Embed Size (px)
Citation preview

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 31 (2006) 713 – 722
EphrinB3 regulates cell proliferation and survival in
adult neurogenesis
Jerome Ricard, Jessica Salinas, Lissette Garcia, and Daniel J. Liebl*
The Miami Project to Cure Paralysis and Department of Neurosurgery, University of Miami School of Medicine, 1095 NW 14th Terrace,
R-48, Miami, FL 33136, USA
Received 10 August 2005; revised 7 December 2005; accepted 2 January 2006
Available online 17 February 2006
Interactions between ephrins and their receptors have been implicated
in many processes during central nervous system development. In the
adult, ephrins and Eph receptors have been implicated in controlling
cell proliferation and neuroblast migration, although there is no direct
evidence for the role of ephrinB3 in these functions. In addition,
activation of Eph receptors has been shown to regulate transduction
pathways important in cell cycle control as well as cell death. We show
that ephrinB3 contributes to the control of cell proliferation and
survival in the adult subventricular zone (SVZ). EphrinB3�/� mice
exhibit a significant increase in dividing cells along the lateral ventricle,
and altered expression of proteins involved in cell cycle regulation.
Gain-of-function approach by infusing soluble ephrinB3-Fc molecules
in ephrinB3�/� can suppress cell proliferation to wild type levels. At the
same time, ephrinB3 also regulates cell survival as greater numbers of
cells die in the SVZ of ephrinB3�/� mice. Together, our results suggest
that ephrinB3 negatively regulates cell cycle progression and cell
apoptosis in the adult subventricular zone.
D 2006 Elsevier Inc. All rights reserved.
Keywords: Ephrin B3; Eph receptors; Stem cells; Subventricular zone;
Apoptosis; Proliferation
Introduction
The subventricular zone (SVZ) is a principle region of
neurogenesis in the adult rodent brain (Alvarez-Buylla and
Garcia-Verdugo, 2002). In normal physiological conditions, neural
stem/progenitor cells in the SVZ give rise to intermediate
progenitors, also named transit-amplifying precursors, which in
turn differentiate into neuroblasts that are destined for the olfactory
bulb (OB). In the OB, these migrating neuroblasts terminally
differentiate into interneurons (Luskin, 1993; Betarbet et al., 1996).
The mechanisms that govern these events are tightly regulated
1044-7431/$ - see front matter D 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.mcn.2006.01.002
* Corresponding author. Fax: +1 305 243 3914.
E-mail address: [email protected] (D.J. Liebl).
Available online on ScienceDirect (www.sciencedirect.com).
during neurogenesis from stem cell to neuron. In particular, to
ensure that there is an adequate pool of cells at each stage of
differentiation, proliferation within these stages requires specific
environmental and intracellular signaling mechanisms. In doing so,
the stem/progenitor cells generate the intermediate progenitors
while maintaining their own pool. The intermediate precursors may
use repeated cell divisions to expand their numbers and thus
greatly amplify the number of neuroblasts produced. Further
divisions within the neuroblast population during migration
contribute to the generation of a large amount of new neurons
(Alvarez-Buylla et al., 2001).
Cell proliferation in the SVZ was shown to be increased by a
variety of growth factors and hormones, including epidermal
growth factor (EGF), fibroblastic growth factor (FGF), vascular
endothelial growth factor (VEGF) and thyroid hormone (Doetsch
et al., 2002a,b; Jin et al., 2002, 2003; Lemkine et al., 2005). Other
molecules involved in the control of neural precursors prolifer-
ation include enhancers such as h1-integrins and retinoic acid
(Leone et al., 2005; Wang et al., 2005) and repressors such as neu-
rofibromin (Dasgupta and Gutmann, 2005). There is a tight link
between cell cycle control and neurogenesis (Ohnuma and Harris,
2003), which is further supported by a reduction in cell proli-
feration and neuroblast generation in mice lacking expression of
the E2F1 transcription factor (Cooper-Kuhn et al., 2002), and by
an increase in the number of proliferating precursors in p107�/�
mice (Vanderluit et al., 2004). An increase in proliferation in the
SVZ is also found in p27Kip1�/� mice and is coupled to a decrease
in neuroblast generation (Doetsch et al., 2002a,b). Similarly, in rats,
a decrease in p27Kip1 expression in the SVZ following a stroke
coincides with an increased cell proliferation (Zhang et al., 2004).
In addition, cell death also controls cell numbers in the SVZ,
probably through the release of yet non-identified diffusible factors
(Agasse et al., 2004). Finally, physiological regulation may also be
provided by sensory signals originating from the OB, where
axotomy-induced deafferentation of the OB leads to an increase
in proliferation in the SVZ (Mandairon et al., 2003). This increase
in proliferation is coupled to an up-regulation of cell death.
Conversely, others have reported decreased proliferation 3 months

J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722714
after bulbectomy (Kirschenbaum et al., 1999). It is still notable that
OB inputs do not appear to be essential for neurogenesis to occur in
the SVZ, suggesting that local cues and not diffusible OB factor
may be required to regulate neurogenic events.
One group of molecules that has been recently implicated in
proliferation of stem/progenitor cells in the SVZ is the ephrin
family and their Eph receptors. Intraventricular infusions of
ephrinB2-Fc or EphB2-Fc molecules lead to increases in the
number of astrocytes (some of which may be the stem/progenitor
cells) and transit-amplifying precursors, as well as a decrease in the
number of neuroblasts after EphB2-Fc injections. RT-PCR experi-
ments show that ephrinB2 and ephrinB3 ligands, as well as EphB1,
EphB2, EphB3 and EphA4 receptors are expressed in the SVZ
(Conover et al., 2000). Furthermore, EphB2 directly activates the
proliferation of SVZ precursors in vitro (Katakowski et al., 2005).
Recently, the A-class was implicated as well in the control of
neurogenesis: ephrinA2 and EphA7 were identified as negative
regulators of neural precursors proliferation (Holmberg et al.,
2005). However, ephrinA1 does not affect telencephalon embry-
onic progenitors proliferation, but rather directs cells towards a
neuronal fate (Aoki et al., 2004). Ephrins are membrane-bound
ligands and tethered to the cell surface, by either a glycosylphos-
phatidylinositol (GPI)-anchor (A-class) or transmembrane domain
(B-class). Eph receptors, which belong to the largest family of
receptor tyrosine kinases, are also subdivided into two A- and B-
classes, based on their preferential binding for one particular group
of ligands. One exception is EphA4 as it binds ligands from both
subclasses. Therefore, receptor activation by ligand binding is a
direct result of cell–cell contact, and interactions between ephrins
and Eph receptors can result in bidirectional signaling from both
the ligand and receptor (Flanagan and Vanderhaeghen, 1998).
Although the binding between ligands and receptors is considered
very promiscuous, specific ligand–receptor interactions exist
within each subclass (Blits-Huizinga et al., 2004).
Here, we examined a novel role for ephrinB3 in neurogenic
proliferation of the SVZ. Examination of gene-targeted knockout
mice for ephrinB3 revealed a significant increase in the number of
proliferating stem/progenitor cells in the SVZ. We also observed
significant increases in cell cycle activators and decreases in cell
cycle inhibitors in the absence of ephrinB3. An increase in
apoptosis in neurogenic regions is associated with the increase in
proliferation in the absence of ephrinB3. We propose that ephrinB3
functions as a negative regulator of cell proliferation and cell
apoptosis in the SVZ during adult neurogenesis.
