadian system. Acute ECSdoes not modify the phase or the period of circadian rhythms. Chronic administration of ECS produces an
increase in motor activity and temperature, a decrease in the amplitude of circadian rhythms, althoughthe period of the free-running rhythm remains unaffected. In conclusion, while chronic ECS does alterthe overt rhythms of motor activity and temperature, it does not modify the functioning of the circadian1
mgwTratiorSi
ariiotrIfsu
0d
pacemaker.
. Introduction
In mammals, circadian rhythms are driven by a central pace-aker located in the suprachiasmatic nucleus (SCN). The SCN
enerates the genetically determined endogenous periodicityhich, under constant environment, is slightly different from 24 h.
o entrain an organism to 24 h light–dark cycle, the pacemakerequires regular environmental signals. In animals, such signalsre mainly provided by light information, which is transmittedo the SCN by retinal ganglion cells [15]. The SCN also receivesnput from other brain regions such as the intergeniculate leaflet
f the thalamus and the raphe nucleus [29]. The latter’s main neu-otransmitter is serotonin, which provides nonphotic input to theCN [28]. The circadian pacemaker is responsible for coordinat-ng physiologic rhythms within the body. This is accomplished by∗ Corresponding author. Tel.: +34 934 024 505; fax: +34 934 035 901.E-mail address: [email protected] (T. Cambras).
srm
dboS
166-4328/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2008.07.021
© 2008 Elsevier B.V. All rights reserved.
set of direct and indirect projections from the SCN onto otheregions of the brain. Under normal conditions most of an organ-sm’s rhythms exhibit the same circadian period, thus reflecting annternal temporal order. However, humans kept under isolation [39]r under forced desynchronization [7], show a divergence betweenhe timing of body rhythms and the light–dark cycle and exhibithythms with different periods between temperature and activity.t has been suggested that this rhythmic disturbance can have pro-ound effects on mood, sleep and health [40] and may attend jet-lag,hift work or even depression [9]. It has also been suggested thatnavoidable disturbances in circadian rhythms can trigger depres-ive episodes in humans [40]. We have proposed that submittingats to T22, situation that induces dissociation in the overt rhythms,ay prove an effective model for human desynchronization [2].
Circadian rhythm alterations have also been described in moodisorders. Depressive patients frequently experience sleep distur-ances, as well as alterations in hormonal secretion (e.g., cortisolr melatonin) and in temperature circadian rhythms [12,33,37].uch changes are mainly characterized by a reduction in rhythm

3 ural B
as(rdattasgtp
afiatipEat[aammEteatca
awetoaamli
2
2
Tirtpmrattd3seltt
ctl
2
1Btir
2
pfdcCCTioatiTie
dcevEnbpttcpa6vpa
cam
au(AiEsAtMtE
paaaam
8 M. Anglès-Pujolràs et al. / Behavio
mplitude, which has been shown to normalize during remis-ion. It is noticeable that an antidepressant, such as agomelatinea melatonergic receptor agonist and 5HT2c receptor antagonist)esynchronizes human circadian rhythms in healthy volunteers andepressed patients [10], as well as rats, which when administeredgomelatine daily were entrained to a 24 h cycle [14]. Moreover,he SCN involves some of the same neurotransmitters thoughto be important in depression, such as serotonin [28,4,22,42,20]nd gamma-aminobutyric acid (GABA) [29]. Taken together, thisuggests that circadian abnormalities may play a role in the patho-enesis of depression. However, how circadian rhythms are relatedo depression still remains unknown and also the effect of antide-ressants on the circadian system.
Electroconvulsive therapy (ECT) is a well-established, effective,nd quick treatment for depression. Indeed, it may be considered arst-line treatment in cases of severe depression, depression char-cterized by specific symptoms (psychosis or catatonia), or suicidalendencies. The intensity of ECT typically administered to patientsnduces a self-sustained after-discharge of cortical neurons thatroduces electroconvulsive seizures. The beneficial effects fromCT may stem not from the convulsion itself, but rather from thenticonvulsant effects of the seizure that result from enhancedransmission of inhibitory neurotransmitters and neuropeptides11]. Electroconvulsive shock (ECS) has been widely used as annimal model of ECT and has, moreover, helped elucidate the ther-peutic profile and side effects of ECT [13]. Although ECS or ECTainly affects hippocampus and neocortex [25], the exact actionechanisms are not yet well known. The mechanism underlying
CS is thought to involve a large number or neurotransmitters sys-ems [11,31,8]. In addition, ECT increases plasma prolactin [24,36],xerts an effect on neuronal plasticity and neurogenesis [1] and itlso reportedly increases the amplitude of the core body tempera-ure in depressed subjects [37]. However, data on ECS’s effects onircadian rhythms remain scare, with an attendant lack of system-tic studies.
In this study, we directly examined the effects of ECS on motorctivity and temperature circadian rhythms in rats. To this end,e carried out three experiments. In the first, we tested the acute
ffects of ECS administration during different times of the subjec-ive day of the animal, while in the second we evaluated the effectsf chronic ECS treatment on free-running circadian rhythms. Welso hypothesized whether ECS would exert different effects onn altered circadian system. Thus, we conducted a third experi-ent examining the effects of chronic ECS in rats submitted to T22
ight–dark cycles, which, as mentioned above, implies dissociationn the rat’s circadian rhythms.
