Embed Size (px)
Citation preview

lable at ScienceDirect
Atherosclerosis 233 (2014) 149e156
Contents lists avai
Atherosclerosis
journal homepage: www.elsevier .com/locate/atherosclerosis
Epicatechin attenuates atherosclerosis and exerts anti-inflammatoryeffects on diet-induced human-CRP and NFkB in vivo
Martine Morrison a,b,c,*, Roel van der Heijden b,c, Peter Heeringa b, Eric Kaijzel d,Lars Verschuren a, Rune Blomhoff e, Teake Kooistra a, Robert Kleemann a
aDepartment of Metabolic Health Research, Netherlands Organisation for Applied Scientific Research (TNO), Zernikedreef 9, 2333 CK Leiden,The NetherlandsbDepartment of Pathology and Medical Biology, University of Groningen, University Medical Center Groningen, Hanzeplein 1 (EA11), 9713 GZ Groningen,The Netherlandsc Top Institute Food and Nutrition, Nieuwe Kanaal 9A, 6709 PA Wageningen, The NetherlandsdMolecular Endocrinology and Molecular Imaging, Department of Endocrinology, Leiden University Medical Center, Albinusdreef 2, 2333 ZA Leiden,The NetherlandseDepartment of Nutrition, Institute of Basic Medical Sciences, Faculty of Medicine, University of Oslo, Sognsvannsveien 9, 0372 Oslo, Norway
a r t i c l e i n f o
Article history:Received 12 October 2013Received in revised form11 December 2013Accepted 22 December 2013Available online 8 January 2014
Keywords:AtherosclerosisCRPEpicatechinInflammationNFkBPolyphenol
Abbreviations: human-CRP, human C-reactive pdisease; E3L, ApoE*3-Leiden; HC, high-cholescholesterol þ epicatechin group; HCA, high-cholesteromatrix metalloproteinase; REF, reference group; SAA,* Corresponding author. TNO Metabolic Health Rese
Leiden, The Netherlands. Tel.: þ31 888 664 448.E-mail address: [email protected] (M. Mor
0021-9150/$ e see front matter � 2014 Elsevier Irelahttp://dx.doi.org/10.1016/j.atherosclerosis.2013.12.027
a b s t r a c t
Objective: Previous studies investigating flavanol-rich foods provide indications for potential car-dioprotective effects of these foods, but the effects of individual flavanols remain unclear. We investi-gated whether the flavanol epicatechin can reduce diet-induced atherosclerosis, with particularemphasis on the cardiovascular risk factors dyslipidaemia and inflammation.Methods: ApoE*3-Leiden mice were fed a cholesterol-containing atherogenic diet with or without epi-catechin (0.1% w/w) to study effects on early- and late-stage atherosclerosis (8w and 20w). In vivo effectsof epicatechin on diet-induced inflammation were studied in human-CRP transgenic mice and NFkB-luciferase reporter mice.Results: Epicatechin attenuated atherosclerotic lesion area in ApoE*3-Leiden mice by 27%, withoutaffecting plasma lipids. This anti-atherogenic effect of epicatechin was specific to the severe lesion types,with no effect on mild lesions. Epicatechin mitigated diet-induced increases in plasma SAA (in ApoE*3-Leiden mice) and plasma human-CRP (in human-CRP transgenic mice). Microarray analysis of aortic geneexpression revealed an attenuating effect of epicatechin on several diet-induced pro-atherogenic in-flammatory processes in the aorta (e.g. chemotaxis of cells, matrix remodelling), regulated by NFkB.These findings were confirmed immunohistochemically by reduced lesional neutrophil content in HCE,and by inhibition of diet-induced NFkB activity in epicatechin-treated NFkB-luciferase reporter mice.Conclusion: Epicatechin attenuates development of atherosclerosis and impairs lesion progression frommild to severe lesions in absence of an effect on dyslipidaemia. The observed reduction of circulatinginflammatory risk factors by epicatechin (e.g. SAA, human-CRP), as well as its local anti-inflammatoryactivity in the vessel wall, provide a rationale for epicatechin’s anti-atherogenic effects.
� 2014 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Cardiovascular disease (CVD) is the leading cause of deathworldwide. Its major underlying pathology is atherosclerosis, a
rotein; CVD, cardiovascularterol group; HCE, high-l þ atorvastatin group; MMP,serum amyloid A.arch, Zernikedreef 9, 2333CK
rison).
nd Ltd. All rights reserved.
complex, multi-factorial disease that is driven by dyslipidaemia andchronic inflammation [1]. Accumulation of lipids and inflammatorycells in the arterial wall promotes formation of mild lesions knownas fatty streaks. Over time, continuous inflammatory stress drivesprogression of these early-stage mild lesions into late-stage severelesions that are characterised by extensive vascular remodelling.
Diet is an important determinant of CVD risk [2,3]. Epidemio-logic studies have provided consistent evidence that the incidenceof CVD is associated with diets rich in saturated fat [4], which maybe mediated by increased levels of circulating inflammatory factors(e.g. liver-derived SAA, CRP, reviewed in Refs. [5,6]). In contrast, adiet rich in fruit and vegetables is associated with decreased levels

M. Morrison et al. / Atherosclerosis 233 (2014) 149e156150
of inflammatory markers [7e9] and may attenuate the develop-ment of CVD [10]. The polyphenols, a large group of bioactivecompounds ubiquitous in plant-derived foods, are considered apotential mediator of these cardioprotective effects [11]. The fla-vanol subclass of the polyphenols in particular has been studiedextensively in relation to its putative cardioprotective properties[12]. The majority of these studies investigated health benefits offlavanol-rich foods such as tea and cocoa, and report reduced risk ofmyocardial infarction [13], CVD [14,15], stroke [16] and athero-sclerotic disease mortality [17]. Results from numerous humanintervention studies on the effects of these flavanol-rich foodsprovide further support for possible cardioprotective effects ofthese foods, although their effects on the CVD risk factorseplasmalipids and inflammatory markerseare not always consistent(recently reviewed in Refs. [18,19]).
