Embed Size (px)
Citation preview

REGULAR PAPER
Elucidating the site of action of oxalate in photosynthetic electrontransport chain in spinach thylakoid membranes
Anjana Jajoo Æ Archna Sahay Æ Pooja Singh Æ Sonal Mathur ÆSergei K. Zharmukhamedov Æ Vyacheslav V. Klimov ÆSuleyman I. Allakhverdiev Æ Sudhakar Bharti
Received: 12 March 2008 / Accepted: 19 May 2008
� Springer Science+Business Media B.V. 2008
Abstract The effects of oxalate on PS II and PS I photo-
chemistry were studied. The results suggested that in
chloride-deficient thylakoid membranes, oxalate inhibited
activity of PS II as well as PS I. To our knowledge, this is the
only anion so far known which inhibits both the photosys-
tems. Measurements of fluorescence induction kinetics, YZ•
decay, and S2 state multiline EPR signal suggested that
oxalate inhibited PS II at the donor side most likely on the
oxygen evolving complex. Measurements of re-reduction of
P700+ signal in isolated PS I particles in oxalate-treated
samples suggested a binding site of oxalate on the donor, as
well as the acceptor side of PS I.
Keywords Electron transport chain � EPR �Fluorescence � Oxalate anion � Photosystem II �Photosystem I
Abbreviations
DCPIP 2,6-Dichlorophenolindophenol
DAD(red) Reduced 3,6-diaminodurene
Fo Initial fluorescence, where all QA are oxidized
Fv Variable fluorescence
MV Methyl viologen
TMPD(red) Reduced N,N,N1,
N1-tetramethyl-p-phenylenediamine
Introduction
Transformation of light energy into chemical energy in the
course of oxygenic photosynthesis takes place with the
contribution of sequentially functioning photosystem II (PS
II) and photosystem I (PS I), the thylakoid membrane
complexes containing reaction center and the core antenna.
PS II is a light-dependent water-plastoquinone oxidore-
ductase enzyme that uses light energy to oxidize water and
is mainly located in the appressed grana stacks. Various
crystallographic investigations of cyanobacterial photo-
system II have provided high resolution structures from 3.8
to 3.0 A (Kamiya and Shen 2003; Ferreira et al. 2004; Loll
et al. 2005) that explain the general arrangement of the
protein matrix and cofactors. The structural and functional
aspects of PS II are interrelated. PS I functions as a light-
driven plastocyanin-Fd oxidoreductase (Chitnis et al.
1995). The structure of the PS I core complex from the
thermophilic cyanobacterium Thermosynechococcus elon-
gates is known at 2.5 A resolution (Jordan et al. 2001).
A large number of organic and inorganic anions affect
various reactions associated with PS II and PS I. Certain
organic anions like glyoxylate, oxalate, and glycolate com-
pete with anions like formate (HCO2-) and bicarbonate
(HCO3-) for binding to the PS II (Petrouleas et al. 1994). It is
well established that bicarbonate is required for the func-
tional activity at the acceptor side of PS II, providing efficient
re-oxidation of the QA. The non-heme Fe between QA and QB
has been shown to play an essential role in bicarbonate
binding (Govindjee and Van Rensen 1993). On the other
hand, bicarbonate requirement for the donor side of PS II has
been clearly demonstrated (Klimov et al. 1995, 1997; All-
akhverdiev et al. 1997). Formate slows down the electron
transfer from QA- to QB and competes with HCO3
- for the
binding to the non-heme iron at the acceptor side of PS II
A. Jajoo (&) � A. Sahay � P. Singh � S. Mathur � S. Bharti
School of Life Sciences, Devi Ahilya University, Vigyan
Bhavan, Khandwa Road, Indore 452017, M.P., India
e-mail: [email protected]
A. Jajoo � S. K. Zharmukhamedov � V. V. Klimov �S. I. Allakhverdiev � S. Bharti
Institute of Basic Biological Problems, Russian Academy
of Sciences, Pushchino, Moscow Region 142290, Russia
123
Photosynth Res
DOI 10.1007/s11120-008-9314-1

(Petrouleas and Diner 1990). Several studies have shown that
HCO2- induces inhibition of water oxidation reactions, as
well as electron transfer on the PS II acceptor side (Stemler
and Lavergne 1997). Interaction of HCO2- with chloride
(Cl-) on the donor and acceptor sides of PS II has been
reported (Feyziev et al. 2000; Jajoo et al. 2005a).
Inorganic anion Cl- has been shown to be an indis-
pensable cofactor involved in the photosynthetic oxygen
evolution (Lindberg and Andreasson 1996). Chloride is
required for high oxygen evolution activity in PS II and is
responsible for the normal electron paramagnetic reso-
nance (EPR) properties of the S2 state (Wincencjusz et al.
1999). Activating Cl- can be replaced by several mono-
valent anions including bromide (Br-), nitrate (NO3-),
iodide (I-) and nitrite (NO2-) with varying efficiency. In
Cl--depleted PS II membranes, substitution of Cl- by Br-,
I-, NO2-, and fluoride (F-) suggests a possible binding site
of these anions at the PS II donor side as well as at the
acceptor side (Jajoo et al. 2005b).