. Materials and methods
.1. Animals and experimental conditions
Forty-six male Wistar rats (Charles River, France) were used for the experiments.he animals arrived at the laboratory at age 4 weeks and were individually housedn transparent cages (25 cm × 25 cm × 12 cm), maintained in three different isolatedooms (one for each experiment, see below), with access to food and water ad libi-um. The motor activity rhythm was detected by activity meters having two crossed,erpendicular infrared beams crossing the cage 7 cm above the floor. The number ofovements was recorded every 15 min. Temperature was measured (with an accu-
acy of 0.125 ◦C) via data loggers (Thermochron®) implanted intraperitoneally in thenimals following administration of isofluorane anesthesia. To avoid alterations inhe animal’s rhythm during the experiment, the surgery was carried out just prioro the start of experiments. Data loggers were programmed to start recording onifferent days depending on the experiment, but always with a sampling period of
0 min. At the end of the experiments, animals were sacrificed and the temperatureensors were removed to acquire the data. Lighting conditions depended on thexperiment. In experiments 1 and 2 rats were maintained under constant dim redight (0.40–2.20 lx). This light intensity was chosen since it not only allowed animalso maintain a robust free-running circadian rhythm, but also permitted researcherso handle them. In experiment 3 rats were submitted to a 22 h period light–darkeTlrsp
rain Research 196 (2009) 37–43
ycle (T22, 11 h-light and 11 h-dark). In this case, darkness was a dim red light withhe same intensity as in the other rooms, while the light phase consisted in indirectight provided by two fluorescent tubes, with an intensity of 300 lx at cage level.
.2. Electroconvulsive shock administration
Following attachment of saline-soaked earclips, ECS consisted of a 1 s,00 pulses/s, 90 mA stimulus of 0.5 ms square-wave pulses delivered using a UGOasile ECT unit, model 57800-001. At these settings, all animals had generalizedonic-clonic seizures with hind limb extension. ECS administrations were performedn an adjacent room without altering the lighting conditions of the experimentaloom.
.3. Experimental procedure
Experiment 1: This experiment was designed to test whether the phase anderiod of motor activity and temperature rhythms changed by ECS applied at dif-
erent times of the subjective day. To do so, 10 rats were maintained under constantim red light for 76 days. Every 7–10 days, each animal received one ECS at differentircadian time (CT), with CT0 serving as the onset of the resting phase for rats andT12 as the onset of the active phase. The CTs assayed were as follows: CT0, CT4, CT8,T12 and CT20. Thus, each rat received a total of five ECS in the whole experiment.o calculate the time of each CT, the day before ECS administration, a line intersect-ng the daily onsets of activity of the last days was drawn to extrapolate the CT12ccurring the following day. The corresponding number of circadian hours was thendded or subtracted to CT12 in order to obtain the exact time of each specific CT. Athe end of the experiment, the CTs at which animals received ECS were revised, andn some cases corrected, based on the entire motor activity and temperature plot.hus, more CTs than initially envisaged were obtained. In this experiment, activ-ty and temperature in eight rats were continuously recorded through the entirexperimental period.
Data analysis consisted in the determination of the period and phase shifts pro-uced by a single ECS. To determine the former, we calculated the period-responseurve (tauRC), which measures the period change produced by the ECS at a differ-nt circadian time. To this end, we calculated a periodogram for each animal andariable with the data that corresponded to 6 days before and 6 days after eachCS application. The tau value from a single stage was then subtracted from theext, thereby obtaining the tau shift value for each CT. These shifts were plottedased on the corresponding CT and the tauRC was subsequently drawn. Since noeriod changes were obtained, to calculate the phase response curve (PRC), this ishe phase shifts produced by a single ECS, we proceed as follows. We first calculatedhe mean period of the endogenous rhythm (tau) for each animal by means of ahi-squared periodogram [35], which was applied to all days of the experimentaleriod. We then conducted cosinor analysis for each animal and variable (motorctivity and temperature) data, with a period equal to tau for 6 days before anddays after ECS application, thereby obtaining the corresponding acrophases. The
alue of one acrophase was subtracted from the next and the phase shift value waslotted according to the CT. We calculated tauRC and PRC for both temperature andctivity data.
Statistical analysis for motor activity and temperature was carried out to cal-ulate the differences from 0, in phase and period shifts, in each CT, by means ofone-sample t-test, and to determine the differences between the various CTs byeans of ANOVAs. In all cases, Bonferroni’s correction was applied.
Experiment 2: This experiment was designed to test the effects of ECS chronicdministration on the free-running rhythms of motor activity and temperature. Wesed 16 male Wistar rats divided in two groups: experimental (n = 8) and controln = 8). During the whole experiment animals were maintained under dim red light.fter 30 days under free-running conditions (pre-ECS stage), each rat of the exper-
mental group received four to six ECS per week for 3 weeks (ECS stage) always anCS per day between 11 and 12 a.m. At this stage, control rats were handled in theame way as the experimental rats, albeit without receiving any electrical current.fterwards, rats were kept under the identical conditions (post-ECS stage) in order
o determine motor activity and temperature rhythms following ECS application.otor activity was recorded for all rats throughout the entire experiment, while
emperature was recorded only for six experimental and five control rats during theCS and post-ECS stages.
Motor activity and temperature data were analyzed separately. To calculate theeriods and percentages of variance (%V) explained by the corresponding rhythm, wepplied a chi squared periodogram [35] to data for 18 days from every stage for eachnimal. %V is a measure for the stability (strength) of the rhythm. The mean periodnd the mean %V were then calculated for each group (experimental and control),)nd each stage (pre-ECS, ECS and post-ECS). In addition, the daily mean values forotor activity and temperature, were calculated for each rat and for each day the
xperiment lasted, to study the evolution of these values along the experiment.hese mean values were calculated for data series corresponding to the tau periodength and expressed as deviation from the mean. To determine variations in thehythm expression, we carried out, for motor activity and temperature data, a powerpectrum analysis to those data sets corresponding to the length of the endogenouseriod (tau). The power content for the first spectrum harmonic (PCH1) reflects the
ural B
alsct
rswipCritcrpgpa
Ttat
duspC
3
Faata
M. Anglès-Pujolràs et al. / Behavio
mplitude of the free-running period. Statistical analysis (ANOVA) involved severalinear models, taking into account period and %V as dependent variables, and thetage of the experiment (pre-ECS, ECS and post-ECS) and group (experimental andontrol) as independent variables. Groups were also compared using the Student’s-test with Bonferroni’s correction.
