Embed Size (px)
Citation preview

www.elsevier.com/locate/pnpbp
Progress in Neuro-Psychopharmacology & B
Electrophysiological substrates of impulsiveness: potential effects
on aggressive behavior
Rebecca J. Houstona,*, Matthew S. Stanfordb
aResearch Institute on Addictions, State University of New York at Buffalo, 1021 Main Street, Buffalo, NY 14203, United StatesbDepartment of Psychology and Neuroscience, Baylor University, Waco, TX 76798, United States
Accepted 19 November 2004
Available online 28 December 2004
Abstract
Previous investigations attempting to examine impulsiveness as a personality construct are likely confounded with a high incidence of
aggressive and antisocial behavior. The present study assessed electroencephalographic activity at rest and during photic stimulation in two
groups: (1) an impulsive group (n=10) scoring high on the Barratt Impulsiveness Scale (BIS-11) and reporting no indication of impulsive
aggressive behavior; and (2) a non-aggressive control group (n=14) scoring within the normal range on the BIS-11. All subjects completed a
brief battery of personality measures related to impulsivity and aggression. Resting EEG was recorded at 9 electrode sites. Photic stimulation
was administered at three frequency levels. The primary findings were consistently lower frontal delta and theta activity in the impulsive
group as well as a different topographical pattern of beta activity between the groups. These differences appeared to be independent of photic
stimulation. Personality analyses indicated significantly greater hostility and lifetime history of aggression in the impulsive group. Taken
together, the personality and EEG results suggest some similarity between the present impulsive group and research on groups regularly
exhibiting premeditated aggression. These results provide unique insight into the construct of impulsivity and its role in the expression of
specific subtypes of aggressive behavior.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Aggression; EEG; Impulsivity
1. Introduction
Impulsivity is generally considered a tendency to respond
quickly to a given stimulus, without deliberation and
evaluation of consequences (Buss and Plomin, 1975).
Moeller et al. (2001) have described it as ba predisposition
toward rapid, unplanned reactions to internal or external
stimuli without regard to the negative consequences of these
reactions to themselves or others.Q As impulsiveness is often
0278-5846/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.pnpbp.2004.11.016
Abbreviations: BIS-11, Barratt Impulsiveness Scale; BDHI, Buss-
Durkee Hostility Inventory; BPAQ, Buss-Perry Aggression Questionnaire;
EEG, electroencephalogram; I7, Impulsiveness Questionnaire; LHAQ,
Lifetime History of Aggression Questionnaire; PSW, positive slow wave;
VEP, visual evoked potential.
* Corresponding author. Tel.: +1 716 887 2579; fax: +1 716 887 2477.
E-mail address: [email protected] (R.J. Houston).
considered a trait personality feature, there have been
numerous attempts to explain and understand this construct
from a biological perspective (Barratt, 1985; Eysenck, 1981;
Gray, 1987). One of the most popular theories regarding
impulsivity is the notion of physiological underarousal
(Barratt, 1985; Eysenck, 1967, 1981; Eysenck and Eysenck,
1985; Gray, 1987; Raine, 1993). This is related to Hebb’s
(1955) concept of an boptimal level of arousal.Q The
hypothesis states that arousal level and sensory intake are
related in such a way that an optimal level of arousal is
maintained. When the arousal level falls below the
optimum, stimulation seeking behavior increases and serves
to raise arousal back up to the optimum. Thus, at rest,
impulsive individuals function at a lower level of physio-
logical arousal, which, according to Hebb (1955), is a
subjectively unpleasant state. As a result, these individuals
iological Psychiatry 29 (2005) 305–313

R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313306
engage in impulsive behavior in order to raise that arousal to
a more optimal level (Barratt, 1985). The notion of
physiological underarousal has also been applied to other
samples often characterized by impulsiveness, such as
aggressive, antisocial, and psychopathic individuals (Convit
et al., 1991; Fishbein et al., 1989; Raine, 1993).
Several studies have demonstrated greater EEG slowing
in individuals scoring high on self-report measures of
impulsivity (Barratt and Patton, 1983; O’Gorman and
Lloyd, 1987; Stenberg, 1992), and most have interpreted
these findings as evidence of low resting cortical arousal in
impulsive individuals (Barratt, 1985). In addition, event-
related potential studies demonstrating reduced P3 ampli-
tudes in impulsive aggressive college student (Gerstle et al.,
1998; Mathias and Stanford, 1999) and prisoner (Barratt et
al., 1997) samples have also contributed to the notion of
physiological underarousal related to impulsivity. However,
it should be noted that studies of individuals characterized
by chronic premeditated aggression, who also score high on
self-report measures of impulsivity, have indicated no
significant differences in P3 amplitude (Barratt et al.,
1997; Stanford et al., 2003). Thus, there appears to be
some discrepancy as to what may be driving the neuro-
physiological findings with regard to impulsivity. Finally,
the augmenting–reducing paradigm, an evoked potential
indication of cortical reactivity to increasing stimulus
intensity, has repeatedly demonstrated significant augment-
ing in impulsive (Barratt et al., 1987; Carrillo de la Pena and
Barratt, 1993), sensation seeking (Zuckerman, 1991),
extraverted (Stenberg et al., 1990), and impulsive aggressive
(Houston and Stanford, 2001) subjects. These results have
been interpreted within the stimulation seeking framework,
but may also suggest greater physiological reactivity in
impulsive subjects.
Studies using autonomic measures have also suggested
low resting arousal in various samples characterized by high
levels of impulsivity including antisocial and delinquent
youth (Farrington, 1997; Gatzke-Kopp et al., 2002),
aggressive (Scarpa and Raine, 1997) and psychopathic
individuals (Hare, 1978; Lykken, 1995), and behavior
disordered children (Lahey et al., 1995; Raine and Jones,
1987). However, a recent study examining heart rate in
college students recruited solely on their self-reported
impulsivity status also demonstrated low resting heart rate
in subjects scoring high in impulsiveness (Mathias and
Stanford, 2003). This investigation also reported greater
initial reactivity upon stimulation in high impulsive subjects
during an arousal challenge task. Consequently there
appears to be evidence from both electrocortical and
autonomic indices supporting the notions of low resting
arousal and greater physiological reactivity in impulsive
individuals.