Of the flavanols, (�)-epicatechin is considered as an importantcandidate that may be responsible for beneficial effects of theseflavanol-rich foods [20], and a strong inverse relationship with CVDmortality has been reported for (�)-epicatechin specifically [21].These results indicate a beneficial role for epicatechin in athero-genesis, all the more so because epicatechin has been shown tohave anti-inflammatory [22] and anti-oxidative [23] propertiesin vivo. In the latter study, Loke et al. reported a small (14%), non-significant effect of epicatechin on total lesion area in apoE�/�
mice, despite clear anti-oxidative effects of epicatechin. As apoE isinvolved in lipid metabolism as well as inflammatory processes,and apoE�/� mice lack this factor completely, the apoE�/� model isnot optimally suited to study diet-induced dyslipidaemia andinflammation in the development of atherosclerosis [24]. Here, westudied the effects of epicatechin on atherosclerosis in ApoE*3-Leiden (E3L) mice, an alternative model for atherosclerosis thatdoes allow for investigation of effects on dyslipidaemia andinflammation throughout the atherogenic process. When fed ahigh-fat/high-cholesterol diet, E3L mice develop mild dyslipidae-mia with a human-like lipoprotein profile, low-grade inflammationand atherosclerosis characterised bymoderate progression throughthe stages of disease, allowing for controlled analysis of early- andlate-stage processes [24,25]. Atorvastatinwas used as a cholesterol-reducing pharmaceutical reference. For a more thorough charac-terisation of the effects of epicatechin on diet-induced inflamma-tion we used specific models of liver and whole-bodyinflammation: human-CRP transgenic mice [26] and NFkB-lucif-erase reporter mice [27], respectively. Histological analysis of early-and late-stage atherosclerosis in E3L mice showed that epicatechinreduces atherosclerosis by attenuating progression to severelesions. Biochemical analyses together with aortic genome profilingprovide indication that these anti-atherogenic effects may atleast partly be explained by epicatechin’s anti-inflammatoryproperties.
2. Materials and methods
2.1. Animal experiments
Experiments were approved by an independent Animal Careand Use Committee and were in compliance with European Com-munity specifications regarding the use of laboratory animals.
2.1.1 Atherosclerosis experimentsFemale ApoE*3-Leiden transgenic (E3L) mice, characterised by
ELISA, were fed an atherogenic Western-type diet for a four-weekrun-in period. This diet contains 15% cocoa butter, 1% corn oil,40.5% sucrose, 20% acid casein, 10% corn starch and 6.2% cellulose(diet-T; AB-Diets, Woerden, the Netherlands), and was supple-mented with 1% (w/w) cholesterol (SigmaeAldrich, Zwijndrecht,
the Netherlands). After run-in on this high-cholesterol (HC) diet,mice were matched for plasma cholesterol and triglycerides into 4treatment groups (n ¼ 15 each). The control group continued toreceive the HC diet, and the epicatechin-treated group (HCE)received the HC diet supplemented with 0.1% (w/w) (�)-epi-catechin (Chromadex Inc., Irvine, USA) for 16 weeks. Two referencegroups were included: a 0.01% (w/w) atorvastatin-treated phar-maceutical reference group (HCA) (Lipitor, Pfizer, Capelle a/d Ijssel,the Netherlands) and an ageing reference group (REF) that receivedthe Western-type diet without cholesterol supplementation.Plasma epicatechin concentration in HCE mice was determined inpooled plasma (n ¼ 15 animals per pool, non-fasted) by HPLC-MS/MS on an Agilent HPLC 1200 Series with a Zorbax SB-Aq column(3.5 mm, 150 mm � 2.1 mm i.d.) (both Agilent Technologies, PaloAlto, USA)) as previously described [28].
EDTA plasma samples were collected from the tail vein after a4 h fasting period. Body weight and food intake were monitoredthroughout the study. Animals were sacrificed by CO2 asphyxiationafter 16 weeks of dietary treatment to collect hearts, aortas andkidneys. Hearts were fixed in formalin and embedded in paraffinfor atherosclerosis analysis. Descending aortas were snap-frozen inliquid nitrogen and stored at �80 �C for microarray analysis.
In an independent early-stage atherosclerosis experiment, fe-male E3L mice were matched for plasma cholesterol and tri-glycerides into an HC and an HCE group (n ¼ 15 each). Mice werefed the diets described above, without a run-in period. After 8weeks of dietary treatment, animals were sacrificed by CO2
asphyxiation and hearts were collected for atherosclerosis analysisas described above.