Experiment 3: In this experiment the effect of chronic ECS application on theat’s dissociated circadian rhythms vis-à-vis motor activity and temperature wastudied. In this case, 20 male Wistar rats (10 experimental and 10 control rats)ere submitted to a 22 h period light–dark cycle (T22) throughout the entire exper-
ment. After 30 days under T22 (pre-ECS stage), experimental rats received 4–6 ECSer week for 3 weeks (ECS stage), always an ECS per day between 11 and 12 a.m.ontrol rats were similarly handled, albeit without electrical current. Finally, ratsemained for 24 more days in the cage to test the rhythms following ECS admin-stration (post-ECS stage). Motor activity was recorded for all the rats throughouthe entire experiment. Temperature was recorded only for six experimental and five
ontrol rats during the pre-ECS and ECS stages. Under T22, two significant circadianhythms were observed regarding motor activity and temperature: one exhibiting aeriod equal to 22 h (light-dependent component, LDC) and another with a periodreater than 24 h (non-light-dependent component, NLDC). Here, to calculate theeriods and %V of the two rhythms (“%VLDC and %VNLDC) we used a periodogramnalysis [35] applied to 19 days data of each stage and for each individual animal.fwtao
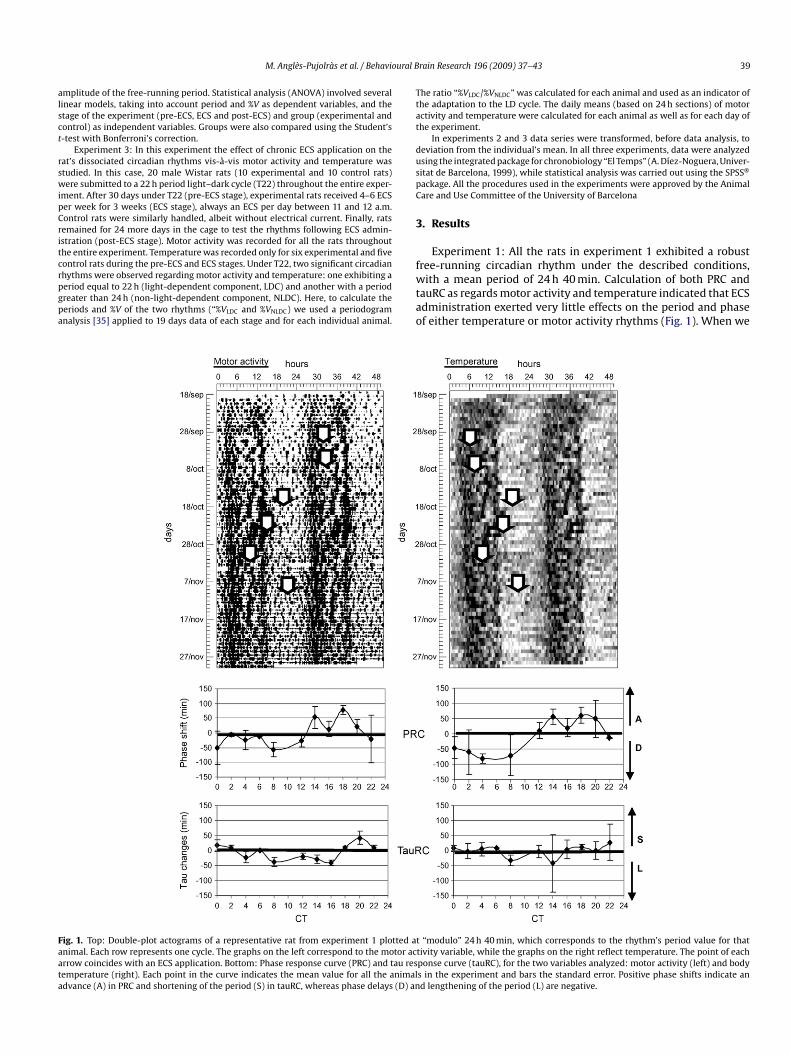
ig. 1. Top: Double-plot actograms of a representative rat from experiment 1 plotted animal. Each row represents one cycle. The graphs on the left correspond to the motor acrrow coincides with an ECS application. Bottom: Phase response curve (PRC) and tau resemperature (right). Each point in the curve indicates the mean value for all the animaldvance (A) in PRC and shortening of the period (S) in tauRC, whereas phase delays (D) an
rain Research 196 (2009) 37–43 39
he ratio “%VLDC/%VNLDC” was calculated for each animal and used as an indicator ofhe adaptation to the LD cycle. The daily means (based on 24 h sections) of motorctivity and temperature were calculated for each animal as well as for each day ofhe experiment.
In experiments 2 and 3 data series were transformed, before data analysis, toeviation from the individual’s mean. In all three experiments, data were analyzedsing the integrated package for chronobiology “El Temps” (A. Díez-Noguera, Univer-itat de Barcelona, 1999), while statistical analysis was carried out using the SPSS®
ackage. All the procedures used in the experiments were approved by the Animalare and Use Committee of the University of Barcelona
. Results
Experiment 1: All the rats in experiment 1 exhibited a robust
ree-running circadian rhythm under the described conditions,ith a mean period of 24 h 40 min. Calculation of both PRC andauRC as regards motor activity and temperature indicated that ECSdministration exerted very little effects on the period and phasef either temperature or motor activity rhythms (Fig. 1). When we
t “modulo” 24 h 40 min, which corresponds to the rhythm’s period value for thattivity variable, while the graphs on the right reflect temperature. The point of eachponse curve (tauRC), for the two variables analyzed: motor activity (left) and bodys in the experiment and bars the standard error. Positive phase shifts indicate and lengthening of the period (L) are negative.
