Embed Size (px)
Citation preview

Environmental determinants of the composition of desert-livingrodent communities in the north-east Badia region of Jordan
Dawn M. Scott and Nigel Dunstone*
Department of Biological Sciences, University of Durham, South Road, Durham City DH1 3LE, U.K.
(Accepted 10 September 1999)
Abstract
The spatial and temporal compositions of rodent communities in habitats of the Badia region of north-east
Jordan were investigated between 1996 and 1997. Live grid-trapping and spot-lighting techniques were
employed to estimate rodent densities. Sixteen environmental variables including soil content, vegetation
cover and relief were recorded within the study sites. TWINSPAN was used to derive six habitat classes
based on the similarity of their rodent communities. These were hammada, harrat, marab, wadi and two
types of sand dunes. The derived classi®cations were similar to the original categorization of habitat types
based on ground-truthed observations of the sites selected from satellite imagery. Multivariate statistical
analyses (TWINSPAN and DECORANA) revealed three major rodent groups in relation to local habitat
types: (1) petrophiles: Gerbillus dasyurus, Acomys russatus and Meriones tristrami; (2) open hammada-
dwelling species: Jaculus jaculus, Gerbillus henleyi and Meriones crassus; (3) psammophiles: Meriones
libycus, Gerbillus nanus and Gerbillus cheesmani. Species richness was highest in marabs and lowest in red
sand dunes. Species diversity (Hill's index) was highest in sand dunes and lowest in red sand dunes. Rodent
biomass was highest in sand dunes and lowest in wadis. CANOCO showed rodent community composition
to be correlated with vegetation parameters and soil content. TWINSPAN grouping and DECORANA
ordination of rodent species provides some evidence for habitat preference, size disparity and competition
as mechanisms of resource partitioning facilitating coexistence.
Key words: rodents, desert-living, environments, petrophiles, psammophiles
INTRODUCTION
Desert-living rodents have frequently been used toexamine the mechanisms underlying the organization ofcommunities (e.g. Rosenzweig & Winakur, 1969). Theemphasis of many of these studies has been concernedwith how resource partitioning facilitates coexistence(Bowers & Brown, 1982; Kotler & Brown, 1988), byinvestigating competition (Heske, Brown & Mistry,1994), morphology (Bowers & Brown, 1982), predation(Kotler, 1984; Bowers & Doley, 1993), foragingbehaviour (Kotler, 1997), diet (Harris, 1986), andmacro- and micro-habitat preferences (Price, 1978;Bowers, 1988). Attempts have also been made to assesshow environmental factors such as resource availability,estimated in terms of seed density (Price & Heinz, 1984),productivity (Abramsky, 1988) and precipitation(Beatley, 1969), affect the composition of rodent com-
munities, i.e. the diversity and relative abundance ofspecies present in a particular location.
Organisms in desert habitats have to cope with spatialheterogeneity, temporal variability and unpredictabilityof resources, as well as competition and predation(Safriel et al., 1989). Therefore the mechanisms under-lying community organization, such as competition, willalso be subject to spatial and temporal variability. In astudy comparing the organization of desert rodent com-munities, Rogovin & Surov (1990) suggested that theunderlying mechanisms may vary between deserts. Intheir study, interspeci®c competition seemed to play amore signi®cant role in the structure of rodent commu-nities in Mexican deserts than those in Mongolia, wherepredators and environmental conditions were thoughtto be more important. However, in an investigation ofseven geographically distinct deserts, Kelt et al. (1996)found that some deserts exhibited similarities, e.g.rodent diversity, and suggested that their communitycomposition may be in¯uenced by common underlyingmechanisms. By examining further communities of
*All correspondence to: N. Dunstone.E-mail: [email protected]
J. Zool., Lond. (2000) 251, 481±494 # 2000 The Zoological Society of London Printed in the United Kingdom

desert-living rodents from different geographical regionsit should be possible to determine whether there aresimilar mechanisms of coexistence operating withindeserts and between deserts on a global level. Thesetrends can thus help to provide an understanding ofwhich patterns of organization reoccur and which aredetermined by unique community composition(Shenbrot & Rogovin, 1995).
Structural and functional convergence of rodentcommunities has been found in similar habitats in geogra-phically isolated deserts. The coexisting rodent speciesseemed to have comparable variations in morphology,foraging behaviour, and more strikingly, similar patternsof body size (J. H. Brown, Reichman & Davidson, 1979).Bowers & Brown (1982) investigated the regular andapparently logarithmic displacement in body size. Theydemonstrated that granivores of similar body sizeoccurred together much less frequently than expected andsuggested that size disparity supported interspeci®ccompetition as a major role in determining the structureof granivorous desert rodent communities.
Analysis of the relationship between the compositionof rodent communities and prevailing environmentalparameters has commonly used ordination and clusteranalysis techniques, although these indirect methodshave generally less strength than direct methods e.g.regression analysis. Krasnov et al. (1996) have investi-gated the rodent communities in the Ramon erosioncirque in Israel, using cluster analysis to separatehabitat types by the similarity of rodent species inha-biting them. Correspondence analysis was used as anindirect ordination technique to determine what envir-onmental factors may be in¯uencing the composition ofrodent communities. Discriminant function analysis orcanonical variate analysis has also been a popularanalytical approach. Shenbrot (1992) investigatedspatial displacement (i.e. the shift of habitat preferencesalong ecological axes) of desert-living species as amechanism facilitating coexistence, using discriminantanalysis to describe `niches' and niche distribution inresource space (Rogovin, Shenbrot & Surov, 1991;Shenbrot & Rogovin, 1995). Canonical correspondenceanalysis (CCA) was originally designed to study plantcommunities and associated environmental gradients(Ter Braak, 1987a,b). It has previously been used inother aspects of vegetation ecology such as succession(Vanzyl & Robertson, 1991) and conservation (Taggart,1994), and has more recently been applied toinvertebrate communities, e.g. microinvertebrates(Ruse, 1996), water beetles (Behr, 1994), earthworms(Roderiguez, Trigo & Cosin, 1997), Nematoda (Soetaertet al., 1995), Odonata (Samways & Steytler, 1996),Polychaeta (Mackie, Parmiter & Tong, 1997); to date ithas not been applied to the analysis of the compositionof mammalian communities.
The aim of the present study was two-fold; ®rst, toinvestigate the spatial and temporal composition ofrodent communities in the north-east Badia region ofJordan, and second, to determine which environmentalparameters may be important in determining such
communities. We adopt a hierarchical analyticalapproach to facilitate a comparison with previousstudies of similar species and/or habitats in an attemptto identify re-occurring patterns in desert communityorganization. In this analysis we use CCA to determinewhich environmental parameters affect the compositionof rodent communities (Ter Braak, 1987a).
MATERIALS AND METHODS
Study area
The investigation was conducted over 2 years within anarea of the north-east Badia region of Jordan (seeFig. 1) which covers a total area of 11 210 km2 in thewestern part of the Jordan panhandle extendingtowards the border with Iraq (Dutton, Clark & Battikhi,1998). The area studied was from Wadi Hashad(32830'N, 37820'E) and Burquh (32837'N, 37857'E) inthe north to Hazim (31835'N, 37815'E) and Qataf®(31849'N, 37832'E) in the south. The climate of thisregion is continental with temperatures ¯uctuatingbetween extremes: summer averages range from 46 8Cmaximum to 15 8C minimum and winter averages from15 8C maximum to 77 8C minimum. Generally theannual rainfall is between 50 and 200 mm and mainlyfalls between October and April, peaking in December(Kirk, 1998). The timing and quantity of rainfall ishighly variable and will directly affect new ephemeraland perennial plant growth, which will subsequentlyaffect yearly seed production and consequently the seedbank; the main food source for granivorous rodents.