Different functions have been proposed for oxalate in
plants, including Ca2+ regulation, ion balance (e.g., Na+
and K+), plant protection, tissue support and heavy metal
detoxification (Nakata 2003). More attention and interest
has been focused on the roles of oxalate in plant abiotic
stress resistance, such as heavy metal toxicity and phos-
phorus deficiency (Dong et al. 2004). In view of its
function as a strong chelator of manganese and other cat-
ions (Schlosser and Hofer 2002), it is necessary to examine
the role of oxalate in photosynthetic reactions also. In the
photosynthetic electron transport chain among many
anions so far studied only oxalate was found to inhibit both
the photosystems (Jajoo and Bharti 1993a). It was, there-
fore, of interest to determine the specific effect and site of
action of oxalate in the photosynthetic electron transport
chain. In this study using oxalate-treated thylakoids, we
present data on: (i) electron transfer rates through PS II; (ii)
chl a fluorescence induction kinetics at room temperature;
(iii) decay kinetics of YZ• by time-resolved EPR at 253 K;
(iv) S2 state multiline EPR spectra; (v) partial reactions
mediated by PS I in the absence and presence of HgCl2;
(vi) re-reduction of P700+ in purified PSI particles. Based
on our results, we report for the first time that oxalate
inhibited PS II at the donor side on the oxygen evolving
complex (OEC) and inhibited PS I by binding on the donor
as well as the acceptor side of PS I.
Materials and methods
Isolation and storage
PS II membranes and thylakoid membranes were prepared
from fresh market spinach following method as described
in Kuwabara and Murata (1982). The membranes were
stored at 77 K with 50% glycerol added until use. Prior to
the experiment, the membranes were thawed slowly at 0�C
and washed with suspension medium containing 0.1 M
sucrose and 50 mM HEPES–NaOH buffer (pH 7.6) in
order to remove glycerol and centrifuged at 4,500g for
10 min. For preparation of ion-deficient membranes (-ion)
which did not contain any exogenous Cl-, the pellet was
suspended in an isolation medium containing 0.33 M
sucrose, 50 mM HEPES–NaOH buffer (pH 7.6). For
preparation of ion-sufficient membranes (+ion) isolation
medium consisted of 0.33 M sucrose, 50 mM HEPES–
NaOH buffer (pH 7.6), 10 mM NaCl and 1 mM MgCl2. All
steps were performed at 0–4�C. The thylakoid membranes
were stored in dark on ice. The chlorophyll content was
measured according to the method as described in Porra
et al. (1989). It is to be emphasized that we did not use
chloride-depleted membranes that require harsher treat-
ments, e.g., high pH, treatment with sulfate to remove
endogenous Cl-.
Purified PS I particles were prepared following the
method of Shuvalov et al. (1976).
Measurements of rates of electron transfer through PS
II and PS I
The activity of PS II was measured spectrophotometrically
as photo-reduction of DCPIP (H2O ? DCPIP reaction).
White light was employed for illuminating the reaction
mixture for 30 s and its absorbance was measured at
605 nm. The PS II activity was expressed in terms of lmol
DCPIP reduced mg chl-1 h-1.
The activity of PS I was monitored polarographically by
YS I model Clark-type oxygen electrode (Yellow Springs,
USA) with a water-jacketed reaction vessel thermostated at
25�C connected to a graphic recorder. The activity of PS I
was measured with either DCPIPH2, TMPD(red) or DAD(red)
as electron donor and methyl viologen (MV) as terminal
auto-oxidizable electron acceptor. The reaction mixture
was constantly stirred with the help of a magnetic stirrer.
Light intensity (150 W m-2) was employed for illuminat-
ing the reaction mixture. The PS I activity was expressed in
terms of lmol oxygen consumed mg chl-1 h-1.
Fluorescence measurements
Fluorescence induction kinetics of chl a was monitored
using a Photosynthetic Efficiency Analyzer (PEA, Hansa-
tech, UK). Re-reduction of P700+ in purified PS I particles
were measured in a 1 cm cuvette using a homemade
phosphoroscopic set-up as described in Klimov et al.
(1982).
Photosynth Res
123
EPR measurements
In order to measure rate of decay of YZ• , time-resolved EPR
measurements were performed using Varian X-band
spectrometer and homemade nitrogen gas-flow cryostat
with a temperature controller. Samples were directly illu-
minated using HOYA-SCHOTT MegaLight-100 to excite
YZ• signals. To measure S2 state multiline EPR signal, CW-
EPR measurements were performed using a Bruker-300E
X-band spectrometer, and an ST4102 standard cavity. An
Oxford-900 continuous gas-flow cryostat and ITC-4 tem-
perature controller were used to regulate the sample
temperature at 6.0 K. Samples (pH 6.8) were illuminated
with 500 mW tungsten-halogen lamp through an 8 cm
thick water filter to induce the formation of the S2 state
multiline signal at 200 K in an ethanol/solid CO2 bath. The
illuminated samples were quickly cooled to 200 K and then
stored at 77 K. The EPR spectra of S2 state multiline sig-
nals were recorded at 6 K. Chlorophyll concentration used
in all EPR measurements was 3–4 mg/ml.