40 M. Anglès-Pujolràs et al. / Behavioural Brain Research 196 (2009) 37–43
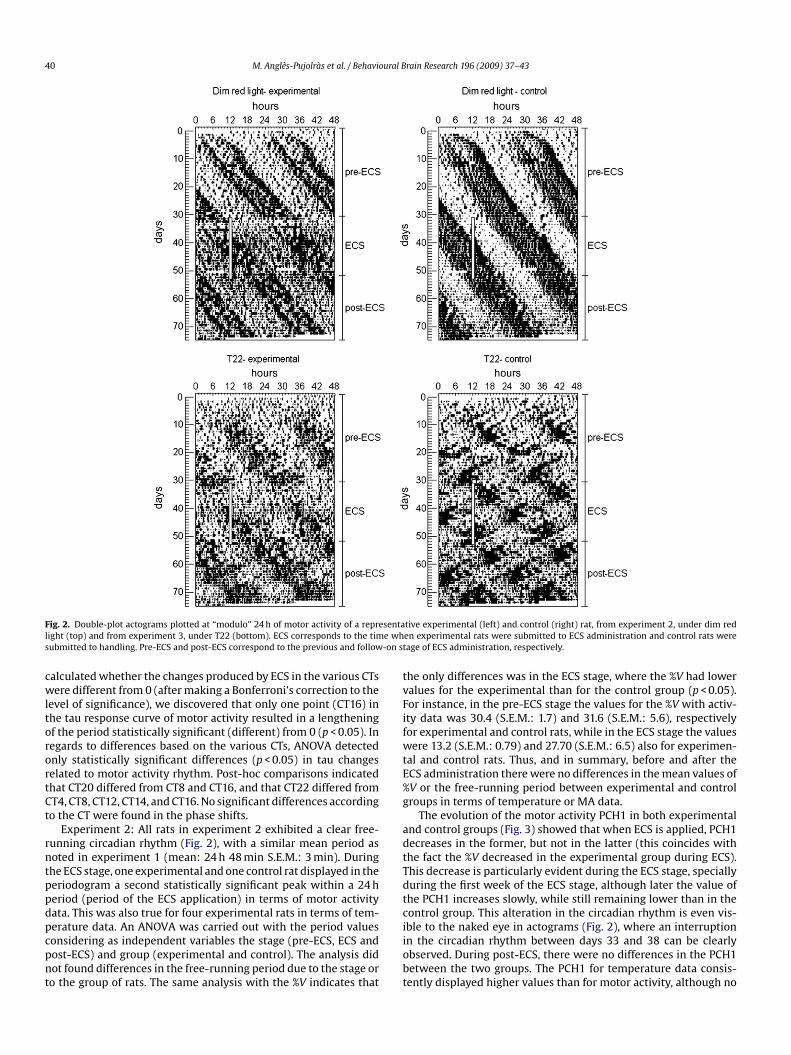
F sental e whs -on st
cwltorortCt
rntppdpcpnt
tvFifwtE%g
adtTdtc
ig. 2. Double-plot actograms plotted at “modulo” 24 h of motor activity of a repreight (top) and from experiment 3, under T22 (bottom). ECS corresponds to the timubmitted to handling. Pre-ECS and post-ECS correspond to the previous and follow
alculated whether the changes produced by ECS in the various CTsere different from 0 (after making a Bonferroni’s correction to the
evel of significance), we discovered that only one point (CT16) inhe tau response curve of motor activity resulted in a lengtheningf the period statistically significant (different) from 0 (p < 0.05). Inegards to differences based on the various CTs, ANOVA detectednly statistically significant differences (p < 0.05) in tau changeselated to motor activity rhythm. Post-hoc comparisons indicatedhat CT20 differed from CT8 and CT16, and that CT22 differed fromT4, CT8, CT12, CT14, and CT16. No significant differences accordingo the CT were found in the phase shifts.
Experiment 2: All rats in experiment 2 exhibited a clear free-unning circadian rhythm (Fig. 2), with a similar mean period asoted in experiment 1 (mean: 24 h 48 min S.E.M.: 3 min). Duringhe ECS stage, one experimental and one control rat displayed in theeriodogram a second statistically significant peak within a 24 heriod (period of the ECS application) in terms of motor activityata. This was also true for four experimental rats in terms of tem-
erature data. An ANOVA was carried out with the period valuesonsidering as independent variables the stage (pre-ECS, ECS andost-ECS) and group (experimental and control). The analysis didot found differences in the free-running period due to the stage oro the group of rats. The same analysis with the %V indicates thatiiobt
tive experimental (left) and control (right) rat, from experiment 2, under dim reden experimental rats were submitted to ECS administration and control rats wereage of ECS administration, respectively.
he only differences was in the ECS stage, where the %V had loweralues for the experimental than for the control group (p < 0.05).or instance, in the pre-ECS stage the values for the %V with activ-ty data was 30.4 (S.E.M.: 1.7) and 31.6 (S.E.M.: 5.6), respectivelyor experimental and control rats, while in the ECS stage the valuesere 13.2 (S.E.M.: 0.79) and 27.70 (S.E.M.: 6.5) also for experimen-
al and control rats. Thus, and in summary, before and after theCS administration there were no differences in the mean values ofV or the free-running period between experimental and controlroups in terms of temperature or MA data.