The landscape is varied, ranging from basalt desert inthe north to the sand dunes of the south. The tertiary±quaternary continental basalt ¯ows and tuffs cover alarge proportion of the area (Dutton et al., 1998). Theprincipal habitat types in this region are:(1) harrat, basalt rock areas which vary in relief fromplains to steep slopes, dominated by large boulders over-lying basaltic soils which tend to be of shallow, sandyloam. Vegetation occurring on the harrat is generallyephemeral e.g. Leysera leyseroides and Graminae spp.;(2) wadis or seasonal rivers, formed as a result of poorin®ltration of the soil, which, although classi®ed assandy loam, contains a high proportion of gravelaccounting for its moderate penetrability. The vegeta-tion within the wadi is typically comprised of Artemisiaherba-alba, Achillea fragrantissima, Astragulus spinosus,Thymus bovii and chenopod species;(3) marab, a low lying area commonly situated at themouth of the wadi having relatively permeable non-saline deposits with a high organic content. Marabs aregenerally vegetated with A. herba-alba, Zilla spinosa andA. fragrantissima, on clay loam soils with a moderateloose soil depth (LSD);(4) qa'a, a low lying area which, due to poor penetr-ability, act as a collection site for rainwater. When therainfall evaporates the area forms large saline ¯ats,which are devoid of vegetation;
D. M. Scott and N. Dunstone482

(5) hammada, or chertz plains, are characterized by anabsence of basalt rocks and have minimal vegetationcover. Where present, this comprises A. herba-alba andephemeral species e.g. Diplotaxis harra, Leysera leyser-iodes and Graminae spp. growing on sandy loam soilsof moderate LSD;(6) sand dunes, semi-stabilized and highly vegetatedwith Haloxylon salicornicum, Raetama raetum, Tamarixaphylla and chenopod species with a deep layer of loosesandy soil.
The vegetation cover of all these habitats can varydramatically between seasons. Some areas with a highpercentage of cover in the spring can become virtuallybarren in the summer.
Rodent species
Previous to this study there was little publishedinformation on the rodent diversity of the Badia region(Searight, 1987). A list of species potentially found inthe area was compiled from distribution maps (Harrison
& Bates, 1991; pers. comm.). These compriseGerbillidae: Meriones crassus (Sundevall, 1842),M. libycus (Lichtenstein, 1823), M. tristrami (Thomas,1892), Gerbillus dasyurus (Wagner, 1842), G. henleyi(De Winton, 1903), G. nanus (Blanford, 1875), G. chees-mani (Thomas, 1919); Dipodinae: Jaculus jaculus(Linnaeus, 1758); and Muridae: Acomys russatus(Wagner, 1840). These range in weight from the 120 gM. libycus to the diminutive G. henleyi (11g). Most ofthese species are nocturnal with the exception ofM. libycus (which is considered mainly diurnal withactivity peaking early morning and mid to late after-noon (Atallah, 1977). Acomys russatus has beenreported as diurnal, only becoming nocturnal when notin competition with other Acomys spp. (Shkolnik,1971). All species are reported to be mainly granivorous,but became more omnivorous in response to changes inresource availability and physiological demands. Jaculusjaculus, G. cheesmani, G. henlyei and G. dasyurus aresolitary, whereas A. russatus, G. nanus, M. libycus andM. crassus are considered to be gregarious (Harrison &Bates, 1991).
483Communities of desert rodents
Fig. 1. The study area in the north-east Badia region of Jordan showing the localities of study sites.
0 50km (approx)
SYRIA
Burquh
Wadi Hashad
Wadi Salma
1200
1000
800
Bqa'awia
Qattafi
600
HazimDhahik
800
SAUDI ARABIA

Field methods
Rodent were trapped between February and September1996 and from February to June 1997. Potentiallysuitable study sites, representative of different habitattypes, were located using a false colour satellite image(bands 7, 4 and 2, path 173, row 32/2N, contact= 1/1 000 000) taken on the 29 March 1992. Sites were®nally selected following ground-truthing of habitatsimmediately before trapping studies.
Trapping was undertaken for 5 consecutive nights ateach site using 48 Sherman traps (236967.5 cm)arranged in a 664 grid con®guration with 2 traps perstation located at 20 m intervals. Traps were baited1±2 h before sunset with a mixture of cereal seeds(typically barley or wheat) and peanut butter andchecked at ®rst light. Species captured were identi®ed,individually marked with a unique hair clip, weighed,morphometrically measured, aged, sexed and sexualstatus determined. All individuals were then released attheir point of capture. Sherman traps were not suitablefor catching jerboas J. jaculus, which have previouslybeen reported as trap-shy (Happold, 1967; J. S. Brownet al., 1994) possibly owing to bipedal locomotion. Cagetraps as used by Hatough-Bouran & Al-Eisawi (1990)were unavailable and not suitable for the study pro-tocol. Jerboa density was estimated by night transects,undertaken by driving at a slow constant speed (5 km/h)along tracks within the study sites. Observers spot-lighted the area with a 1 000 000 candlepower search-light. The numbers sighted 1 h after dark, over a settransect length (5 km) and breadth (50 m) were used toestimate density.
As a result of the temporal variability in rodentdensities and environmental variables, samples taken atdifferent times (e.g. months or, more usually, seasons)were considered independent (see Table 1). Three trap-ping sessions were undertaken to show seasonal andannual variation in rodent densities and distributions:spring 1996, summer 1996 and spring 1997. Six gridsrepresentative of 4 habitat types (harrat, marab, wadiand sand dunes) were trapped during each session. In1997, a further 6 habitat grids were sampled. Two gridsin Hazim (hammada and sand dunes) and 2 grids inWadi Salma (wadi and harrat) were trapped at monthlyintervals to investigate temporal variability within agiven season. A total of 12 grids of 0.8 ha were sampledover the 3 sessions. Trapping was not conductedbetween October and February.
Environmental variables
Sixteen environmental parameters were measured at 528(1 m2) plots within the study sites (Table 2). Threevariables were scored on arbitrary scales: local relief(where a sloping angle of 0±158= 1, 16±308= 2, 31±458= 3 and > 458= 4); human presence (1 = low to5 = high); and grazing level (1 = low to 5 = high). Percen-tage covers of perennials, ephemerals, basalt, rocks and
stones were estimated visually in each of the plots. Theheights of perennial and ephemeral plants weremeasured to the nearest centimetre. Loose soil depth(LSD) was estimated by measuring the penetration of agraduated spike to which a ®xed pressure was applied.Altitude measurements were taken using a GarminGPS 45, with up to 5 position ®xes taken to verifythe constancy of the altitude reading which was latercompared with local topographical maps.
Soil samples were collected from each plot by takingcores (4 cm diameter65 cm deep). Five randomsamples were selected and used to assess particle size.The dried samples were ®rst passed through a 2 mmsieve and reduced to 0.5 g using a soil divider. Eachsample was then treated with 2 ml of 20% volumehydrogen peroxide and placed in a boiling water bathfor 2 h to remove the organic component. Distilledwater was added to the tubes and subsequently sampleswere centrifuged for 4 min at 40 000 rev/min. Anyresidual organic component was then decanted from thesample. Samples were then treated with 2 ml of awetting agent (10% sodium hexametaphosphate) andstirred. Particle size was measured using a Coulter LS230 laser granulometer, keeping the obscuration atc. 12%. Particle size classi®cations for estimation ofpercentages of clay, silt, sand and gravel were according
D. M. Scott and N. Dunstone484
Table 1. Description of sample sites
Sampleno. Site name Season Year Habitat type
1 Dhahik Spring 96 Tamarix dunes2 Dhahik Summer 96 Tamarix dunes3 Dhahik Spring 97 Tamarix dunes4 Bqa awia Spring 96 Marab5 Bqa awia Summer 96 Marab6 Bqa awia Spring 97 Marab7 Wadi Hashad Spring 96 Marab A8 Wadi Hashad Summer 96 Marab A9 Wadi Hashad Spring 97 Marab A
10 Wadi Hashad Spring 97 Marab B11 Hazim Spring 96 Sand dunes12 Hazim Summer 96 Sand dunes13 Hazim Spring, March 97 Sand dunes14 Hazim Spring, April 97 Sand dunes15 Hazim Spring, May 97 Sand dunes16 Hazim Spring, March 97 Hammada17 Hazim Spring, April 97 Hammada18 Hazim Spring, May 97 Hammada19 Qataf® Spring 97 Red dunes20 Burquh Spring 97 Hammada21 Burquh Spring 97 Harrat22 Wadi Salma Spring 96 Harrat23 Wadi Salma Spring, March 97 Harrat24 Wadi Salma Spring, April 97 Harrat25 Wadi Salma Spring, May 97 Harrat26 Wadi Salma Spring 96 Marab27 Wadi Salma Summer 96 Marab28 Wadi Salma Summer 96 Wadi29 Wadi Salma Spring, March 97 Wadi30 Wadi Salma Spring, April 97 Wadi31 Wadi Salma Spring, May 97 Wadi

to Church, Mclean & Wolcott (1987). Soil types werenamed from the resulting proportions of particle size ineach sample using a soil texture classi®cation chart(after FAO, 1977).