As noted, physiological underarousal is a popular theory
for explaining the expression of aggressive and violent
behavior as well (Convit et al., 1991; Drake et al., 1992;
Fishbein et al., 1989; Gatzke-Kopp et al., 2001; Houston
and Stanford, 2001). This is not surprising as impulsivity
and aggression are behavioral constructs that are difficult to
separate. As a result, many of the aforementioned con-
clusions on impulsivity were drawn from studies using
samples in which high levels of aggressive behavior were
anticipated. Conversely, the few psychophysiological stud-
ies that have examined subjects based on impulsiveness did
not account for potential mediating effects of aggression or
related factors (Barratt and Patton, 1983; O’Gorman and
Lloyd, 1987; Mathias and Stanford, 2003; Stenberg, 1992).
The current study presents a different approach in the
psychophysiological assessment of impulsivity. In order to
address the issues of physiological underarousal and
reactivity associated with impulsiveness and how it might
relate to aggressive behavior, the authors examined electro-
encephalographic activity at rest and during a photic
stimulation task in subjects scoring high on a self-report
measure of impulsivity (BIS-11; Patton et al., 1995) and
reporting no indication of impulsive aggressive behavior.
Because neurobiological deficits have been repeatedly
demonstrated in impulsive aggressive samples, the authors
chose to specifically exclude this subtype of aggressive
behavior rather than aggression in general. This approach
may be useful in attempting to separate impulsivity as a
personality construct from a specific form of aggressive
behavior associated with a lack of behavioral control.
Furthermore, the criteria for chronic impulsive aggressive
behavior have been validated in several previous studies in
prisoner, community, and college student samples, whereas
determining cutoffs for generalized aggression would be
somewhat arbitrary.
2. Methods
2.1. Subjects
Subjects (n=24) were recruited from undergraduate
courses at a mid-sized southern university as part of a
multiple-study screening process. A two page questionnaire
containing the Barratt Impulsiveness Scale (BIS-11; Patton
et al., 1995), a set of questions concerning impulsive
aggressive criteria (e.g. over the past 6 months, have you
had episodes where you were enraged or angry in such a
way that you felt it was excessive or inappropriate to the
situation; Houston and Stanford, 2001 for details on
impulsive aggression criteria in this type of sample) and
other demographic information was distributed in these
courses.
The criteria for inclusion in the impulsive group were the
following: (i) a total score of z74 on the BIS-11 (greater
than 1 S.D. above published norms; Patton et al., 1995) and
(ii) negative responses to all impulsive aggression criteria,
which included 0 outbursts in the last 6 months and scoring
b8 on the Irritability subscale of the Buss–Durkee Hostility
Inventory (BDHI; Buss and Durkee, 1957). Normal control

R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313 307
subjects were drawn from those subjects whose BIS-11
scores were within 1 S.D. above or below the standardized
mean and also responded negatively to all impulsive
aggression criteria (including a score of b8 on the BDHI
Irritability subscale). In addition, all recruited subjects were
right-handed.
Of the 2118 students who completed the screening
questionnaire, 57 met criteria for the impulsive group while
161 were classified as potential control subjects. Subjects
who met some but not all criteria or did not provide
adequate contact information were removed from further
use in this investigation. Subjects who met criteria were
solicited via telephone to participate in the remainder of the
study. Potential subjects were asked a few brief questions
over the phone in order to exclude individuals that were
taking psychoactive medication or had a history of head
injury or seizure. Of the 57 impulsive subjects, 12 refused
participation and 21 reported significant head injury, seizure
activity, or regular psychoactive medication. We were
unsuccessful in contacting 5 of the potential subjects. As a
result, 19 subjects agreed to participate. Seven subjects did
not show up for their appointments and subsequent attempts
to reschedule were unsuccessful.
Participating subjects underwent approximately 2 h of
personality and psychophysiological assessment. Upon
arrival, each subject was asked to read and sign a consent
form. Then each subject was interviewed about psychiatric,
substance abuse, and medical history. Subjects reporting any
Axis I psychiatric diagnoses or significant neurological/
medical problems were excluded from further participation
(n=2 from the impulsive group). Thus, data were collected
for the following groups: impulsive group (n=10, 4M/6F;
mean age 20.3; S.D. 1.2; mean years of education 13.0, S.D.
0.8) and normal control group (n=14, 9M/5F; mean age
19.4, S.D. 1.0; mean years of education 13.1, S.D. 1.0).
2.2. Personality assessment
Subjects were administered the following personality
instruments:
1) I7 Impulsiveness Questionnaire (I7; Eysenck et al.,
1985); the I7 Impulsiveness Questionnaire is a 54-item
questionnaire assessing impulsivity, venturesomeness,
and empathy.
2) Buss–Perry Aggression Questionnaire (BPAQ; Buss
and Perry, 1992); The Buss–Perry Aggression Ques-
tionnaire is a 29-item instrument consisting of four
subscales: Physical Aggression, Verbal Aggression,
Anger, and Hostility. The subscale scores can also be
summed for a total aggression score.
3) Lifetime History of Aggression Questionnaire (LHAQ;
Coccaro et al., 1997); the Lifetime History of Aggres-
sion Questionnaire is administered in an interview
format and assesses incidence of aggressive behavior
from age 13 through adulthood. The LHAQ is
composed of three subscales: the aggression subscale,
the consequences/antisocial behavior subscale, and the
self directed aggression subscale. The aggression
subscale pertains to incidences of aggression such as
verbal and physical fights, throwing/breaking objects,
and childhood temper tantrums. The consequences/
antisocial subscale pertains to the individual’s history of
disciplinary action at school and work as well as any
arrests, convictions, and illegal activities, regardless of
aggressive content. The self directed aggression sub-
scale pertains to acts of self-mutilation and suicide
attempts.
2.3. Electrophysiological assessment
Each subject was seated in a comfortable chair in a sound
and light attenuated room. The scalp was prepared by
application of rubbing alcohol and a mildly abrasive gel
(NuPrep). An electrocap was fitted onto each subject and
EEG data were collected for 9 electrode sites (F3, Fz, F4,
C3, Cz, C4, P3, Pz, and P4). Electrodes were referenced to
linked ears. An eye blink electrode was affixed below the
left eye to allow for differentiation of artifact and removal of
data collected during eye blinks. Impedance for each
electrode was less than 5 Kohms. Filter bandpass was set
at 0.5 and 35 Hz. Five minutes of resting EEG was recorded
with eyes closed.
