Embed Size (px)
Citation preview

B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
. sc iencedi rec t . com
ava i lab le a t wwwjournal homepage: www.elsevier .com/ locate /b iocon
Evaluating ipe (Tabebuia, Bignoniaceae) logging in Amazonia:Sustainable management or catalyst for forest degradation?
Mark Schulzea,b,c,*, James Groganb,d, Chris Uhle, Marco Lentinia,b, Edson Vidalb,f
aSchool of Forest Resources and Conservation, P.O. Box 110760, University of Florida, Gainesville, FL 32611-0760, USAbInstituto do Homem e Meio Ambiente da Amazonia, R. Domingos Marreiros, No. 2020, Bairro Fatima, Belem, Para 66060-160, BrazilcInstituto Floresta Tropical, Caixa Postal 13077, Belem, PA 66040-970, BrazildYale School of Forestry and Environmental Studies, New Haven, CT 06511, USAeDepartment of Biology, 208 Mueller Lab, Pennsylvania State University, University Park, PA 16802, USAfESALQ/USP, Avenue Padua Dias, 11, Departamento de Ciencias Florestais, Sao Dimas, CEP 13418-900, Piracicaba/SP, Brazil
A R T I C L E I N F O
Article history:
Received 27 February 2008
Received in revised form
28 May 2008
Accepted 2 June 2008
Available online 21 July 2008
Keywords:
CITES
Population structure and dynamics
Post-logging recovery
Reduced-impact logging
Sustained yield
Tabebuia
0006-3207/$ - see front matter � 2008 Elsevidoi:10.1016/j.biocon.2008.06.003
* Corresponding author: Address: School of32611-0760, USA. Tel.: +1 573 268 0714.
E-mail addresses: [email protected] (M. S(M. Lentini), [email protected] (E. Vidal).
A B S T R A C T
Prized for their dense, rot-resistant wood, Tabebuia impetiginosa and T. serratifolia (vernacular
name = ipe) are among the most valuable Amazonian timbers. We analyzed the geograph-
ical extent, spread and trajectory of ipe logging in Brazilian Amazonia, and evaluated har-
vest pressure on this forest resource. We also examine Tabebuia population response to
reduced-impact logging, a more ecologically benign alternative to destructive conventional
harvest practices in Amazonia. Based on eight years of population monitoring at multiple
sites in the eastern Brazilian Amazon, we project second harvest ipe yields in forests logged
using RIL under legally allowable (90% of commercial stems) and reduced (70%) harvest
intensities.
In recent years ipe harvests have declined or ceased in the majority of old logging fron-
tiers in eastern Amazonia while spreading to new logging frontiers in central and south-
western Amazonia. With current timber market prices, transportation infrastructure and
harvesting costs, logging of ipe would be profitable in an estimated 63% of the Brazilian
Amazon; in the more remote logging frontiers only logging of ipe and a few other high-
value timbers is currently profitable. All populations of T. impetiginosa and T. serratifolia in
northeastern forests showed drastic population declines over multiple RIL harvests in sim-
ulations, with no indication of population recovery over the long term. We conclude from
study of Tabebuia populations in eastern Amazonia and modeling of response to logging
that these two species are endangered by logging activity and merit additional protection
under forest legislation.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Since the 1980s logging has been an important catalyst for
deforestation in many regions of Amazonia. Much of the
er Ltd. All rights reserved
Forest Resources and Co
chulze), jgrogan@crocker
deforestation along the southern rim of Amazonia has been
facilitated by loggers penetrating primary forests in search
of sparsely distributed big-leaf mahogany (Swietenia macrophy-
lla) trees, in the process providing access to ranchers and
.
nservation, P.O. Box 110760, University of Florida, Gainesville, FL
.com (J. Grogan), [email protected] (C. Uhl), [email protected]

Table 1 – Timber exports in 2004 from eight statescomprising the Brazilian Legal Amazon showing thedollar value of ipe exportsa and the percent of total ipeexports by dollar value
State Total exports(US$)
Ipe exportsb
(US$)Ipe %
of total
Acre 5,440,552 124,316 2.3
Amapa 42,311,249 – –
Amazonas 24,139,739 314,467 1.3
Maranhao 12,681,694 208,458 1.6
Mato Grosso 197,596,200 30,857,124 15.6
Para 543,441,974 31,811,577 5.9
Rondonia 113,456,363 19,496,032 17.2
Roraima 3,660,606 45,618 1.2
Legal Amazon 942,728,377 82,857,592 8.8
a Source: SECEX (2005).
b Tabebuia are generally not distinguished commercially by spe-
cies. Although ipe may refer to any species in the genus, T. impeti-
ginosa and T. serratifolia are the most commonly logged species in
Amazonia.
2072 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
farmers who transform forests to pasture and farm fields
(Verıssimo et al., 1995; Fearnside, 1997; Asner et al., 2005,
2006). Today, mahogany is recognized as a threatened species
under Appendix II of the Convention on International Trade
in Endangered Species; significant commercial populations
of mahogany exist only in remote regions of Amazonia, pri-
marily in the southwest (Grogan et al., 2008). Meanwhile,
the logging industry has migrated from the impoverished
landscapes along the so-called arc of deforestation to new for-
est frontiers in central and western Amazonia, pursuing high-
value timbers that occur in these regions and are not subject
to the legal protections belatedly bestowed upon mahogany.
Many of the characteristics rendering mahogany vulnerable
to commercial extinction are shared by other high-value tim-
ber species in Amazonia (Grogan et al., 2008).
Efforts to transform an uncontrolled and destructive Ama-
zonian logging industry into an engine for sustainable eco-
nomic development have achieved tangible, if limited,
successes: certified ‘well-managed’ forest area has increased
to >5 million ha since Forest Stewardship Council (FSC) certi-
fication arrived in the region in the 1990s (primarily in Bolivia
and Brazil; FSC, 2007); a growing segment of the timber indus-
try has demonstrated interest in reduced-impact logging (RIL)
techniques as an economically viable alternative to haphaz-
ard conventional or predatory logging; and big-leaf mahogany
was listed on Appendix II of the UN-chartered Convention on
International Trade in Endangered Species of Fauna and Flora
(CITES) in 2002, necessitating more stringent harvest regula-
tions and tighter controls on timber exports (Grogan and Bar-
reto, 2005). This progress notwithstanding, the vast majority
(95–98%) of timber harvesting in Amazonia continues to occur
by conventional logging that is often a precursor to forest
conversion to ranching or farming (Lentini et al., 2003, 2005;
Verıssimo and Barreto, 2004). This boom-and-bust exploita-
tion cycle has been well described elsewhere: loggers migrate
to new frontiers as timber stocks in historic logging centers
dwindle, instigating new cycles of timber mining, forest clear-
ing, and land conflicts in what were previously intact forests
on unclaimed public lands (Uhl et al., 1991, 1997; Verıssimo
et al., 1992, 1995, 2002a; Grogan et al., 2002; Verıssimo and Bar-
reto, 2004). The pursuit of high-value timber species for ex-
port markets drives much of this expansion into heretofore
unlogged regions of Amazonia (Verıssimo et al., 1995; Lentini
et al., 2005).
Tabebuia (ipe, pronounced ‘‘ee-pay’’) are the new mahog-
any. These are prized neotropical timber species whose
dense, rot-resistant wood is exported primarily to North
America for use in boardwalks and residential decking.
Although the commercial name ipe encompasses several Tab-
ebuia species, T. impetiginosa and T. serratifolia comprise the
bulk of ipe extraction from Amazonia and are the focus of this
study. Widely advertised on websites for outdoor construction
materials, ipe is the most common tropical ‘species’ in the $3
billion residential decking market in the US, and its domi-
nance is expected to continue for the near future as restric-
tions on chemical treatment of woods reduce market share
of temperate zone species (Metafore, 2004). Responding to ex-
port prices (FOB – Free On Board) ranging from US$ 400 to
500 per m3 of sawn timber (Lentini et al., 2005), logging com-
panies profitably extract ipe from remote Amazonian regions
where timber harvests would not otherwise be feasible.