Statistical analysis: multivariate methods
Two-way Indicator Species Analysis (TWINSPAN;Hill, 1979) was used to group samples on the basis oftheir rodent species composition, and to group rodentspecies on the basis of their spatial distribution. Valuesused in the analyses are averages (n = 20) from each site.All parameters in the program were set at default valuesin the analyses, as standard (Hill, 1979).
De-trended correspondence analysis (DCA) was used(DECORANA program; Hill, 1979) as an ordinationtechnique to identify rodent and site groupings, and todetermine which environmental gradients were re¯ectedin these groupings. The relationship between speciesgroups and environmental variables was assessed bycomparing ecological parameters between sites withinthe study area. Species-density estimates were used inthe analysis and all parameters were set to the programdefault values.
CCA was employed using the FORTRAN computerprogram CANOCO (Ter Braak, 1988). CCA is amethod of arranging species along environmentalvariables (Ter Braak, 1987a). All variables used in theanalysis were quantitative, and their values werestandardized to mean 0 and variance 1. Variablesshowing co-linearity in the initial program outputwere excluded from further analysis. The data werenot transformed. This ordination technique was usedto detect the relationship between environmentalvariables and rodent species community composition.A Monte Carlo permutation test (Manly, 1990) wasused to examine the signi®cance of the correlationbetween the ®rst canonical axis and rodent communitycomposition.
RESULTS
Habitat types
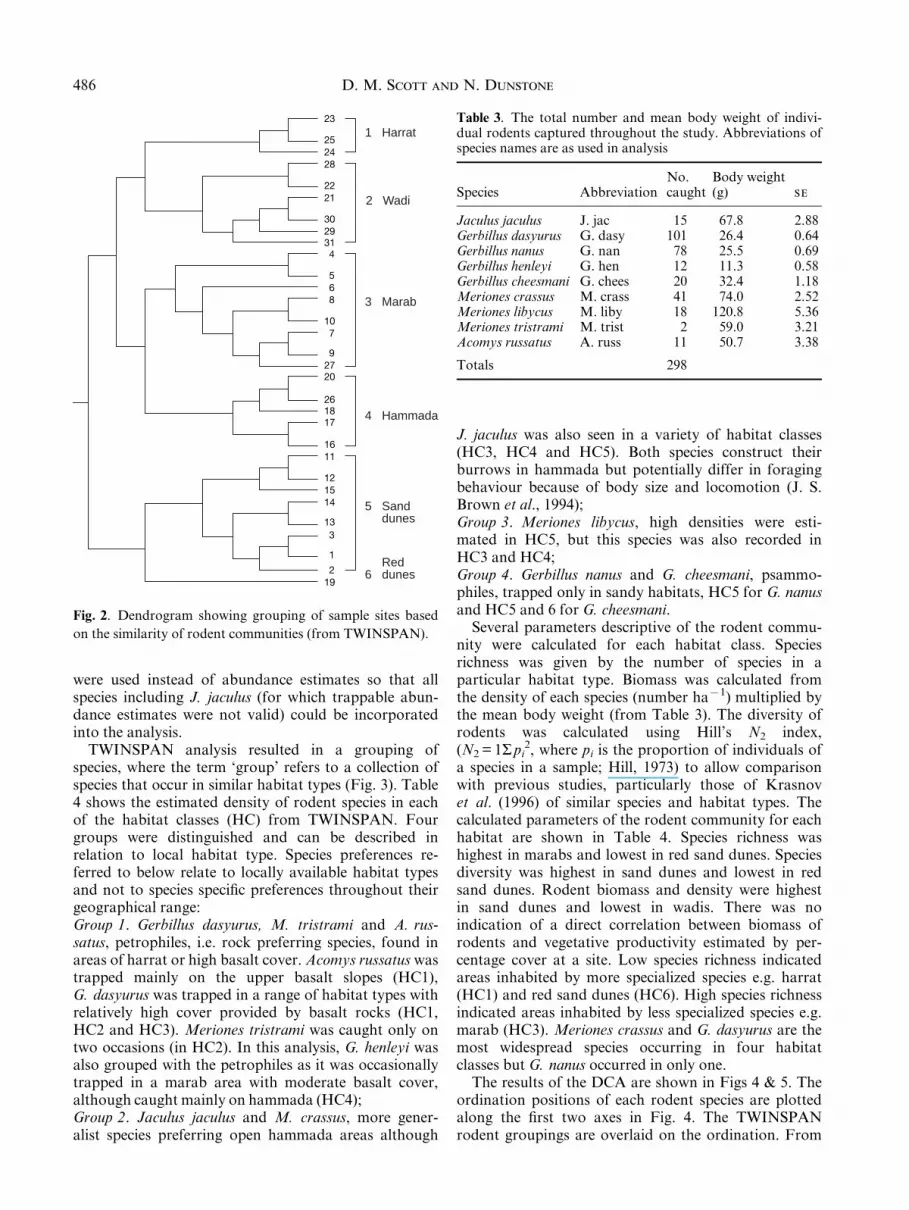
TWINSPAN analysis was used to derive habitat classes(HC) based on the similarity of their rodent communitydensities. Figure 2 shows the grouping of sample sitesinto habitat classes. The derived classi®cations weresimilar to the original categorization of habitat typesbased on ground-truthed observations of the sitesselected from the false colour satellite image. Six mainhabitat classes were: (HC1) basalt slopes (harrat);(HC2) wadi and areas with high basalt cover; (HC3)marab; (HC4) hammada; (HC5) sand dunes; (HC6) redsand dunes. Most of the temporal and spatial samplesgrouped as expected. For example, the monthly samplesfrom Wadi Salma basalt slopes all grouped into HC1.The second class, which consisted of samples from wadihabitat at Wadi Salma and from the basalt plains ofWadi Salma and Burquh, arises because the lowerbasalt ¯ats are more similar in species composition towadis than the upper basalt slopes. HC3 comprisedsamples collected over different seasons from three sites(Bqa awia, Hashad and Salma) demonstrating temporaland spatial consistency within this habitat class. Thehammada (HC4) showed one anomaly; marab habitatat Wadi Salma sampled in spring 1996 was grouped inthis class along with samples collected from hammadahabitat at Hazim and Burquh. Variable classi®cation oftemporal samples from the same site indicates annualand seasonal variability in rodent community composi-tion. A red sand dune class (HC6) was identi®ed asdistinct from HC5 (other sand dunes). This probablyresulted from differences in species richness at eachsite, HC5 having ®ve species present and HC6 onlyhaving one.