Results
Expression of B-class ephrins in the adult SVZ and RMS regions
Neurogenesis continues to persist in the subventricular zone into
adulthood, where stem/progenitor cells give rise to neuroblasts that
migrate through the rostral migratory stream (RMS) to become
interneurons in the olfactory bulb. Previous reports have implicated
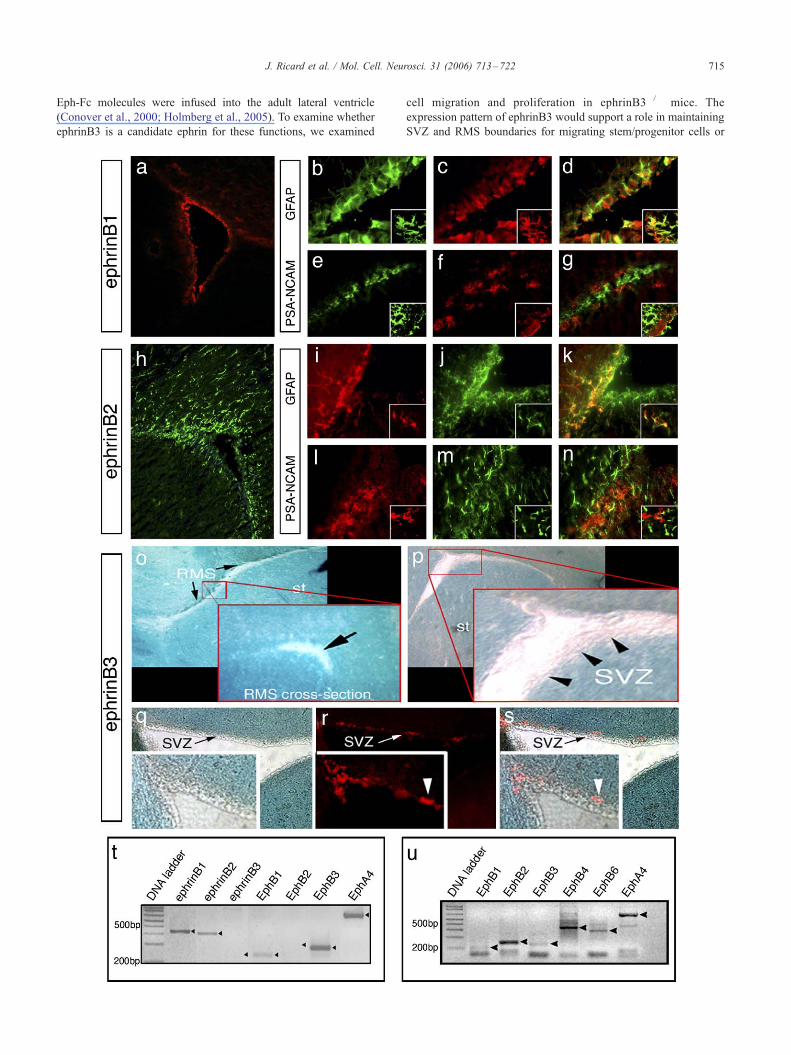
Fig. 1. EphrinB ligands are expressed in the adult subventricular zone. EphrinB1
GFAP (green, b, d) but not PSA-NCAM (green, e, g). EphrinB2 (green, h, j, m)
localizes with GFAP (red, i, k) but not PSA-NCAM (red, l, n). Insets: high-mag
staining in ephrinB3hgal mice) is excluded from the RMS (o, sagittal section; inse
devoid of ephrinB3 contains PSA-NCAM-stained neuroblasts (red, r, s), which see
receptors detected by RT-PCR in cultured neurospheres. (u) Expression of Eph re
a role for B-class ephrins in the adult rodent SVZ, however, it is
unclear which ephrin(s) may be involved (Conover et al., 2000). To
further elucidate the role of ephrins in the SVZ, we initially
examined the expression of this family of ligands in the subven-
tricular zone. All three ephrinBs were observed in the SVZ or
surrounding tissues, where ephrinB1 and ephrinB2 were expressed
on cells residing in the SVZwhile ephrinB3 was expressed in tissues
surrounding the SVZ and RMS (Fig. 1). In particular, ephrinB1 was
observed on cells that resided in the SVZ along the striatal, septal,
and cortical sides (Fig. 1a). In addition, ephrinB1 was found to be
co-expressed with GFAP (Figs. 1b–d), an astrocytic marker found
also in stem/progenitor cells in the SVZ, but not PSA-NCAM (Figs.
1e–g). EphrinB2 was expressed in cells that resided in and around
the SVZ and corpus callosum (CC) (Fig. 1h). Like ephrinB1,
ephrinB2 was also co-expressed with GFAP and not PSA-NCAM;
however, some of the GFAP-expressing cells have a ramified
appearance that likely represents mature astrocytes while other
GFAP-expressing cells appeared to be morphologically similar to
stem/progenitor cells (Figs. 1i–n). In addition, we cannot rule out
the possibility that ephrinB1 and ephrinB2may also be expressed on
ependymal cells, since cross-reactivity with anti-GFAP in such cells
antibodies has been shown previously (Doetsch et al., 2002a,b). Co-
expression of ephrinB1 and ephrinB2 with GFAP was confirmed
using confocal microscopy (Figs. 1b–g insets, i–n insets). To
examine ephrinB3 expression, we took advantage of a transgenic
knock-in mouse where h-galactosidase (ephrinB3hgal) replaces thecytoplasmic domain. X-gal staining was observed throughout the
brain, including the cortex, striatum, septum, and CC; however, we
did not observe staining within the SVZ or RMS (Figs. 1o, p). Figs.
1q–s represent an overlay image where PSA-NCAM-expressing
neuroblasts where found to mainly reside along a border of
ephrinB3 expression in the SVZ. To confirm whether ephrinB1
and ephrinB2 but not ephrinB3 are present in stem/progenitor cell
populations, we examined mRNA expression in purified neuro-
spheres. RT-PCR analysis using RNA from cultured neurospheres
generated from adult SVZ cells showed expression of ephrinB1 and
ephrinB2 but not ephrinB3, supporting our immunohistochemical
observations (Fig. 1t). These findings also support the presence of
ephrinB1 and ephrinB2 in the stem/progenitor cell population. We
also found that EphB1, EphB3, and EphA4 receptors were all
expressed in the neurospheres, which specifically contain stem/
progenitor cell populations. Further analysis of carefully dissected
SVZ tissue revealed that in addition to EphB1, EphB3, and EphA4
mRNA expression, EphB2, EphB4, and EphB6 mRNA transcripts
were also localized to tissues in or closely associated to the SVZ
(Fig. 1u). These studies demonstrate that multiple ephrins and Eph
receptors are present in and/or around the SVZ, and may play
important roles in regulating adult neurogenesis.
Deletion of ephrinB3 results in cell proliferation and not migration
abnormalities in the subventricular zone
Previous reports have implicated ephrins in regulating cell
migration and proliferation in the adult SVZ when ephrin-Fc or
(red, a, c, f) is expressed in the walls of the ventricle, and co-localizes with
is expressed along the ventricle and in the corpus callosum, and also co-
nification confocal images. Expression of ephrinB3 (identified using X-gal
t: cross-section of the RMS) and SVZ (p, q). The area lining the ventricle
m to align along the ephrinB3 border (s). (t) Expression of ephrins and Eph
ceptors detected by RT-PCR in carefully dissected SVZ tissues.