Meanwhile export markets for Tabebuia are well developed
and growing: there was a 500% increase from 1998 to 2004
in ipe timber exports from the Brazilian Amazon, and ipe ac-
counted for ca. 9% of the total value of wood exports from the
Brazilian Amazon in 2004 (Table 1; SECEX, 2005).
Conventional selective logging in the Brazilian Amazon
typically resembles a mining operation characterized by poor
planning and execution. Over-exploitation of commercial
species combined with excessive damages inflicted on forest
stands during harvest can compromise future production po-
tential as well as the ecological integrity of logged forests
(Johns et al., 1996; Holdsworth and Uhl, 1997; Uhl et al.,
1997; Nepstad et al., 1999; Vidal, 2004). Reduced-impact log-
ging practices mitigate forest-level damages and improve
operational cost effectiveness through improved inventory
and infrastructural planning (Barreto et al., 1998; Boltz et al.,
2001; Holmes et al., 2002). Yet even operating under RIL guide-
lines, loggers can legally harvest up to 90% of commercial-
sized individuals of a given species in Brazil on the assump-
tion that future harvests will be supplied by recruitment of to-
day’s sub-merchantable individuals (juvenile and pole-sized
trees, saplings, and seedlings) into commercial size classes
during proposed 25–35 yr cutting cycles (Brazil, 2006). Poor
logging practices and disregard for basic principles of sus-
tained resource use may lead to depletion and regional com-
mercial extinction of desirable timber species, as has been
observed for mahogany throughout much of its natural range
(Rodan et al., 1992; Verıssimo et al., 1995; Gullison et al., 1996;
Snook, 1996; Grogan et al., 2002; Kometter et al., 2004).
In this paper we describe mounting logging pressure on
natural Tabebuia populations in the Brazilian Legal Amazon.
We examine geographical range, density patterns, population
dynamics, and prospects for sustained timber yield at current
and hypothetical extraction rates for the two most widely dis-
tributed and heavily exploited ipes, T. impetiginosa (ipe roxo)
and T. serratifolia (ipe amarelo). We ask: what is the geograph-
ical extent and annual rate of Tabebuia exploitation in Brazil-

B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2073
ian Amazonia? Can ipe populations persist in the face of cur-
rent exploitation rates? Would reduced-impact logging sys-
tems adequately manage these species if widely adopted
across this vast region? Finally, we consider ipe logging in
the context of regional conservation issues, and propose mea-
sures that could reduce logging pressure on these valuable
species.
2. Methods
The questions above demand synthesis of several data
sources. Geographical distribution and abundance patterns
across the Amazon are critical to understanding Tabebuia
ecology, and also inform analyses of current and potential
logging pressure. Population structures from large-scale
inventories at sites spanning the Brazilian Amazon and mul-
tiple-year observations of population dynamics at a subset of
these sites allow for projections of Tabebuia population re-
sponse to logging and ipe timber yields under current and po-
tential harvest regulations. Official statistics on Brazilian ipe
exports in different years can be used to identify trends in
ipe harvests and to generate estimates of the minimum forest
area logged annually to supply export volume. Detailed sur-
veys of timber processing centers across the Brazilian Ama-
zon, including production data, timber prices and extraction
and transport costs, show trends in geographical patterns of
ipe exploitation and are the foundation for models of the
profitability of ipe extraction across the Amazon. In combina-
tion, these analyses provide a first assessment of the impact
logging has had on Tabebuia populations, the potential to reg-
ulate sustainable ipe harvests, and the relationship between
demand for ipe timber and the spread of logging to new forest
frontiers. Below, we detail the sources and methods used to
acquire and analyze the datasets for each component of this
study.
2.1. Study species
Tabebuia impetiginosa and T. serratifolia occur from Brazil’s
Atlantic Forest through the core region of Amazonia, into
Central America and up to and including areas of Mexico.
Both species are canopy emergent trees attaining heights up
to 50 m and stem diameters approaching 2 m. Adult morphol-
ogy and wood anatomy vary markedly across Brazilian
Amazonia in response to as yet poorly understood climatic
and edaphic factors. Regional populations along the southern
and southeastern Amazonian rim tend to be shorter in stat-
ure with less desirable timber characteristics. Both species oc-
cur at much reduced stature in central Brazilian cerrado
(savanna), and again as large trees in remnant Atlantic Forest
fragments. T. impetiginosa is also common in the vast Pantanal
wetland straddling Brazil’s border with Bolivia (Hasse and Hir-
ooka, 1998).
Both ipes considered here are deciduous species renowned
for spectacular floral displays on leafless crowns (purple and
yellow for T. impetiginosa and T. serratifolia, respectively). Seeds
are small, winged, and wind-dispersed, produced annually or
supra-annually, with limited dispersal range (ca. 150 m; Schu-
lze, 2003). Germination occurs rapidly with the first rains after
dispersal; no year-to-year soil storage is possible. Seedlings
are light demanding, requiring some level of forest canopy
opening for optimal sustained growth. Reproductive ecology
and age or stem size at first fruiting are poorly understood
for both species. As well, little is known about age- or size-
specific growth and mortality rates by natural populations
(Gentry, 1992; Justiniano et al., 2000).
2.2. Distribution and abundance
Species distributions in the Brazilian Amazon were mapped
based on RadamBrasil (1974) plot-level and sub-region-level
data plus collection lists from herbaria (Gentry, 1992; MOBOT,
2005; NYBG, 2005). RadamBrasil is an extensive resource
inventory conducted in the 1970s by the Brazilian Ministry
of Mines and Energy, covering the entire Legal Amazon and
including forest tree (P30 cm diameter) inventories in 2,364
1-ha plots. Tabebuia density patterns are estimated based on
data from RadamBrasil (1974) and Schulze (2003). Overall spe-
cies ranges were estimated by plotting all RadamBrasil census
plots that contained the species and the locations of all her-
barium specimen collections in a GIS database (ArcView 3.2;
ESRI, 1999). For both species, known locations spanned the
entire Brazilian Amazon, although this does not imply that
the species are distributed uniformly within the range. Den-
sity patterns within the RadamBrasil dataset were used to
tentatively demarcate regions of relatively high, medium
and low mean densities. The data provide a coarse view of
density patterns, but are limited by the relatively small size
and number of sample plots.
2.3. Population structures and dynamics
Tabebuia population structures were examined in seven large-
scale (204–11,370 ha) inventories at sites that span the Brazil-
ian Amazon (Table 2). While all sites receive <2200 mm of
rainfall annually and are seasonally dry during 1–6 months
of the year, underlying geomorphology and soil physical and
chemical properties vary widely among sites (IBGE, 2003;
Schulze, 2003; Grogan and Galvao, 2006). As a consequence,
forest structure ranges from tall closed evergreen canopies
in eastern forests (sites A–C) to tall open evergreen forests
in the southwest (E, G), low broken semi-evergreen forest in
the southeast (D), and open semi-deciduous forest in Rondo-
nia (F). At two of these sites (F, G), commercial inventories
only included stems >30 cm diameter, and all Tabebuia species
were lumped under the commercial name ipe; these invento-
ries only provide information on the ratio of commercial to
sub-merchantable stems available to loggers. At the remain-
ing sites, Tabebuia impetiginosa and T. serratifolia (and other
Tabebuia species) were distinguished, and lower minimum
inventory diameters (5 cm or 20 cm for site E) yield more com-
plete information on species populations. At the three east-
ern sites (A–C), stratified subsampling of smaller size classes
provided full population structures, from seedling to adult.
Analyses of population dynamics are based on data from
study of Tabebuia seedlings, juveniles, and adult trees in un-
logged and logged forests at four sites in the eastern Amazo-
nian state of Para (sites A–C and Moju at 2�11’S, 48�49’W).
Forest area included in the study totaled 1625 ha (Table 3).
At each site, sample populations were censused for growth,

Ta
ble
2–
Ta
beb
uia
inv
en
tory
site
sin
the
Bra
zili
an
Am
azo
n
Sit
ea
Mu
nic
ipa
lity
,S
tate
Lo
cati
on
Inven
tory
are
a(h
a)
Sp
eci
es
iden
tifi
cati
on
T.im
pet
igin
osa
den
sity
(>30
cmh
a�
1)
T.se
rrati
foli
ad
en
sity
(>30
cmh
a�
1)
A.