The evolution of the motor activity PCH1 in both experimentalnd control groups (Fig. 3) showed that when ECS is applied, PCH1ecreases in the former, but not in the latter (this coincides withhe fact the %V decreased in the experimental group during ECS).his decrease is particularly evident during the ECS stage, speciallyuring the first week of the ECS stage, although later the value ofhe PCH1 increases slowly, while still remaining lower than in theontrol group. This alteration in the circadian rhythm is even vis-
ble to the naked eye in actograms (Fig. 2), where an interruptionn the circadian rhythm between days 33 and 38 can be clearlybserved. During post-ECS, there were no differences in the PCH1etween the two groups. The PCH1 for temperature data consis-ently displayed higher values than for motor activity, although no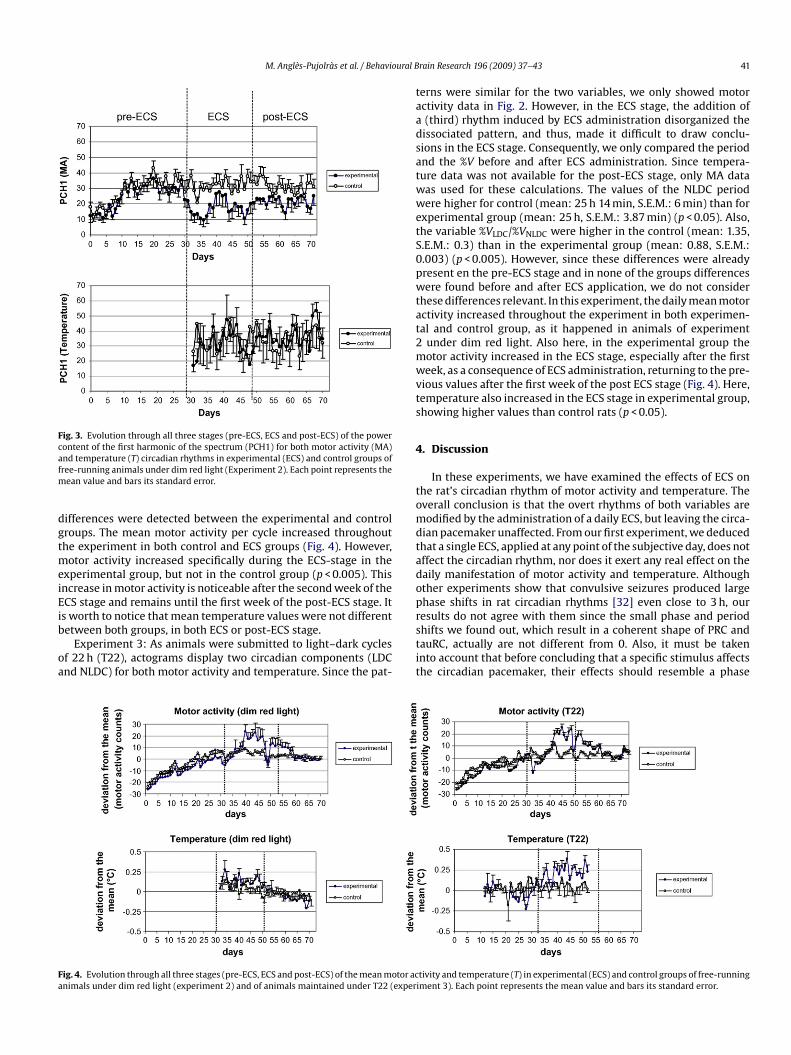
M. Anglès-Pujolràs et al. / Behavioural B
Fig. 3. Evolution through all three stages (pre-ECS, ECS and post-ECS) of the powercontent of the first harmonic of the spectrum (PCH1) for both motor activity (MA)afm
dgtmeiEib
oa
taadsatwwetS0pwtat2mwvts
4
tomdtadopresults do not agree with them since the small phase and period
Fa
nd temperature (T) circadian rhythms in experimental (ECS) and control groups ofree-running animals under dim red light (Experiment 2). Each point represents the
ean value and bars its standard error.
ifferences were detected between the experimental and controlroups. The mean motor activity per cycle increased throughouthe experiment in both control and ECS groups (Fig. 4). However,
otor activity increased specifically during the ECS-stage in thexperimental group, but not in the control group (p < 0.005). Thisncrease in motor activity is noticeable after the second week of theCS stage and remains until the first week of the post-ECS stage. Its worth to notice that mean temperature values were not different
etween both groups, in both ECS or post-ECS stage.Experiment 3: As animals were submitted to light–dark cyclesf 22 h (T22), actograms display two circadian components (LDCnd NLDC) for both motor activity and temperature. Since the pat-
stit
ig. 4. Evolution through all three stages (pre-ECS, ECS and post-ECS) of the mean motor acnimals under dim red light (experiment 2) and of animals maintained under T22 (exper
rain Research 196 (2009) 37–43 41
erns were similar for the two variables, we only showed motorctivity data in Fig. 2. However, in the ECS stage, the addition of(third) rhythm induced by ECS administration disorganized theissociated pattern, and thus, made it difficult to draw conclu-ions in the ECS stage. Consequently, we only compared the periodnd the %V before and after ECS administration. Since tempera-ure data was not available for the post-ECS stage, only MA dataas used for these calculations. The values of the NLDC periodere higher for control (mean: 25 h 14 min, S.E.M.: 6 min) than for
xperimental group (mean: 25 h, S.E.M.: 3.87 min) (p < 0.05). Also,he variable %VLDC/%VNLDC were higher in the control (mean: 1.35,.E.M.: 0.3) than in the experimental group (mean: 0.88, S.E.M.:.003) (p < 0.005). However, since these differences were alreadyresent en the pre-ECS stage and in none of the groups differencesere found before and after ECS application, we do not consider
hese differences relevant. In this experiment, the daily mean motorctivity increased throughout the experiment in both experimen-al and control group, as it happened in animals of experiment
under dim red light. Also here, in the experimental group theotor activity increased in the ECS stage, especially after the firsteek, as a consequence of ECS administration, returning to the pre-
ious values after the first week of the post ECS stage (Fig. 4). Here,emperature also increased in the ECS stage in experimental group,howing higher values than control rats (p < 0.05).