Selection of EEG segments for analysis was done by
visual inspection, and segments containing artifact (i.e. eye
movements, eye blinks, etc.) were excluded. The process of
eliminating data was performed blind to subject group
assignment. For the resting EEG condition, the average
length of the combined artifact-free segments was 243.21 s.
Signal amplification was conducted using the NE-3 ampli-
fier and the sampling rate was 128 Hz. Artifact-free
segments were subjected to spectral analysis by a Fast
Fourier Transform. Absolute EEG power was calculated by
collapsing the frequency spectrum into EEG frequency
bands of delta (1–4 Hz), theta (5–7 Hz), alpha (8–12 Hz),
and beta (13–20 Hz) (Davidson et al., 2000).
2.4. Photic stimulation
The subject was instructed to focus on the center of a
photostimulator positioned approximately 25 cm from the
subject’s eyelids. The photic stimulation was a red flicker
with flash intensity of 1.40 kcd/m2. The stimulus frequen-
cies used were 5, 10, and 15 Hz. The stimulus train at each
frequency was applied for 30 s with a 10 s time interval. All
subjects were instructed to remain awake and keep their
eyes closed throughout the testing period. EEG power
during photic stimulation was analyzed using the procedures
described above for the resting EEG condition. Average
lengths of the combined artifact-free segments for each
stimulus frequency were as follows: 5 Hz, 23.48 s; 10 Hz,
23.95 s; 15 Hz, 24.03 s.
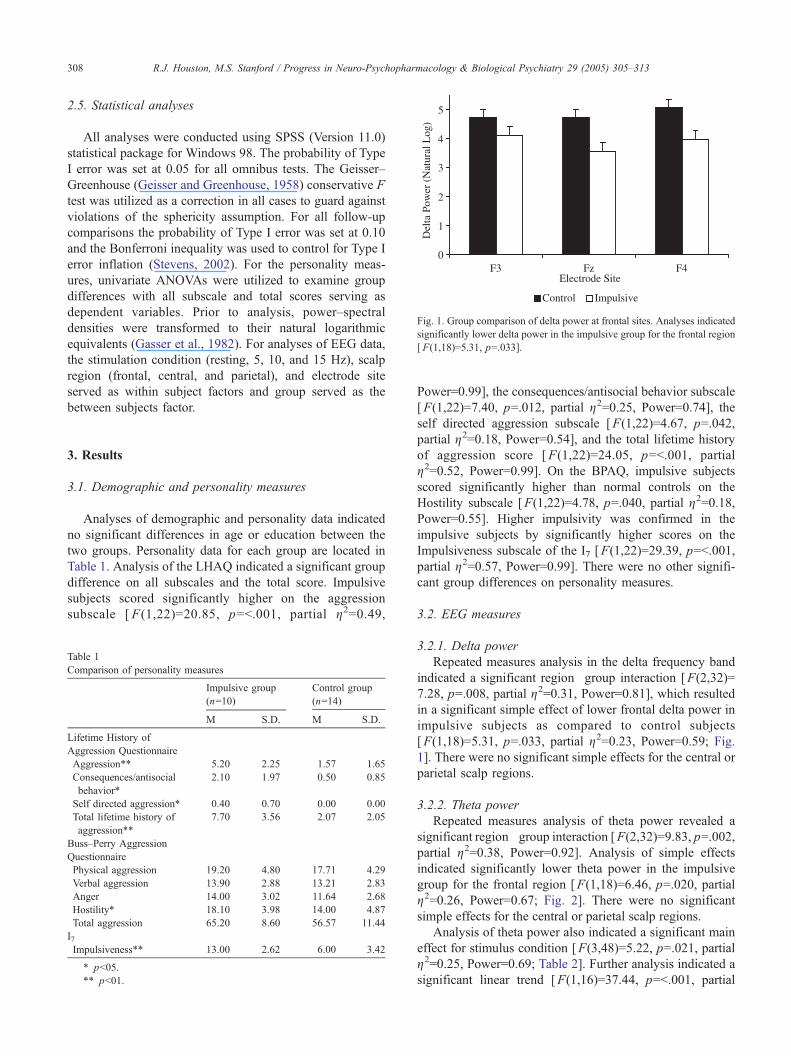
Fig. 1. Group comparison of delta power at frontal sites. Analyses indicated
significantly lower delta power in the impulsive group for the frontal region
[ F(1,18)=5.31, p=.033].
R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313308
2.5. Statistical analyses
All analyses were conducted using SPSS (Version 11.0)
statistical package for Windows 98. The probability of Type
I error was set at 0.05 for all omnibus tests. The Geisser–
Greenhouse (Geisser and Greenhouse, 1958) conservative F
test was utilized as a correction in all cases to guard against
violations of the sphericity assumption. For all follow-up
comparisons the probability of Type I error was set at 0.10
and the Bonferroni inequality was used to control for Type I
error inflation (Stevens, 2002). For the personality meas-
ures, univariate ANOVAs were utilized to examine group
differences with all subscale and total scores serving as
dependent variables. Prior to analysis, power–spectral
densities were transformed to their natural logarithmic
equivalents (Gasser et al., 1982). For analyses of EEG data,
the stimulation condition (resting, 5, 10, and 15 Hz), scalp
region (frontal, central, and parietal), and electrode site
served as within subject factors and group served as the
between subjects factor.
3. Results
3.1. Demographic and personality measures
Analyses of demographic and personality data indicated
no significant differences in age or education between the
two groups. Personality data for each group are located in
Table 1. Analysis of the LHAQ indicated a significant group
difference on all subscales and the total score. Impulsive
subjects scored significantly higher on the aggression
subscale [ F(1,22)=20.85, p=b.001, partial g2=0.49,
Table 1
Comparison of personality measures
Impulsive group
(n=10)
Control group
(n=14)
M S.D. M S.D.