Faze
nd
aA
gro
sete
Pa
rago
min
as,
Pa
ra3
�00’
S,
47
�20’
W210
Bo
tan
ica
l0.1
60.3
1
B.
Faz.
Ca
ua
xi
Go
ian
esi
a,
Pa
ra3
�40’
S,
48
�20’
W1200
Bo
tan
ica
l0.0
80.1
0
C.
Flo
na
Ta
pa
jos
km
83
Belt
err
a,
Pa
ra3
�04’
S,
54
�15’
W400
Bo
tan
ica
l0.1
90.3
0
D.
Ma
rajo
ara
Pa
ud
’Arc
o,
Pa
ra7
�50’
S,
50
�16’
W204
Bo
tan
ica
lN
ot
pre
sen
t0.3
1
E.
Faz.
Sa
oJo
rge
Sen
aM
ad
ure
ira
,A
cre
9�
25’
S,
68
�38’
W685
Bo
tan
ica
lN
ot
pre
sen
t0.4
5
F.Fa
z.Im
acu
lad
aII
Ch
up
ingu
aia
,R
on
do
nia
12
�43’
S,
61
�00’
W1015
Co
mm
erc
ial
All
ipe
spp
=1.1
4
G.
Faz.
Seri
nga
lLıb
rea
,A
ma
zon
as
8�
44’
S,
68
�59’
W’
11,3
70
Co
mm
erc
ial
All
ipe
spp
=0.0
4
aS
ites
A–D
inven
tori
ed
for
stu
dy
of
tim
ber
speci
es
po
pu
lati
on
dy
na
mic
s(S
chu
lze,
2003;G
roga
n,u
np
ub
lish
ed
);si
teE
inven
tori
ed
as
pa
rto
fa
stu
dy
on
ma
na
gem
en
to
fSw
iete
nia
macr
oph
ylla
(Gro
ga
n,
un
pu
bli
shed
);si
tes
Fa
nd
Gin
ven
tori
ed
for
aco
mm
erc
ial
tim
ber
op
era
tio
n(R
.O
liveir
a,
un
pu
bli
shed
).
2074 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
mortality and recruitment annually for 3-5 yr beginning in
1993, 1996, or 1997 and at supra-annual (bi- or tri-annual)
intervals up through 2005. Short-term (2-yr) data from site D
were not included in analyses, but were used along with pub-
lished rates from other studies for comparison of mean Tab-
ebuia growth rates recorded across the Amazon.
As part of a larger study of commercial species regenera-
tion, T. impetiginosa and T. serratifolia seedlings and saplings
were inventoried in seven logged, 100-ha forest blocks at sites
A–C, with at least 60 felling gaps sampled per block and 1 ha
of skid trails per block at sites A and B, for a total of 540 gaps
and 5.6 ha of skid trails. In 360 one- to three-yr-old logging
gaps, seedling presence and density were compared to adult
distributions, and overall seedling density per ha in each har-
vest block was estimated based on this relationship and the
distribution of all logging gaps relative to adults (Schulze,
2003). Tabebuia seedlings and saplings were tagged and cens-
used annually for growth and mortality in 480 gaps over a per-
iod of 6 yr. Experimental introduction of seeds and seedlings
into 29 logging gaps at site B provided additional information
on seedling growth rates and mortality. Methods and results
of post-logging regeneration studies are detailed elsewhere
(Schulze, 2003; Schulze, 2008). Here we use information on
Tabebuia gap colonization and recruitment to model popula-
tion recovery following logging.
In the presentation and modeling exercises to follow, den-
sities and population structures of the two species are treated
separately, while growth and mortality rates are pooled due to
lack of significant differences between species.
2.4. Modeling population response to logging and volumeaccumulation
We constructed projection models of second-harvest poten-
tial for Tabebuia under best-practices RIL by combining data
on population structure and dynamics with observations on
gap colonization and recruitment rates (Table 4). At three of
four study sites we also collected data on logging intensity
and impacts on Tabebuia populations. Models were applied
to representative populations of each species in 1000-ha man-
agement blocks at each site. Starting population structures
were derived using means from 200- to 400-ha tree samples
and 20- to 40-ha juvenile sub-samples.
First-harvest intensity (year 0 in simulations) was as-
sumed to be either 90% of trees P50 cm diameter or 70% of
trees P70 cm diameter, with trees smaller than the chosen
minimum felling diameter considered sub-merchantable.
The higher harvest intensity and lower minimum felling
diameter are typical of Tabebuia harvests we have observed
in eastern Amazonia, and are consistent with Brazilian regu-
lations stipulating 10% seed tree retention. The lower harvest-
ing intensity and higher minimum felling diameter simulates
a biologically realistic alternative to current logging practices,
and is similar to the approach taken to regulate harvests of
threatened big-leaf mahogany in the Brazilian Amazon and
elsewhere (Brazil, 2003; Grogan et al., 2008; Grogan and
Schulze, 2008).
Commercially defective trees, those with major stem hol-
lows or poor form, may or may not be logged. In RIL opera-
tions, hollow trees are often left standing if little

Table 3 – Total sample sizes for Tabebuia impetiginosa and T. serratifolia at three sites* in eastern Amazonia
Size class – forest environment Sample areaa Sample # (Tabebuia stems)
impetiginosa serratifolia
Trees P10 cm diam – unlogged forest 1000 ha 112 128
Trees P10 cm diam – logged forest 625 ha 40 50
Juvenilesb – unlogged forest 100 ha 9 12
Juveniles – logged forest 10 ha + 590 logging gaps 139 117
Established seedlingsc – unlogged forest 75 ha 31 44
Established seedlings – 1–6 yr logged forest 5 ha skid trails + 350 gaps 396 591
Established seedlings – 10–15 yr logged forest 240 logging gaps 9 14
1st year seedlingsd – unlogged forest 15 crown shadows/sp. 2573 2525
1st year seedlings – logged forest 71 logging gaps 398 3131
a Area used for quantitative sample of each stem size class. Juveniles and seedlings were added to growth monitoring as encountered outside
quantitative sample plots.
b Juveniles are stems P2 cm and <10 cm diameter.
c Established seedlings are P50 cm height and <2 cm diameter.
d 1st year seedlings are those monitored from germination through the end of the 1st year of growth.
* Sites A–C used for study of Tabebuia population dynamics.
Table 4 – Observed mortality and growth rates for Tabebuia in unlogged and logged forests in Para, Brazil*
Size class – forest environment Annual mortality rate Annual growth ratea
Trees P10 cm diameter – unlogged forest 0.7% 0.19 (0.33) ± 0.22
Trees P10 cm diameter – logged forest 2.5% (yr 1–3); 1.3% (yr 3–8) 0.31 (0.47) ± 0.28
Juvenilesb – unlogged forest 9.2% 0.19 (0.32) ± 0.08
Juveniles – logged forest 6.3% 0.43 (0.60) ± 0.27
Established seedlingsc – unlogged forest 16.4% 0.09 (0.25) ± 0.03
Established seedlings – 1–6 yr logged forest 11.9% 0.36 (0.56) ± 0.30
Established seedlings – 10–15 yr logged forest 9.7% 0.09 (0.14) ± 0.13
1st year seedlingsd – unlogged forest 81–97% 0.05 (0.10) ± 0.02
1st year seedlings – logged forest 5–22% 0.43 (0.57) ± 0.23
a Diameter growth (cm) reported for all size classes except 1st year seedlings (height growth in meters). Numbers in parentheses are 75th
percentile values, ± SD.
b Juveniles are stems P2 cm and <10 cm diameter.
c Established seedlings are P50 cm height and <2 cm diameter.
d 1st year seedlings are those monitored from germination through the end of the 1st year of growth.
* Growth and mortality data are from sites A–C.