Results and discussion
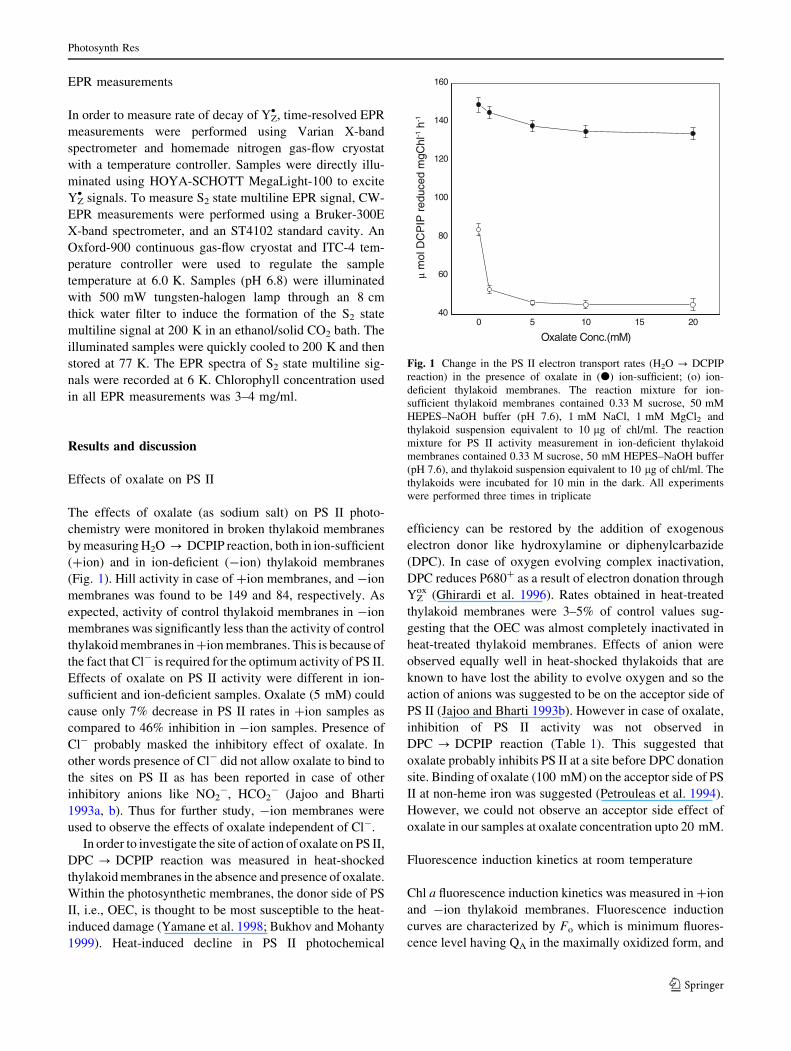
Effects of oxalate on PS II
The effects of oxalate (as sodium salt) on PS II photo-
chemistry were monitored in broken thylakoid membranes
by measuring H2O ? DCPIP reaction, both in ion-sufficient
(+ion) and in ion-deficient (-ion) thylakoid membranes
(Fig. 1). Hill activity in case of +ion membranes, and -ion
membranes was found to be 149 and 84, respectively. As
expected, activity of control thylakoid membranes in -ion
membranes was significantly less than the activity of control
thylakoid membranes in +ion membranes. This is because of
the fact that Cl- is required for the optimum activity of PS II.
Effects of oxalate on PS II activity were different in ion-
sufficient and ion-deficient samples. Oxalate (5 mM) could
cause only 7% decrease in PS II rates in +ion samples as
compared to 46% inhibition in -ion samples. Presence of
Cl- probably masked the inhibitory effect of oxalate. In
other words presence of Cl- did not allow oxalate to bind to
the sites on PS II as has been reported in case of other
inhibitory anions like NO2-, HCO2
- (Jajoo and Bharti
1993a, b). Thus for further study, -ion membranes were
used to observe the effects of oxalate independent of Cl-.
In order to investigate the site of action of oxalate on PS II,
DPC ? DCPIP reaction was measured in heat-shocked
thylakoid membranes in the absence and presence of oxalate.
Within the photosynthetic membranes, the donor side of PS
II, i.e., OEC, is thought to be most susceptible to the heat-
induced damage (Yamane et al. 1998; Bukhov and Mohanty
1999). Heat-induced decline in PS II photochemical
efficiency can be restored by the addition of exogenous
electron donor like hydroxylamine or diphenylcarbazide
(DPC). In case of oxygen evolving complex inactivation,
DPC reduces P680+ as a result of electron donation through
YZox (Ghirardi et al. 1996). Rates obtained in heat-treated
thylakoid membranes were 3–5% of control values sug-
gesting that the OEC was almost completely inactivated in
heat-treated thylakoid membranes. Effects of anion were
observed equally well in heat-shocked thylakoids that are
known to have lost the ability to evolve oxygen and so the
action of anions was suggested to be on the acceptor side of
PS II (Jajoo and Bharti 1993b). However in case of oxalate,
inhibition of PS II activity was not observed in
DPC ? DCPIP reaction (Table 1). This suggested that
oxalate probably inhibits PS II at a site before DPC donation
site. Binding of oxalate (100 mM) on the acceptor side of PS
II at non-heme iron was suggested (Petrouleas et al. 1994).
However, we could not observe an acceptor side effect of
oxalate in our samples at oxalate concentration upto 20 mM.