Lifetime History of
Aggression Questionnaire
Aggression** 5.20 2.25 1.57 1.65
Consequences/antisocial
behavior*
2.10 1.97 0.50 0.85
Self directed aggression* 0.40 0.70 0.00 0.00
Total lifetime history of
aggression**
7.70 3.56 2.07 2.05
Buss–Perry Aggression
Questionnaire
Physical aggression 19.20 4.80 17.71 4.29
Verbal aggression 13.90 2.88 13.21 2.83
Anger 14.00 3.02 11.64 2.68
Hostility* 18.10 3.98 14.00 4.87
Total aggression 65.20 8.60 56.57 11.44
I7Impulsiveness** 13.00 2.62 6.00 3.42
* pb05.
** pb01.
Power=0.99], the consequences/antisocial behavior subscale
[F(1,22)=7.40, p=.012, partial g2=0.25, Power=0.74], the
self directed aggression subscale [F(1,22)=4.67, p=.042,
partial g2=0.18, Power=0.54], and the total lifetime history
of aggression score [F(1,22)=24.05, p=b.001, partial
g2=0.52, Power=0.99]. On the BPAQ, impulsive subjects
scored significantly higher than normal controls on the
Hostility subscale [F(1,22)=4.78, p=.040, partial g2=0.18,
Power=0.55]. Higher impulsivity was confirmed in the
impulsive subjects by significantly higher scores on the
Impulsiveness subscale of the I7 [F(1,22)=29.39, p=b.001,
partial g2=0.57, Power=0.99]. There were no other signifi-
cant group differences on personality measures.
3.2. EEG measures
3.2.1. Delta power
Repeated measures analysis in the delta frequency band
indicated a significant region�group interaction [F(2,32)=
7.28, p=.008, partial g2=0.31, Power=0.81], which resulted
in a significant simple effect of lower frontal delta power in
impulsive subjects as compared to control subjects
[F(1,18)=5.31, p=.033, partial g2=0.23, Power=0.59; Fig.1]. There were no significant simple effects for the central or
parietal scalp regions.
3.2.2. Theta power
Repeated measures analysis of theta power revealed a
significant region�group interaction [F(2,32)=9.83, p=.002,
partial g2=0.38, Power=0.92]. Analysis of simple effects
indicated significantly lower theta power in the impulsive
group for the frontal region [F(1,18)=6.46, p=.020, partial
g2=0.26, Power=0.67; Fig. 2]. There were no significant
simple effects for the central or parietal scalp regions.
Analysis of theta power also indicated a significant main
effect for stimulus condition [F(3,48)=5.22, p=.021, partial
g2=0.25, Power=0.69; Table 2]. Further analysis indicated a
significant linear trend [F(1,16)=37.44, p=b.001, partial
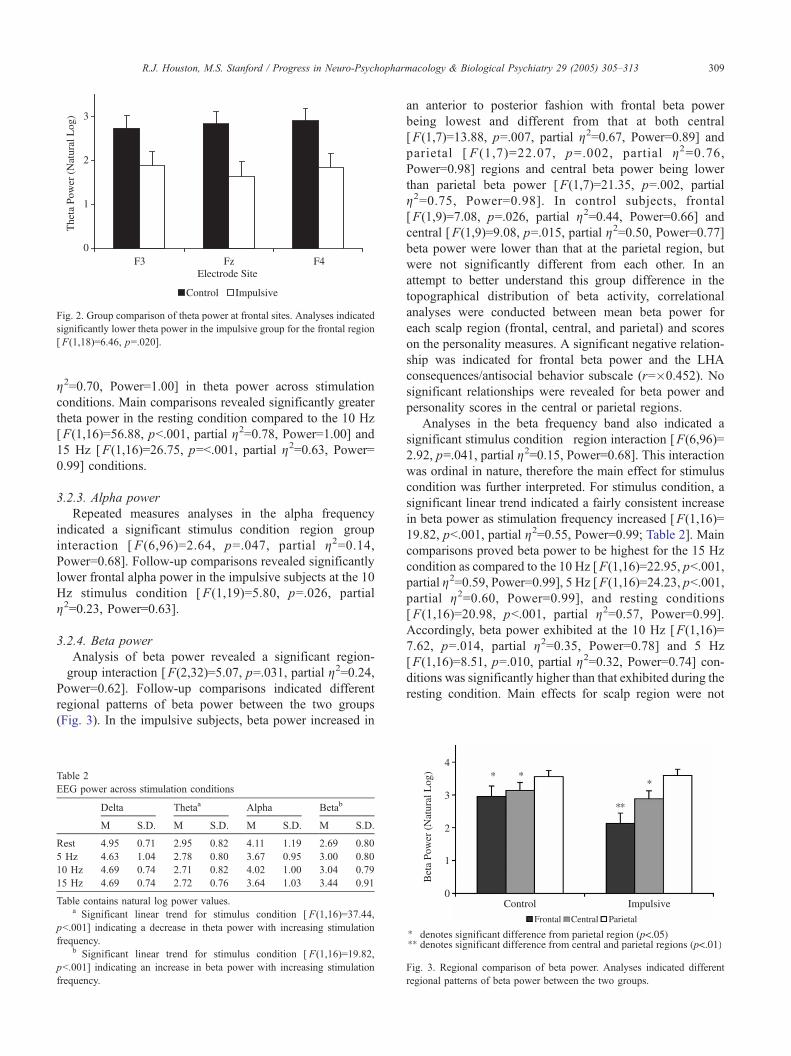
Fig. 2. Group comparison of theta power at frontal sites. Analyses indicated
significantly lower theta power in the impulsive group for the frontal region
[ F(1,18)=6.46, p=.020].
R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313 309
g2=0.70, Power=1.00] in theta power across stimulation
conditions. Main comparisons revealed significantly greater
theta power in the resting condition compared to the 10 Hz
[F(1,16)=56.88, pb.001, partial g2=0.78, Power=1.00] and
15 Hz [F(1,16)=26.75, p=b.001, partial g2=0.63, Power=0.99] conditions.
3.2.3. Alpha power
Repeated measures analyses in the alpha frequency
indicated a significant stimulus condition�region�group
interaction [ F(6,96)=2.64, p=.047, partial g2=0.14,
Power=0.68]. Follow-up comparisons revealed significantly
lower frontal alpha power in the impulsive subjects at the 10
Hz stimulus condition [F(1,19)=5.80, p=.026, partial
g2=0.23, Power=0.63].