B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2075
commercial value is expected. However, for high-value spe-
cies there remains a strong incentive to fell trees with hollows
at the base in the hope of obtaining a sound log near the
crown (Grogan et al., 2008). It is therefore unclear whether
Tabebuia trees are more commonly spared or felled in RIL
operations when a hollow is detected at the base. Further-
more, minimum seed tree retention requirements may be
met with sound or hollow stems; some forest certifiers explic-
itly require loggers to designate sound trees, but otherwise
this aspect of seed tree retention remains an open question
in Brazil. In the first round of projections, we assumed that
hollows were as common in Tabebuia populations as for com-
mercial trees as a group, that hollow trees were not felled, and
that all seed trees were free of defect. Before simulating har-
vests, we randomly selected 25% of trees P50 cm diameter for
removal from harvest consideration due to commercially
defective (hollow) stems (Holmes et al., 2002; Gourlet-Fleury
et al., 2004; Zweede, personal communication). Lacking com-
parable data on the rate of defect in sub-merchantable popu-
lations, we opted for the conservative approach of assuming
all sub-merchantable stems were of good form and free of
hollows. Inventories from the study sites have found 12–37%
of sub-merchantable stems with obvious defects, with an
additional unknown fraction possessing cryptic hollows (i.e.,
undetectable in inventories) that may compromise merchant-
ability if they reach harvestable size (Valle et al., 2006; Schulze
and Vidal, unpublished data; Zweede, personal communica-
tion). Thus, all of our second harvest projections likely overes-
timate timber volume to some degree. In subsequent runs, we
tested the effect of alternative assumptions about hollows by
simulating harvests in which: (A) hollow trees occurred as
above, but seed tree retention rules could be met by designat-
ing hollow trees as seed trees; and (B) all Tabebuia trees were
considered defect free (Supplemental Table 1).
Growth and mortality rates of trees surviving the first har-
vest were modeled under our best estimate of initially ele-
vated post-logging population-level growth (75th percentile
annual diameter increment in forest samples, by life stage)
and mortality rates, with growth rates declining after 10 yr
to the median observed increment, and tree mortality declin-
ing after 3 yr and again after 10 yr (Supplemental Table 2).
Projected mortality rates for smaller size classes began with

2076 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
observed rates for each class, with rates changing as the co-
hort was projected to transition from one size class to the
next. These growth and mortality patterns provide the most
realistic estimate of post-logging population response given
currently available data (Silva et al., 1995, 1996; de Graaf
et al., 1999; de Graaf, 2000; Oliveira, 2000; Schulze, 2003; Vidal,
2004; Oliveira, 2005). An additional growth scenario was mod-
eled using optimistic assumptions: applying the mean 75th
percentile diameter growth rate (0.42 cm yr�1) throughout
the projection period, that is, no decline in growth rate over
time (total mortality in smaller size classes was also reduced
as trees transitioned between size classes more rapidly).
Our assumption of equal growth rates for all stems in a
size class is a deliberate simplification of growth patterns.
Tree populations display wide variation in growth rates; thus,
our projection likely overestimates transition rates among
size classes, while underestimating the growth of outstanding
individuals. For purposes of estimating volume and popula-
tion recovery under legal harvests, we favored simple projec-
tions over a more sophisticated model with more
assumptions. The high-growth scenario simulates the
maximum conceivable growth response, as all individuals
are essentially considered outstanding specimens, and there-
fore captures the most optimistic post-logging recovery
outcome.
To project second cutting cycle Tabebuia timber yields, 90%
and 70% harvestable volume of non-defective commercial-
sized adults was calculated after 30 and 60 yr (two possible
cutting cycles). Commercial stem volume per diameter size
class was estimated using the mean of four volume equa-
tions:V = e�7.62812+2.1809 · ln(diam) (Silva and Carvalho, 1984);
V = 0.0775 + 0.05179 · (diam2 · CommHgt) (Queiroz, 1984 cited
in Rolim et al., 2006); V = BasalArea · CommHgt · 0.7(Hei-
nsdijk and Bastos, 1963; Brown et al., 1989); and
V ¼ 0:5179� D100
� �2CH þ 0:0775 (Phillips et al., 2004). We pres-
ent the total harvestable volume in each scenario, although
presumably loggers would be subject to the same retention
criteria in the second harvest as in the first, meaning actual
harvests would be roughly 10% (or 30%) lower than the har-
vestable volumes we report.
2.5. Projecting long-term population recovery
The recovery of Tabebuia populations over the long term will
be dependent on regeneration in logged forest stands. Projec-
tions of growth and recruitment of existing stems as de-
scribed above demonstrated that low densities, slow growth,
and high mortality of existing juveniles in the forest under-
story result in extremely low recruitment rates to adult size.
We assume seedling recruitment to be largely restricted to
logging gaps and other disturbed zones within the 100-ha
management block (11% of total area). This assumption is
based on recorded first-year mortality rates of 90% for seed-
lings establishing in the understory (with 4-yr mortality of
100% for shaded seedlings; Schulze, 2003). Stems recruiting
in logging gaps and other disturbed sites will thus be largely
responsible for the eventual completion of a harvest rotation,
and for the long-term persistence of Tabebuia populations
subjected to logging. This recruitment in logging gaps will
take longer than the 30- and 60-yr periods relevant to second
harvest projections, and therefore was modeled separately
from volume recovery.
Three estimates of the time required for seedlings to re-
cruit to commercial size were made by applying the following
growth rates observed in logging gaps during 6 yr following
harvest: (1) mean growth rate of all Tabebuia stems; (2) 90th
percentile growth rate; and (3) maximum observed growth
rate. Projections of stem sizes at year 15 were also compared
to observed sizes of Tabebuia stems in 15-yr-old logging gaps.
Projections of the number of stems surviving to commercial
size in a typical 100-ha management block were then made
by applying observed annual mortality rates to three esti-
mates of total seedling number per block based on: (1) overall
mean density per hectare; (2) lower 95% confidence interval of
mean density; and (3) upper 95% confidence interval of mean
density. Observed gap mortality rates were applied for 10, 20
and 30 yr before assuming that rates dropped to those ob-
served for adult Tabebuia trees.
2.6. Exploitation rates and regional logging pressure
The total ipe roundwood harvest for export in 2004 was esti-
mated by converting reported export weight (158,628,644 kg;
SECEX, 2005) to export volume at 850-1050 kg m�3 (Souza
et al., 1997; Loureiro et al., 2000). Conversion of export volume
to harvested roundwood volume was based on 42% process-
ing efficiency and 36% of processed wood meeting export
standards (Lentini et al., 2003, 2005; Sobral, personal
communication).
The forest area required to meet this export demand was
estimated by dividing per hectare ipe yield into estimated to-
tal roundwood harvest volume. Yield per hectare was deter-
mined based on estimates of 0.34 commercial-sized trees of
Tabebuia per hectare (RadamBrasil, 1974); the percentage of
commercial-sized trees without defects (50–80%; Holmes
et al., 2002; Grogan and Schulze, 2008; Zweede, unpublished
data); and 5.3–7.8 m3 of roundwood volume per harvested tree
(diameter- and height-based volume equations as above;
diameter distributions and tree heights from this study and
Schulze (2003)).
Economic feasibility of Tabebuia extraction at regional
scales – that is, the extent of their natural ranges that can
be profitably logged through conventional selective high-
grading – was inferred from spatial models of the economic
accessibility of logging in the Brazilian Amazon (Stone, 1998;
Verıssimo et al., 1998; Lentini, 2007). These models estimate
the feasibility of timber extraction based on prices of logs pro-
cessed in sawmills subtracting transportation, logging, trans-
action costs for harvesting activities, and a minimum profit
level that loggers will impose over the log price. During mod-
eling procedures, an algorithm developed in a GIS environ-
ment (ArcView 3.2a) estimates that a given forest cell is
feasible for logging if the difference between logwood prices
(at the urban centers where milling occurs) and the costs de-
scribed above is greater than or equal to zero. Logwood prices
for ipe were estimated in 2004 as US$ 42.5 per m3 on average,
ranging from US$ 29 to 73 depending on the region within
Amazonia (standard deviation US$ 28.9). Harvesting costs var-
ied between US$ 9.0 and US$ 13.7 per m3 depending on the re-
gion, and transaction costs were assumed to be fixed and

B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2077
equal to US$ 3.9 per m3 (Barreto, 2002). Minimum profit levels
in logging activity were assumed to be 10%, based on an
extensive survey carried out in Para State in 1998 by Verıssimo
et al. (2002b). Transportation costs between milling centers
and harvestable forests are highly variable in the Amazon,
depending on the type of log transport surface (rivers, roads)
and extant infrastructure in each region. Modeling accounted
for different costs depending on the quality of roads in each
region, including expected costs of the construction of new
roads to access valuable timber (more details are given in Len-
tini (2007)). The location of milling centers harvesting Tab-
ebuia in 2004 and the economic data used in the modeling
were collected in an extensive survey conducted in 143 mill-
ing localities (Lentini et al., 2005).