Fluorescence induction kinetics at room temperature
Chl a fluorescence induction kinetics was measured in +ion
and -ion thylakoid membranes. Fluorescence induction
curves are characterized by Fo which is minimum fluores-
cence level having QA in the maximally oxidized form, and
0 5 10 15 2040
60
80
100
120
140
160
µ m
ol D
CP
IP r
educ
ed m
gChl
-1 h
-1
Oxalate Conc.(mM)
Fig. 1 Change in the PS II electron transport rates (H2O ? DCPIP
reaction) in the presence of oxalate in (d) ion-sufficient; (o) ion-
deficient thylakoid membranes. The reaction mixture for ion-
sufficient thylakoid membranes contained 0.33 M sucrose, 50 mM
HEPES–NaOH buffer (pH 7.6), 1 mM NaCl, 1 mM MgCl2 and
thylakoid suspension equivalent to 10 lg of chl/ml. The reaction
mixture for PS II activity measurement in ion-deficient thylakoid
membranes contained 0.33 M sucrose, 50 mM HEPES–NaOH buffer
(pH 7.6), and thylakoid suspension equivalent to 10 lg of chl/ml. The
thylakoids were incubated for 10 min in the dark. All experiments
were performed three times in triplicate
Photosynth Res
123
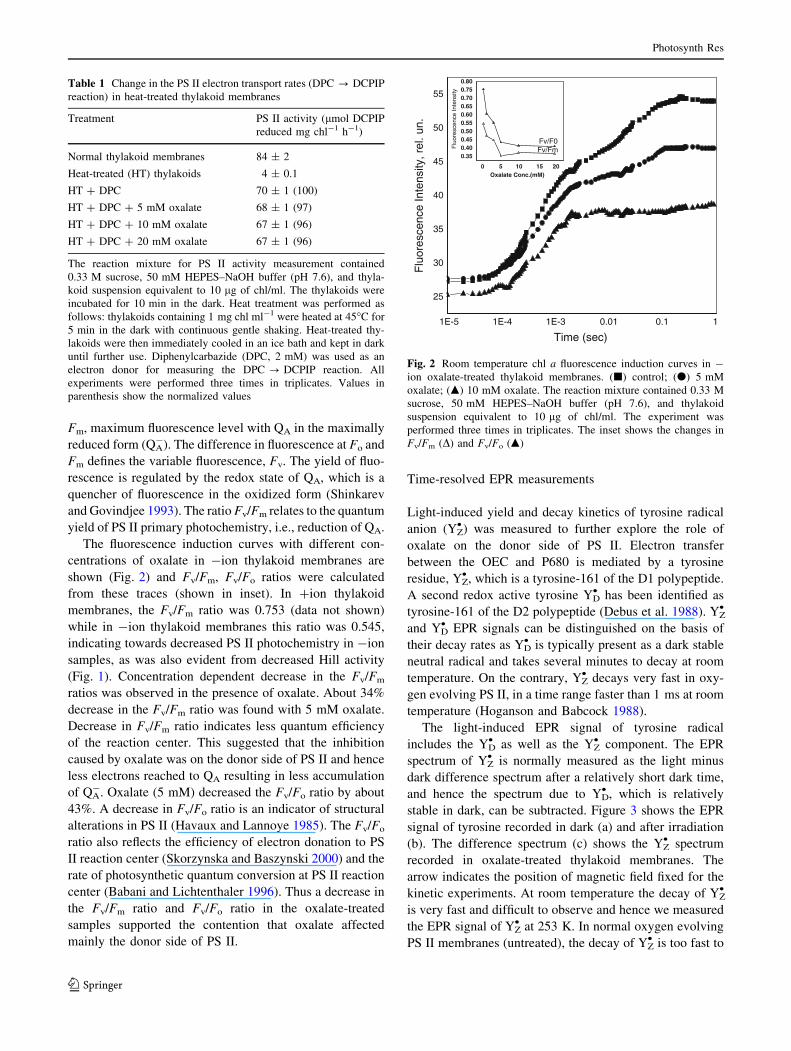
Fm, maximum fluorescence level with QA in the maximally
reduced form (QA-). The difference in fluorescence at Fo and
Fm defines the variable fluorescence, Fv. The yield of fluo-
rescence is regulated by the redox state of QA, which is a
quencher of fluorescence in the oxidized form (Shinkarev
and Govindjee 1993). The ratio Fv/Fm relates to the quantum
yield of PS II primary photochemistry, i.e., reduction of QA.
The fluorescence induction curves with different con-
centrations of oxalate in -ion thylakoid membranes are
shown (Fig. 2) and Fv/Fm, Fv/Fo ratios were calculated
from these traces (shown in inset). In +ion thylakoid
membranes, the Fv/Fm ratio was 0.753 (data not shown)
while in -ion thylakoid membranes this ratio was 0.545,
indicating towards decreased PS II photochemistry in -ion
samples, as was also evident from decreased Hill activity
(Fig. 1). Concentration dependent decrease in the Fv/Fm
ratios was observed in the presence of oxalate. About 34%
decrease in the Fv/Fm ratio was found with 5 mM oxalate.
Decrease in Fv/Fm ratio indicates less quantum efficiency
of the reaction center. This suggested that the inhibition
caused by oxalate was on the donor side of PS II and hence
less electrons reached to QA resulting in less accumulation
of QA-. Oxalate (5 mM) decreased the Fv/Fo ratio by about
43%. A decrease in Fv/Fo ratio is an indicator of structural
alterations in PS II (Havaux and Lannoye 1985). The Fv/Fo
ratio also reflects the efficiency of electron donation to PS
II reaction center (Skorzynska and Baszynski 2000) and the
rate of photosynthetic quantum conversion at PS II reaction
center (Babani and Lichtenthaler 1996). Thus a decrease in
the Fv/Fm ratio and Fv/Fo ratio in the oxalate-treated
samples supported the contention that oxalate affected
mainly the donor side of PS II.
Time-resolved EPR measurements
Light-induced yield and decay kinetics of tyrosine radical
anion (YZ• ) was measured to further explore the role of
oxalate on the donor side of PS II. Electron transfer
between the OEC and P680 is mediated by a tyrosine
residue, YZ• , which is a tyrosine-161 of the D1 polypeptide.
A second redox active tyrosine YD• has been identified as
tyrosine-161 of the D2 polypeptide (Debus et al. 1988). YZ•
and YD• EPR signals can be distinguished on the basis of
their decay rates as YD• is typically present as a dark stable
neutral radical and takes several minutes to decay at room
temperature. On the contrary, YZ• decays very fast in oxy-
gen evolving PS II, in a time range faster than 1 ms at room
temperature (Hoganson and Babcock 1988).