J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722 715
Eph-Fc molecules were infused into the adult lateral ventricle
(Conover et al., 2000; Holmberg et al., 2005). To examine whether
ephrinB3 is a candidate ephrin for these functions, we examined
cell migration and proliferation in ephrinB3�/� mice. The
expression pattern of ephrinB3 would support a role in maintaining
SVZ and RMS boundaries for migrating stem/progenitor cells or
J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722716
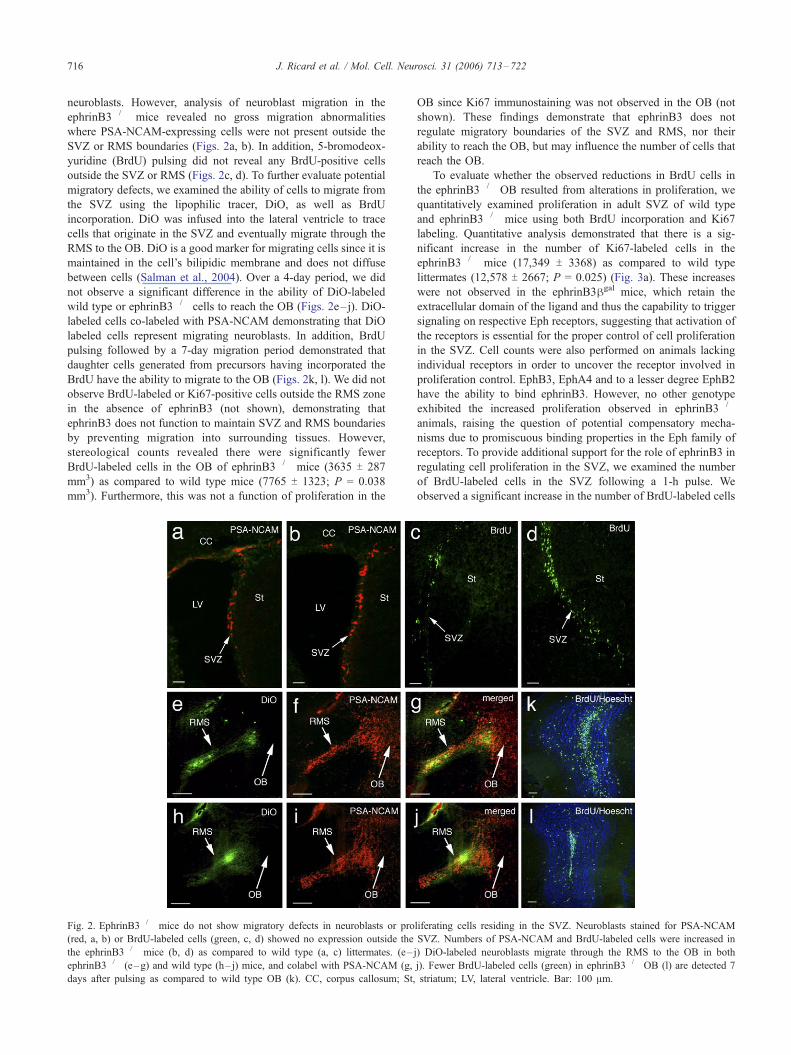
neuroblasts. However, analysis of neuroblast migration in the
ephrinB3�/� mice revealed no gross migration abnormalities
where PSA-NCAM-expressing cells were not present outside the
SVZ or RMS boundaries (Figs. 2a, b). In addition, 5-bromodeox-
yuridine (BrdU) pulsing did not reveal any BrdU-positive cells
outside the SVZ or RMS (Figs. 2c, d). To further evaluate potential
migratory defects, we examined the ability of cells to migrate from
the SVZ using the lipophilic tracer, DiO, as well as BrdU
incorporation. DiO was infused into the lateral ventricle to trace
cells that originate in the SVZ and eventually migrate through the
RMS to the OB. DiO is a good marker for migrating cells since it is
maintained in the cell’s bilipidic membrane and does not diffuse
between cells (Salman et al., 2004). Over a 4-day period, we did
not observe a significant difference in the ability of DiO-labeled
wild type or ephrinB3�/� cells to reach the OB (Figs. 2e–j). DiO-
labeled cells co-labeled with PSA-NCAM demonstrating that DiO
labeled cells represent migrating neuroblasts. In addition, BrdU
pulsing followed by a 7-day migration period demonstrated that
daughter cells generated from precursors having incorporated the
BrdU have the ability to migrate to the OB (Figs. 2k, l). We did not
observe BrdU-labeled or Ki67-positive cells outside the RMS zone
in the absence of ephrinB3 (not shown), demonstrating that
ephrinB3 does not function to maintain SVZ and RMS boundaries
by preventing migration into surrounding tissues. However,
stereological counts revealed there were significantly fewer
BrdU-labeled cells in the OB of ephrinB3�/� mice (3635 T 287
mm3) as compared to wild type mice (7765 T 1323; P = 0.038
mm3). Furthermore, this was not a function of proliferation in the
Fig. 2. EphrinB3�/� mice do not show migratory defects in neuroblasts or pro
(red, a, b) or BrdU-labeled cells (green, c, d) showed no expression outside the
the ephrinB3�/� mice (b, d) as compared to wild type (a, c) littermates. (e–
ephrinB3�/� (e–g) and wild type (h– j) mice, and colabel with PSA-NCAM (g,
days after pulsing as compared to wild type OB (k). CC, corpus callosum; St,
OB since Ki67 immunostaining was not observed in the OB (not
shown). These findings demonstrate that ephrinB3 does not
regulate migratory boundaries of the SVZ and RMS, nor their
ability to reach the OB, but may influence the number of cells that
reach the OB.
To evaluate whether the observed reductions in BrdU cells in
the ephrinB3�/� OB resulted from alterations in proliferation, we
quantitatively examined proliferation in adult SVZ of wild type
and ephrinB3�/� mice using both BrdU incorporation and Ki67
labeling. Quantitative analysis demonstrated that there is a sig-
nificant increase in the number of Ki67-labeled cells in the
ephrinB3�/� mice (17,349 T 3368) as compared to wild type
littermates (12,578 T 2667; P = 0.025) (Fig. 3a). These increases
were not observed in the ephrinB3hgal mice, which retain the
extracellular domain of the ligand and thus the capability to trigger
signaling on respective Eph receptors, suggesting that activation of
the receptors is essential for the proper control of cell proliferation
in the SVZ. Cell counts were also performed on animals lacking
individual receptors in order to uncover the receptor involved in
proliferation control. EphB3, EphA4 and to a lesser degree EphB2
have the ability to bind ephrinB3. However, no other genotype
exhibited the increased proliferation observed in ephrinB3�/�
animals, raising the question of potential compensatory mecha-
nisms due to promiscuous binding properties in the Eph family of
receptors. To provide additional support for the role of ephrinB3 in
regulating cell proliferation in the SVZ, we examined the number
of BrdU-labeled cells in the SVZ following a 1-h pulse. We
observed a significant increase in the number of BrdU-labeled cells
liferating cells residing in the SVZ. Neuroblasts stained for PSA-NCAM
SVZ. Numbers of PSA-NCAM and BrdU-labeled cells were increased in
j) DiO-labeled neuroblasts migrate through the RMS to the OB in both
j). Fewer BrdU-labeled cells (green) in ephrinB3�/� OB (l) are detected 7
striatum; LV, lateral ventricle. Bar: 100 Am.
J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722 717
in the ephrinB3�/� mice (7295 T 653) as compared to wild type
mice (3847 T 1238; P = 0.044) (Fig. 3b). This increased
proliferation in the ephrinB3�/� mice correlated with an increase
in the number of neuroblasts present in the SVZ (Fig. 3c). We also
examined vimentin expression (a marker not specific for cell
proliferation but known to be up-regulated in germinal zones
(Alonso, 2001)), and found increased expression of vimentin in the
ephrinB3�/� mice as compared to wild type mice (Figs. 3d, e). In
contradiction to the reduction of BrdU-labeled cells in the OB,
these findings support the role of ephrinB3 in regulating cell
proliferation that results in increased numbers of neuroblasts in the
SVZ of ephrinB3�/� mice.