3. Results
3.1. Geographic range and abundance
Both Tabebuia are widely distributed across the Brazilian Ama-
zon, typically co-existing within a given region or forest. Both
species generally occur at low densities; in the extensive
RadamBrasil (1974) forest inventory, average density of trees
P30 cm diameter in regions where Tabebuia were present
was 0.11 and 0.32 trees ha�1 for T. impetiginosa and T. serratifo-
lia, respectively (Fig. 1). Both species occurred at relatively
high-density compared to the overall species-level density
in RadamBrasil plots in transitional forests of south and
southwestern Brazilian Amazonia (Fig. 1). This pattern is sup-
ported by Bolivian forest inventory data in which these two
species occurred at relatively high densities (mean of 2.5
and 0.45 trees ha�1 for T. impetiginosa and T. serratifolia, respec-
tively; Justiniano et al., 2000). In the large-scale inventories at
sites A–G, Tabebuia species were present at low densities sim-
ilar to those found in the RadamBrasil survey, with T. serrati-
folia occurring at highest density at site E in the western
state of Acre (Table 2).
3.2. Population structure and dynamics
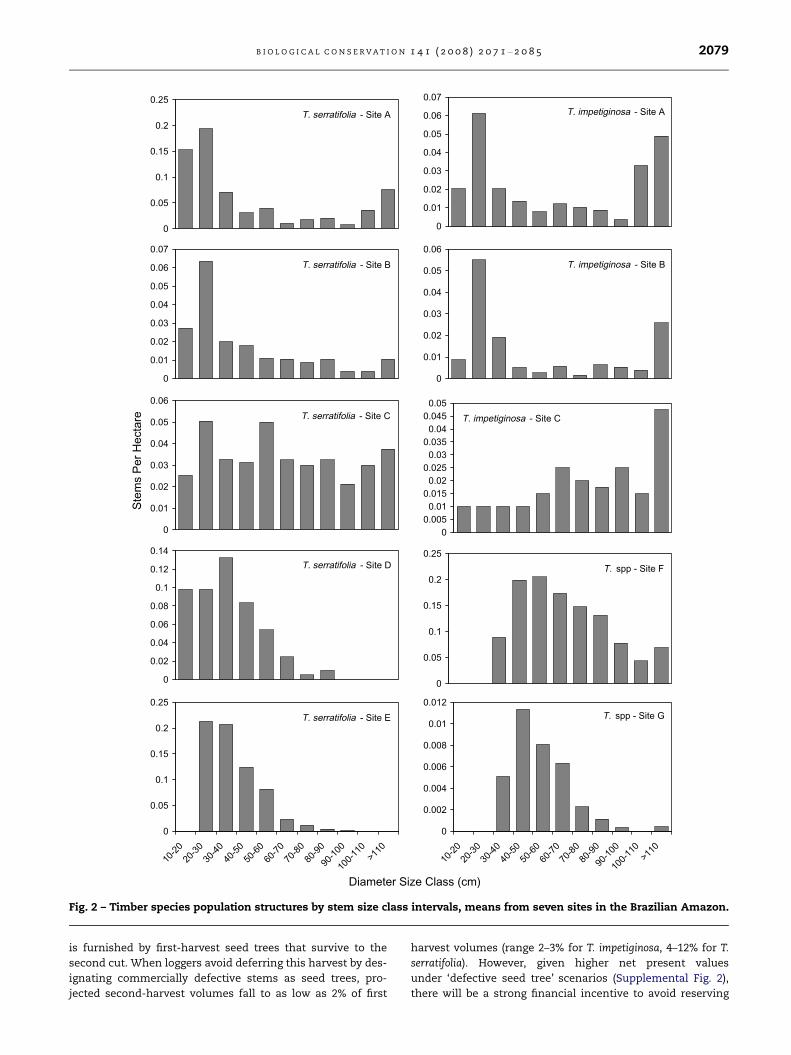
The frequency distribution of stem diameter size classes,
from pole-sized juveniles to large adults, determines timber
yields during first and second harvests. Sub-merchantable
trees plus commercial-sized trees (P50 cm diameter) surviv-
ing the first harvest must comprise the second harvest com-
monly anticipated after 30 yr of growth. Tabebuia population
structures in the northeastern Amazon are typically unbal-
anced towards large diameter size classes, with commer-
cial-sized trees equal to or outnumbering sub-merchantable
trees (P20 cm diameter) in six of 10 populations (Fig. 2). In
transitional forests in southeastern and southwestern
Amazonia (sites D and E), T. serratifolia attains a smaller max-
imum adult size and has a higher ratio of sub-merchantable
to commercial stems.
The rate at which sub-merchantable trees achieve harvest-
able size is determined by growth capacity and environmen-
tal conditions. Mean growth rates for all diameter size
classes of Tabebuia are low in both unlogged and logged for-
ests (Table 4 and Supplemental Fig. 1). Sub-merchantable Tab-
ebuia trees growing at 90th percentile rates in unlogged and
recently logged forests achieve diameter increments of 0.49
and 0.70 cm yr�1, respectively. That is, only sub-merchantable
trees of at least 20 cm diameter can achieve commercial size
during anticipated 30-yr intervals between harvests. Nine out
of ten stems will grow slower than these optimal rates.
While observed mortality rates by Tabebuia are low (<1%
annually) compared to other tropical timber species, post-log-
ging mortality by surviving trees increases to 2.5% due to
stem damage and crown exposure to storm winds (Table 4;
Schulze and Zweede, 2006). If mortality rates decline to nor-
mal within 10 yr of logging, approximately 28% of surviving
trees can be expected to die during anticipated 30-yr intervals
between harvests.
Timber production beyond the second harvest will depend
on seed production by surviving trees and seedling growth
through sub-merchantable to commercial stem size classes.
Advance seedling regeneration is rarely present in the forest
understory at the time of first harvest due to low growth
and high mortality rates in closed forest (Table 4). Height
growth rates by Tabebuia seedlings in logging gaps are also
low (24 ± 15 cm yr�1; ± values indicate SD throughout the
text). Even seedlings growing at 90th percentile rates in-
creased heights by only 1 m yr�1. Five years after logging gaps
were opened in eastern Amazonia, 90% of seedlings remained
in the lower strata of gap vegetation, shaded by faster growing
pioneer species (Schulze, 2003).
3.3. Harvest projections: post-logging recovery
After removing 90% of commercial-sized adults in the first
harvest – currently allowable under Brazilian forest legislation
– commercial volume would not recover to pre-harvest levels
within 60 yr under all model scenarios (Table 5). Even under
the rapid growth projection, at least 60 yr would be required
for an equivalent volume to be extracted in the second har-
vest. If harvest intensity were restricted to only 70% of har-
vestable trees and the minimum felling diameter were
raised to 70 cm, populations would require less time to re-
cover wood volume, although 30 yr (the cutting cycle widely
assumed to be sustainable for Amazonian forests harvested
using RIL) would not be sufficiently long to ensure an equiva-
lent harvest of T. impetiginosa during the second cut (Table 5).
Assumptions about the defect rate in commercial popula-
tions affected total harvestable volume in both the first and
second harvest, but did not substantively alter estimates of
percent volume recovery between first and second harvests
(Supplemental Fig. 2). When all commercial trees were con-
sidered sound under the Brazilian regulation scenario (10%
retention rate), harvestable volume at year 30 was <21% of
first-harvest volume at each of the three sites for both Tab-
ebuia species. A similar lack of substantive differences be-
tween no-defect and defect projections was observed under
all combinations of growth (probable, fast) and harvest (Bra-
zilian legal standard, restricted harvest) scenarios.