3.2.4. Beta power
Analysis of beta power revealed a significant region-�group interaction [F(2,32)=5.07, p=.031, partial g2=0.24,Power=0.62]. Follow-up comparisons indicated different
regional patterns of beta power between the two groups
(Fig. 3). In the impulsive subjects, beta power increased in
Table 2
EEG power across stimulation conditions
Delta Thetaa Alpha Betab
M S.D. M S.D. M S.D. M S.D.
Rest 4.95 0.71 2.95 0.82 4.11 1.19 2.69 0.80
5 Hz 4.63 1.04 2.78 0.80 3.67 0.95 3.00 0.80
10 Hz 4.69 0.74 2.71 0.82 4.02 1.00 3.04 0.79
15 Hz 4.69 0.74 2.72 0.76 3.64 1.03 3.44 0.91
Table contains natural log power values.a Significant linear trend for stimulus condition [ F(1,16)=37.44,
pb.001] indicating a decrease in theta power with increasing stimulation
frequency.b Significant linear trend for stimulus condition [ F(1,16)=19.82,
pb.001] indicating an increase in beta power with increasing stimulation
frequency.
an anterior to posterior fashion with frontal beta power
being lowest and different from that at both central
[F(1,7)=13.88, p=.007, partial g2=0.67, Power=0.89] and
parietal [ F (1,7)=22.07, p=.002, partial g2=0.76,Power=0.98] regions and central beta power being lower
than parietal beta power [F(1,7)=21.35, p=.002, partial
g2=0.75, Power=0.98]. In control subjects, frontal
[F(1,9)=7.08, p=.026, partial g2=0.44, Power=0.66] and
central [F(1,9)=9.08, p=.015, partial g2=0.50, Power=0.77]
beta power were lower than that at the parietal region, but
were not significantly different from each other. In an
attempt to better understand this group difference in the
topographical distribution of beta activity, correlational
analyses were conducted between mean beta power for
each scalp region (frontal, central, and parietal) and scores
on the personality measures. A significant negative relation-
ship was indicated for frontal beta power and the LHA
consequences/antisocial behavior subscale (r=�0.452). No
significant relationships were revealed for beta power and
personality scores in the central or parietal regions.
Analyses in the beta frequency band also indicated a
significant stimulus condition�region interaction [F(6,96)=
2.92, p=.041, partial g2=0.15, Power=0.68]. This interaction
was ordinal in nature, therefore the main effect for stimulus
condition was further interpreted. For stimulus condition, a
significant linear trend indicated a fairly consistent increase
in beta power as stimulation frequency increased [F(1,16)=
19.82, pb.001, partial g2=0.55, Power=0.99; Table 2]. Main
comparisons proved beta power to be highest for the 15 Hz
condition as compared to the 10 Hz [F(1,16)=22.95, pb.001,
partial g2=0.59, Power=0.99], 5 Hz [F(1,16)=24.23, pb.001,partial g2=0.60, Power=0.99], and resting conditions
[F(1,16)=20.98, pb.001, partial g2=0.57, Power=0.99].
Accordingly, beta power exhibited at the 10 Hz [F(1,16)=
7.62, p=.014, partial g2=0.35, Power=0.78] and 5 Hz
[F(1,16)=8.51, p=.010, partial g2=0.32, Power=0.74] con-
ditions was significantly higher than that exhibited during the
resting condition. Main effects for scalp region were not
Fig. 3. Regional comparison of beta power. Analyses indicated different
regional patterns of beta power between the two groups.

R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313310
explored in light of the aforementioned group�region
interaction.
4. Discussion
The present study yielded four primary findings for the
impulsive group: (1) a greater lifetime incidence of
aggressive behavior; (2) higher BPAQ hostility scores; (3)
lower frontal delta and theta activity; and (4) a different
regional pattern of beta activation.
4.1. Impulsivity and aggression
With regard to the greater lifetime history of aggressive
behavior in the impulsive group, it is important to acknowl-
edge that aggressive behavior, in conjunction with high
impulsivity, is common throughout the clinical and empiri-
cal literature regardless of the predominant type of
aggressive behavior (Lee and Coccaro, 2001). Furthermore,
impulsivity is a defining characteristic in several research
groups associated with aggressive behavior that is not
necessarily impulsive or affective in nature such as
premeditated aggression (Barratt et al., 1997; Stanford et
al., 2003) and psychopathy (Cleckley, 1976; Hare, 1993).
As a result, the underlying question is whether impul-
siveness, as a personality trait, differentially influences the
expression of different types of aggressive behavior.
Unfortunately, the data presented at this time cannot provide
an unequivocal answer, but it does allow for some intriguing
suggestions. For example, the impulsive subjects did not
differ from normal controls on most subscales of the BPAQ,
such as physical and verbal aggression, thus corroborating
the ability to control their behavior with respect to
aggression. The higher hostility scores, combined with a
greater lifetime history of aggressive acts, may imply a
personality profile similar to that of the antisocial individual,
who is more likely to commit controlled, premeditated acts
of aggression (Stanford et al., 2003). Interestingly, the
majority of the aggressive acts reported by impulsive
subjects on the LHAQ lacked an affective or reactive
component. This observation is purely anecdotal and cannot
be considered empirical evidence; it is, however, highly
consistent with the greater hostility scores in the impulsive
group.
4.2. Electrocortical findings: interpretation and implications
The findings of lower frontal delta and theta activity in
the impulsive subjects are particularly striking considering
the large body of work demonstrating increased slow wave
activity in impulsive and aggressive samples (Barratt and
Patton, 1983; Convit et al., 1991; Gatzke-Kopp et al., 2001;
O’Gorman and Lloyd, 1987; Stenberg, 1992). Of course,
these previous investigations did not examine the constructs
of impulsivity and impulsive aggression independently.
Historically, decreases in the delta and theta frequency
bands have been associated with greater cortical activation.