Rodents
During the study 298 individuals of nine species ofrodents were captured. The mean body weight wascalculated from captures of all adult individuals ofspecies from all habitats (Table 3). This was done toexamine body mass between species and size disparitywithin groups. Gerbillus dasyurus was the most com-monly trapped gerbil species, and Meriones crassus wasthe most commonly captured jird species. Merionestristrami was only rarely encountered. There was sometemporal variability in trap success. The trappingsuccess (number of captures/number of trap nights) was14.2%, 10.81.% and 7.75%, for spring 1996, summer1996 and spring 1997, respectively. In spring 1997 thetrap success rate for March, April and May was 7.9%,4.7% and 10.6%, respectively. Owing to the low capturerates the most suitable method of estimating density wasto use the total number of individuals captured within atrapping grid. The overall trapping area was calculatedfrom the grid con®guration plus a 10 m boundary stripsurrounding the grid (Blair, 1942). Density estimates
485Communities of desert rodents
Table 2. The 16 environmental variables recorded at each plot
Abbreviation Variable Unit
Alt Altitude mRelief Angle of slope of ground ArbitraryGrazing Level of grazing pressure ArbitraryHumans Level of human disturbance Arbitrary% Clay Clay content of soil %% Silt Silt content of soil %% Sand Sand content of soil %% Gravel Gravel content of soil %LSD Loose soil depth mm% Perenn Perennial cover %PerHt Perennial height cm% Epheme Ephemeral cover %EphHt Ephemeral height cm% Basalt Cover of basalt rocks %% Rock Cover of other rocks (>5 cm) %% Stone Cover of stones (between 1 and 5 cm) %
were used instead of abundance estimates so that allspecies including J. jaculus (for which trappable abun-dance estimates were not valid) could be incorporatedinto the analysis.
TWINSPAN analysis resulted in a grouping ofspecies, where the term `group' refers to a collection ofspecies that occur in similar habitat types (Fig. 3). Table4 shows the estimated density of rodent species in eachof the habitat classes (HC) from TWINSPAN. Fourgroups were distinguished and can be described inrelation to local habitat type. Species preferences re-ferred to below relate to locally available habitat typesand not to species speci®c preferences throughout theirgeographical range:Group 1. Gerbillus dasyurus, M. tristrami and A. rus-satus, petrophiles, i.e. rock preferring species, found inareas of harrat or high basalt cover. Acomys russatus wastrapped mainly on the upper basalt slopes (HC1),G. dasyurus was trapped in a range of habitat types withrelatively high cover provided by basalt rocks (HC1,HC2 and HC3). Meriones tristrami was caught only ontwo occasions (in HC2). In this analysis, G. henleyi wasalso grouped with the petrophiles as it was occasionallytrapped in a marab area with moderate basalt cover,although caught mainly on hammada (HC4);Group 2. Jaculus jaculus and M. crassus, more gener-alist species preferring open hammada areas although
J. jaculus was also seen in a variety of habitat classes(HC3, HC4 and HC5). Both species construct theirburrows in hammada but potentially differ in foragingbehaviour because of body size and locomotion (J. S.Brown et al., 1994);Group 3. Meriones libycus, high densities were esti-mated in HC5, but this species was also recorded inHC3 and HC4;Group 4. Gerbillus nanus and G. cheesmani, psammo-philes, trapped only in sandy habitats, HC5 for G. nanusand HC5 and 6 for G. cheesmani.
Several parameters descriptive of the rodent commu-nity were calculated for each habitat class. Speciesrichness was given by the number of species in aparticular habitat type. Biomass was calculated fromthe density of each species (number ha71) multiplied bythe mean body weight (from Table 3). The diversity ofrodents was calculated using Hill's N2 index,(N2 = 1Spi
2, where pi is the proportion of individuals ofa species in a sample; Hill, 1973) to allow comparisonwith previous studies, particularly those of Krasnovet al. (1996) of similar species and habitat types. Thecalculated parameters of the rodent community for eachhabitat are shown in Table 4. Species richness washighest in marabs and lowest in red sand dunes. Speciesdiversity was highest in sand dunes and lowest in redsand dunes. Rodent biomass and density were highestin sand dunes and lowest in wadis. There was noindication of a direct correlation between biomass ofrodents and vegetative productivity estimated by per-centage cover at a site. Low species richness indicatedareas inhabited by more specialized species e.g. harrat(HC1) and red sand dunes (HC6). High species richnessindicated areas inhabited by less specialized species e.g.marab (HC3). Meriones crassus and G. dasyurus are themost widespread species occurring in four habitatclasses but G. nanus occurred in only one.
The results of the DCA are shown in Figs 4 & 5. Theordination positions of each rodent species are plottedalong the ®rst two axes in Fig. 4. The TWINSPANrodent groupings are overlaid on the ordination. From
D. M. Scott and N. Dunstone486
232524
2822
21302931
456
810
79
27
1 Harrat
2 Wadi
3 Marab
202618
4 Hammada
5 Sand dunes
Red6 dunes
1716
111215
1413
312
19
Fig. 2. Dendrogram showing grouping of sample sites based
on the similarity of rodent communities (from TWINSPAN).
Table 3. The total number and mean body weight of indivi-dual rodents captured throughout the study. Abbreviations ofspecies names are as used in analysis
No. Body weightSpecies Abbreviation caught (g) se
Jaculus jaculus J. jac 15 67.8 2.88Gerbillus dasyurus G. dasy 101 26.4 0.64Gerbillus nanus G. nan 78 25.5 0.69Gerbillus henleyi G. hen 12 11.3 0.58Gerbillus cheesmani G. chees 20 32.4 1.18Meriones crassus M. crass 41 74.0 2.52Meriones libycus M. liby 18 120.8 5.36Meriones tristrami M. trist 2 59.0 3.21Acomys russatus A. russ 11 50.7 3.38
Totals 298
23
252428
2221
3029314
568
107
92720
261817
1611
121514
13
3
1
219

the clustering of species shown by DCA, the speciesgroupings can be categorized on the basis of their localhabitat preferences: (1) petrophiles; (2) generalist - openhammada species; (3) psammophiles. The ordination ofsites is plotted in Fig. 5. The habitat classi®cations areoverlaid on the ordination to show site groupings. Ingeneral, the site groupings remain clear, re¯ecting thehabitat classes with distinct rodent communities.However, there is spatial and temporal overlap of somesite groupings, indicating that these had highly variablerodent communities that were not unique to a particularhabitat class. As the DCA ordination results andTWINSPAN groupings are comparable this suggeststhat environmental parameters may have a majorin¯uence on species composition. The ®rst DCA axiscan be interpreted as representing the gradient frombasalt to sand.
CCA was carried out to investigate further the rela-tionships between species distributions andenvironmental variables suggested by TWINSPAN andDCA. CANOCO was then used to determine whichenvironmental factors were most highly correlated. Theordinations of environmental variables in the ®rst twocanonical axes were plotted using bi-plot scores fromCCA (Fig. 6). The ®rst canonical axis correlated mosthighly with basalt cover, soil sand content, percentage
silt, perennial height and ephemeral height. This axisaccounted for 42.9% of the variance in weighted averageof the species for each environmental variable (eigen-value = 0.895, r = 0.99). The second axis related toaltitude and local relief and accounted for 23.8% of thevariance, (eigenvalue = 0.492, r = 0.93). The third andfourth axes accounted for < 25% of the total varianceand will not be considered further. A Monte Carlopermutation test (Manly, 1990) showed that the ®rstcanonical axis was signi®cantly correlated to speciesdistribution patterns (P < 0.01). The clusters of sites andspecies derived from the DCA (Figs 4 & 5) can berelated to environmental variables in Fig. 6. The groupon the left hand side of Fig. 4 comprises the petrophilerodents associated with harrat (HC1) and wadi (HC2)habitats of Fig. 5. These sites have high basalt cover,more ephemeral cover and height and tend to have ahigher soil content of silt and clay. The second rodentgroup, found in open hammada (HC4), is located in thecluster in the middle of Fig. 4. This group is associatedwith low basalt cover and low perennial cover. Thethird group on the far right hand side comprises thepsammophilous rodent species and sand dune habitats(HC5 and HC6) which are associated with a largepercentage of sand content in the soil and increasedperennial height.