The light-induced EPR signal of tyrosine radical
includes the YD• as well as the YZ
• component. The EPR
spectrum of YZ• is normally measured as the light minus
dark difference spectrum after a relatively short dark time,
and hence the spectrum due to YD• , which is relatively
stable in dark, can be subtracted. Figure 3 shows the EPR
signal of tyrosine recorded in dark (a) and after irradiation
(b). The difference spectrum (c) shows the YZ• spectrum
recorded in oxalate-treated thylakoid membranes. The
arrow indicates the position of magnetic field fixed for the
kinetic experiments. At room temperature the decay of YZ•
is very fast and difficult to observe and hence we measured
the EPR signal of YZ• at 253 K. In normal oxygen evolving
PS II membranes (untreated), the decay of YZ• is too fast to
Table 1 Change in the PS II electron transport rates (DPC ? DCPIP
reaction) in heat-treated thylakoid membranes
Treatment PS II activity (lmol DCPIP
reduced mg chl-1 h-1)
Normal thylakoid membranes 84 ± 2
Heat-treated (HT) thylakoids 4 ± 0.1
HT + DPC 70 ± 1 (100)
HT + DPC + 5 mM oxalate 68 ± 1 (97)
HT + DPC + 10 mM oxalate 67 ± 1 (96)
HT + DPC + 20 mM oxalate 67 ± 1 (96)
The reaction mixture for PS II activity measurement contained
0.33 M sucrose, 50 mM HEPES–NaOH buffer (pH 7.6), and thyla-
koid suspension equivalent to 10 lg of chl/ml. The thylakoids were
incubated for 10 min in the dark. Heat treatment was performed as
follows: thylakoids containing 1 mg chl ml-1 were heated at 45�C for
5 min in the dark with continuous gentle shaking. Heat-treated thy-
lakoids were then immediately cooled in an ice bath and kept in dark
until further use. Diphenylcarbazide (DPC, 2 mM) was used as an
electron donor for measuring the DPC ? DCPIP reaction. All
experiments were performed three times in triplicates. Values in
parenthesis show the normalized values
25
30
35
40
45
50
55
Flu
ores
cenc
e In
tens
ity, r
el. u
n.
Time (sec)
0.350.400.450.500.550.600.650.700.750.80
Flu
ores
cenc
e In
tens
ity
Oxalate Conc.(mM)
Fv/FmFv/F0
50 10 15 20
1E-5 1E-4 1E-3 0.01 0.1 1
Fig. 2 Room temperature chl a fluorescence induction curves in -
ion oxalate-treated thylakoid membranes. (j) control; (d) 5 mM
oxalate; (m) 10 mM oxalate. The reaction mixture contained 0.33 M
sucrose, 50 mM HEPES–NaOH buffer (pH 7.6), and thylakoid
suspension equivalent to 10 lg of chl/ml. The experiment was
performed three times in triplicates. The inset shows the changes in
Fv/Fm (D) and Fv/Fo (m)
Photosynth Res
123
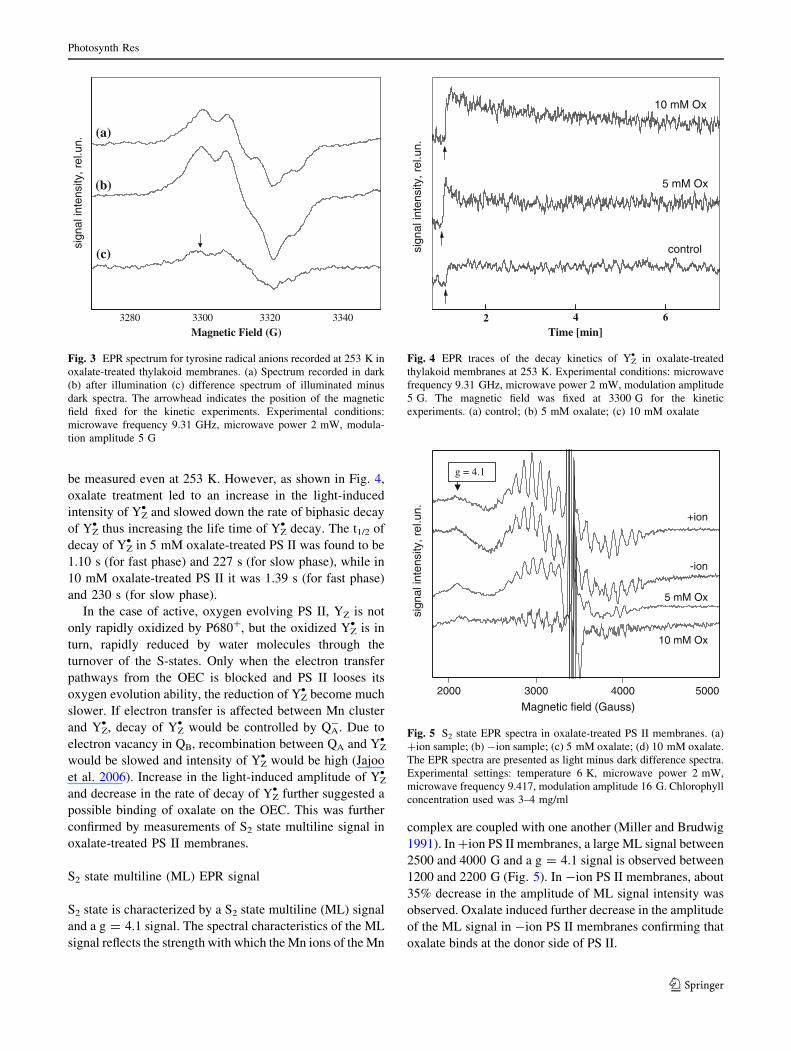
be measured even at 253 K. However, as shown in Fig. 4,
oxalate treatment led to an increase in the light-induced
intensity of YZ• and slowed down the rate of biphasic decay
of YZ• thus increasing the life time of YZ
• decay. The t1/2 of
decay of YZ• in 5 mM oxalate-treated PS II was found to be
1.10 s (for fast phase) and 227 s (for slow phase), while in
10 mM oxalate-treated PS II it was 1.39 s (for fast phase)
and 230 s (for slow phase).