Suppressed theta has been repeatedly associated with
hypervigilance and attention (Beatty et al., 1974; Beatty
and O’Hanlon, 1980; Valentino et al., 1993). Decreases in
delta activity in response to nicotine administration have
also been interpreted as evidence of greater cortical activity
and alertness (Knott and Harr, 1996; Pritchard, 1991). Thus,
the present findings could potentially be interpreted as
evidence of greater alertness or attention in this group as
compared to controls. Moreover, group discrepancies in the
frontal region are consistent with a multitude of previous
studies indicating frontal lobe dysfunction in association
with impulsive behavior (Fuster, 1997).
The implication of hypervigilance or enhanced attention
in the current impulsive group is remarkably similar to one
of the more popular theories of physiological function in
psychopath samples (Blackburn, 1979; Raine, 1993). This
research contends that psychopaths exhibit better processing
of relevant stimuli or enhanced attention as indexed by
larger P3 event-related potential and positive slow wave
(PSW) amplitudes (Flor et al., 2002; Raine and Venables,
1988; Raine, 1989, 1993). Given that previous research has
indicated a negative correlation between P3 amplitude and
EEG power in the lower frequency bands (Barratt, 1985),
the present findings could suggest a similarity between our
impulsive group and a psychopath-like sample. As noted,
psychopaths are more likely to commit controlled, premedi-
tated acts of aggression (Williamson et al., 1987; Wood-
worth and Porter, 2002). Thus, it is possible that the current
impulsive group may represent a type of bpremeditated
aggressor,Q possibly similar to a psychopath sample.
In addition, theta rhythms have been linked with various
aspects of cognitive and memory function (Basar, 1998;
Basar et al., 1998). Basar et al. (2001) have concluded that
bspontaneous theta activity of the frontal cortex is a factor
influencing the amplitudes of frontal VEPsQ (p. 198). As
visual event-related potential (VEPs) deficits have been
associated with impulsiveness (Barratt et al., 1987; Carrillo
de la Pena, 1992; Carrillo de la Pena and Barratt, 1993), the
lower frontal theta activity in the current impulsive group
could have important implications in terms of abnormal
cognitive function in these individuals. Future research
might incorporate methodology better designed to inves-
tigate the relationship between these EEG oscillations and
cognitive function in an attempt to further disentangle the
underlying substrates of impulsive and aggressive behavior.
With regard to the group differences in regional beta
activation, correlational analyses indicated a significant
relationship between frontal beta power and the LHA
consequences/antisocial behavior subscale. This subscale
directly assesses behaviors that are often exhibited by
antisocial individuals (i.e. rule violations, arrests) (Coccaro
et al., 1997). Thus, these results are consistent with research
demonstrating less frontal beta reactivity in relation to
antisocial behavior (Finn et al., 2000). Again, this could

R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313 311
provide further support for a similarity between the
impulsive group in the current study and an antisocial or
premeditated profile.
As for the photic stimulation, this task did not
demonstrate greater reactivity in the impulsive group. Given
that EEG power, as measured in the present study, is a
relatively non-specific measure of cortical function, it is
possible that measuring it in response to such stimulation
may be ineffective in determining reactivity differences,
especially in a high functioning sample. Previous studies
using a similar paradigm have demonstrated a diminished
capacity to respond in disorders such as schizophrenia (Jin
et al., 2000) and Alzheimer’s disease (Wada et al., 1998).
Therefore, another influential factor in using this paradigm
may be the necessity of more severe pathology.
4.3. Study limitations
The present results should be interpreted with some
limitations in mind. This study is based on a small sample
size, and despite fairly robust effects, replication of these
results could benefit substantially from a larger sample.
This is particularly true as this approach is different from
previous EEG studies of impulsiveness. The two groups
examined here also differed with respect to gender; the
impulsive group contained a higher ratio of female
subjects. It is unlikely that this significantly impacted the
present results, particularly since recent work has demon-
strated lower theta power in males as compared to females
(Clarke et al., 2001). However, it would be more
appropriate to have gender-matched groups. Furthermore,
although we have interpreted these results as potential
evidence of similarity between the present impulsive group
and antisocial or psychopathic groups, the current study
did not employ any direct measures of psychopathy or
related features aside from impulsivity and aggression.
Continuing work in this area would benefit from a more
extensive battery of personality assessments. For example,
a more extensive examination of psychopathy, criminal
history, and other relevant antisocial behaviors would
prove useful in disentangling these constructs. Finally,
future EEG work in this area should also explore the use of
narrow or individually adjusted frequency bands (Kli-
mesch, 1999) to determine whether differences in cortical
activation in these groups may be restricted to a more
specific frequency.
5. Conclusions
The current findings do raise the question as to the exact
role of impulsivity in the expression of different types of
aggressive behavior. A key element in this distinction is the
notion of behavioral control, particularly in the face of
emotion. Individuals high in impulsivity may be unable to
control certain behavioral impulses, but this phenomenon is
not necessarily related to emotion. Therefore, while some
individuals (i.e. antisocials or premeditated aggressors) are
high in impulsiveness, they can exhibit violent behavior of a
controlled nature, unrelated to their emotional state. In
contrast, impulsive aggressive individuals clearly display a
lack of behavioral control specifically when angered,
agitated or upset. The screening questions utilized in this
study explicitly referred to a loss of behavioral control when
enraged, thus excluding those potential subjects who
endorsed such behavior control problems. As a result, the
significantly higher lifetime history of aggression and
hostility scores in the present impulsive group, coupled
with the suggestion of enhanced attention or vigilance in
frontal areas reinforces the suggestion of similarity between
this group and premeditated or antisocial aggressive groups.
Taken as a whole, these results provide intriguing insight
into the role of impulsivity as a personality construct, its
underlying physiological substrates and its relationship with
specific types of aggressive behavior.
Acknowledgement
This research was supported by the Dreyfus Health
Foundation, The Rogosin Institute, New York Hospital-
Cornell Medical Center.
References
Barratt, E.S., 1985. Impulsiveness subtraits: arousal and information
processing. In: Spence, J.T., Izard, C.E. (Eds.), Motivation, emotion,
and personality. Elsevier Science Publishers, New York, pp. 137–143.