2.1.2. Diet-induced inflammation experimentsHuman-CRP transgenic mice carry a 31-kb human DNA frag-
ment containing the human-CRP gene including the entire human-CRP promoter. Male mice were characterised by human-CRP ELISA(R&D Systems, Abingdon, UK) andmatched into 2 treatment groups(n ¼ 10e14) based on plasma human-CRP. Mice were fed standardmouse chow (ssniff� R/M-H, ssniff Spezialdiäten, Soest, Germany)without or with epicatechin (0.06% w/w) for 2 weeks followed by ahigh-fat diet (45% kcal% fat D12451, Research Diets, NewBrunswick,USA) without (HF) or with (�)-epicatechin (0.1% w/w, HFE) for 3weeks. To correct for the higher food intake on the chow dietcompared with the fat-containing diets, the epicatechin concen-tration was adjusted to achieve a comparable daily intake. EDTAplasma was collected from the tail vein (non-fasted) before andafter HFD-feeding for human-CRP measurements. NFkB-luciferasereporter mice carry a transgene for the firefly luciferase gene,coupled with three binding sites for NF-kB. This transgene isflanked by insulator sequences from the chicken [b]-globin gene toreduce genomic interference [27]. Female mice were characterisedby RT-PCR. NF-kB activity was monitored non-invasively by in vivomolecular imaging in an IVIS spectrum in vivo imaging system(Caliper Life Sciences, Hopkinton, USA). Mice were anaesthetisedusing 2.5% isoflurane, and ventral fur was removed by shaving.Images were acquired 5 min after intraperitoneal injection of D-Luciferin (150 mg/kg, Caliper Life Sciences), with an exposure timeof 30 s. Living imaging 4 software (Caliper Life Sciences) was used toquantify the signal. Mice were matched into 2 experimental groupsaccording to baseline NFkB activity. Mice were fed HC diet (n ¼ 15)or HCE diet (n¼ 7) for 7 days after which diet-induced NFkB activitywas determined.
2.2. Histological, biochemical and microarray analyses
A detailed description of histological, biochemical andmicroarray analyses is provided in Supplement 1. Briefly,

M. Morrison et al. / Atherosclerosis 233 (2014) 149e156 151
atherosclerosis was scored histologically using an adaptedgrading method for human atherosclerosis, and the plasma pa-rameters and histological and RT-PCR analyses of renal pathologywere quantified as described [29]. Illumina microarray analysis ofaortic gene expression was performed following establishednormalisation and quality control protocols [25] as well aspathway analysis [30].
2.3. Statistical analysis
All data are presented as mean � SD. The atherosclerosis ex-periments investigated the null hypothesis that epicatechin maynot exert beneficial anti-atherosclerotic effects relative to the un-treated control (HC). Significance of differences between HCE andHC animals was tested using one-sided t-tests. The referencegroups (REF and HCA) were not required to test the null hypothesisand were used to control the experimental conditions of the dis-ease model. The experiments in the inflammation models investi-gated the potential effects of epicatechin in preventing diet-induced inflammation. Paired one-sided t-tests were used tocalculate the significance of induction of inflammation within eachgroup. Statistical tests were performed using SPSS software(version 20, IBM, Armonk USA) or Graphpad Prism software(version 5.03, Graphpad Software Inc., La Jolla, USA). Ap-value � 0.05 was considered statistically significant.
Fig. 1. Effects of 16-week epicatechin-treatment (0.1% w/w in atherogenic HC diet) on atheaortic root area showing smaller and less severe atherosclerotic lesions in HCE compared wivalve area (n ¼ 15/group). C: plasma cholesterol levels over time-course of the intervention.REF: reference, HC: high cholesterol, HCE: high cholesterol þ epicatechin, HCA: high chole
3. Results
3.1. Epicatechin attenuates atherosclerosis development withouteffect on plasma lipids
All treatments were well tolerated and food intake and bodyweight were comparable in HC and HCE (Supplement 2)throughout the 16-week study period. The calculated daily dose ofepicatechin in HCE was 110� 11 mg/kg body weight, based on foodintake measurements. HCE diet-feeding resulted in an averageplasma epicatechin concentration of 4.2 mM, i.e. within the micro-molar range achievable in humans [31]. The atherogenic HC-dietinduced pronounced atherosclerosis in HC relative to REF, with atotal atherosclerotic lesion area in the aortic root of515,190 � 265,747 mm2 compared with 11,797 � 11,822 mm2 in REF(Fig. 1a). Epicatechin treatment significantly attenuated the devel-opment of atherosclerosis by 27% (375,495 � 205,386 mm2, Fig. 1aeb). As expected, atherosclerosis development was strongly reducedin atorvastatin-treated reference mice (100,148 � 108,685 mm2).There were no effects of epicatechin on the microvasculature (HC-induced kidney injury) as assessed histologically (not shown).
Dyslipidaemia is a major risk factor for atherosclerosis develop-ment. HC-diet feeding strongly increased plasma cholesterol levels(17.05� 2.84mM) relative to REF (6.24� 0.66mM)within the first 4weeks and levels remained stable for the rest of the study, with an
rosclerosis and plasma lipids in E3L mice. A: representative photomicrographs of theth HC. B: total atherosclerotic lesion area quantified in serial cross-sections of the aorticD: lipoprotein profile for cholesterol distribution in VLDL, LDL and HDL-sized particles.sterol þ atorvastatin. Data are mean � SD. *p � 0.05 compared with HC.

Fig. 2. Effects of epicatechin on diet-induced inflammation in E3L mice and human-CRP transgenic mice. A: HC diet increases plasma SAA concentration during athero-genesis in E3L mice, epicatechin attenuates this effect (n ¼ 15/group). B: SAA exposure(area under curve) in E3L mice. C: HF-diet induces plasma human-CRP levels inhuman-CRP transgenic mice (HF; n ¼ 14) which is prevented by epicatechin (HFE;n ¼ 10). Data are mean � SD. *p < 0.05 compared with HC, #p � 0.05 for increasewithin group.
M. Morrison et al. / Atherosclerosis 233 (2014) 149e156152
averageof18.14�3.17mM(Fig.1c). The increase inplasmacholesterolobserved inHCwas confined to the atherogenic VLDL- and LDL-sizedparticles (Fig. 1d). Addition of epicatechin to the diet had no effect ontotal plasma cholesterol levels and lipoprotein profile, except a smalltransient decrease in total cholesterol after 4 weeks of treatment. Inline with its well-known lipid-lowering properties, atorvastatintreatment strongly and rapidly decreased plasma cholesterol andlevels remained low until the end of the study (average8.38 � 1.60 mM, a 54% reduction). Atorvastatin treatment reducedcholesterol in the VLDL and LDL-sized particles specifically. Plasmatriglyceride levels were not affected by epicatechin and were slightlyreduced by atorvastatin treatment (Supplement 2).