We also performed a primary neurosphere assay in order to
determine whether the increase in proliferation and neuroblasts
observed in the ephrinB3�/� animals could be linked to the
presence of greater numbers of progenitors. A defined number of
cells were plated from wild type and ephrinB3�/� SVZ and the
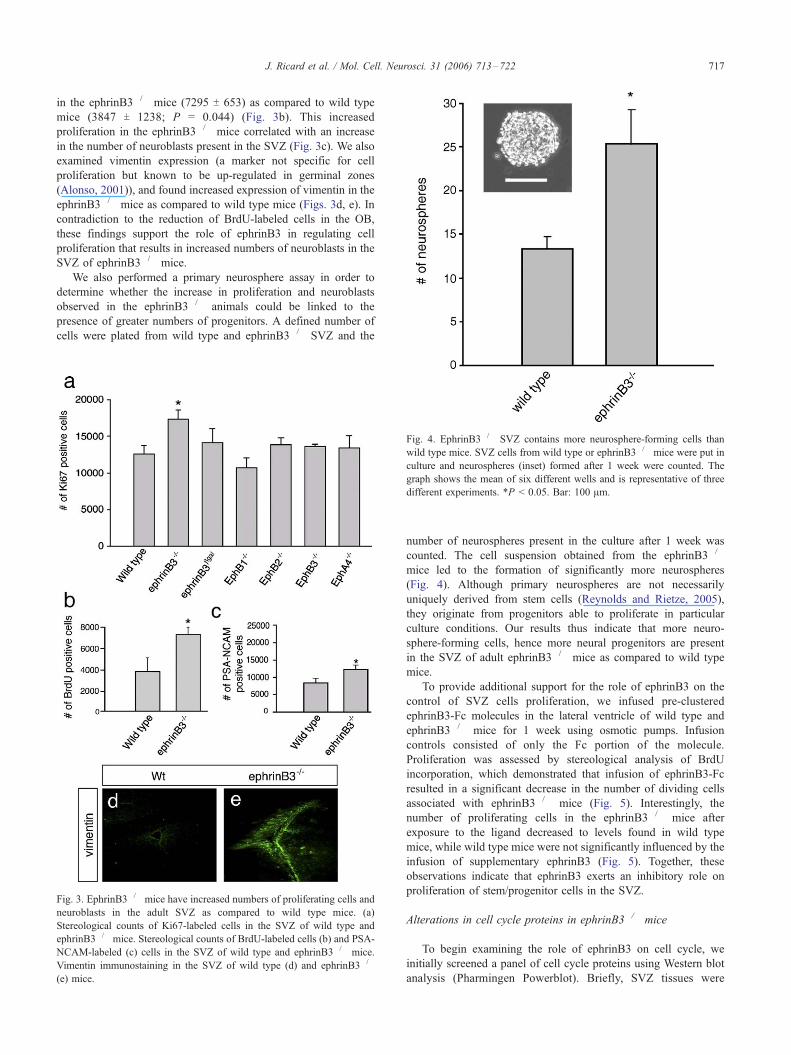
Fig. 3. EphrinB3�/� mice have increased numbers of proliferating cells and
neuroblasts in the adult SVZ as compared to wild type mice. (a)
Stereological counts of Ki67-labeled cells in the SVZ of wild type and
ephrinB3�/� mice. Stereological counts of BrdU-labeled cells (b) and PSA-
NCAM-labeled (c) cells in the SVZ of wild type and ephrinB3�/� mice.
Vimentin immunostaining in the SVZ of wild type (d) and ephrinB3�/�
(e) mice.
Fig. 4. EphrinB3�/� SVZ contains more neurosphere-forming cells than
wild type mice. SVZ cells from wild type or ephrinB3�/� mice were put in
culture and neurospheres (inset) formed after 1 week were counted. The
graph shows the mean of six different wells and is representative of three
different experiments. *P < 0.05. Bar: 100 Am.
number of neurospheres present in the culture after 1 week was
counted. The cell suspension obtained from the ephrinB3�/�
mice led to the formation of significantly more neurospheres
(Fig. 4). Although primary neurospheres are not necessarily
uniquely derived from stem cells (Reynolds and Rietze, 2005),
they originate from progenitors able to proliferate in particular
culture conditions. Our results thus indicate that more neuro-
sphere-forming cells, hence more neural progenitors are present
in the SVZ of adult ephrinB3�/� mice as compared to wild type
mice.
To provide additional support for the role of ephrinB3 on the
control of SVZ cells proliferation, we infused pre-clustered
ephrinB3-Fc molecules in the lateral ventricle of wild type and
ephrinB3�/� mice for 1 week using osmotic pumps. Infusion
controls consisted of only the Fc portion of the molecule.
Proliferation was assessed by stereological analysis of BrdU
incorporation, which demonstrated that infusion of ephrinB3-Fc
resulted in a significant decrease in the number of dividing cells
associated with ephrinB3�/� mice (Fig. 5). Interestingly, the
number of proliferating cells in the ephrinB3�/� mice after
exposure to the ligand decreased to levels found in wild type
mice, while wild type mice were not significantly influenced by the
infusion of supplementary ephrinB3 (Fig. 5). Together, these
observations indicate that ephrinB3 exerts an inhibitory role on
proliferation of stem/progenitor cells in the SVZ.
Alterations in cell cycle proteins in ephrinB3�/� mice
To begin examining the role of ephrinB3 on cell cycle, we
initially screened a panel of cell cycle proteins using Western blot
analysis (Pharmingen Powerblot). Briefly, SVZ tissues were
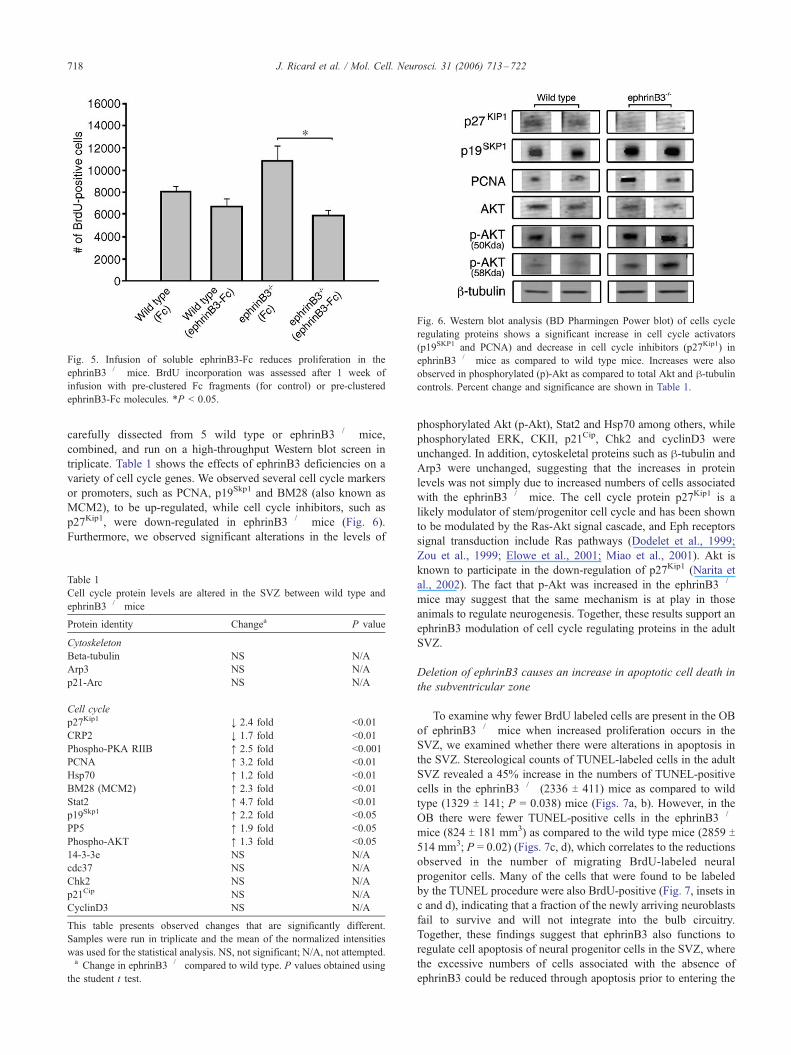
Fig. 5. Infusion of soluble ephrinB3-Fc reduces proliferation in the
ephrinB3�/� mice. BrdU incorporation was assessed after 1 week of
infusion with pre-clustered Fc fragments (for control) or pre-clustered
ephrinB3-Fc molecules. *P < 0.05.
Fig. 6. Western blot analysis (BD Pharmingen Power blot) of cells cycle
regulating proteins shows a significant increase in cell cycle activators
(p19SKP1 and PCNA) and decrease in cell cycle inhibitors (p27Kip1) in
ephrinB3�/� mice as compared to wild type mice. Increases were also
observed in phosphorylated (p)-Akt as compared to total Akt and h-tubulincontrols. Percent change and significance are shown in Table 1.
J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722718
carefully dissected from 5 wild type or ephrinB3�/� mice,
combined, and run on a high-throughput Western blot screen in
triplicate. Table 1 shows the effects of ephrinB3 deficiencies on a
variety of cell cycle genes. We observed several cell cycle markers
or promoters, such as PCNA, p19Skp1 and BM28 (also known as
MCM2), to be up-regulated, while cell cycle inhibitors, such as
p27Kip1, were down-regulated in ephrinB3�/� mice (Fig. 6).