487Communities of desert rodents
Table 4. Density (no. ha71) of rodent species in the derived habitat classes. Species richness (no. of species), overall density,diversity (Hill's N2 index) and biomass values (g)
Habitat class
HC1 HC2 HC3 HC4 HC5 HC6Species Harrat Wadi Marab Hammada Sand dunes Red dunes
Jaculus jaculus 0.00 0.00 0.09 0.08 0.08 0.00Gerbillus dasyurus 11.25 6.32 7.37 0.21 0.00 0.00Gerbillus nanus 0.00 0.00 0.00 0.00 10.86 0.00Gerbillus henleyi 0.00 0.00 0.26 2.50 0.00 0.00Gerbillus cheesmani 0.00 0.00 0.00 0.00 2.71 8.75Meriones crassus 0.00 0.21 3.46 2.46 1.04 0.00Meriones libycus 0.00 0.00 0.13 0.25 2.03 0.00Meriones tristrami 0.00 0.35 0.00 0.00 0.00 0.00Acomys russatus 4.17 0.00 0.13 0.00 0.00 0.00
Species richness 2.00 3.00 6.00 5.00 5.00 1.00Site density 15.42 6.88 11.41 5.49 16.71 8.75Diversity 1.63 1.14 1.73 1.91 1.96 1.00Biomass 534.71 193.91 476.33 245.26 695.45 308.00
Meriones tristramiAcomys russatusGerbillus dasyurusGerbillus henleyi
Meriones crassusJaculus jaculus
Meriones tlibycusGerbillus nanusGerbillus dcheesmani
1
2
3
4
Fig. 3. Dendrogram showing grouping of rodent species based on distribution (from TWINSPAN).
Meriones tristrami
Acomys russatusGerbillus disyurus
Gerbillus henleyiMeriones crassus
Jaculus jaculusMeriones libycusGerbillus nanus
Gerbillus cheesmani

DISCUSSION
The north-east Badia is considered depauperate forrodent biodiversity. Many of the species whose distribu-tions from published maps (Harrison & Bates, 1991)were thought to encompass this area were not encoun-tered during this study. For example, Allactaga
euphratica has previously been reported to occur at lowdensities in Shaumari Wildlife Reserve, a protected areasituated close to the Badia region (Hatough-Bouran &Al-Eisawi, 1990). Lower productivity of deserts inunprotected areas and interspeci®c competition mayaccount for the absence of this and other potentialspecies. Additionally, species at the limit of their
D. M. Scott and N. Dunstone488
200
180
100
20
0
0 50 100 150 200 250 300 350 400 450 500
40
60
80
160
120
140
222128
3029
31
2523
24
Wadi
Harrat
4
5
8
6
7
27
10
9
2620
18 16Marab
Hammada
17
Sand dunes
Red dunes
21
3
121113
14
15
19
Fig. 5. Site ordination based on rodent similarity using DCA. Grouping corresponds to habitat classi®cations in Fig. 2.
Acomys russatus
Jaculus jaculus
Gerbillus dasyurus
Gerbillus cheesmani
Gerbillus henleyi
Gerbillus nanus
Meriones crassus
Meriones libycus
Meriones tristrami
1
2
3
300
250
200
150
100
50
0
–50
–50
0 50 100 150 200 250 300 350 400 450 500
Fig. 4. Positions of rodent species along the ®rst two ordination axes from DCA. TWINSPAN groupings are hand drawn for
illustrative purposes only. Circled ®gures refer to the categories of groups: 1, petrophiles; 2, generalists; 3, psammophiles.

geographical range may be only encountered whereconditions are particularly favourable.
Trap dimensions were selected to encompass the sizerange of all species likely to be encountered. Before thestudy, a preliminary investigation was undertaken toassess an ef®cient trapping period. No species wereexclusively diurnal and the employed trapping period(early evening to dawn) recorded all individuals asencountered during 24 h trapping. The restricted periodchosen also avoided the death of animals resultingfrom extreme diurnal temperatures. Disturbance bylivestock and adverse weather conditions caused invali-dation and hence exclusion of some grid samples fromthe analysis.
Grid trapping was undertaken within homogeneousareas of chosen habitats, since border areas betweenhabitats may contain other species and could in¯uencecommunity composition of the sample area. Forexample, only G. cheesmani was captured at Qatta®although M. libycus was sighted close-by and oneindividual A. russatus was recorded in the borderingbasalt area. Furthermore, grid-based live-trappingtechniques may encounter individuals whilst they areusing the area for one particular aspect of their ecology,e.g. preliminary radio-tracking studies suggest thatM. crassus differed in the habitats used for foraging andburrowing (T. Maddox, pers. comm.). Hence, spatialvariation in community composition and structure canbe viewed on a macrohabitat and microhabitat scale.
The seed bank and productivity
Seeds are an important and limited resource forgranivorous rodents and the competition for seeds,which is directly affected by rodent density, has beenshown to in¯uence community structure (Brown &Lieberman, 1973). The seed bank is highly dependent onannual productivity which is itself related to rainfall(Reichman, 1984). The winter of 1994±95 in Jordan hadan above average rainfall, which resulted in a productivespring and a likely high level of input into the seed bankduring spring 1996. The winter of 1995±96, however,was drier and resulted in a lower vegetation cover in thefollowing spring. This could have negatively affected theseed bank for spring 1997. As yet there is little informa-tion available on which species of seed are preferentiallyforaged by desert rodents. Hence, at this stage it is notpossible to correlate plant and rodent species distribu-tions. An investigation is ongoing to assess the seedoutput of different species of plants with the aim ofevaluating resource availability within differenthabitats.
Evidence that the extent of the seed bank affectsrodent density is re¯ected in the trap success, whichdecreased by 50% from spring 1996 to spring 1997.However, annual or seasonal periods of naturally highresource availability may in¯uence trapping success asspecies become less dependent on bait as a food sourceand hence less trap-prone. The decrease in trapping
489Communities of desert rodents
300
300200
200
100
1000
0
–100
–100
–200
–200
–300
–300
–400
Axi
s 2
Axis 1
% Stone
Grazing% Gravel
% Epheme
EphHt
% Clay% Silt
% Basalt
Alt
Relief
% Perenn
PerHt
LSD
% Sand
Humans
Fig. 6. Ordination of environmental variables in the ®rst two canonical axes using bi-plot scores from CANOCO (CCA). See
Table 2 for meaning of abbreviations.

success from spring to summer 1996 may be the result ofincreased food availability, i.e. the new seed production.Density estimates ¯uctuate temporally as well asspatially, and in this study varied dramatically betweenmonths, seasons and years. Annual rainfall mayaccount for some of the temporal ¯uctuations in densityand community composition. Most desert rodents breedduring late winter with juveniles recruited into thepopulation during spring (Harrison & Bates, 1991). Thisrecruitment and subsequent dispersal may causemonthly ¯uctuations in population density and mayaccount for the observed seasonal decrease in trappingsuccess.
An animal's response to variations in the environmentcan result in spatial and temporal patterns of commu-nity composition (Yajun et al., 1997). Habitat use mayalso ¯uctuate temporally depending on the variability ofenvironmental factors, e.g. a year of high rainfall andconsequent high productivity may allow the incursionof a new, highly competitive species, into the communitywhich may become dominant. Intersexual differencesin habitat use by dominant species have also beendemonstrated (Monamy, 1997).
In this study M. libycus was grouped with the psam-mophiles but was trapped at low density in a variety ofhabitat types (see Table 3). Meriones libycus is a domi-nant species and because of its size and high-densitycolonies it is possibly restricted to areas of high produc-tivity, such as sand dunes with a high perennial cover. Itis therefore envisaged that a highly productive yearwould allow this species to be more generalist in itshabitat selection than previously recorded. The ¯ex-ibility of generalist species to exploit different habitattypes is an adaptation to the highly unpredictable andvariable desert ecosystems. The generalist species seemto be the most successful in terms of their geographicaldistributions, e.g. Jaculus jaculus, M. libycus andM. crassus are all widely distributed throughout thesouth Palaearctic desert belt (Niethammer, 1985).