These results indicate that the anti-atherogenic effect of epi-catechin is independent of an effect on plasma lipids. We thereforecontinued by investigating the effects of epicatechin on inflam-mation, a second important risk factor in atherosclerosis.
3.2. Epicatechin does not affect monocyte adhesion to theendothelium
An important inflammatory process in atherosclerosis is theattachment of monocytes to the endothelial cell layer, which ismediated by adhesion molecules. Epicatechin feeding did notreduce plasma levels of the endothelial cell adhesion molecules sE-selectin and sVCAM-1 (Supplement 3) and did not reduce mono-cyte expression of the adhesion molecules CD11a, CD11b, CD49dand CD162 as analysed by flow cytometry (Supplement 3). In linewith these findings, histological quantification of the number ofmonocytes adherent to the endothelial cell layer revealed thatmonocyte adhesion to the endothelium was not decreased inepicatechin-treated animals (not shown).
3.3. Epicatechin attenuates diet-induced expression of circulatinginflammatory markers
Epicatechin-treated animals showed lower SAA levels than HCthroughout the study period, and this effect reached significance atweek 16 (Fig. 2a). Because SAA, secreted by the liver, can participatein disease progression throughout the development of atheroscle-rosis, the total SAA exposure (area under the curve) was calculated.SAA exposure was also significantly reduced compared with HC(Fig. 2b). This anti-inflammatory effect was substantiated in anindependent experiment in mice expressing human-CRP, anotherliver-derived acute phase response protein and CVD risk factor. Incontrol animals, HF-feeding significantly induced plasma human-CRP levels, while supplementation with epicatechin completelyprevented this diet-induced increase in plasma human-CRP(Fig. 2c). Together, these data demonstrate that epicatechinquenched the formation of the liver-derived pro-atherogenic fac-tors SAA and human-CRP.
3.4. Effect of epicatechin on lesion severity in early- and late-stageatherosclerosis
To provide insight into which stage of the atherosclerotic dis-ease process may be affected by epicatechin, we performed arefined morphological analysis of lesion severity. This analysisrevealed that the anti-atherosclerotic effect of epicatechin wasspecific to the severe lesions (type IV and V), the area of which was27% lower in HCE than in HC (Fig. 3a). There was no differencebetween HC and HCE animals in the total area of mild lesions (typeI-III, Fig. 3a), but epicatechin-treated animals did have significantlymore lesion-free segments (10% in HCE compared with 2% in HC,Fig. 3b). To investigatewhether this effect is attributable to an effectof epicatechin on early atherosclerosis development, E3Lmicewere
fed the HC or HCE diet for 8 weeks, which resulted in a greaterrelative contribution of mild lesions to the total lesion area (Fig. 3c).There was no effect of epicatechin on the prevalence of mild lesionsand, consistent with the long-term experiment, epicatechin tendedto reduce the severe lesion types as well as the total lesion area(Fig. 3ced). The overall reduction in atherosclerotic lesion area was40% (Fig. 3d). Together, these data confirm the anti-atheroscleroticpotential of epicatechin and suggest a specific effect of epicatechinon the progression from mild to severe lesions, a transition that ischaracterised by progression of the atherosclerotic plaque into themedia of the vessel wall (representative photomicrographs shownin Fig. 3e). This lesion progression requires matrix remodelling,which is mediated by processes related to vascular inflammation.
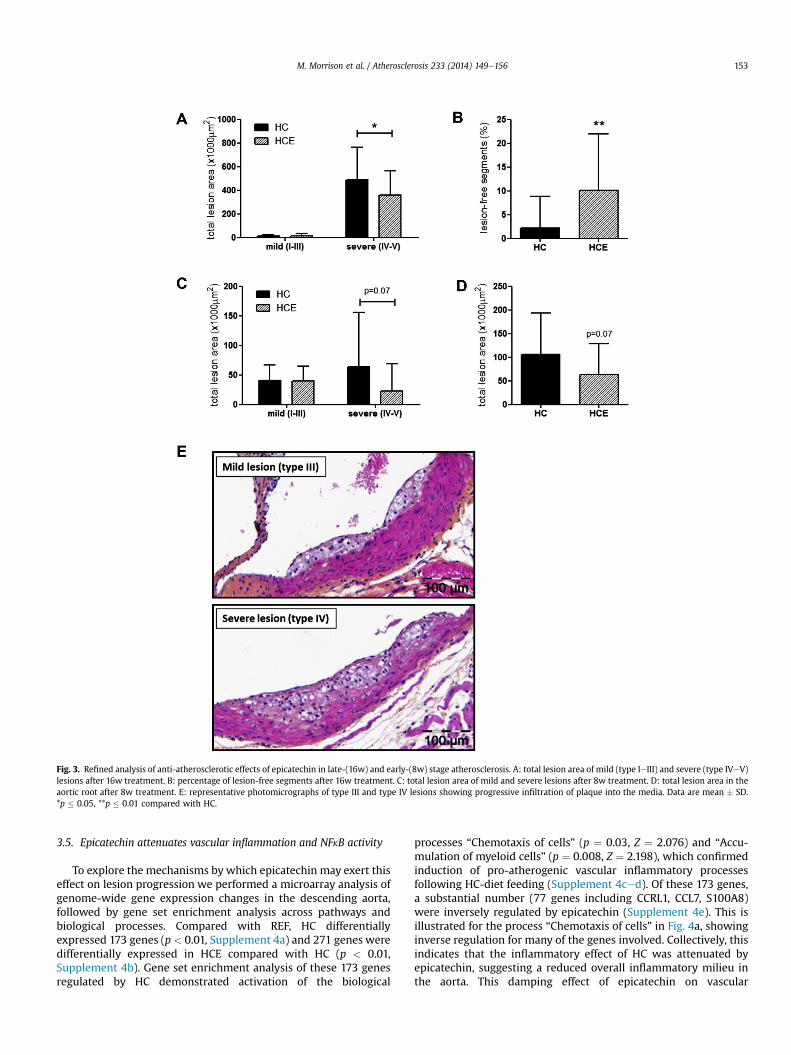
Fig. 3. Refined analysis of anti-atherosclerotic effects of epicatechin in late-(16w) and early-(8w) stage atherosclerosis. A: total lesion area of mild (type IeIII) and severe (type IVeV)lesions after 16w treatment. B: percentage of lesion-free segments after 16w treatment. C: total lesion area of mild and severe lesions after 8w treatment. D: total lesion area in theaortic root after 8w treatment. E: representative photomicrographs of type III and type IV lesions showing progressive infiltration of plaque into the media. Data are mean � SD.*p � 0.05, **p � 0.01 compared with HC.