In the case of active, oxygen evolving PS II, YZ is not
only rapidly oxidized by P680+, but the oxidized YZ• is in
turn, rapidly reduced by water molecules through the
turnover of the S-states. Only when the electron transfer
pathways from the OEC is blocked and PS II looses its
oxygen evolution ability, the reduction of YZ• become much
slower. If electron transfer is affected between Mn cluster
and YZ• , decay of YZ
• would be controlled by QA-. Due to
electron vacancy in QB, recombination between QA and YZ•
would be slowed and intensity of YZ• would be high (Jajoo
et al. 2006). Increase in the light-induced amplitude of YZ•
and decrease in the rate of decay of YZ• further suggested a
possible binding of oxalate on the OEC. This was further
confirmed by measurements of S2 state multiline signal in
oxalate-treated PS II membranes.
S2 state multiline (ML) EPR signal
S2 state is characterized by a S2 state multiline (ML) signal
and a g = 4.1 signal. The spectral characteristics of the ML
signal reflects the strength with which the Mn ions of the Mn
complex are coupled with one another (Miller and Brudwig
1991). In +ion PS II membranes, a large ML signal between
2500 and 4000 G and a g = 4.1 signal is observed between
1200 and 2200 G (Fig. 5). In -ion PS II membranes, about
35% decrease in the amplitude of ML signal intensity was
observed. Oxalate induced further decrease in the amplitude
of the ML signal in -ion PS II membranes confirming that
oxalate binds at the donor side of PS II.
sign
al in
tens
ity, r
el.u
n.
3340332033003280
(c)
(b)
(a)
Magnetic Field (G)
Fig. 3 EPR spectrum for tyrosine radical anions recorded at 253 K in
oxalate-treated thylakoid membranes. (a) Spectrum recorded in dark
(b) after illumination (c) difference spectrum of illuminated minus
dark spectra. The arrowhead indicates the position of the magnetic
field fixed for the kinetic experiments. Experimental conditions:
microwave frequency 9.31 GHz, microwave power 2 mW, modula-
tion amplitude 5 G
sign
al in
tens
ity, r
el.u
n.
10 mM Ox
5 mM Ox
control
642Time [min]
Fig. 4 EPR traces of the decay kinetics of YZ• in oxalate-treated
thylakoid membranes at 253 K. Experimental conditions: microwave
frequency 9.31 GHz, microwave power 2 mW, modulation amplitude
5 G. The magnetic field was fixed at 3300 G for the kinetic
experiments. (a) control; (b) 5 mM oxalate; (c) 10 mM oxalatesi
gnal
inte
nsity
, rel
.un.
10 mM Ox
5 mM Ox
-ion
+ion
Magnetic field (Gauss)5000400030002000
g = 4.1
Fig. 5 S2 state EPR spectra in oxalate-treated PS II membranes. (a)
+ion sample; (b) -ion sample; (c) 5 mM oxalate; (d) 10 mM oxalate.
The EPR spectra are presented as light minus dark difference spectra.
Experimental settings: temperature 6 K, microwave power 2 mW,
microwave frequency 9.417, modulation amplitude 16 G. Chlorophyll
concentration used was 3–4 mg/ml
Photosynth Res
123

Effects of oxalate on PS I
A concentration response of oxalate on PS I activity was
measured. PS I rates were inhibited significantly in the
presence of oxalate (Table 2). There was about 20% inhi-
bition in PS I activity with 5 mM oxalate. To precisely
locate the site of action of oxalate on PS I, PS I mediated
partial electron transfer chain reactions (DCPIPH2 ? MV,
TMPD(red) ? MV, and DAD(red) ? MV) were measured
(Table 3), where the addition of 5 mM oxalate caused
17%, 40% and 36% inhibition, respectively. DAD(red) and
TMPD(red) are known to donate electrons near cytochrome
f in the electron transport chain. DCPIPH2 donate electrons
at two sites, one directly to P700 and the other via plas-
tocyanin (Jajoo and Bharti 1993a, b).
In order to pinpoint the site of action of oxalate on PS I,
we carried out the above mentioned partial reactions in the
presence of mercuric chloride (HgCl2). Mercury specifi-
cally inhibits PS I rates by binding to plastocyanin as well
at the acceptor side (FA/FB). In the presence of HgCl2,
62%, 78%, and 84% inhibition in DCPIPH2 ? MV,
TMPD(red) ? MV, and DAD(red) ? MV reactions were
observed, respectively. On addition of oxalate and HgCl2together, predominant effects of oxalate were observed
(Table 3). In Table 3, we observe that the donor systems
give different maximum rates, whereas in the presence of
oxalate (5 mM), all donor systems give about the same
rates. This suggests that oxalate imposes the same rate
limiting step regardless of the donor. It implies that oxalate
works on the acceptor or reducing side of PS I. Oxalate
could form insoluble salt with HgCl2 thus preventing
inhibition caused by HgCl2. However, this possibility is
ruled out as according to standard chemical data both
mercury chloride (Ksp = 1.4 9 10-18) and mercury oxa-
late (Ksp = 1.7 9 10-13) have very low solubility. Steady
state measurements of photo-induced absorbance changes
at 700 nm related to photoaccumulation of the oxidized
primary donor of PS I were carried out using purified PSI
particles. If we assume a monophasic decay in all cases and
take the half-time value on X-axis, then as shown in Fig. 6,
rate of re-reduction of P700+ is a little slower in oxalate-
treated (5 mM) PS I particles as compared to control. The
back reaction of dark reduction of preliminary primary
electron donor of PS, chlorophyll P700, is characterized by
a half-life of 11.0 ± 0.5 s in the absence of any other
additions, while it was 15.0 ± 0.5 s and 25.7 ± 0.5 s in
the presence of 5 mM and 10 mM oxalate respectively.