Barratt, E.S., Patton, J.H., 1983. Impulsivity: cognitive, behavioral and
psychophysiological correlates. In: Zuckerman, M. (Ed.), Biological
bases of sensation seeking, impulsivity, and anxiety. Lawrence Ear-
lbaum Associates, Hillsdale, NJ, pp. 77–122.
Barratt, E.S., Pritchard, W.S., Faulk, D.M., Brandt, M.E., 1987. The
relationship between impulsiveness subtraits, trait anxiety, and visual
N100 augmenting/reducing: a topographic analysis. Pers. Individ.
Differ. 8, 43–51.
Barratt, E.S., Stanford, M.S., Kent, T.A., Felthous, A., 1997. Neuro-
psychological and cognitive psychophysiological substrates of impul-
sive aggression. Biol. Psychiatry 41, 1045–1061.
Basar, E., 1998. Brain function and oscillations: integrative brain function,
vol. 2. Springer Verlag, Berlin.
Basar, E., Rahn, E., Demiralp, T., Schqrmann, M., 1998. Spontaneous
EEG theta activity controls frontal visual evoked potential ampli-
tudes. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials 108,
101–109.
Basar, E., Schqrmann, M., Sakowitz, O., 2001. The selectively distributed
theta system: functions. Int. J. Psychophysiol. 39, 197–212.
Beatty, J., O’Hanlon, J., 1980. Operant control of posterior theta rhythm
and vigilance performance: repeated treatments and transfer of training.
In: Birbaumer, N., Kimmel, H. (Eds.), Biofeedback and self-regulation.
Lawrence Erlbaum Associates, Hillsdale, NJ, pp. 247–258.
Beatty, J., Greenberg, A., Deibler, W.P., O’Hanlon, J.F., 1974. Operant
control of occipital theta rhythm affects performance in a radar
monitoring task. Science 183, 871–873.
Blackburn, R., 1979. Cortical and autonomic arousal in primary and
secondary psychopaths. Psychophysiology 16, 143–150.

R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313312
Buss, A.H., Durkee, A., 1957. An inventory for assessing different kinds of
hostility. J. Consult. Psychol. 21, 343–349.
Buss, A.H., Perry, M., 1992. The aggression questionnaire. J. Pers. Soc.
Psychol. 63, 452–459.
Buss, A.H., Plomin, R., 1975. A temperament theory of personality
development. John Wiley & Sons, New York.
Carrillo de la Pena, M.T., 1992. ERP augmenting/reducing and sensation
seeking: a critical review. Int. J. Psychophysiol. 12, 211–220.
Carrillo de la Pena, M.T., Barratt, E.S., 1993. Impulsivity and ERP
augmenting-reducing. Pers. Individ. Differ. 15, 25–32.
Clarke, A.R., Barry, R.J., McCarthy, R., Selikowitz, M., 2001. Age and
sex effects in the EEG: development of the normal child. Clin.
Neurophysiol. 112, 806–814.
Cleckley, H., 1976. The mask of sanity, 5th ed. Mosby, St. Louis, MO.
Coccaro, E.F., Berman, M.E., Kavoussi, R.J., 1997. Assessment of life
history aggression: development and psychometric characteristics.
Psychiatry Res. 73, 147–157.
Convit, A., Czober, P., Volavka, J., 1991. Lateralized abnormality in the
EEG of persistently violent psychiatric inpatients. Biol. Psychiatry 3,
121–138.
Davidson, R.J., Jackson, D.C., Larson, C.L., 2000. Human electro-
encephalography. In: Cacioppo, J.T., Tassinary, L.G., Berntson, G.G.
(Eds.), Handbook of psychophysiology 2nd ed. Cambridge University
Press, New York, pp. 27–52.
Drake, M.E., Hietter, S.A., Pakalnis, A., 1992. EEG and evoked potentials
in episodic-dyscontrol syndrome. Neuropsychobiology 26, 125–128.
Eysenck, H.J., 1967. The biological basis of personality. Charles C.
Thomas, Springfield, IL.
Eysenck, H.J., 1981. General features of the model. In: Eysenck, H.J. (Ed.),
A model for personality. Springer-Verlag, New York.
Eysenck, HJ., Eysenck, M.W., 1985. Personality and individual differences:
a natural science approach. Plenum Press, New York.
Eysenck, S.B.G., Pearson, P.R., Easting, G., Allsopp, J.F., 1985.
Impulsiveness and venturesomeness: their position in a dimensional
system of personality description. Pers. Individ. Differ. 6, 613–619.
Farrington, D.P., 1997. The relationship between low resting heart rate
and violence. In: Raine, A., Brennan, P.A., Farrington, D.P., Mednick,
S.A. (Eds.), Biosocial bases of violence. Plenum Press, New York,
pp. 89–105.
Finn, P.R., Ramsey, S.E., Earleywine, M., 2000. Frontal EEG response to
threat, aggressive traits and a family history of alcoholism: a
preliminary study. J. Stud. Alcohol 61, 38–45.
Fishbein, D.H., Herning, R.I., Pickworth, W.B., Haertzen, C.A., Hickey,
J.E., Jaffe, J.H., 1989. EEG and brainstem auditory evoked response
potentials in adult male drug abusers with self-reported histories of
aggressive behavior. Biol. Psychiatry 26, 595–611.
Flor, H., Birbaumer, N., Hermann, C., Ziegler, S., Patrick, C., 2002.
Aversive Pavlovian conditioning in psychopaths: peripheral and central
correlates. Psychophysiology 39, 505–518.
Fuster, J.M., 1997. The prefrontal cortex: anatomy, physiology and
neuropsychology of the frontal lobe. 3rd ed. Raven, New York.
Gasser, T., Bacher, P., Mocks, J., 1982. Transformation towards normal
distribution of broad spectral parameters of the EEG. Electroencepha-
logr. Clin. Neurophysiol. 53, 119–124.
Gatzke-Kopp, L.M., Raine, A., Buchsbaum, M., LaCasse, L., 2001.
Temporal lobe deficits in murderers: EEG findings undetected by
PET. J. Neuropsychiatry Clin. Neurosci. 13, 486–491.