M. Morrison et al. / Atherosclerosis 233 (2014) 149e156 153
3.5. Epicatechin attenuates vascular inflammation and NFkB activity
To explore the mechanisms by which epicatechin may exert thiseffect on lesion progression we performed a microarray analysis ofgenome-wide gene expression changes in the descending aorta,followed by gene set enrichment analysis across pathways andbiological processes. Compared with REF, HC differentiallyexpressed 173 genes (p < 0.01, Supplement 4a) and 271 genes weredifferentially expressed in HCE compared with HC (p < 0.01,Supplement 4b). Gene set enrichment analysis of these 173 genesregulated by HC demonstrated activation of the biological
processes “Chemotaxis of cells” (p ¼ 0.03, Z ¼ 2.076) and “Accu-mulation of myeloid cells” (p ¼ 0.008, Z ¼ 2.198), which confirmedinduction of pro-atherogenic vascular inflammatory processesfollowing HC-diet feeding (Supplement 4ced). Of these 173 genes,a substantial number (77 genes including CCRL1, CCL7, S100A8)were inversely regulated by epicatechin (Supplement 4e). This isillustrated for the process “Chemotaxis of cells” in Fig. 4a, showinginverse regulation for many of the genes involved. Collectively, thisindicates that the inflammatory effect of HC was attenuated byepicatechin, suggesting a reduced overall inflammatory milieu inthe aorta. This damping effect of epicatechin on vascular

Fig. 4. Effects of epicatechin on vascular inflammation. AeB: inversely regulated genes in the process ‘chemotaxis of cells’ as determined by aortic microarray analysis and Ingenuitypathway analysis. A: subset of differentially regulated genes by HC compared with REF (all p < 0.01) showing upregulation (red) of most genes involved. B: downregulation (green)of the same subset of genes by HCE compared with HC (CCRL1: p < 0.01) indicating inverse anti-inflammatory effect of epicatechin. C: representative photomicrographs ofatherosclerotic lesions immunohistochemically stained for myeloperoxidase-positive neutrophils. D: quantification of lesional neutrophils in HC and HCE, showing reduced numberof neutrophils in HCE. Data are mean � SD. *p � 0.05 compared with HC.
M. Morrison et al. / Atherosclerosis 233 (2014) 149e156154
inflammation was substantiated by results from immunohisto-chemical staining of lesional neutrophils, which revealed adecrease in the number of neutrophils in lesions from epicatechin-treated animals (Fig. 4bec). The attenuation of HC-induced vascularinflammation by epicatechinwas accompanied by effects on severalgenes involved in matrix degradation and remodelling (Adam4,Adam21, Adam22, MMP12, MMP13 and TIMP1; all upregulated byHC, p< 0.05), the induction of which was globally quenched in HCE(Supplement 4f) providing further mechanistic support for theobserved effects on lesion progression.
As many of the quenched chemotaxis- and matrix remodelling-associated genes as well as the liver-derived factors SAA andhuman-CRP are controlled by the inflammatory master regulatorNFkB, we investigated whether epicatechin would quench HC diet-induced NFkB activation in vivo. An independent experiment wasperformed in NFkB-luciferase reporter mice using non-invasivebioluminescence imaging. Relative to baseline, HC diet-inducedactivation of NFkB was mainly observed in the abdominal andheart region (Fig. 5). Epicatechin supplementation to the diet
indeed prevented this activation of NFkB, and photon countsremained comparable to baseline levels. This further substantiatesthe anti-inflammatory capacity of epicatechin in vivo.
4. Discussion
In the present study, we investigated the effects of the flavanolepicatechin on diet-induced early- and late-stage atherosclerosisand the cardiovascular risk factors dyslipidaemia and diet-inducedinflammation. Previous studies investigating the effects of flavanol-rich foods have provided indications for potential cardioprotectiveeffects of these foods [18,19], but the effects of individual flavanolsremain unclear. Here, we have shown that epicatechin can atten-uate the development of atherosclerosis, independently of effectson plasma lipids. Refined analysis of aortic lesions at early and latestages of the pathogenesis showed that epicatechin specificallyreduces the progression frommild to severe lesions. This effect maybe mediated by attenuation of several HC-induced pro-atherogenicinflammatory factors in the liver (SAA, human-CRP) as well as

Fig. 5. Effects of epicatechin on diet-induced inflammation in NFkB-luciferase reportermice. A: representative capture of NFkB activity at baseline, after HC-feeding and HCE-feeding. B: quantification of whole-body NFkB activity relative to baseline (photoncounts/cm2/second). Data are mean � SD. #p � 0.05 for increase within group.