Mechanisms of coexistence
Desert rodent communities are complex and themechanisms behind their composition are not discrete(Kotler & Brown, 1988). Differences in habitatpreferences morphology and competitive ability havepreviously been reported to aid coexistence (Bowers &Brown, 1982; Reichman, 1984; Kotler, 1992, 1997). Inthe present study there seems to be evidence to supportthese three methods of resource partitioning as amechanism facilitating coexistence and thereforeimportant in determining community structure.
Habitat preferences
TWINSPAN and DCA derived rodent groups werecorrelated with the ®rst environmental axis (see Fig. 6),and this result was con®rmed statistically by CCA. The
grouping of rodents in relation to habitat types suggestsdistinct local habitat preferences. For example, bothanalyses suggested that A. russatus is restricted to harratand trapping studies have demonstrated that it is onlypresent on the upper-most basalt slopes. This preferencecould be due to the availability of suitable refugesaffording protection from predators and against theharsh environmental conditions (Hughes, Ward &Perrin, 1994). Similarly, G. nanus was only found insand dune habitats (Table 3), even though hammadawas available nearby. Meriones crassus tends to burrowon hammada and therefore may show a compromisein habitat selection between substrates suitable forburrowing, foraging and protection from predators.Jaculus jaculus may show a preference for a particularhabitat, i.e. hammada, owing to its bipedal locomotion,since this may well affect foraging economics andanti-predator behaviour (Lemen & Rosenzweig, 1978;Thompson, 1982).
Morphology
Body size and form have previously been associatedwith microhabitat use by desert rodents (J. S. Brownet al., 1994). Large bipedal species tend to be associatedwith open microhabitats, whereas smaller quadrupedsmore frequently occur in areas with cover. However,this does not seem to be the case for macrohabitatselection, which encompasses an array of microhabitattypes, demonstrated in this study (see Tables 3 & 4).
Within each derived rodent group there was consider-able size disparity and degree of specialization tohabitat type, which potentially facilitates coexistence(Bowers, 1982). For example, only one Meriones speciesis represented in each group (Fig. 3). Where there aretwo or more species from the same family there tends tobe either a variation in body weight or in specialization.The ®rst example of this is seen in group 4 fromTWINSPAN (Fig. 2), where G. nanus and G. cheesmanihave comparable body weights, but G. cheesmani hasspecialist adaptations to sandy habitats, i.e. hairy feet.The second example from TWINSPAN is seen inGroup 1, where G. dasyurus and G. henleyi both occur,but differ in body weight by over 100% (see Table 3).Size disparity is also evident within habitat classes (seeTables 3 & 4) with size differences following similarlogarithmic patterns of spacing as described by Bowers& Brown (1982). Gerbillus nanus and G. dasyurus, whichhave the most similar body size, did not inhabit thesame habitat class.
Competitive ability
Competition may also account for the fact that only oneMeriones species is represented in each group. Merionestristrami was only caught on a few occasions, and thisrarity could be the result of competition with otherMeriones species. Meriones tristrami was included in the
D. M. Scott and N. Dunstone490

petrophile group but may be exhibiting competitivedisplacement into a less productive habitat in the pre-sence of a more successful competitor. Acomys russatushas previously been reported as diurnal (Shkolnik,1971), but showed predominantly nocturnal behaviourduring this study. Although activity patterns can varyseasonally, a preference for nocturnal activity possiblyresulted from reduced competition with other Acomysspecies (Shkolnik, 1971), since no other Acomys specieswere captured during this study.
Multivariate analyses
Rodent-de®ned habitat types derived from TWINSPANcorrespond to ground-truthed descriptions of habitatsamples. The grouping of rodents generally followedwhat was expected from ®eld observations. The excep-tion was G. henleyi, which was categorized into thepetrophiles group, although trapping results indicatedthat it occurred more frequently and at higher density inhammada. The species has previously been recorded asdwelling in hammada (Krasnov et al., 1996) and itsgrouping by TWINSPAN was probably because of itsoccurrence in marab habitats and its associations withspecies occurring in rocky habitats, e.g. G. dasyurus,which was also trapped in marab and hammada.Gerbillus henleyi and M. tristrami were not capturedfrequently, and further evidence is required to groupthese species more precisely.
As species compositions were compared usingstandardized measurements between rather than withinsites, integration of the two methodologies to estimatedensities was considered justi®able for this analysis.Additionally, J. jaculus grouping and ordination isapproximately where they would be envisaged from the®eld results.
The TWINSPAN grouping (Figs 2 & 3) indicatedthat community structuring is not simply based onenvironmental factors which classify habitat types(Table 2), but is complex in nature with a variety ofin¯uential factors. Anomalies in TWINSPAN resultsemphasize the need for temporal replicates when deter-mining community composition. The speciescomposition of communities and the mechanisms thatdetermine them are dynamic. Temporal variations mayresult from the resources available at that time, whichcould be a consequence of environmental conditions at,or before the time of sampling (Ellis et al., 1997).Temporal constancy of rodent communities seems to bemore prevalent in areas that have specialist species (e.g.harrat) rather than generalist species (e.g. hammada). Inhabitat types which do not possess a discrete rodentfauna (e.g. marab) there tend to be greater ¯uctuationsin community composition (Fig. 5). A longer termstudy is required to further investigate the temporal¯uctuations in rodent composition.
Environmental variables are usually highly intercorre-lated and so it is dif®cult or impossible to separate outindependent effects (Ter Braak, 1988). The CANOCO
program tests for co-linearity so variables showinglinear relationships can be excluded from further ana-lyses. The habitat groupings in DCA shows that somesites have a variable composition of rodent species andcannot be associated exclusively with any particularde®ned rodent group. Habitats that are not located onthe extremes of the environmental axes (see Fig. 5)tend to have a higher species richness, e.g. marabsand hammadas. The CCA showed similar patterns ofdistribution of species and sites as the DCA andindicated the most important environmental variablescontributing to their distribution.
Comparisons with previous studies of rodent communitycomposition
A comparison between studies conducted in Israel(Krasnov et al., 1996) and the present work suggestsdifferences in habitat selection by G. nanus andG. dasyurus. These differences could be accounted forby species divergence into various habitat types in¯u-enced by several factors, e.g. availability of differenthabitats.
In Israel, Gerbillus nanus was recorded to be a gener-alist species found on hammada and in wadis, whereasin the present study it was only found in sandy habitats.This habitat shift may have been caused by potentialcompetition with a similar sized species, G. dasyurus,which, in the Badia is more adapted to basaltic habitats,such as wadis. Species showing such divergence seem topossess adaptations to the habitats they occupy, forexample, in Jordan, G. dasyurus tended to be morepetrophilic than in Israel. The Jordanian specimensseem to be a darker morph which could be anadaptation to inhabiting a black basalt habitat. Thepredominant colour of the surrounding habitat has beensuggested to be a potentially important factor in theformation of `substrate races' (Harrison & Seaton-Browne, 1969). Acomys russatus lewisi, found in thisstudy, has been previously identi®ed as a subspeciesspeci®c to basaltic habitats (Atallah, 1967). The differ-ences in community composition between studies re¯ectthe adaptations of species to a different range of habitattypes and competition between different species.
Some similarities and differences between the rodentcommunities found in these two deserts could beexplained by their biogeographic histories, e.g.G. gerbillus, a strictly psammophlious species in Israel(Krasnov et al., 1996), does not occur in north-eastBadia. Here its niche seems to be ®lled by anotherpsammophile of similar body size, G. cheesmani.