. Discussion
In these experiments, we have examined the effects of ECS onhe rat’s circadian rhythm of motor activity and temperature. Theverall conclusion is that the overt rhythms of both variables areodified by the administration of a daily ECS, but leaving the circa-
ian pacemaker unaffected. From our first experiment, we deducedhat a single ECS, applied at any point of the subjective day, does notffect the circadian rhythm, nor does it exert any real effect on theaily manifestation of motor activity and temperature. Althoughther experiments show that convulsive seizures produced largehase shifts in rat circadian rhythms [32] even close to 3 h, our
hifts we found out, which result in a coherent shape of PRC andauRC, actually are not different from 0. Also, it must be takennto account that before concluding that a specific stimulus affectshe circadian pacemaker, their effects should resemble a phase
tivity and temperature (T) in experimental (ECS) and control groups of free-runningiment 3). Each point represents the mean value and bars its standard error.

4 ural B
rwecaIemirpataam
auErltTp
biTcaotfitttcimri
stcEdnagtanp
tegtoamr
letpgamteds[ssimish
si[cipmsGMrtiwdioptpancfpt
octsoctadtice
2 M. Anglès-Pujolràs et al. / Behavio
esponse curve with that includes a dead zone, as well as zonesith phase advances and delays [18], which has not thus far been
ncountered out. In any way, both experiments suggest that thehanges in phase or tau, are most likely due to day-to-day fluctu-tions in the effector system, rather than in the brain pacemaker.t must also be taken into account that the rats used in the presentxperiment, are “normal”, this is, they are not depressive animalodels, and ECS has been reported, to have an effect, by increas-
ng the levels of NPY in the hypothalamus, only in flinder sensitiveats and not in control rats [17]. Our results are consistent withrevious experiments in the sense that the rat circadian systemppears little affected by non-photic stimuli [3]. Thus, we concludehat the effects of a single ECS on the circadian pacemaker, if any,re insignificant. This could be related to the fact that repeateddministrations of ECS are more likely to impact the therapeuticechanisms of ECT than would a single ECS [11].The lack of effect on the circadian pacemaker became even more
pparent when we carried out the second experiment, in which anndisturbed free-running rhythm remained unaffected after theCS treatment. Thus, daily ECS is not able to entrain the circadianhythms of rats, as other antidepressant agents, such as agome-atine [23], do. In this case, neither the period nor the phase ofhe free-running rhythms were affected by daily ECS application.he free-running rhythm after ECS application maintains the samehase and period the animal had before ECS application.
However, although the free-running rhythm does not changeefore and after the ECS application, it must be noticed that dur-
ng the ECS stage strong alterations in the overt rhythm are found.hese alterations are seen as a decrease in the amplitude of the cir-adian rhythm (detected by the PCH1) and by an increase in motorctivity. During the ECS administration, there was a noticeable lossf circadian rhythmicity, which was stronger during the first days ofhe ECS stage. However, in the post-ECS stage, the levels are not dif-erent from the control group. This lack of rhythmicity is reflectedn the actogram as strong disturbances of the circadian pattern, buthat lasted only the first days in the ECS stage. It must be noticedhat this loss of rhythmicity is also seen during the first week ofhe ECS stage in temperature data. However, in this variable, weould not find differences between experimental and control ratsn the ECS stage neither in the post-ECS stage. This lack of rhyth-
icity lasted only a few days and afterwards the circadian rhythmecovered the same phase and period as before ECS administration,ndicating no effect on the circadian clock.
The loss of rhythmicity is also visible in the third experiment, inpite of the complex rhythmic pattern of the animals under T22. Inhis case, the animals follow a dissociated behavioural pattern withlear manifestation of two circadian rhythms. The administration ofCS produced a strong disturbance in the circadian structure, ren-ering the circadian pattern disorganized. Although here, we couldot calculated the amplitude of the rhythms, since there is inter-ction between the two rhythms and the ECS administration, thatenerates a third rhythm, it is worth to notice that the disorganiza-ion of the rhythmic pattern lasted the first week of the ECS stage,nd later the rhythmic pattern is restored. Taking into account theon-light dependent component, we found no differences in theeriod before and after ECS application
It is interesting the effect that the administration of ECS has onhe mean levels of motor activity and temperature. Looking to thevolution of the motor activity in both experimental and controlroups under dim red light (experiment 2) it can be noticed that
he increase in motor activity actually takes places after more thanne week of ECS administration, and that lasted for another weekfter the end of the ECS stage. This suggests that the increase inotor activity do not correspond only to the manipulation of theat, (actually, we have measured the mean activity in the 4 h fol-
pmAmo
rain Research 196 (2009) 37–43
owing the ECS application, and these values did not differ betweenxperimental and control groups), but perhaps to a general state inhe animal’s behaviour. Thus, alterations in motor activity and tem-erature levels are due not only to reactivity to ECS but also to theeneral levels of these two variables. It is worth noting that daily ECSpplication did not modify the levels of temperature in the experi-ental animals under dim red light. This is important, since in the
hird experiment, not only motor activity but also temperature lev-ls increased because of ECS. This may stem the fact that ECS affectsissociated rats more than free-running rats, perhaps because oftress mechanisms. Since stress produces temperature increases27,38], it could be argued that dissociated animals are more sen-itive to external stimuli such as ECS, which could act then as aort of stressful factor. Thus, chronic ECS, added to the disturbancenduced by T22 results in a stressful state that induces hyperther-
ia. Perhaps restoration of the normal circadian rhythmicity couldmply a recovery from stress, which may be the cause of depressivetates. This fits with the supposition that rhythmic alterations inumans may trigger exacerbation of affective disorders [19].