Rate of reduction of preliminary primary electron donor of
PS I, chlorophyll P700, was 0.91 9 10-3 mol mg-1 h-1
in the absence of other additions, while it was 0.67 9
10-3 mol mg-1 h-1 and 0.39 910-3 mol mg-1 h-1 in the
presence of 5 mM and 10 mM oxalate, respectively.
Comparison of this data with Table 3 suggests that the
main effect of 5 mM oxalate is on the reducing side of PS
I, but higher concentration of oxalate (10 mM) may also
start to affect the oxidizing side as well.
This study reports for the first time that: (i) unlike all
other anions reported so far, only oxalate inhibits both PS I
and PS II, (ii) oxalate shows its inhibitory effects on PS I
and PS II only when exogenous chloride is not present in
the medium, (iii) the site of action of oxalate is on the
Table 2 Changes in the PS I mediated electron transport rate
(DCPIPH2 ? MV reaction) in the presence of oxalate
Treatment PS I activity (lmol oxygen
consumed mg chl-1 h-1)
Control 409 ± 5 (100)
1 mM oxalate 384 ± 6 (94)
5 mM oxalate 339 ± 5 (83)
10 mM oxalate 339 ± 7 (83)
20 mM oxalate 331 ± 5 (81)
The reaction mixture for PS I measurement contained 0.1 M Sucrose,
20 mM HEPES–NaOH buffer (pH 7.6), 3 mM ascorbate, 5 lM
DCMU, 0.1 mM DCPIP, 0.1 mM methyl viologen, 5 lM sodium
azide and thylakoid suspension equivalent to 20 lg chl/ml. All
experiments were performed three times in triplicates. Values in
parenthesis show the normalized values
Table 3 Changes in various partial reactions mediated by PS I in the presence of oxalate and HgCl2
Treatment PS I activity (lmol oxygen consumed mg chl-1 h-1)
DCPIPH2 ? MV TMPD(red) ? MV DAD(red) ? MV
Control 409 ± 5 (100) 617 ± 16 (100) 502 ± 8 (100)
5 mM oxalate 339 ± 9 (83) 376 ± 4 (60) 322 ± 12 (64)
10 nmol HgCl2 155 ± 8 (38) 136 ± 4 (22) 75 ± 6 (16)
5 mM oxalate + 10 nmol HgCl2 327 ± 9 (78) 364 ± 3 (59) 274 ± 12 (61)
The reaction mixture for PS I measurement contained 0.1 M Sucrose, 20 mM HEPES–NaOH buffer (pH 7.6), 3 mM ascorbate, 5 lM DCMU,
0.1 mM methyl viologen, 5 lM sodium azide and thylakoid suspension equivalent to 20 lg chl/ml. DCPIPH2, TMPD(red) and DAD(red) were
used as electron donors. Concentrations of DCPIPH2, DAD and TMPD were 0.1 mM, 0.05 mM and 0.05 mM, respectively. Values in paren-
thesis show normalized values
Photosynth Res
123

donor side of PS II, (iv) Oxalate inhibits PS I by binding to
donor as well as the acceptor side of PS I.
Acknowledgment This work was supported by the Indo-Russian
Joint project INT/ILTP/B-6.27 and by grants from the Russian
Foundation for Basic Research.
References
Allakhverdiev SI, Yruela I, Picorel R, Klimov VV (1997)
Bicarbonate is an essential constituent of the water-oxidizing
complex of photosystem II. Proc Natl Acad Sci U S A 94:
5050–5054
Babani F, Lichtenthaler HK (1996) Light-induced and age dependent
development of chloroplasts in etiolated barley leaves as
visualized by determination of photosynthetic pigments, CO2
assimilation rates and different kinds of chlorophyll fluorescence
ratios. J Plant Physiol 148:555–566
Bukhov NG, Mohanty P (1999) Elevated temperature stress effects on
photosystems: characterization and evaluation of the nature of
heat-induced impairments. In: Singhal GS, Renger G, Sopory
SK, Irrgang KD, Govindjee (eds) Concepts in photobiology;
photosynthesis and photomorphogenesis. Narosa Publishers,
New Delhi, pp 617–648
Chitnis PR, Xu Q, Chitnis VP, Nechushtai R (1995) Function and
organization of photosystem I polypeptides. Photosynth Res
44:23–40
Debus RJ, Barry BA, Sithole I, Babcock GT, McIntosh L (1988)
Directed mutagenesis indicates that the donor to P680+ in
photosystem II is tyrosine-161 of the D1 polypeptide. Biochemistry
27:9071–9074
Dong D, Peng X, Yan L (2004) Organic acid exudation induced by
phosphorus deficiency and/or aluminium toxicity in two con-
trasting soybean genotypes. Physiol Plant 122:190–199
Ferreira KN, Iverson TM, Maghlaoui K, Barber J, Iwata S (2004)
Architecture of the photosynthetic oxygen evolving center.
Science 303:1831–1838
Feyziev YM, Yoneda D, Yoshii T, Katsuta N, Kawamori A,
Watanabe Y (2000) Formate-induced inhibition of the water-
oxidizing complex of photosystem II studied by EPR. Biochem-
istry 39:3848–3855
Ghirardi ML, Lutton TW, Seibert M (1996) Interactions between
diphenylcarbazide, zinc, cobalt and manganese on the oxidizing
side of photosystem II. Biochemistry 35:1820–1828
Govindjee, Van Rensen JJS (1993) Photosystem II reaction centers
and bicarbonate. In: Deisenhofer J, Norris JR (eds) The photo-
synthetic reaction center, vol I. Academic Press, New York,
pp 357–389
Havaux M, Lannoye R (1985) In vivo chlorophyll fluorescence and
delayed light emission as rapid screening techniques for stress
tolerance in crop plants. Z Pflanzen 95:1–13
Hoganson CW, Babcock GT (1988) Electron transfer events near the
reaction center in O2-evolving photosystem II preparations.