Furthermore, we observed significant alterations in the levels of
Table 1
Cell cycle protein levels are altered in the SVZ between wild type and
ephrinB3�/� mice
Protein identity Changea P value
Cytoskeleton
Beta-tubulin NS N/A
Arp3 NS N/A
p21-Arc NS N/A
Cell cycle
p27Kip1 , 2.4 fold <0.01
CRP2 , 1.7 fold <0.01
Phospho-PKA RIIB j 2.5 fold <0.001
PCNA j 3.2 fold <0.01
Hsp70 j 1.2 fold <0.01
BM28 (MCM2) j 2.3 fold <0.01
Stat2 j 4.7 fold <0.01
p19Skp1 j 2.2 fold <0.05
PP5 j 1.9 fold <0.05
Phospho-AKT j 1.3 fold <0.05
14-3-3e NS N/A
cdc37 NS N/A
Chk2 NS N/A
p21Cip NS N/A
CyclinD3 NS N/A
This table presents observed changes that are significantly different.
Samples were run in triplicate and the mean of the normalized intensities
was used for the statistical analysis. NS, not significant; N/A, not attempted.a Change in ephrinB3�/� compared to wild type. P values obtained using
the student t test.
phosphorylated Akt (p-Akt), Stat2 and Hsp70 among others, while
phosphorylated ERK, CKII, p21Cip, Chk2 and cyclinD3 were
unchanged. In addition, cytoskeletal proteins such as h-tubulin and
Arp3 were unchanged, suggesting that the increases in protein
levels was not simply due to increased numbers of cells associated
with the ephrinB3�/� mice. The cell cycle protein p27Kip1 is a
likely modulator of stem/progenitor cell cycle and has been shown
to be modulated by the Ras-Akt signal cascade, and Eph receptors
signal transduction include Ras pathways (Dodelet et al., 1999;
Zou et al., 1999; Elowe et al., 2001; Miao et al., 2001). Akt is
known to participate in the down-regulation of p27Kip1 (Narita et
al., 2002). The fact that p-Akt was increased in the ephrinB3�/�
mice may suggest that the same mechanism is at play in those
animals to regulate neurogenesis. Together, these results support an
ephrinB3 modulation of cell cycle regulating proteins in the adult
SVZ.
Deletion of ephrinB3 causes an increase in apoptotic cell death in
the subventricular zone
To examine why fewer BrdU labeled cells are present in the OB
of ephrinB3�/� mice when increased proliferation occurs in the
SVZ, we examined whether there were alterations in apoptosis in
the SVZ. Stereological counts of TUNEL-labeled cells in the adult
SVZ revealed a 45% increase in the numbers of TUNEL-positive
cells in the ephrinB3�/� (2336 T 411) mice as compared to wild
type (1329 T 141; P = 0.038) mice (Figs. 7a, b). However, in the
OB there were fewer TUNEL-positive cells in the ephrinB3�/�
mice (824 T 181 mm3) as compared to the wild type mice (2859 T514 mm3; P = 0.02) (Figs. 7c, d), which correlates to the reductions
observed in the number of migrating BrdU-labeled neural
progenitor cells. Many of the cells that were found to be labeled
by the TUNEL procedure were also BrdU-positive (Fig. 7, insets in
c and d), indicating that a fraction of the newly arriving neuroblasts
fail to survive and will not integrate into the bulb circuitry.
Together, these findings suggest that ephrinB3 also functions to
regulate cell apoptosis of neural progenitor cells in the SVZ, where
the excessive numbers of cells associated with the absence of
ephrinB3 could be reduced through apoptosis prior to entering the
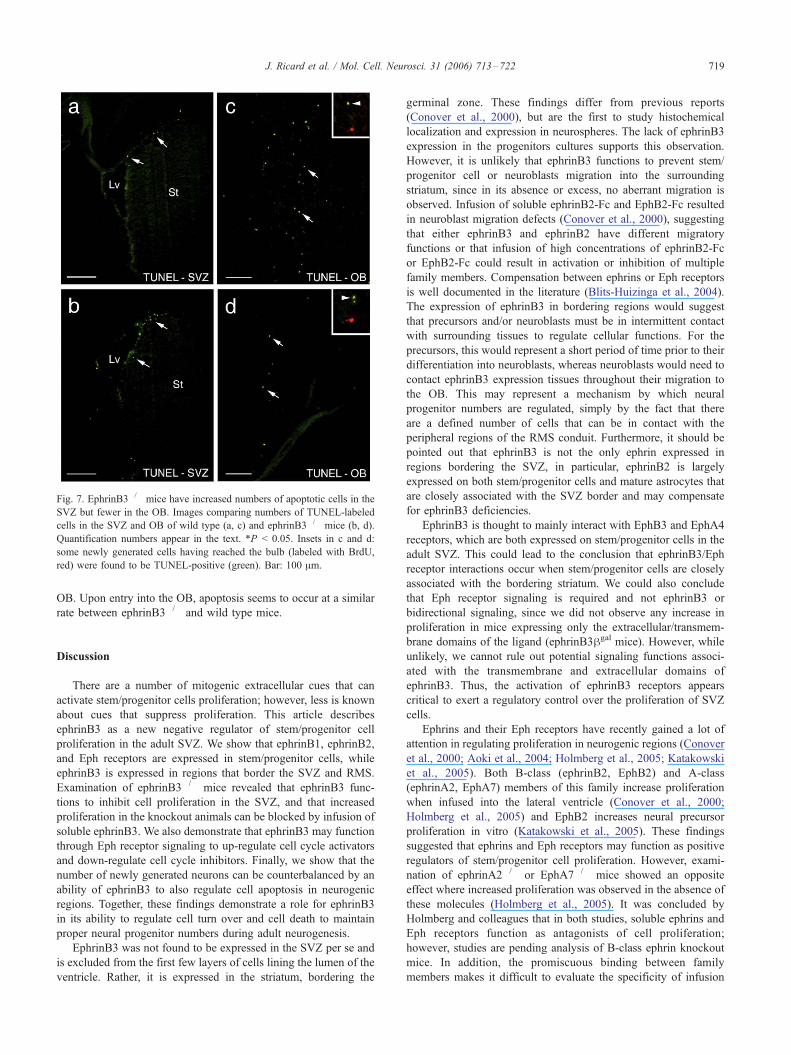
Fig. 7. EphrinB3�/� mice have increased numbers of apoptotic cells in the
SVZ but fewer in the OB. Images comparing numbers of TUNEL-labeled
cells in the SVZ and OB of wild type (a, c) and ephrinB3�/� mice (b, d).
Quantification numbers appear in the text. *P < 0.05. Insets in c and d:
some newly generated cells having reached the bulb (labeled with BrdU,
red) were found to be TUNEL-positive (green). Bar: 100 Am.
J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722 719
OB. Upon entry into the OB, apoptosis seems to occur at a similar
rate between ephrinB3�/� and wild type mice.
Discussion
There are a number of mitogenic extracellular cues that can
activate stem/progenitor cells proliferation; however, less is known
about cues that suppress proliferation. This article describes
ephrinB3 as a new negative regulator of stem/progenitor cell
proliferation in the adult SVZ. We show that ephrinB1, ephrinB2,
and Eph receptors are expressed in stem/progenitor cells, while
ephrinB3 is expressed in regions that border the SVZ and RMS.
Examination of ephrinB3�/� mice revealed that ephrinB3 func-
tions to inhibit cell proliferation in the SVZ, and that increased
proliferation in the knockout animals can be blocked by infusion of
soluble ephrinB3. We also demonstrate that ephrinB3 may function
through Eph receptor signaling to up-regulate cell cycle activators
and down-regulate cell cycle inhibitors. Finally, we show that the
number of newly generated neurons can be counterbalanced by an
ability of ephrinB3 to also regulate cell apoptosis in neurogenic
regions. Together, these findings demonstrate a role for ephrinB3
in its ability to regulate cell turn over and cell death to maintain
proper neural progenitor numbers during adult neurogenesis.
EphrinB3 was not found to be expressed in the SVZ per se and
is excluded from the first few layers of cells lining the lumen of the
ventricle. Rather, it is expressed in the striatum, bordering the
germinal zone. These findings differ from previous reports
(Conover et al., 2000), but are the first to study histochemical
localization and expression in neurospheres. The lack of ephrinB3
expression in the progenitors cultures supports this observation.