Granivorous rodent communities show species con-vergence towards similar niches and coexisting speciesseem to show comparable differences in morphology,body size and foraging behaviour (Brown et al., 1979).This is thought to be a mechanism facilitating coexis-tence by reducing interspeci®c competition. Stone et al.(1996) compared deserts across the American Southwestand identi®ed competition as an underlying mechanism
491Communities of desert rodents

in structuring local rodent communities. Competitionwas also evident in the present study, but to what extentthis in¯uenced community composition has yet to beinvestigated. We noted considerable similarity in thepartitioning of body sizes and morphology, to thatfound by Brown (1975) in his comparisons of NorthAmerican deserts. Body sizes tend to fall into discretecategories in the range from 5 to 120 g (Table 3). Ineach category there are usually only one or two repre-sentatives; where there are two or more representatives,there seems to be variation in either morphology,locomotion (bipedal and quadrupedal) or in habitatpreferences. Although foraging behaviour has not yetbeen investigated for rodents in the north-east Badia ofJordan, it is likely that body size and morphology willin¯uence foraging behaviour leading to similar patternsin variation.
Asian desert-living species show a greater degree ofdivergence and specialization between bipedal andquadrupedal rodents than those from North Americandeserts, although feeding modes were similar on bothcontinents (Shenbrot et al., 1994). Rodent diversity inAsian and American deserts was similar to that found inthe present study (2±4 species per site), although ourresults failed to show a correlation between rodentdiversity and biomass.
For rodent communities in general, vegetation coverand related productivity seem to be the main environ-mental determinants of community composition (Bianet al., 1994; Adler, 1995; Oguge, 1995). In the presentstudy and other desert systems (south Bukhara desert,Uzbekistan, Shenbrot, 1992; Asian deserts, Shenbrotet al., 1994; Anix desert, China, Liu et al., 1994; Negevhighlands, Israel, Krasnov et al., 1996) soil structure andcontent have also been shown to be important factors.
Acknowledgements
The research was funded by the Darwin Initiative, TheMarion Zunz Award, The Trevelyan Trust and TheBritish and Foreign School Society. We are grateful tothe Badia Research and Development Project, HigherCouncil for Science & Technology, Royal Society forthe Conservation of Nature in Jordan and the Centrefor Overseas Research and Development (Durham Uni-versity), Royal Geographical Society and the Instituteof British Geographers for providing logistical support.Our gratitude extends especially to Professor BrianHuntley for advice on multivariate analysis and statis-tical techniques, Professor Peter Evans for invaluablecomments on the draft manuscript and continualsupport and advice. Dr Tusi Butter®eld and Dr RusHoelzel also commented on the manuscript. We wouldalso like to thank the research assistants who partici-pated with the survey: Tom Maddox, Kate Lister, AnneBurrows, Mike O' Connor, Rachel Freer and KarenTaylor. Thanks goes to Kevin Brown and the Darwinvegetation team for supplying information and con-®rming identi®cations. We also thank the Harrison
Zoological Museum, Sevenoaks, Kent, for con®rmingidenti®cation and training. Finally we thank the BorderPolice at Hashad and Hazim, and all the Bedouin wemet for their over¯owing hospitality.
REFERENCES
Abramsky, Z. (1988). The role of habitat and productivity instructuring desert rodent communities. Oikos 52: 107±114.
Adler, G. H. (1995). Habitat relations within lowland grasslandrodent communities in Taiwan. J. Zool. (Lond.) 237: 563±576.
Atallah, S. I. (1967). A new species of spiny mouse (Acomys) fromJordan. J. Mammal. 48(2): 258±261
Atallah, S. I. (1977). Mammals of the Eastern Mediterraneanregion; their ecology, systematics and zoogeographical relation-ships. Saeugetierkd. Mitt. 25: 241±320
Beatley, J. C. (1969). Dependence of desert rodents on winterannuals and precipitation. Ecology 50: 721±724.
Behr, H. (1994). Communities of coexisting species of the water-beetle genus Hydroporus in two northern German areas(Coleoptera, Dytiscidae). Int. Rev. Gesamten Hydrobiol. 79:337±355.
Bian, J., Fan, N., Jing, Z. & Shi, Y. (1994). Studies on thesuccessive relation between small mammal community andplant community in alpine meadow. Acta Theriol. 14: 209±216.
Blair, W. F. (1942). Home ranges and populations of the meadowvole in southern Michigan. J. Wildl. Manage. 4: 141±161.
Bowers, M. A. (1982). Foraging behaviour in heteromyid rodents:®eld evidence for resource partitioning. J. Mammal. 63:361±367.
Bowers, M. A. (1988). Seed removal experiments on desertrodents ± The microhabitat by moonlight effect. J. Mammal.69: 201±204.
Bowers, M. A. & Brown, J. H. (1982). Body size and coexistencein desert rodents ± chance or community structure. Ecology 63:391±400.
Bowers, M. A. & Doley, J. L. (1993). Predation hazard and seedremoval by small mammals ± microhabitat versus patch scaleeffects. Oecologia 94: 247±254.
Brown, J. H. (1975). Geographical ecology of desert rodents. InEcology and evolution of communities. Cody, M. L. &Diamond, J. M. (Eds). Cambridge, MA: Harvard UniversityPress.
Brown, J. H. & Lieberman, G. (1973). Resource utilization andcoexistence of seed-eating rodents in sand dune habitats.Ecology 54: 788±797.
Brown, J. H., Reichman, O. J. & Davidson, D. W. (1979).Granivory in desert ecosystems. Annu. Rev. Ecol. Syst. 10:201±227.
Brown, J. S. Kotler, B. P. & Mitchell, W. A. (1994). Foragingtheory, patch use and the structure of a Negev desert granivorecommunity. Ecology 75(8): 2286±2300.
Church, M. A., Mclean, D. G. & Wolcott, J. F. (1987). River bedgravel: sampling and analysis. In Sediment transport in gravel-bed rivers. Thorne, C. R., Bathurst, J. C. & Hey, R. D. (Eds).Chichester: Wiley.
Dutton, R. W., Clark, J. I. & Battikhi, A. M. (1998). Arid landresources and their management. London: Kegan Paul.
Ellis, L. M., Crawford, C. S. & Morales, M. C. Jr (1997). Rodentcommunities in native and exotic riparian vegetation in theMiddle Rio Grande Valley of central New Mexico. Southwest.Nat. 42: 13±19.
FAO (1977). Guidelines for soil pro®les description. Rome: Foodand Agricultural Organisation of the United Nations,
Happold, D. C. D. (1967). Biology of the jerboa, Jaculus jaculusbutleri (Rodentia, Dipodidae), in the Sudan. J. Zool. (Lond.)151: 257±275.
D. M. Scott and N. Dunstone492

Harris, J. H. (1986). Microhabitat segregation in 2 desert rodentspecies ± the relation of prey availability to diet. Oecologia(Berl.) 68: 417±421.
Harrison, D. L. & Bates, P. (1991). The Mammals of Arabia. 2ndedn. Sevenoaks: Harrison Zoological Museum.
Harrison, D. L. & Seaton-Browne, C. J. (1969). The in¯uence ofsoil colour on subspeciation of mammals in eastern Arabia.Zool. J. Linn. Soc. 48: 467±470
Hatough-Bouran, A. M. & Al-Eisawi, D. (1990). Rodent ecologyin the Shaumari Wildlife Reserve of Jordan. Dirasat Ser. BPure Appl. Sci. 17(3): 62± 85.
Heske, E. J., Brown, J. H. & Mistry, S. (1994). Long-termexperimental study of a Chihuahuan desert rodent community± 13 years of competition. Ecology 75: 438±445.
Hill, M. O. (1973). Diversity and evenness: a unifying notationand its consequences. Ecology 54: 427±432.
Hill, M. O. (1979). DECORANA: a FORTRAN program fordetrended correspondence analysis and reciprocal averaging.Ithaca, NY: Section of Ecology and Systematics, Cornell Uni-versity.
Hughes, J. J., Ward, D. & Perrin, M. R. (1994). Predation riskand competition affect habitat selection and activity of Namibdesert gerbils. Ecology 75: 1397±1405.