In contrast, allowing loggers to meet seed tree retention
requirements with defective trees increased projected first-
harvest volume and value but reduced second-harvest pros-
pects (Supplemental Fig. 3). This result highlights the large
contribution that seed trees retained during the first harvest
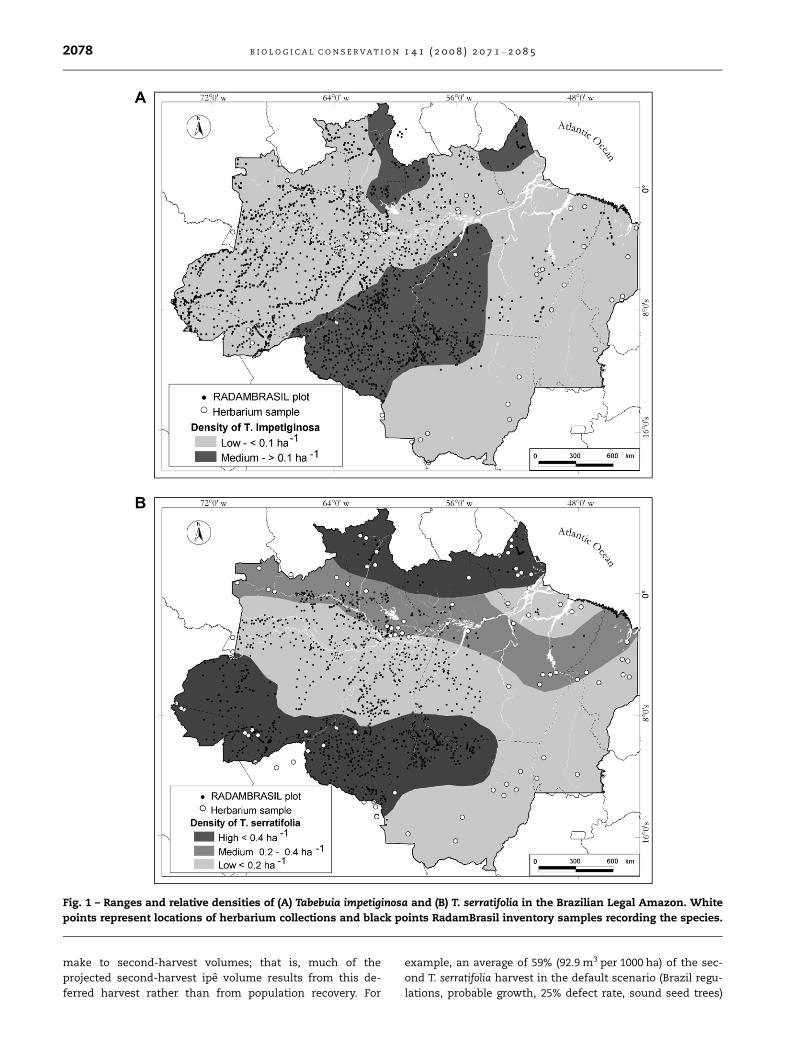
Fig. 1 – Ranges and relative densities of (A) Tabebuia impetiginosa and (B) T. serratifolia in the Brazilian Legal Amazon. White
points represent locations of herbarium collections and black points RadamBrasil inventory samples recording the species.
2078 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
make to second-harvest volumes; that is, much of the
projected second-harvest ipe volume results from this de-
ferred harvest rather than from population recovery. For
example, an average of 59% (92.9 m3 per 1000 ha) of the sec-
ond T. serratifolia harvest in the default scenario (Brazil regu-
lations, probable growth, 25% defect rate, sound seed trees)
T. serratifolia - Site A
0
0.05
0.1
0.15
0.2
0.25T. impetiginosa - Site A
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
T. serratifolia - Site B
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07T. impetiginosa - Site B
0
0.01
0.02
0.03
0.04
0.05
0.06
T. serratifolia - Site C
0
0.01
0.02
0.03
0.04
0.05
0.06
T. impetiginosa - Site C
00.005
0.010.015
0.020.025
0.030.035
0.040.045
0.05
T. serratifolia - Site D
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
T. spp - Site F
0
0.05
0.1
0.15
0.2
0.25
T. serratifolia - Site E
0
0.05
0.1
0.15
0.2
0.25
10-20
20-30
30-40
40-50
50-60
60-70
70-80
80-90
90-10
0
100-1
10>1
10
T. spp - Site G
0
0.002
0.004
0.006
0.008
0.01
0.012
10-20
20-30
30-40
40-50
50-60
60-70
70-80
80-90
90-10
0
100-1
10>1
10
Stem
s Pe
r Hec
tare
Diameter Size Class (cm)
Fig. 2 – Timber species population structures by stem size class intervals, means from seven sites in the Brazilian Amazon.
B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2079
is furnished by first-harvest seed trees that survive to the
second cut. When loggers avoid deferring this harvest by des-
ignating commercially defective stems as seed trees, pro-
jected second-harvest volumes fall to as low as 2% of first
harvest volumes (range 2–3% for T. impetiginosa, 4–12% for T.
serratifolia). However, given higher net present values
under ‘defective seed tree’ scenarios (Supplemental Fig. 2),
there will be a strong financial incentive to avoid reserving

Table 5 – Estimated timber volume available from commercial-sized trees of Tabebuia impetiginosa and T. serratifolia atsecond harvest as a percent of the volume extracted during the first harvest at sites in eastern Amazonia
% First-harvest volume at estimated growth rates
Harvestintensity (%)
Minimum fellingdiameter (cm)
Years after1st harvest
Probable (%) Fast (%)
T. impetiginosa 90a 50 30 12.4–13.6b 12.9–14.3
60 12.6–15.9 16.6–35.9
70c 70 30 43.1–49.9 43.8–51.3
60 40.3–49.3 48.9–62.0
T. serratifolia 90 50 30 15.2–24.1 16.5–26.2
60 18.5–29.8 29.3–63.7
70 70 30 41.9–57.3 43.5–60.2
60 42.8–61.7 58.4–99.0
Based on observed initial population structures, and estimated growth and mortality rates of trees P10 cm diameter and juveniles P50 cm tall.
a Estimated first-harvest volume extracted at 90% rate varied by site from 357 to 832 m3 per 1000 ha for T. impetiginosa and 269 to
1270 m3 per 1000 ha for T. serratifolia.
b Estimates were calculated for each site separately and are presented here as ranges.
c Estimated first-harvest volume extracted at 70% rate varied by site from 263 to 654 m3 per 1000 ha for T. impetiginosa and 174 to
913 m3 per 1000 ha for T. serratifolia.
0
10
20
30
40
50
60
70
80
90
1 10 20 30 40 50 60 70 80 90 100Year after logging gap formation
Num
ber
per
100
hect
ares
Mean density
Lowest observed density
Highest observed density
0
10
20
30
40
50
60
0 10 20 30 40 50 60 70 80 90 100Year after logging gap formation
Dia
met
er a
t bre
ast h
eigh
t (cm
)
Mean growth90th percentile growthMaximum growth
Minimum legal commercial size = 50 cm dbh
X Maximum observed size in 15-year-old logging gaps
A
B
Fig. 3 – Projection of Tabebuia (A) survival and (B) recruitment
in logging gaps based on observed sizes, densities, growth
and mortality in 2–year-old logging gaps in RIL stands at
three sites (A–C) in Para, Brazil. The X in (B) represents the
maximum observed size of a Tabebuia sapling observed in a
survey of 180 15-year-old logging gaps.
2080 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
commercially sound seed trees, to the extent that this is leg-
ally or technically feasible.
3.4. Regeneration and population recovery
Observed Tabebuia regeneration following logging at study
sites in eastern Amazonia was patchy, with low overall seed-
ling and sapling densities in the high-light disturbed zones
within logged forest (5.7 ± 3.8 stems ha�1). Only at the highest
observed seedling densities and the lowest possible mortality
rates would a sufficient number of stems survive post-logging
gap succession to eventually yield densities of at least three
commercial-sized trees per 100 ha (Fig. 3a), the minimum
density necessary to allow harvest under current regulations
(Brazil, 2006). This population-level recovery through seedling
recruitment to commercial size classes would require approx-
imately 100 yr (Fig. 3b).