However, it is unlikely that ephrinB3 functions to prevent stem/
progenitor cell or neuroblasts migration into the surrounding
striatum, since in its absence or excess, no aberrant migration is
observed. Infusion of soluble ephrinB2-Fc and EphB2-Fc resulted
in neuroblast migration defects (Conover et al., 2000), suggesting
that either ephrinB3 and ephrinB2 have different migratory
functions or that infusion of high concentrations of ephrinB2-Fc
or EphB2-Fc could result in activation or inhibition of multiple
family members. Compensation between ephrins or Eph receptors
is well documented in the literature (Blits-Huizinga et al., 2004).
The expression of ephrinB3 in bordering regions would suggest
that precursors and/or neuroblasts must be in intermittent contact
with surrounding tissues to regulate cellular functions. For the
precursors, this would represent a short period of time prior to their
differentiation into neuroblasts, whereas neuroblasts would need to
contact ephrinB3 expression tissues throughout their migration to
the OB. This may represent a mechanism by which neural
progenitor numbers are regulated, simply by the fact that there
are a defined number of cells that can be in contact with the
peripheral regions of the RMS conduit. Furthermore, it should be
pointed out that ephrinB3 is not the only ephrin expressed in
regions bordering the SVZ, in particular, ephrinB2 is largely
expressed on both stem/progenitor cells and mature astrocytes that
are closely associated with the SVZ border and may compensate
for ephrinB3 deficiencies.
EphrinB3 is thought to mainly interact with EphB3 and EphA4
receptors, which are both expressed on stem/progenitor cells in the
adult SVZ. This could lead to the conclusion that ephrinB3/Eph
receptor interactions occur when stem/progenitor cells are closely
associated with the bordering striatum. We could also conclude
that Eph receptor signaling is required and not ephrinB3 or
bidirectional signaling, since we did not observe any increase in
proliferation in mice expressing only the extracellular/transmem-
brane domains of the ligand (ephrinB3hgal mice). However, while
unlikely, we cannot rule out potential signaling functions associ-
ated with the transmembrane and extracellular domains of
ephrinB3. Thus, the activation of ephrinB3 receptors appears
critical to exert a regulatory control over the proliferation of SVZ
cells.
Ephrins and their Eph receptors have recently gained a lot of
attention in regulating proliferation in neurogenic regions (Conover
et al., 2000; Aoki et al., 2004; Holmberg et al., 2005; Katakowski
et al., 2005). Both B-class (ephrinB2, EphB2) and A-class
(ephrinA2, EphA7) members of this family increase proliferation
when infused into the lateral ventricle (Conover et al., 2000;
Holmberg et al., 2005) and EphB2 increases neural precursor
proliferation in vitro (Katakowski et al., 2005). These findings
suggested that ephrins and Eph receptors may function as positive
regulators of stem/progenitor cell proliferation. However, exami-
nation of ephrinA2�/� or EphA7�/� mice showed an opposite
effect where increased proliferation was observed in the absence of
these molecules (Holmberg et al., 2005). It was concluded by
Holmberg and colleagues that in both studies, soluble ephrins and
Eph receptors function as antagonists of cell proliferation;
however, studies are pending analysis of B-class ephrin knockout
mice. In addition, the promiscuous binding between family
members makes it difficult to evaluate the specificity of infusion

J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722720
experiments. Our studies address both these points through
analysis of cell proliferation in the SVZ of ephrinB3�/� mice.
We found that ephrinB3 functions as a negative regulator of stem/
progenitor cell proliferation in the SVZ, and unlike the previous
studies we have found that infusion of clustered ephrinB3 func-
tions to suppress proliferation, hence restoring proliferation to
levels similar to the ones found in wild type animals. The fact that
ephrinB3 infusion does not have a significant effect on prolifer-
ation in wild type mice supports the specificity of the function of
ephrinB3 in regulating cell proliferation and suggests that ephrinB3
may only partially regulate cell proliferation. This is confirmed by
the alterations in cell cycle proteins in SVZ tissues, where cell
cycle activators (e.g., PCNA) are up-regulated while cell cycle
inhibitors (e.g., cyclin-dependent kinase inhibitor p27Kip1) are
down-regulated. One could argue that the increased numbers of
proliferating cells in the SVZ are due to an impairment of
neuroblast chain migration in the RMS, since fewer cells reach
the OB in the ephrinB3�/� mice. While we cannot completely
exclude this possibility, the increased expression of vimentin in the
ephrinB3�/� SVZ, an intermediate filament associated with
precursor and glial cells but not PSA-NCAM-positive neuroblasts
(Doetsch et al., 1997), supports the role of ephrinB3 in stem/
progenitor cell proliferation. Moreover, the changes in cell cycle
proteins detected by Western blot analysis also support a
proliferative function for ephrinB3.
One perplexing observation from our studies is the lack of
effect on proliferation when the receptors of ephrinB3 are knocked
out. Neither EphB3 nor EphA4 deletion resulted in the similar
increases in proliferation. One would expect that if the interaction
between an ephrin and its receptor(s) triggers a signal to regulate
cell proliferation, the lack of either signaling partner would result
in the same effect. This was the case for ephrinA2 and EphA7, as
mice lacking the expression of either one similarly exhibited more
proliferation in the SVZ (Holmberg et al., 2005). The absence of
proliferation increase in mice lacking EphA4 or EphB3 may
suggest that the two receptors could compensate for each other, and
that the deletion of both of them would be required to observe any
changes in proliferation. This hypothesis is supported by the fact
that both receptors are expressed on stem/progenitors cell in the
adult SVZ.
An increase in cell death was observed in the ephrinB3�/� mice
in parallel to increases in cell proliferation. This has been observed
in several cases when proliferation was up-regulated in the SVZ
(Belvindrah et al., 2002; Doetsch et al., 2002a,b; Vanderluit et al.,
2004). Conversely, in mice lacking E2F1 expression, a decrease in
the proliferation level in the SVZ occurs simultaneously with a
decrease in TUNEL-positive cells (Cooper-Kuhn et al., 2002).
However, some studies have shown opposite trends between
proliferation and cell death in the case of PTEN (Groszer et al.,
2001), or no effect in apoptosis as in the case of VEGF (Jin et al.,
2002). More likely, fluctuations in proliferation precede alterations
in cell death, since apoptosis is known to be one of the regulatory
components that control stem cell numbers and progeny output
(Sommer and Rao, 2002). This would ensure constant levels of
new neurons to meet the specific needs of the OB. Recently, A-
class ephrins and their receptors have been implicated in apoptotic
events (Dohn et al., 2001; Depaepe et al., 2005). In particular,
EphA7 and ephrinA5 have been shown to control neural progenitor
cell death, where EphA7�/� mice exhibit decreased cell death
(Depaepe et al., 2005). This would suggest that ephrinB3 function
in an opposite manner to suppress the apoptotic response.
Together, ephrinB3 may be a key regulator of regulating of both
cellular proliferation and apoptosis needed to maintain proper
neuronal numbers in the OB.
Here, we have identified ephrinB3 as a new negative regulator
of cell proliferation and positive regulator of cell survival within
the adult SVZ, where stem/progenitor cells expressing Eph
receptors interact with bordering tissues expressing ephrinB3 in
the adult SVZ. These findings demonstrate a novel function for
ephrinB3 and reinforce the importance of the ephrin/Eph families
as endogenous modulators of neurogenesis.
Experimental methods
Animals and tissue preparation
The generation of the mutant CD1 mice has been described
(Henkemeyer et al., 1996; Orioli et al., 1996; Cowan et al., 2000;
Yokoyama et al., 2001). Genotyping was performed by PCR analysis. Mice
were perfused intracardially with 4% paraformaldehyde. Brains were
removed and post-fixed for 2 h (for X-gal or Ki67 staining) or overnight
in the same solution, cryopreserved in 25% sucrose, and frozen in OCT
(Tissue Tek).
Staining procedures
For immunostaining, sections were washed with PBS, permeabilized for
10 min in 0.2% Triton-X100 and blocked in 5% BSA for 30 min. Primary
antibodies were applied for 1 h at room temperature (polyclonal anti-GFAP:
1/1000, Dako; monoclonal anti-GFAP: 1/1000, Pharmingen; anti-PSA-
NCAM: 1/1000, Chemicon) or overnight at 4-C (anti-Ki67: 1/25, Dako).