The SCN can be entrained by external stimuli other than light,uch as serotonergic innervations from the raphe [6,30]. ECTnvolves massive discharge encompassing large areas of the brain21], and thus there are several pathways by which ECT could affectircadian rhythms. ECT induces transcriptional changes primarilyn the locus coeruleus [5], whose main neurotransmitter is nore-inephrine. The locus coeruleus is a well-known stress responseodulator since it receives input from other neurotransmitter
ystems providing information about homeostasis (e.g. serotonin,ABA, corticotropine releasing factor, dopamine and Glutamate).oreover, GABA is produced most by, if not all, the SCN neu-
ons [29]. It has been demonstrated that GABA concentrations inhe occipital cortex of depressed patients are lower and that theyncrease following ECT treatment [34]. Some, or all, of these path-ays may be responsible for alterations in overt rhythms. Theisturbance in the motor activity and temperature rhythms follow-
ng ECS administration may stem from the effects of ECS on areasf the brain that control both variables. However, the results of theresent experiment clearly show that the robust changes in overtemperature and activity rhythms are not mirrored by changes inacemaker function; i.e., either period or phase are affected by ECSdministration. This not only agrees with the fact that ECT doesot greatly alter the hypothalamus area [16], but also suggests thatortex inputs do not significantly affect the circadian system. There-ore, changes in temperature and activity are apparently caused byeripheral alterations to the circadian timing system, which maskshe output of the unaffected pacemaker.
One must be careful when drawing conclusions about the statef this internal clock when overt rhythms are measured, sincehanges in amplitude do not necessarily reflect alterations inhe circadian pacemaker. While some experiments reveal changestemming from stress or depression, evident in measurements ofvert circadian rhythms [26,38], more careful analysis [27] indi-ates that these alterations occur only in the overt rhythms ratherhan in the pacemaker itself. Our experiments show similar results,nd indicate that extensive recordings of circadian rhythms, before,uring and after a manipulation represent the only reliable wayo detect real changes in a pacemaker. In any case, this does notnvalidate the hypothesis contending that a relationship betweenircadian rhythms and depression exists, or the fact that ECT re-stablishes the amplitude of the circadian rhythms in depressed
atients [37]. While ECS does not affect the entire circadian pace-aker, it does induce reversible alterations in the system output.lthough, we did not study the effect of ECS on depressive ani-al models, it could be suggested that the antidepressant effectsf ECT do not affect the circadian pacemaker. Instead, via effectors

ural B
srsya
A
cHd
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[39] Wever RA. The circadian system of man. Results of experiments under temporal
M. Anglès-Pujolràs et al. / Behavio
ystems, it could help to re-establish the altered overt circadianhythms observed in depression to their previous norms. Our studyuggests the importance of carrying out careful behavioural anal-ses when addressing the functionality of the circadian system innimal models of depression.
cknowledgments
This work was financially supported by the Ministerio de Edu-ación y Ciencia (BFU2008-00199) and by the Spanish Ministry ofealth, Instituto de Salud Carlos III, Centro de Investigación en Rede Salud Mental (CIBER-SAM).
eferences
[1] Altar CA, Laeng P, Jurata LW, Brockman JA, Lemire A, Bullard J, et al. Electro-convulsive seizures regulate gene expression of distinct neurotrophic signalingpathways. J Neurosci 2004;24:2667–77.
[2] Cambras T, Weller JR, Anglès-Pujoràs M, Lee ML, Christopher A, Díez-NogueraA, et al. Circadian desynchronization of core body temperature and sleep stagesin the rat. Proc Natl Acad Sci USA 2007;104:7634–9.
[3] Canal-Corretger MM, Cambras T, Díez-Noguera A. Tau and phase responsecurves for non-photic stimuli in blinded rats. Biol Rhythm Res 2003;34:91–9.
[4] Carlsson A, Svenbnerhold L, Winblad B. Seasonal and circadian monoaminevariations in human brains examined post mortem. Acta Psychiatr Scand1980;61:75–85.
[5] Conti B, Maier R, Barr AM, Morale MC, Lu X, Sanna PP, et al. Region-specific transcriptional changes following the three antidepressant treatmentselectro convulsive therapy, sleep deprivation and fluoxetine. Mol Psychiatry2007;12:167–89.
[6] Cutrera RA, Kalsbeek A, Pevet P. Specific destruction of the serotoninergic affer-ents to the suprachiasmatic nuclei prevents triazolam-induced phase advancesof hamster activity rhythms. Behav Brain Res 1994;62:21–8.
[7] Czeisler CA, Duffy JF, Shanahan TL, Brown EN, Mitchell JF, Rimmer DW, et al.Stability, precision and near-24-h period of the human circadian pacemaker.Science 1999;284:2177–81.
[8] Dremenkov E, Gur E, Lerer B, Newman ME. Effects of chronica antidepressantsand electroconvulsive shock on serotonergic neurotransmission in the rat hip-pocampus. Prog Neuro-Psychopharmacol Biol Psychiatry 2003;27:729–39.
[9] The relevance of circadian rhythms for human welfare. In: Chronobiology. Bio-logical Timekeeping 2004;325–56.
10] Duval F, Lebowitz BD, Macher JP. Pharmacological aspects. Treatments indepression. In: Macher JP, Crocq MA, editors. Dialogues Clin Neurosci: Depres-sion 2006;8:191–206.
11] Eitan R, Lerer B. Nonpharmacological, somatic treatments of depression: elec-troconvulsive therapy and novel brain stimulation modalities. Dialogues ClinNeurosci Depression 2006;8:241–53.
12] Ford DE, Kamerow DB. Epidemiologic study of sleep disturbances and psychi-atric disorders. An opportunity for prevention? JAMA 1989;262:1479–84.
13] Green AR, Nutt DJ. Psychopharmacology of repeated seizures: possible rele-vance to the mechanisms of action of ECT. In: Iversen LL, Snyder SH, editors.Handbook of psychopharmacology. New York: Plenum Press; 1987. p. 375–419.