Kelt, D. A., Brown, J. H., Heske, E. J., Marquet, P. A., Morton,S. R., Reid, J. R. W., Rogovin, K. A. & Shenbrot, G. (1996).Community structure of desert small mammals ± comparisonsacross four continents. Ecology 77: 746±761.
Kirk, A. (1998). A synthesis of climatic data with speci®c interestin the precipitation record. In Arid land resources and theirmanagement. Dutton, R. W., Clark, J. I. & Battikhi, A. M(Eds). London: Kegan Paul.
Kotler, B. P. (1972). Behavioural resource depression and decay-ing perceived risk of predation in two species of coexistinggerbils. Behav. Ecol. Sociobiol. 30: 239±244.
Kotler, B. P. (1984). Predation risk and the structure of desertrodent communities. Ecology 65: 689±701.
Kotler, B. P. (1997). Patch use by gerbils in a risky environment:manipulating food and safety to test four models. Oikos 78:274±282.
Kotler, B. P. & Brown, J. S. (1988). Environmental heterogeneityand the coexistence of desert rodents. Annu. Rev. Ecol. Syst. 19:281±307.
Krasnov, B. Shenbrot, G., Khokhlova, I. & Ivanitskaya, E.(1996). Spatial patterns of rodent communities in the RamonErosion Cirque, Negev Highlands, Israel. J. Arid Environ. 32:319±327.
Lemen, C. A. & Rozenzweig, M. L. (1978). Microhabitat selectionin two species of heteromyid rodents. Oecologia (Berl.) 33:127±135.
Liu, D., Liu, N., Song, Z., Pan, F. & Ning, R. (1994). A detrendedcorrespondence analysis of community structure of desertrodent and environmental factors in Anxi. Acta Theriol. Sin.12: 108±116.
Mackie, A. S. Y., Parmiter, C. & Tong, L. K. Y. (1997). Distribu-tion and diversity of Polychaeta in the southern Irish Sea. Bull.Mar. Sci. 60: 467±481.
Manly, B. F. (1990). Randomization and Monte Carlo methods inbiology. London: Chapman & Hall.
Monamy, V. (1997). Sexual differences in habitat use by Rattuslutreolus (Rodentia: Muridae) the emergence of patterns innative rodent community structure. Aust. Mammal. 20: 43±48.
Niethammer, J. (1985). Rodent distributions in the Middle East.In Proceedings of the symposium on the fauna and zoogeographyof the Middle East. Mainz Krupp, F., Schneider, W. & Kinzel-bach, R. (Eds). Wiesbaden: Ludwig Verlag.
Oguge, N. O. (1995). Diet, seasonal abundance and microhabitatsof Praomys (Mastomys) natalensis (Rodentia: Muridae) andother small rodents in a Kenyan sub-humid grassland commu-nity. Afr. J. Ecol. 33: 211±223.
Price, M. V. (1978). The role of microhabitat in structuring desertrodent communities. Ecology 59: 910±921.
Price, M. V. & Heinz, K. M. (1984). Effects of body size, seeddensity and soil characteristics on rates of seed harvest byheteromyid rodents. Oecologia (Berl.) 61: 420±425.
Reichman, O. J. (1984). Spatial and temporal variation of seeddistributions in Sonoran desert soils. J. Biogeogr. 11: 1±11.
Roderiguez, T., Trigo, D. & Cosin, D. D. (1997). Biogeographicalzonation of the western Iberian peninsula on the basis of thedistribution of earthworm species. J. Biogeogr. 24: 893±901.
Rogovin, K. A. & Surov, A. V. (1990). Morpho-ecologicalstructure of desert rodent communities in Central Asia andSouth-western North America ± a multivariate approach. ActaTheriol. 35: 225±239.
Rogovin, K., Shenbrot, G. & Surov, A. (1991). Analysis of spatialorganisation of a desert rodent community in Bolson DeMapimi, Mexico. J. Mammal. 72: 347±359.
Rosenzweig, M. L. & Winakur, J. (1969). Population ecology ofdesert rodent communities: habitats and environmentalcomplexity. Ecology 50: 558±572.
Ruse, L. P. (1996). Multivariate techniques relating macroinverte-brate and environmental data from a river catchment. WaterRes. 30: 3017±3024.
Safriel, U. N., Ayal, Y., Kotler, B. P., Lubin, Y., Olsvigwhittaker,L. & Pinshow, B. (1989). What's special about desert ecology ±introduction. J. Arid Environ. 17: 125±130.
Samways, M. & Steytler, N. S. (1996). Dragon¯y (Odonata)distribution patterns in urban forest landscapes and recommen-dations for riparian management. Biol. Conserv. 78: 279±288.
Searight, A. (1987). Some records of mammals from North-Eastern Jordan. In Proceedings of the symposium on the faunaand zoogeography of the Middle East. Mainz Krupp, F.,Schneider, W. & Kinzelbach, R. (Eds). Wiesbaden: LudwigVerlag.
Shenbrot, G. I. (1992). Spatial structure and niche patterns of arodents community in the South Bukhara desert (Middle Asia).Ecography 15: 347±357.
Shenbrot, G. I. & Rogovin, K. A. (1995). Temporal variation inspatial-organization of a rodents community in the Southwes-tern Kyzyl-Kum desert (Middle Asia). Ecography 18: 370±383.
Shenbrot, G. I., Rogovin, K. A. & Heske, E. J. (1994). Compar-ison of niche-packing and community organization in desertrodents in Asia and North-America. Aust. J. Zool. 42: 479±499.
Shkolnik, A.(1971). Diurnal activity in a small desert rodent. Int.J. Biometrics 15: 115±120
Soetaert, K., Vincx, M., Wittoeck, J. & Tulkens, M. (1995).Meiobenthic distributions and nematode community structurein ®ve European estuaries. Hydrobiologia 31: 185±206.
Stone, L., Dayan, T. & Simberloff, D. (1996). Community-wideassembly patterns unmasked ± the importance of speciesdiffering geographical ranges. Am. Nat. 148: 997±1015.
Taggart, J. B. (1994). Ordination as an aid in determiningpriorities for plant community protection. Biol. Conserv. 8:135±141.
Ter Braak, C. J. F. (1987a). Ordination. In Data analysis incommunity and landscape ecology. Jongman, R. H. G., TerBraak, C. J. F. & Van Tongeren, D. F. R. (Eds). Wageningen:Pudoc.
Ter Braak, C. J. F. (1987b). The analysis of vegetation±environ-ment relationships by canonical correspondence analysis. Vege-tatio 64: 69±77.
Ter Braak, C. J. F. (1988). CANOCO ± A FORTAN program forcanonical community ordination by partial detrended canonicalcorrespondences analysis, principal component analysis and re-dundancy analysis. Wageningen: TNO Institute of AppliedComputer Science.
Thompson, S. D. (1982). Microhabitat utilization and foragingbehaviour of bipedal and quadrupedal heteromyid rodents.Ecology 63: 1303±1312.
493Communities of desert rodents

Yajun, Z., Tinzheng, W., Jingang, L., Yue, Z., Fadao, T. &Juxiang, Z. (1997). Dynamics of rodent communities in agricul-tural area of Loess Plateau in the west of Henan Province:primary analyses for the pattern on temporal and spatial scale.Acta Theriol. Sin. 17: 197±203.
Vanzyl, P. & Robertson, B. L. (1991). Seasonal and species-environmental succession on an eastern cape rocky shore.S. Afr. J. Sci. 87: 327±330.
D. M. Scott and N. Dunstone494




















![Public Sphere and Civil Society in Jordan: "Jordan First" as a Case Study (2006) [Hebrew]](https://img.pdfslide.net/doc/110x75/631531d1511772fe45103fba/public-sphere-and-civil-society-in-jordan-jordan-first-as-a-case-study-2006.jpg)