We observed no significant difference in the immunostaining pattern in the
SVZ between the monoclonal and polyclonal GFAP antibodies (not
shown). The detection of Ki67 required antigen retrieval prior to the
staining (sections placed in 10 mM citrate pH 6.0 and heated in a
microwave oven twice for 4 min). Staining for ephrinB1 (1/100; Santa
Cruz) and ephrinB2 (1/100; Santa Cruz) was performed on fresh frozen
tissue (isopentane at dry ice temperature). The specificity of the antibodies
was first assessed on recombinant ligands (100 ng/lane ephrinB1-Fc,
ephrinB2-Fc, and ephrinB3-Fc molecules; R&D Systems) by Western blot,
where both anti-ephrinB1 and anti-ephrinB2 were found to be specific for
their respective antigens (not shown). Alexa Fluor 488- or Alexa Fluor 594-
conjugated secondary antibodies (Molecular Probes) were used for 30 min
at room temperature.
EphrinB3hgal mice (expressing fusion proteins where h-galactosidasereplaces the intracytoplasmic domain of the ephrin) were used to visualize
the areas of expression of ephrinB3. X-gal staining (5 mM potassium ferro/
ferricyanide, 2 mM MgCl2, 1 mg/ml X-gal in PBS) was performed at room
temperature until a strong blue color was visible.
TUNEL staining was performed on PFA-fixed tissue using the ApopTag
Plus Fluorescein In Situ detection kit (Chemicon) according to the
manufacturer’s instructions.
BrdU injection and detection
Mice were injected intraperitoneally with 50 Ag BrdU per gram of body
weight once for 1 h for stereological counts or three times (2 h apart) before
waiting for 7 days for migration studies. Prior to the immunostaining (anti-
BrdU: 1/25, Roche), sections were incubated in 2N HCl for 30 min at 37-Cand neutralized in 0.1 M borate pH 8.5.
Stereological procedures
Cell counts were performed on 30 Am-thick cryostat brain sections (the
first section was collected when the rostral part of the lateral ventricle was
reached). Staining was performed on 10 sections every fifth section (total

J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722 721
thickness of the examined tissue was 1500 Am). For neuroblasts counts,
PSA-NCAM was revealed with an Alexa Fluor 594 secondary antibody, the
nuclei were counterstained using Sytox Green (1/100,000, Molecular
Probes), and the tissue was analyzed under a dual FITC/Rhodamine filter
on a Zeiss Axiophot microscope equipped with a CCD camera. The
analysis was conducted using the StereoInvestigator software (Micro-
BrightField) to estimate the total number of cells in the volume examined.
Western blot analysis
Western blot analysis was performed by BD Pharmingen using the
Power blot system. Protein samples were prepared according to Pharmingen
instructions. Briefly, proteins were separated on 4–15% gradient SDS-
polyacrylamide gels. After transfer to Immobilon-P membrane (Millipore),
the membrane was blocked for 1 h with blocking buffer (LI-COR). Primary
antibodies were applied to the membrane using a manifold that isolates 40
different channels. Primary antibodies were allowed to bind for 1 h at 37-C.The secondary antibodies were Alexa 680-conjugated goat anti-mouse
(Molecular Probes) and IRDye 800-conjugated goat anti-rabbit (Rockland).
The membrane was scanned on an Odyssey Infrared Imaging System (LI-
COR Biosciences) and the raw measured intensities were normalized
against intensities obtained from a standardization cocktail of proteins.
Samples were run in triplicate. Antibodies from the Cell Cycle and
Phosphoproteins sets (BD Pharmingen) were chosen for the study.
Neurosphere cultures
Adult neural progenitor cells were isolated from the lateral ventricle
wall of 2 month old CD1 mice. 1 mm coronal slices corresponding to
anteroposterior coordinates from 1 mm to 0 relative to bregma were cut.
The ventricle wall on the striatal side was dissected out and the tissue was
dissociated in 1.33 mg/ml trypsin, 0.7 mg/ml hyaluronic acid and 0.2 mg/ml
kynurenic acid in HBSS, and cells were grown in DMEM-F12 medium
supplemented with B27 (Life Technologies) in the presence of 20 ng/ml
EGF (Life Technologies) and 10 ng/ml bFGF (Chemicon). Half of the
medium was replaced every 4 days with fresh mitogens. For the primary
neurosphere assay, 2000 cells (from the SVZ tissue of 3 pooled mice) were
plated in 6-well plates and cultured for 7 days, after which the numbers of
neurospheres formed were counted.
Intracerebral infusions
EphrinB3-Fc ligands (R&D Systems; 140 Ag/ml in PBS) were pre-
clustered for 2 h at room temperature (anti-human-Fc ratio 10:1) and were
infused over a 7-day period using osmotic pumps (100 Al volume, rate 0.5
Al/h, connected to brain infusion devices (Alzet) at the following
coordinates (from bregma: lateral 0.5 mm; posterior �0.7 mm). Clustered
Fc fragments were used as controls. For tracing, 1 Al of a 2 mg/ml solution
of DiO (3,3V-dihexadecyloxacarbocyanine perchlorate; Molecular Probes)
was injected in the ventricle (coordinates as above) using a Hamilton
syringe. Animals were analyzed 4 days after injection.
Acknowledgments
We thank Dr. Mark Henkemeyer for the gracious gift of the
ephrinB3 knockout mice. We also thank Dr. Steven Kernie for
critical reading of the manuscript. This work was supported by The
Miami Project to Cure Paralysis, Ralph C. Wilson Sr./Ralph C.
Wilson Jr. Medical Research Foundation, and the National
Institutes of Health, NINDS (NS049545/NS30291).
References
Agasse, F., Roger, M., Coronas, V., 2004. Neurogenic and intact or
apoptotic non-neurogenic areas of adult brain release diffusible
molecules that differentially modulate the development of subventric-
ular zone cell cultures. Eur. J. Neurosci. 19, 1459–1468.
Alonso, G., 2001. Proliferation of progenitor cells in the adult rat brain
correlates with the presence of vimentin-expressing astrocytes. Glia 34,
253–266.
Alvarez-Buylla, A., Garcia-Verdugo, J.M., 2002. Neurogenesis in adult
subventricular zone. J. Neurosci. 22, 629–634.
Alvarez-Buylla, A., Garcia-Verdugo, J.M., Tramontin, A.D., 2001. A
unified hypothesis on the lineage of neural stem cells. Nat. Rev.,
Neurosci. 2, 287–293.
Aoki, M., Yamashita, T., Tohyama, M., 2004. EphA receptors direct the
differentiation of mammalian neural precursor cells through a mitogen-
activated protein kinase-dependent pathway. J. Biol. Chem. 279,
32643–32650.
Belvindrah, R., Rougon, G., Chazal, G., 2002. Increased neurogenesis in
adult mCD24-deficient mice. J. Neurosci. 22, 3594–3607.
Betarbet, R., Zigova, T., Bakay, R.A., Luskin, M.B., 1996. Dopaminergic
and GABAergic interneurons of the olfactory bulb are derived from the
neonatal subventricular zone. Int. J. Dev. Neurosci. 14, 921–930.
Blits-Huizinga, C.T., Nelersa, C.M., Malhotra, A., Liebl, D.J., 2004.
Ephrins and their receptors: binding versus biology. IUBMB Life 56,
257–265.
Conover, J.C., Doetsch, F., Garcia-Verdugo, J.M., Gale, N.W., Yancopoulos,
G.D., Alvarez-Buylla, A., 2000. Disruption of Eph/ephrin signaling
affects migration and proliferation in the adult subventricular zone. Nat.
Neurosci. 3, 1091–1097.
Cooper-Kuhn, C.M., Vroemen, M., Brown, J., Ye, H., Thompson,
M.A., Winkler, J., Kuhn, H.G., 2002. Impaired adult neurogenesis
in mice lacking the transcription factor E2F1. Mol. Cell. Neurosci.
21, 312–323.
Cowan, C.A., Yokoyama, N., Bianchi, L.M., Henkemeyer, M., Fritzsch, B.,
2000. EphB2 guides axons at the midline and is necessary for normal
vestibular function. Neuron 26, 417–430.
Dasgupta, B., Gutmann, D.H., 2005. Neurofibromin regulates neural stem
cell proliferation, survival, and astroglial differentiation in vitro and in
vivo. J. Neurosci. 25, 5584–5594.