14] Guardiola-Lemaitre B. Agonistes et antagonistes des récepteurs mélatonin-ergiques: effets pharmacologiques et perspectives thérapeutiques. Ann PharmFr 2005;63:385–400.
15] Hattar S, Lucas RJ, Mrosovsky N, Thompson S, Douglas RH, Hankins MW, et al.Melanopsin and rod-cone photoreceptive systems account for all major acces-sory visual functions in mice. Nature 2003;424:76–81.
16] Ji RR, Schlaepfer TE, Aizenman CD, Epstein CM, Qiu D, Huang JC, et al. Repetitivetranscranial magnetic stimulation activates specific regions in rat brain. ProcNatl Acad Sci USA 1998;95:15635–40.
[
[
rain Research 196 (2009) 37–43 43
17] Jimenez-Vasquez P, Overstreet DH, Mathé AA. J Psychiat. Neuropeptide Y inmale and female brains of flinders sensitive line, a rat model of depression.Effects of electroconvulsive stimuli. J Psychiat Res 2000;4:405–12.
18] Johnson CH. Phase response curves: what can they tell us about circadianclocks? In: Hiroshige T, Honma K, editors. Circadian clocks from cell to human.Sapporo, Hokkaido: Univ Press; 1992. p. 209–19.
19] Katz G, Durst R, Xislin Y, Barel Y, Knobler HY. Psychiatric aspects of jet-lag:review and hypothesis. Med Hypotheses 2001;56:20–3.
20] Kennaway DJ. Light, neurotransmitters and the suprachiasmatic nucleus controlof pineal melatonin production in the rat. Biol Signals Recept 1997;6:247–54.
21] Kety S. Effects of repeated electroconvulsive shock on brain catecholamines. In:Fink M, Kety S, McGaugh JWTA, editors. Psychobiology of convulsive therapy.Washington, DC: Winston and Sons; 1974.
22] Lambert GW, Reid C, Kaye DM, Jennings GL, Esler MD. Effect of sunlight andseason on serotonin turnover in the brain. Lancet 2002;360:1840–2.
23] Martinet L, Guardiola-Lemaitre B, Mocaer E. Entrainment of circadian rhythmsby S-20008, a melatonin agonist, is dose and plasma concentration dependent.Pharmacol Biochem Behav 1996;54:713–8.
24] Markianos M, Hatzimanolis J, Lykouras L. Relationship between prolactinresponses to ECT and dopaminergic and serotonergic responsivity in depressedpatients. Eur Arch Psychiatry Clin Neurosci 2002;252:166–71.
25] Epstein CM. Electroconvulsive therapy: sixty years of progress and acomparison with transcranial magnetic stimulation and vagal nerve stim-ulation. Neuropsychopharmacology: the fifth generation of progress 2000.p. 1097–108.
26] Meerlo P, de Boer SF, Koolhaas JM, Daan S, Van den Hoofdakker RH. Changes indaily rhythms of body temperature and activity after a single social defeat inrats. Physiol Behav 1996;59:735–9.
27] Meerlo P, van den Hoofdakker RH, Koolhaas JM, Daan S. Stress-induced changesin circadian rhythms of body temperature and activity in rats are not caused bypacemaker changes. J Biol Rhythms 1997;12:80–92.
28] Moore RY, Speh JC. Serotonin innervation of the primate suprachiasmaticnucleus. Brain Res 2004;1010:169–73.
29] Moore RY, Speh JC, Leak RK. Suprachiasmatic nucleus organization. Cell TissueRes 2002;309(1):89–98.
30] Morin LP. SCN organization reconsidered. J Biol Rhythms 2007;22(1):3–13.31] Newman ME, Shapira B, Lerer B. Evaluation of central serotonergic func-
tion in affective and related disorders by the fenfluramine challenge test:a critical review. Int J Neuropsychopharmacol Biol Psychiatry 1998;27:729–39.
32] Quigg M, Straume M, Smith T, Menaker M, Bertram EH. Seizures induce phaseshifts of rat circadian rhythms. Brain Res 2001;913:165–9.
33] Rubin RT, Heist EK, McGeoy SS, Hanada K, Lesser IM. Neuroendocrine aspectsof primary endogenous depression. XI. Serum melatonin measures in patientsand matched control subjects. Arch Gen Psychiatry 1992;49:558–67.
34] Sanacora G, Fenton LR, Fasula MK, Rothman DL, Levin Y, Krystal JH, et al. Cor-tical gamma-aminobutyric acid concentration in depressed patients receivingcognitive behavioral therapy. Biol Psychiatry 2006;59:284–6.
35] The chi square periodogram: its utility for analysis of circadian rhythms. J TheorBiol 1978;8:72(1):131–60.
36] Swartz CM. Related neuroendocrine effects of electroconvulsive therapy (ECT).Psychopharmacol Bull 1997;33:265–71.
37] Szuba MP, Guze BH, Baxter Jr LR. Electroconvulsive therapy increases circa-dian amplitude and lowers core body temperature in depressed subjects. BiolPsychiatry 1997;15;42:1130–7.
38] Ushijima K, Morikawa T, To H, Higuchi S, Ohdo S. Chronobiological disturbanceswith hyperthermia and hypercortisolism induced by chronic mild stress in rats.Behav Brain Res 2006;173:326–30.
isolation. New York: Springer-Verlag; 1979.40] Wirz-Justice A. Biological rhythms in mood disorders. Neuropsychopharmacol-
ogy: the fifth generation of progress; 2000.42] Wirz-Justice A. Biological rhythm disturbances in mood disorders. Int Clin Psy-
chopharmacol 2006;21:11–5.





