M. Morrison et al. / Atherosclerosis 233 (2014) 149e156 155
atherogenic processes in the aorta (e.g. immune cell chemotaxisand cellular movement; matrix remodelling), all of which may atleast partly be related to the observed quenching effect of epi-catechin on NFkB.
Our atherosclerotic lesion analysis showed that epicatechin canreduce the development of atherosclerosis, resulting in a reductionof total lesion area by 27%. The observed reduction in atheroscle-rosis was not accompanied by an effect of epicatechin on athero-genic lipids or lipoproteins. A similar lipid-independent anti-atherogenic effect has been reported for another polyphenolquercetin under comparable experimental conditions in E3L mice[26]. The pharmaceutical reference compound atorvastatin didreduce plasma cholesterol and atherosclerotic lesion development,thus confirming that cholesterol modulation is possible underthese conditions. These results suggest that the anti-atherogeniceffect of epicatechin may be mediated by attenuation of otherrisk factors such as inflammation.
Here, we demonstrate that epicatechin can mitigate diet-inducedinflammation in atherogenesis. In E3L mice, epicatechin attenuatedHC-diet-induced SAA expression. This acute phase protein is pre-dominantly formed in the liver, indicates increased CVD risk inhumans [32], and is considered a potential mediator of atherogenesis[33,34]. SAA may contribute to atherosclerosis progression by stim-ulating lipoprotein retention in the vessel wall [35] and by inducingthe expression of MMPs [36] involved in degradation of the extra-cellular matrix. Indeed, SAA directly accelerates atherosclerosisprogression independent of plasma lipids [37]. We analysed the ef-fect of epicatechin on liver-derived inflammatory factors in moredetail using human-CRP transgenicmice [38]. (N.B. mouse-CRP is notan inflammationmarker). Like SAA, human-CRP is a strong predictorof CVD [39] and pharmaceutical reduction of human-CRP levels re-duces risk of CVD independently of effects on plasma cholesterol[40]. The observed quenching effect of epicatechin on human-CRP
expression is in line with reports of lower plasma CRP levels inhumans consuming foods rich in epicatechin [7,8].
Refined histological analysis of lesion severity in late-stage aswellas early-stage atherosclerosis in E3L mice indicates that epicatechinaffects the progression of lesions from mild to severe lesion typesspecifically. This lesion progression is characterised by invasion ofthe plaque into the media of the vessel wall, a complex process thatis driven by local intravascular inflammation andmediated by factorsthat orchestrate chemotaxis and cellular movement, as well as byremodelling of the extracellular matrix. Microarray analysis of geneexpression in the aorta revealed a mitigating effect of epicatechin onHC-induced changes in aortic gene expression, including genes thatcontribute to the pathways crucial for chemotaxis and cellularmovement (e.g. CCRL1, CCL7, S100A8) and vascular remodelling(e.g. MMP12, MMP13). To our knowledge, microarray analysis of theeffects of epicatechin in aorta has not been performed previously.Notably, epicatechin did not act on a specific set of ‘target’ genes or asingle inflammatory process, but reduced the overall inflammatorytone within the vasculature, suggesting a damping effect on a moregeneric inflammatory event upstream of these genes. In reflection ofthis reduced inflammatorymilieu in epicatechin-treated animals, weobserved a reduction in the number of lesional myeloperoxidase-positive neutrophilic cells in HCE, a cell type that has been shownto accumulate most prominently in atherosclerotic plaques in re-gions of high inflammatory activity [41]. Furthermore, neutrophilsare a source of large amounts of matrix-degrading proteases that cancontribute to matrix degradation and remodelling and are recog-nised as a cell type that has an important contributory role inatherosclerosis progression (reviewed in Ref. [42]).
Many of the HC-induced genes quenched by epicatechin treat-ment are transcriptionally regulated by the inflammatory tran-scription factor NFkB. As the liver-derived factors SAA and human-CRP are also controlled by NFkB [43,44], we sought direct evidenceof the potential of epicatechin to inhibit NFkB activation in vivo.Imaging of NFkB activation in NFkB-luciferase reporter mice showedthat epicatechin can indeed prevent diet-induced activation of thistranscription factor, providing a potential lipid-independent mech-anismbywhich epicatechinmay exert its anti-atherosclerotic effects.Therapeutic benefits of NFkB inhibition have been demonstrated inearlier studies on the anti-atherogenic effects of systemic [45,46] andendothelial cell-specific [47] inhibition of NFkB, and pharmaceuticalattenuation of vascular NFkB activity resulted in reduced lesionprogression from mild towards complex severe lesions [46],consistent with the effects observed here. To our knowledge, noprevious studies have investigated the effects of epicatechin on NFkBactivation in vivo. Results from in vitro studies provide conflictingevidence for the potential NFkB-inhibitory effect of epicatechin [48e50] that may be explained by differences in the choice of cell type,stimulus for NFkB activation, concentration of epicatechin used andincubation and exposure times.
Together, the results of the present study provide evidence forbeneficial anti-inflammatoryeffectsof theflavanol epicatechin in liverand aorta and a reduction of atherosclerosis, with inhibited progres-sion to severe lesions. Our findings substantiate cardioprotective as-sociations from epidemiological studies on flavanol-rich foods andimplicate epicatechin as a possible mediator of these effects.
Conflict of interest
The authors declare that they have no conflict of interest.
Funding
The study was funded by TI Food and Nutrition, a public-privatepartnership on pre-competitive research in food and nutrition.

M. Morrison et al. / Atherosclerosis 233 (2014) 149e156156
The funders had no role in study design, data collection and anal-ysis, decision to publish, or preparation of the manuscript.