Depaepe, V., Suarez-Gonzalez, N., Dufour, A., Passante, L., Gorski, J.A.,
Jones, K.R., Ledent, C., Vanderhaeghen, P., 2005. Ephrin signalling
controls brain size by regulating apoptosis of neural progenitors. Nature
435, 1244–1250.
Dodelet, V.C., Pazzagli, C., Zisch, A.H., Hauser, C.A., Pasquale, E.B.,
1999. A novel signaling intermediate, SHEP1, directly couples Eph
receptors to R-Ras and Rap1A. J. Biol. Chem. 274, 31941–31946.
Doetsch, F., Garcia-Verdugo, J.M., Alvarez-Buylla, A., 1997. Cellular
composition and three-dimensional organization of the subven-
tricular germinal zone in the adult mammalian brain. J. Neurosci.
17, 5046–5061.
Doetsch, F., Petreanu, L., Caille, I., Garcia-Verdugo, J.M., Alvarez-Buylla,
A., 2002a. EGF converts transit-amplifying neurogenic precursors in
the adult brain into multipotent stem cells. Neuron 36, 1021–1034.
Doetsch, F., Verdugo, J.M., Caille, I., Alvarez-Buylla, A., Chao, M.V.,
Casaccia-Bonnefil, P., 2002b. Lack of the cell-cycle inhibitor p27Kip1
results in selective increase of transit-amplifying cells for adult neuro-
genesis. J. Neurosci. 22, 2255–2264.
Dohn, M., Jiang, J., Chen, X., 2001. Receptor tyrosine kinase EphA2 is
regulated by p53-family proteins and induces apoptosis. Oncogene 20,
6503–6515.
Elowe, S., Holland, S.J., Kulkarni, S., Pawson, T., 2001. Downregulation of
the Ras-mitogen-activated protein kinase pathway by the EphB2
receptor tyrosine kinase is required for ephrin-induced neurite retrac-
tion. Mol. Cell. Biol. 21, 7429–7441.
Flanagan, J.G., Vanderhaeghen, P., 1998. The ephrins and Eph receptors in
neural development. Annu. Rev. Neurosci. 21, 309–345.
Groszer, M., Erickson, R., Scripture-Adams, D.D., Lesche, R., Trumpp, A.,
Zack, J.A., Kornblum, H.I., Liu, X., Wu, H., 2001. Negative regulation
of neural stem/progenitor cell proliferation by the Pten tumor suppressor
gene in vivo. Science 294, 2186–2189.

J. Ricard et al. / Mol. Cell. Neurosci. 31 (2006) 713–722722
Henkemeyer, M., Orioli, D., Henderson, J.T., Saxton, T.M., Roder, J.,
Pawson, T., Klein, R., 1996. Nuk controls pathfinding of commissural
axons in the mammalian central nervous system. Cell 86, 35–46.
Holmberg, J., Armulik, A., Senti, K.A., Edoff, K., Spalding, K., Momma,
S., Cassidy, R., Flanagan, J.G., Frisen, J., 2005. Ephrin-A2 reverse
signaling negatively regulates neural progenitor proliferation and
neurogenesis. Genes Dev. 19, 462–471.
Jin, K., Zhu, Y., Sun, Y., Mao, X.O., Xie, L., Greenberg, D.A., 2002.
Vascular endothelial growth factor (VEGF) stimulates neurogenesis in
vitro and in vivo. Proc. Natl. Acad. Sci. U. S. A. 99, 11946–11950.
Jin, K., Sun, Y., Xie, L., Batteur, S., Mao, X.O., Smelick, C., Logvinova,
A., Greenberg, D.A., 2003. Neurogenesis and aging: FGF-2 and HB-
EGF restore neurogenesis in hippocampus and subventricular zone of
aged mice. Aging Cell 2, 175–183.
Katakowski, M., Zhang, Z., Decarvalho, A.C., Chopp, M., 2005. EphB2
induces proliferation and promotes a neuronal fate in adult subven-
tricular neural precursor cells. Neurosci. Lett. 385, 204–209.
Kirschenbaum, B., Doetsch, F., Lois, C., Alvarez-Buylla, A., 1999. Adult
subventricular zone neuronal precursors continue to proliferate
and migrate in the absence of the olfactory bulb. J. Neurosci. 19,
2171–2180.
Lemkine, G.F., Raji, A., Alfama, G., Turque, N., Hassani, Z., Alegria-
Prevot, O., Samarut, J., Levi, G., Demeneix, B.A., 2005. Adult neural
stem cell cycling in vivo requires thyroid hormone and its alpha
receptor. FASEB J. 19, 863–865.
Leone, D.P., Relvas, J.B., Campos, L.S., Hemmi, S., Brakebusch, C.,
Fassler, R., Ffrench-Constant, C., Suter, U., 2005. Regulation of neural
progenitor proliferation and survival by {beta}1 integrins. J. Cell Sci.
118, 2589–2599.
Luskin, M.B., 1993. Restricted proliferation and migration of postnatally
generated neurons derived from the forebrain subventricular zone.
Neuron 11, 173–189.
Mandairon, N., Jourdan, F., Didier, A., 2003. Deprivation of sensory
inputs to the olfactory bulb up-regulates cell death and prolifera-
tion in the subventricular zone of adult mice. Neuroscience 119,
507–516.
Miao, H., Wei, B.R., Peehl, D.M., Li, Q., Alexandrou, T., Schelling, J.R.,
Rhim, J.S., Sedor, J.R., Burnett, E., Wang, B., 2001. Activation of
EphA receptor tyrosine kinase inhibits the Ras/MAPK pathway. Nat.
Cell Biol. 3, 527–530.
Narita, Y., Nagane, M., Mishima, K., Huang, H.J., Furnari, F.B.,
Cavenee, W.K., 2002. Mutant epidermal growth factor receptor
signaling down-regulates p27 through activation of the phosphatidy-
linositol 3-kinase/Akt pathway in glioblastomas. Cancer Res. 62,
6764–6769.
Ohnuma, S., Harris, W.A., 2003. Neurogenesis and the cell cycle. Neuron
40, 199–208.
Orioli, D., Henkemeyer, M., Lemke, G., Klein, R., Pawson, T., 1996. Sek4
and Nuk receptors cooperate in guidance of commissural axons and in
palate formation. EMBO J. 15, 6035–6049.
Reynolds, B.A., Rietze, R.L., 2005. Neural stem cells and neurospheres—
Re-evaluating the relationship. Nat. Methods 2, 333–336.
Salman, H., Ghosh, P., Kernie, S.G., 2004. Subventricular zone neural
stem cells remodel the brain following traumatic injury in adult mice.
J. Neurotrauma 21, 283–292.
Sommer, L., Rao, M., 2002. Neural stem cells and regulation of cell
number. Prog. Neurobiol. 66, 1–18.
Vanderluit, J.L., Ferguson, K.L., Nikoletopoulou, V., Parker, M.,
Ruzhynsky, V., Alexson, T., McNamara, S.M., Park, D.S., Rudnicki,
M., Slack, R.S., 2004. p107 regulates neural precursor cells in the
mammalian brain. J. Cell Biol. 166, 853–863.
Wang, T.W., Zhang, H., Parent, J.M., 2005. Retinoic acid regulates
postnatal neurogenesis in the murine subventricular zone-olfactory bulb
pathway. Development 132, 2721–2732.
Yokoyama, N., Romero, M.I., Cowan, C.A., Galvan, P., Helmbacher, F.,
Charnay, P., Parada, L.F., Henkemeyer, M., 2001. Forward signaling
mediated by ephrin-B3 prevents contralateral corticospinal axons from
recrossing the spinal cord midline. Neuron 29, 85–97.
Zhang, R., Zhang, Z., Tsang, W., Wang, L., Chopp, M., 2004. Down-
regulation of p27kip1 increases proliferation of progenitor cells in adult
rats. NeuroReport 15, 1797–1800.
Zou, J.X., Wang, B., Kalo, M.S., Zisch, A.H., Pasquale, E.B., Ruoslahti, E.,
1999. An Eph receptor regulates integrin activity through R-Ras. Proc.
Natl. Acad. Sci. U. S. A. 96, 13813–13818.





