3.5. Exploitation rates and regional harvest pressure
Based on Brazilian government data tracking timber exports
(SECEX, 2005), we estimate that 1,104,000 m3 of ipe round-
wood were harvested in 2004 to generate 167,000 m3 of ex-
port-grade timber. These estimates are comparable to
mahogany volumes extracted from Brazilian Amazonia dur-
ing the ‘mahogany rush’ of the late 1980s and early 1990s
(Browder, 1987; Verıssimo et al., 1995; Grogan et al., 2002). At
landscape-scale densities recorded by RadamBrasil (1974)
and Schulze (2003), approximately 649,000 ha of forest were
selectively logged in 2004 to meet this export total (assuming
a defect rate of 25%, mean values of 6.6 m3 tree�1 harvested,
and 950 kg m�3; minimum and maximum area estimates de-
rived from extremes of these three values were 373,000 and
1,354,000 ha, respectively). This estimated forest area repre-
sents ca. 40% of the total forest area logged annually in Brazil-
ian Amazonia (Nepstad et al., 1999; Asner et al., 2005).
Sawmill surveys in 2004 revealed that ipe timber was
being extracted from 93 of the 143 milling localities in
Amazonia, while most timber processing centers where
ipe logging was not recorded (41 of 50) were located in

B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2081
old frontier areas of eastern Para and north-central Mato
Grosso (Fig. 4). Of these 50 milling localities, 80% were still
producing ipe timber in 1998, suggesting that by 2004 ipe
was exhausted in these regions. With few exceptions, ipe
timber was an important product in the western logging
frontiers in 2004. Sawmill surveys indicate a wave of ipe
logging moving from the original eastern logging centers
west into more remote regions over time. Where access
and infrastructure are poor and the pool of harvest species
restricted, ipe species dominate selective harvests. In well-
developed logging centers with large species lists, ipe is ab-
sent from or a small component of harvests.
Ipe’s role as a critical resource in new logging frontiers is
predicted by models of logging profitability. Even though Tab-
ebuia occur at extremely low densities, these species can be
extracted profitably from all but the remotest 36% of Brazilian
Amazonia, corresponding to approximately 178 million km2
(Fig. 4). Over most of the Amazon, economic barriers are
now insufficient to prevent pursuit of new sources of high-va-
lue ipe timber to replace dwindling production in old sawmill
centers.
Fig. 4 – Map of the Brazilian Legal Amazon showing urban cent
triangles) ipe timber in 2004. The majority (80%) of the milling ce
eastern and southern borders of Amazonia, were processing ip
extraction of ipe species is profitable based on prices of sawn i
logging centers. Medium gray areas represent regions where tra
unprofitable. Although the ranges of Tabebuia impetiginosa and
species are not present in all forests in the region and understa
4. Discussion
Tabebuia impetiginosa and T. serratifolia populations in the
Amazon present considerable challenges for sustainable
management. Both species generally occur at densities much
lower than shade-tolerant timber species such as Manilkara
huberi (Schulze et al., 2005), and as low as the threatened
mahogany (0.18 trees ha�1 in RadamBrasil (1974)). Moreover,
population structures are weighted towards large, very old
adults, meaning relatively few young trees are present in
stands. This pattern is typical for light-demanding long-lived
timber species in the Amazon, including mahogany (Gullison
et al., 1996; Jennings et al., 2001; Grogan et al., 2002), and is
frequently cited as a major limitation for sustainable timber
production (e.g., Fredericksen and Putz, 2003). In the case of
Tabebuia, unfavorable population distribution patterns are
compounded by slow median and maximum growth rates.
Thus, in many Amazonian forests, ipe presents a ‘perfect
storm’ of unfavorable ecological characteristics from the per-
spective of the forest manager concerned with sustained tim-
ber production.
ers processing (white triangles) and not processing (black
nters not processing ipe in that year, all located closer to the
e timber in 1998. The map also shows areas where the
pe timber and transportation costs between forests and
nsport limitations currently render extraction of ipe timber
T. serratifolia encompass the entire Legal Amazon, these
nding of species’ distribution patterns remains limited.

2082 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
4.1. Can 30-yr cutting cycles be sustained?
One frequently cited benefit of reduced-impact logging (RIL) is
that cutting cycles of 30 yr will be possible when RIL is em-
ployed, compared to 100 yr or more when logging is un-
planned (Uhl et al., 1997). The widely held assumption that
30-yr cutting cycles will be sustainable is based on stand-level
volume increment from a limited number of small plots in
Amazonia (Barreto et al., 1998; Alder and Silva, 2000; Keller
et al., 2004; Valle et al., 2006). These estimates do not consider
logging impacts on timber species populations or population
recovery rates. Given Tabebuia population dynamics at our
eastern Amazonian sites, it is not possible to maintain timber
production at current harvest levels on 30-yr cutting cycles, or
even at substantially reduced logging intensities with the cur-
rent log-and-leave RIL model. Similar population structures
(Fig. 2), slow growth rates (Dauber et al., 2003; Supplemental
Table 3), and poor regeneration at other Amazonian sites
(Fredericksen and Mostacedo, 2000; Justiniano et al., 2000;
Heuberger et al., 2002) indicate that our results are broadly
applicable across Tabebuia ranges in Amazonia (population
dynamics may be quite different outside Amazon forests,
such as in cerrado and pantanal regions). New silvicultural
models will be necessary to sustainably manage Tabebuia
and similar timber species.
4.2. Will populations persist under logging pressure?
In order to predict long-term effects of logging on Tabebuia
populations, we must understand seedling dynamics in dis-
turbed areas (logging gaps and skid trails) within logged for-
ests. Given low densities of sub-merchantable trees,
saplings, and seedlings in unlogged forest, seedling perfor-
mance in logging gaps will largely determine these species’
capacity to recover from logging disturbance. Even under
the best regeneration scenario – the highest observed seed-
ling densities in logging gaps and lowest observed mortality
rates – only one ipe tree per 10 ha could be expected to survive
to adult size, and this only after a century or more of growth.
A more probable scenario is that only the rare individual
establishing in the right location in a gap with appropriate
conditions will survive and recruit to commercial size in
logging gaps that are left to natural successional processes.
We therefore cannot assume that natural regeneration fol-
lowing logging will be sufficient to enable population recov-
ery from the initial harvest. Thus we conclude that
sustainable logging of Tabebuia will require some combina-
tion of enrichment planting and long-term silvicultural
tending of established seedlings in high-light environments
within logged stands.
Demand for ipe timber is responsible for the logging of a
vast region of Amazonia each year, much of this logging is
in new frontier regions, and the expansion of ipe logging into
previously intact forest regions will likely increase as demand
for ipe increases and supplies from established logging cen-
ters dwindle. Ipe harvest volumes in 2004 were comparable
to mahogany volumes extracted from Brazilian Amazonia
during the ‘mahogany rush’ of the late 1980s and early
1990s (Browder, 1987; Verıssimo et al., 1995; Grogan et al.,
2002). Accurate estimates of total annual ipe consumption,
including domestic and clandestine production, are not avail-
able; it is likely, however, that the total area of forest logged
for ipe in 2004 was greater than our estimations from export
data. Because ipe stocks have been largely depleted in old
frontier regions such as eastern Para state (Verıssimo and Bar-
reto, 2004), most current production must come from new
logging frontiers in western Para, southern Amazonas, north-
western Mato Grosso, and Acre.
Similar to mahogany since the early 1970s, Tabebuia log-
ging is lucrative in primary forests at great distances from
established wood processing centers, drawing industrial tim-
ber operations into the core of the Amazon Basin (Verıssimo
et al., 1992, 2002; Lentini et al., 2003, 2005). Planned or ongoing
projects to extend the network of all-weather roads into the
core of Amazonia (Laurance et al., 2001; Nepstad et al., 2004)
would remove most of the remaining economic barriers to
ipe extraction.
A general expansion of export-oriented forestry has been
observed in Brazil (Lentini et al., 2005) and is predicted to con-
tinue as Amazonian timber fills the void left by depletion of
southeast Asian timber stocks (Nepstad et al., 2004). We
emphasize that the vast majority of logging in Amazonia does
not following RIL protocols or even the legal standards used
as the basis for the projections presented here (Schulze
et al., 2008). That is, deleterious impacts on populations har-
vested by conventional or predatory logging practices are
undoubtedly higher than we report here due to higher inten-
sity harvests, absence of seed trees, and greater damage in-
flicted on the residual stand (Johns et al., 1996; Holmes
et al., 2002).