Acknowledgements
Wewould like to thank Wim van Duyvenvoorde, Erik Offerman,Karin Toet and Ivo Que for their excellent technical assistance.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.atherosclerosis.2013.12.027.
References
[1] Weber C, Noels H. Atherosclerosis: current pathogenesis and therapeuticoptions. Nat Med 2011;17:1410e22.
[2] Estruch R, Ros E, Salas-Salvado J, et al. Primary prevention of cardiovasculardisease with a Mediterranean diet. N Engl J Med 2013;368:1279e90.
[3] Nettleton JA, Polak JF, Tracy R, Burke GL, Jacobs Jr DR. Dietary patterns andincident cardiovascular disease in the Multi-Ethnic Study of Atherosclerosis.Am J Clin Nutr 2009;90:647e54.
[4] Siri-Tarino PW, Sun Q, Hu FB, Krauss RM. Saturated fatty acids and risk ofcoronary heart disease: modulation by replacement nutrients. Curr Athe-roscler Rep 2010;12:384e90.
[5] Chait A, Han CY, Oram JF, Heinecke JW. Thematic review series: the immunesystem and atherogenesis. Lipoprotein-associated inflammatory proteins:markers or mediators of cardiovascular disease? J Lipid Res 2005;46:389e403.
[6] Gregor MF, Hotamisligil GS. Inflammatory mechanisms in obesity. Annu RevImmunol 2011;l29:415e45.
[7] Esmaillzadeh A, Kimiagar M, Mehrabi Y, Azadbakht L, Hu FB, Willett WC. Fruitand vegetable intakes, C-reactive protein, and the metabolic syndrome. Am JClin Nutr 2006;84:1489e97.
[8] Holt EM, Steffen LM, Moran A, et al. Fruit and vegetable consumption and itsrelation to markers of inflammation and oxidative stress in adolescents. J AmDiet Assoc 2009;109:414e21.
[9] Detopoulou P, Panagiotakos DB, Chrysohoou C, et al. Dietary antioxidant ca-pacity and concentration of adiponectin in apparently healthy adults: theATTICA study. Eur J Clin Nutr 2010;64:161e8.
[10] van’t Veer P, Jansen MC, Klerk M, Kok FJ. Fruits and vegetables in the preventionof cancer and cardiovascular disease. Public Health Nutr 2000;3:103e7.
[11] Arts IC, Hollman PC. Polyphenols and disease risk in epidemiologic studies.Am J Clin Nutr 2005;81:317Se25S.
[12] Heiss C, Keen CL, Kelm M. Flavanols and cardiovascular disease prevention.Eur Heart J 2010;31:2583e92.
[13] Geleijnse JM, Launer LJ, Van der Kuip DA, Hofman A, Witteman JC. Inverseassociation of tea and flavonoid intakes with incident myocardial infarction:the Rotterdam Study. Am J Clin Nutr 2002;75:880e6.
[14] Kuriyama S. The relation between green tea consumption and cardiovasculardisease as evidenced by epidemiological studies. J Nutr 2008;138:1548Se53S.
[15] Buijsse B, Feskens EJ, Kok FJ, Kromhout D. Cocoa intake, blood pressure, andcardiovascular mortality: the Zutphen Elderly Study. Arch Intern Med2006;166:411e7.
[16] Larsson SC, Virtamo J, Wolk A. Chocolate consumption and risk of stroke: aprospective cohort of men and meta-analysis. Neurology 2012;79:1223e9.
[17] Ivey KL, Lewis JR, Prince RL, Hodgson JM. Tea and non-tea flavonol intakes inrelation to atherosclerotic vascular disease mortality in older women. Br JNutr 2013:1e8.
[18] Arranz S, Valderas-Martinez P, Chiva-Blanch G, et al. Cardioprotective effectsof cocoa: clinical evidence from randomized clinical intervention trials inhumans. Mol Nutr Food Res 2013;57(6):936e47.
[19] Hooper L, Kroon PA, Rimm EB, et al. Flavonoids, flavonoid-rich foods, andcardiovascular risk: a meta-analysis of randomized controlled trials. Am J ClinNutr 2008;88:38e50.
[20] Knaze V, Zamora-Ros R, Lujan-Barroso L, et al. Intake estimation of total andindividual flavan-3-ols, proanthocyanidins and theaflavins, their food sourcesand determinants in the European Prospective Investigation into Cancer andNutrition (EPIC) study. Br J Nutr 2012;108:1095e108.
[21] Arts IC, Jacobs Jr DR, Harnack LJ, Gross M, Folsom AR. Dietary catechins inrelation to coronary heart disease death among postmenopausal women.Epidemiology 2001;12:668e75.
[22] Noll C, Lameth J, Paul JL, Janel N. Effect of catechin/epicatechin dietary intakeon endothelial dysfunction biomarkers and proinflammatory cytokines inaorta of hyperhomocysteinemic mice. Eur J Nutr 2013;52:1243e50.
[23] Loke WM, Proudfoot JM, Hodgson JM, et al. Specific dietary polyphenolsattenuate atherosclerosis in apolipoprotein E-knockout mice by alleviating
inflammation and endothelial dysfunction. Arterioscler Thromb Vasc Biol2010;30:749e57.
[24] Zadelaar S, Kleemann R, Verschuren L, et al. Mouse models for atherosclerosisand pharmaceutical modifiers. Arterioscler Thromb Vasc Biol 2007;27:1706e21.
[25] Kleemann R, Verschuren L, van Erk MJ, et al. Atherosclerosis and liverinflammation induced by increased dietary cholesterol intake: a combinedtranscriptomics and metabolomics analysis. Genome Biol 2007;8:R200.