Biochemistry 27:5848–5855
Jajoo A, Bharti S (1993a) Effect of anions on photosystem I-mediated
electron transport in spinach chloroplast. J Exp Bot 44:785–790
Jajoo A, Bharti S (1993b) On the participation of chloride in
bicarbonate-induced reversal of anion inhibition of photosystem
II electron transport in spinach thylakoid membranes. Physiol
Plant 88:78–84
Jajoo A, Bharti S, Kawamori A (2005a) Interactions of chloride and
formate at the donor and the acceptor side of photosystem II. J
Bioenerg Biomembr 37:49–54
Jajoo A, Bharti S, Kawamori A (2005b) EPR characteristic of
chloride-depleted photosystem II membranes in the presence of
other anions. Photochem Photobiol Sci 4:459–462
Jajoo A, Katsuta N, Kawamori A (2006) An EPR study of the pH
dependence of formate effects on Photosystem II. Plant Physiol
Biochem 44:186–192
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauss N
(2001) Three-dimensional structure of cyanobacterial photosys-
tem I at 2.5 A resolution. Nature 411:909–917
Kamiya N, Shen JR (2003) Crystal structure of oxygen-evolving
photosystem II from Thermosynechococcus vulcanus at 3.7-A
resolution. Proc Natl Acad Sci U S A 100:98–103
Klimov VV, Allakhverdiev SI, Shuvalov VV, Krasnovsky AA (1982)
Effect of extraction and re-addition of manganese on light
reactions of photosystem II preparations. FEBS Lett 148:
307–312
Klimov VV, Allakhverdiev SI, Feyziev YM, Baranov SV (1995)
Bicarbonate requirement for the donor side of PS II. FEBS Lett
363:251–255
Klimov VV, Baranov SV, Allakhverdiev SI (1997) Bicarbonate
protects the donor side of photosystem II against photoinhibition
and thermoinactivation. FEBS Lett 418:243–246
Kuwabara T, Murata N (1982) Inactivation of photosynthetic oxygen
evolution and concomitant release of three polypeptides from
photosystem II particles of spinach chloroplasts. Plant Cell
Physiol 23:533–539
Lindberg K, Andreasson LE (1996) A one-site, two-state, model for
the binding of anions in photosystem II. Biochemistry 35:
14259–14267
Loll B, Kern J, Saenger W, Zouni A, Biesiadka J (2005) Towards
complete cofactor arrangement in the 3.0 A resolution structure
of photosystem II. Nature 438:1040–1044
Miller AF, Brudwig GW (1991) A guide to electron paramagnetic
resonance spectroscopy of photosystem II membranes. Biochim
Biophys Acta 1056:1–18
Nakata PA (2003) Advances in our understanding of calcium oxalate
crystal formation and function in plants. Plant Sci 164:901–909
10 mM Ox
5 mM Ox
control
∆A
at 7
00 n
m, r
el.u
n.
Time (sec)0 20 40 60 80 100
Fig. 6 Re-reduction of P700+ in oxalate-treated PS I particles. The
reaction mixture contained 0.33 M sucrose, 50 mM HEPES–NaOH
buffer (pH 7.6), TMPD 100 lM, MV 100 lM and thylakoid
suspension equivalent to 10 lg of chl/ml
Photosynth Res
123

Petrouleas V, Diner BA (1990) Formation by NO of nitrosyl adducts
of redox components of the photosystem II reaction center I NO
binds to the acceptor-side non-heme iron. Biochim Biophys Acta
1015:131–140
Petrouleas V, Deligiannakis Y, Diner BA (1994) Binding of
carboxylate anions at the non-heme Fe(II) of PS II. Competition
with bicarbonate and effects on the QA/QB electron transfer rate.
Biochim Biophys Acta 1188:271–277
Porra RJ, Thompson WJ, Kriedemann PE (1989) Determination of
accurate extinction coefficient and simultaneous equations for
assaying chlorophyll a and b with four different solvents:
verification of the concentration of chlorophyll by atomic
absorption spectroscopy. Biochim Biophys Acta 975:384–394
Schlosser D, Hofer C (2002) Laccase-catalysed oxidation of Mn2+ in
the presence of natural Mn3+ chelators as a novel source of
extracellular H2O2 production and its impact on manganese
peroxidase. Appl Environ Microbiol 68(7):3514–3521
Shinkarev VP, Govindjee (1993) Insight into the relationship of
chlorophyll a fluorescence yield to the concentration of its
natural quencher in oxygenic photosynthesis. Proc Natl Aca Sci
U S A 90:7466–7469
Shuvalov VA, Klimov VV, Krasnovskii AA (1976) Investigation of
primary photoprocesses in light fragment of chloroplasts. Mol
Biol Russ 10(2):326–339
Skorzynska E, Baszynski T (2000) Does Cd use Ca channels to
penetrate into chloroplasts?—a preliminary study. Acta Physiol
Plant 22:171–178
Stemler AJ, Lavergne J (1997) Evidence that formate destabilizes the
S1 state of the oxygen-evolving mechanism in photosystem II.
Photosynth Res 51:83–92
Wincencjusz H, Yocum CF, Gorkom JV (1999) Activating anions
that replace Cl- in the O2-evolving complex of photosystem II
slow the kinetics of the terminal step in water oxidation and
destabilize the S2 and S3 states. Biochemistry 38:3719–3725
Yamane Y, Kashino Y, Koike H, Satoh H (1998) Effects of high
temperature on the photosynthetic systems in spinach oxygen-
evolving activities, fluorescence characteristics and the denatur-
ation process. Photosynth Res 57:51–59
Photosynth Res
123





