Gatzke-Kopp, L.M., Raine, A., Loeber, R., Stouthamer-Loeber, M.,
Steinhauer, S., 2002. Serious delinquent behavior, sensation
seeking and electrodermal arousal. J. Abnorm. Child Psychol.
30, 477–486.
Geisser, S., Greenhouse, S.W., 1958. An extension of Box’s results on the
use of the F distribution in multivariate analysis. Ann. Math. Stat. 29,
885–891.
Gerstle, J.E., Mathias, C.W., Stanford, M.S., 1998. Auditory P300 and self-
reported impulsive aggression. Prog. Neuro-Psychopharmacol. Biol.
Psychiatry 22, 575–583.
Gray, J.A., 1987. Perspectives on anxiety and impulsivity: a commentary.
J. Res. Pers. 21, 493–509.
Hare, R.D., 1978. Electrodermal and cardiovascular correlates of psychop-
athy. In: Hare, R.D., Schalling, D. (Eds.), Psychopathic behavior:
approaches to research. Wiley, New York, pp. 107–144.
Hare, R.D., 1993. Without conscience: the disturbing world of psychopaths
among us. Pocket Books, New York.
Hebb, D.O., 1955. Drives and the CNS (conceptual nervous system).
Psychol. Rev. 62, 243–254.
Houston, R.J., Stanford, M.S., 2001. Mid-latency evoked potential in self-
reported impulsive aggression. Int. J. Psychophysiol. 40, 1–15.
Jin, Y., Castellanos Jr., A., Solis Jr., E.R., Potkin, S.G., 2000. EEG resonant
responses in schizophrenia: a photic driving study with improved
harmonic resolution. Schizophr. Res. 44, 213–220.
Klimesch, W., 1999. EEG alpha and theta oscillations reflect cognitive and
memory performance: a review and analysis. Brain Res. Rev. 29,
169–195.
Knott, V.J., Harr, A., 1996. Assessing the topographic EEG changes
associated with aging and acute/long-term effects of smoking. Neuro-
psychobiology 33, 210–222.
Lahey, B.B., McBurnett, K., Loeber, R., Hart, E.L., 1995. Psychobiology.
In: Sholevar, G. (Ed.), Conduct disorders in children and adolescents.
Ameri. Psychiat. Press, Washington, pp. 27–44.
Lee, R., Coccaro, E., 2001. The neuropsychopharmacology of criminality
and aggression. Can. J. Psychiatry 46, 35–44.
Lykken, D., 1995. The antisocial personalities. Lawrence Erlbaum,
Hillsdale, NJ.
Mathias, C.W., Stanford, M.S., 1999. P300 under standard and surprise
conditions in self-reported impulsive aggression. Prog. Neuro-Psycho-
pharmacol. Biol. Psychiatry 23, 1037–1051.
Mathias, C.W., Stanford, M.S., 2003. Impulsiveness and arousal: heart rate
under conditions of rest and challenge in healthy males. Pers. Individ.
Differ. 35, 355–371.
Moeller, F.G., Barratt, E.S., Dougherty, D.M., Schmitz, J.M., Swann,
A.C., 2001. Psychiatric aspects of impulsivity. Am. J. Psychiatr. 158,
1783–1793.
O’Gorman, J.G., Lloyd, J.E.M., 1987. Extraversion, impulsiveness, and
EEG alpha activity. Pers. Individ. Differ. 8, 169–174.
Patton, J.H., Stanford, M.S., Barratt, E.S., 1995. Factor structure of the
Barratt Impulsiveness Scale. J. Clin. Psychol. 51, 768–774.
Pritchard, W.S., 1991. Electroencephalographic effects of cigarette
smoking. Psychopharmacology 104, 485–490.
Raine, A., 1989. Evoked potentials and psychopathy. Int. J. Psychophysiol.
8, 1–16.
Raine, A., 1993. The psychopathology of crime: criminal behavior as a
clinical disorder. Academic Press, San Diego.
Raine, A., Jones, F., 1987. Attention, autonomic arousal and personality
in behaviorally disordered children. J. Abnorm. Child Psychol. 14,
583–599.
Raine, A., Venables, P.H., 1988. Enhanced P3 evoked potentials and
longer P3 recovery times in psychopaths. Psychophysiology 25,
30–38.
Scarpa, A., Raine, A., 1997. Psychophysiology of anger and violent
behavior. Psychiatr. Clin. North Am. 20, 375–394.
Stanford, M.S., Houston, R.J., Villemarette-Pittman, N.R., Greve, K.W.,
2003. Premeditated aggression: clinical assessment and cognitive
psychophysiology. Pers. Individ. Differ. 34, 773–781.
Stenberg, G., 1992. Personality and the EEG: arousal and emotional
arousability. Pers. Individ. Differ. 13, 1097–1113.
Stenberg, G., Rosen, I., Risenberg, J., 1990. Attention and personality in
augmenting/reducing of visual evoked potentials. Pers. Individ. Differ.
11, 1243–1254.
Stevens, J., 2002. Applied multivariate statistics for the social sciences.
Lawrence Erlbaum Assoc, Mahwah, NJ.
Valentino, D.A., Arruda, J.E., Gold, S.M., 1993. Comparison of QEEG and
response accuracy in good vs. poorer performers during a vigilance task.
Int. J. Psychophysiol. 15, 123–134.

R.J. Houston, M.S. Stanford / Progress in Neuro-Psychopharmacology & Biological Psychiatry 29 (2005) 305–313 313
Wada, Y., Nanbu, Y., Kikuchi, M., Koshino, Y., Hashimoto, T., Yamaguchi,
N., 1998. Abnormal functional connectivity in Alzheimer’s disease:
intrahemispheric EEG coherence during rest and photic stimulation.
Eur. Arch. Psychiatry Clin. Neurosci. 248, 203–208.
Williamson, S., Hare, R.D., Wong, S., 1987. Violence: criminal psycho-
paths and their victims. Can. J. Behav. Sci. 19, 454–462.
Woodworth, M., Porter, S., 2002. In cold blood: characteristics of criminal
homicides as a function of psychopathy. J. Abnorm. Psychology 111,
436–445.
Zuckerman, M., 1991. Psychobiology of personality. Cambridge University
Press, New York.





