[26] Kleemann R, Verschuren L, Morrison M, et al. Anti-inflammatory, anti-proliferative and anti-atherosclerotic effects of quercetin in human in vitroand in vivo models. Atherosclerosis 2011;218:44e52.
[27] Carlsen H, Haugen F, Zadelaar S, et al. Diet-induced obesity increases NF-kappaB signaling in reporter mice. Genes Nutr 2009;4:215e22.
[28] Yakala GK, Wielinga PY, Suarez M, et al. Effects of chocolate supplementationon metabolic and cardiovascular parameters in ApoE3L mice fed a high-cholesterol atherogenic diet. Mol Nutr Food Res 2013;57(11):2039e48.
[29] Wielinga PY, Yakala GK, Heeringa P, Kleemann R, Kooistra T. Beneficial effectsof alternate dietary regimen on liver inflammation, atherosclerosis and renalactivation. PLoS One 2011;6:e18432.
[30] Verschuren L, Radonjic M, Wielinga PY, et al. Systems biology analysis un-ravels the complementary action of combined rosuvastatin and ezetimibetherapy. Pharmacogenet Genomics 2012;22:837e45.
[31] Manach C, Williamson G, Morand C, Scalbert A, Remesy C. Bioavailability andbioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies.Am J Clin Nutr 2005;81:230Se42S.
[32] Ridker PM, Hennekens CH, Buring JE, Rifai N. C-reactive protein and othermarkers of inflammation in the prediction of cardiovascular disease inwomen. N Engl J Med 2000;342:836e43.
[33] King VL, Thompson J, Tannock LR. Serum amyloid A in atherosclerosis. CurrOpin Lipidol 2011;22:302e7.
[34] O’Brien KD, Chait A. Serum amyloid A: the "other" inflammatory protein. CurrAtheroscler Rep 2006;8:62e8.
[35] O’Brien KD, McDonald TO, Kunjathoor V, et al. Serum amyloid A and lipo-protein retention in murine models of atherosclerosis. Arterioscler ThrombVasc Biol 2005;25:785e90.
[36] Lee HY, Kim MK, Park KS, et al. Serum amyloid A stimulates matrix-metalloproteinase-9 upregulation via formyl peptide receptor like-1-mediated signaling in human monocytic cells. Biochem Biophys Res Com-mun 2005;330:989e98.
[37] Dong Z, Wu T, Qin W, et al. Serum amyloid A directly accelerates the pro-gression of atherosclerosis in apolipoprotein E-deficient mice. Mol Med2011;17:1357e64.
[38] Ciliberto G, Arcone R, Wagner EF, Ruther U. Inducible and tissue-specificexpression of human C-reactive protein in transgenic mice. EMBO J 1987;6:4017e22.
[39] Kaptoge S, Di Angelantonio E, Lowe G, et al. for the Emerging Risk FactorsCollaboration. C-reactive protein concentration and risk of coronary heartdisease, stroke, and mortality: an individual participant meta-analysis. Lancet2010;375:132e40.
[40] Nissen SE, Tuzcu EM, Schoenhagen P, et al. Statin therapy, LDL cholesterol, C-reactive protein, and coronary artery disease. N Engl J Med 2005;352:29e38.
[41] Rotzius P, Thams S, Soehnlein O, et al. Distinct infiltration of neutrophils inlesion shoulders in ApoE-/- mice. Am J Pathol 2010;177:493e500.
[42] Soehnlein O. Multiple roles for neutrophils in atherosclerosis. Circ Res2012;110:875e88.
[43] Jensen LE, Whitehead AS. Regulation of serum amyloid A protein expressionduring the acute-phase response. Biochem J 1998;334(Pt 3):489e503.
[44] Kleemann R, Gervois PP, Verschuren L, Staels B, Princen HM, Kooistra T.Fibrates down-regulate IL-1-stimulated C-reactive protein gene expression inhepatocytes by reducing nuclear p50-NFkappa B-C/EBP-beta complex for-mation. Blood 2003;101:545e51.
[45] Chiba T, Kondo Y, Shinozaki S, et al. A selective NFkappaB inhibitor, DHMEQ,reduced atherosclerosis in ApoE-deficient mice. J Atheroscler Thromb2006;13:308e13.
[46] de Vries-van der Weij J, Toet K, Zadelaar S, et al. Anti-inflammatory salicylatebeneficially modulates pre-existing atherosclerosis through quenching of NF-kappaB activity and lowering of cholesterol. Atherosclerosis 2010;213:241e6.
[47] Gareus R, Kotsaki E, Xanthoulea S, et al. Endothelial cell-specific NF-kappaBinhibition protects mice from atherosclerosis. Cell Metab 2008;8:372e83.
[48] Mackenzie GG, Carrasquedo F, Delfino JM, Keen CL, Fraga CG, Oteiza PI. Epi-catechin, catechin, and dimeric procyanidins inhibit PMA-induced NF-kappaBactivation at multiple steps in Jurkat T cells. FASEB J 2004;18:167e9.
[49] Al-Hanbali M, Ali D, Bustami M, et al. Epicatechin suppresses IL-6, IL-8 andenhances IL-10 production with NF-kappaB nuclear translocation in wholeblood stimulated system. Neuro Endocrinol Lett 2009;30:131e8.
[50] Granado-Serrano AB, Martin MA, Haegeman G, Goya L, Bravo L, Ramos S.Epicatechin induces NF-kappaB, activator protein-1 (AP-1) and nucleartranscription factor erythroid 2p45-related factor-2 (Nrf2) viaphosphatidylinositol-3-kinase/protein kinase B (PI3K/AKT) and extracellularregulated kinase (ERK) signalling in HepG2 cells. Br J Nutr 2010;103:168e79.





