4.3. Ipe logging as a catalyst for forest degradation andclearing
In recent years there has been a clear shift in the geographical
range of ipe logging, as stocks in older eastern frontiers were
exhausted and logging operations spread to central and wes-
tern Amazonia. Even without planned expansion of the trans-
portation network, the majority of the Brazilian Amazon is
already financially accessible to ipe logging. Left to market
forces, ipe logging will continue to spread to unlogged forest
regions wherever these species can be profitably harvested.
As logging roads and sawmills penetrate new regions they
provide access and incentive for colonists and land specula-
tors to follow (Fearnside, 1997; Laurance et al., 2001). This dy-
namic has been well documented for Amazonia (Asner et al.,
2006), and in much of the so-called ‘arc of deforestation’ the
pursuit of big-leaf mahogany served as the catalyst for the
process of land-clearing, serial logging and burning that has
resulted in a landscape where islands of degraded forest
(heavily logged and in many cases repeatedly burned) persist
precariously within a matrix of used and abandoned pastures
and agricultural fields (Holdsworth and Uhl, 1997; Gerwing,
2002). With ipe as a new catalyst, the wave of forest degrada-
tion and clearing powered by largely unregulated logging now
threatens to wash across the heart of Amazonia. The implica-
tions of uncontrolled logging of ipe extend well beyond the
potential depletion of commercial stocks of Tabebuia species,

B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2083
and could undermine government efforts to achieve forest
conservation by bringing order to the Amazon frontier.
4.4. Regulation of ipe logging
Industrial logging is depleting Tabebuia populations through-
out Amazonia. Illegal harvest of high-value species such as
Tabebuia continues to drive expansion of logging frontiers into
regions previously inaccessible to logging. Current best-prac-
tices forestry, even if uniformly adopted, would not alleviate
the pressure on Tabebuia populations because harvest intensi-
ties and silvicultural systems under RIL have not been ad-
justed to species-level population dynamics. Solving these
problems will require eliminating incentives for illegal logging
of ipe while improving standards for legal management of
this renewable resource.
In Brazil, concern about populations of high-value species
has led to specific legal protection for Brazilnut (Bertholettia ex-
celsa), virola (Virola duckei), and more recently, mahogany (Bra-
zil, 1994, 1999, 2003). Brazilnut and virola were protected
through domestic government policies, whereas protection
for mahogany stemmed from concern over international
trade of a threatened species. Mahogany’s recent listing on
CITES Appendix II forced the design and adoption of manage-
ment requirements more stringent than for any other timber
species in Amazonia, while making it more difficult to market
illegal wood (Blundell, 2004). This has slowed the spread of
uncontrolled mahogany logging into new regions and raised
hope that S. macrophylla might still be sustainably managed
within some portion of its natural geographic range (Grogan
and Barreto, 2005).
Like mahogany, the two species of Tabebuia discussed here
merit greater protection than provided by current generic for-
estry laws in Brazil and other Amazonian countries. The eco-
logical similarities between Tabebuia and S. macrophylla are
striking. Tabebuia appear as vulnerable to uncontrolled log-
ging as mahogany. The growing international demand for
and rising value of ipe timber is creating pressure on this for-
est resource that may soon rival the pursuit of ‘green gold’, as
mahogany is widely known. International demand facilitates
much of this uncontrolled logging and is one factor (albeit
indirect) in the expansion of illegal logging into the heart of
Amazonia. Clearly, eradication or drastic reduction of illegal
logging – possible through improved monitoring and enforce-
ment as well as market avoidance of wood of uncertain origin
– is fundamental to conserving forest resources, including
Tabebuia. However, protecting Tabebuia populations will re-
quire development of higher standards for ipe management
– reduced harvest intensity and requirements for manage-
ment of regeneration – much like those for the threatened
mahogany. Tabebuia species may also warrant consideration
for international regulatory protection under the CITES char-
ter. Unsustainable logging of mahogany was not slowed in
Brazil until systematic efforts to restrict harvest, transport
and trade were implemented at the national and interna-
tional levels (Grogan and Barreto, 2005). These efforts came
too late for most mahogany populations and forests within
the species’ range (Grogan et al., 2008). Will we repeat history
with ipe?
Acknowledgements
We especially thank L. Rogerio Oliveira for providing inven-
tory data from Rondonia and Amazonas, and Rodney Salo-
mao for assistance with preparation of the economic
accessibility model in Fig. 4. Funding for various phases of
this research was provided by the US Agency for International
Development, the National Science Foundation (graduate fel-
lowship), the Charles A. and Anne Morrow Lindbergh Founda-
tion, the Conservation, Food and Health Foundation, the
International Tropical Timber Organization, the Lincoln Park
Zoo Scott Neoptropic Fund, and the Milton and Miriam Han-
dler Foundation. Among many contributors in the field, we
thank Elaelson Santos, Josieldo Pantoja, Manoel Rodrigues
Vitorino, Marcelo Galdino, Maria Nascimento Rodrigues, Mig-
uel Alves de Jesus, Miguel Lopes, and Waldemir Ribeiro da
Cruz.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,
in the online version, at doi:10.1016/j.biocon.2008.06.003.
R E F E R E N C E S
Alder, D., Silva, J.N.M., 2000. An empirical cohort model formanagement of terra firme forests in the Brazilian Amazon.Forest Ecology and Management 130, 141–157.
Asner, G.P., Knapp, D.E., Broadbent, E.N., Oliveira, P.J.C., Keller, M.,Silva, J.N., 2005. Selective logging in the Brazilian Amazon.Science 310, 480–482.
Asner, G.P., Broadbent, E.N., Oliveira, P.J.C., Keller, M., Knapp, D.E.,Silva, J.N., 2006. Logging and deforestation trends in theBrazilian Amazon. Proceedings of the National Academy ofSciences 103, 12947–12950.
Barreto, P., Amaral, P., Vidal, E., Uhl, C., 1998. Costs and benefits offorest management for timber production in easternAmazonia. Forest Ecology and Management 108, 9–26.
Barreto, P., 2002. Manejo florestal para producao de madeira naAmazonia: situacao e perspectivas. IMAZON, Belem, PA, Brazil.<http://www.imazon.org.br>.
Blundell, A.G., 2004. A review of the CITES listing of big-leafmahogany. Oryx 38, 1–7.
Boltz, F., Carter, D.R., Holmes, T.P., Pereira Jr., R., 2001. Financialreturns under uncertainty for conventional and reduced-impact logging in permanent production forests of theBrazilian Amazon. Ecological Economics 39, 387–398.
Brazil, 1994. Decreto No. 1282, 19 October 1994. Regulamenta osartigos 15, 19, 20 e 21, da Lei n. 4771 de 15 de setembro de 1965,e da outras providencias. <http://www.lei.adv.br/1282-94.htm>.
Brazil, 1999. Instrucao Normativa No. 01, 8 January 1999.Ministerio do Meio Ambiente, Instituto Brasileiro do MeioAmbiente e dos Recursos Naturais Renovaveis (IBAMA).<http://ibama2.ibama.gov.br>.
Brazil, 2003. Instrucao Normativa No. 07, 22 August 2003. InstitutoBrasileiro do Meio Ambiente e dos Recursos NaturaisRenovaveis (IBAMA), Ministerio do Meio Ambiente, Brasılia,DF, Brazil. <http://ibama2.ibama.gov.br>.
Brazil, 2006. Instrucao Normativa No. 05, 11 December 2006.Ministerio do Meio Ambiente, Instituto Brasileiro do Meio

2084 B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5
Ambiente e dos Recursos Naturais Renovaveis (IBAMA).<http://ibama2.ibama.gov.br>.
Browder, J.O., 1987. Brazil’s export promotion policy (1980–1984):impacts on the Amazon’s industrial wood sector. The Journalof Developing Areas 21, 285–304.
Brown, S., Gillespie, A., Lugo, A., 1989. Biomass estimationmethods for tropical forests with applications to forestinventory data. Forest Science 35, 881–902.
Dauber, E., Fredericksen, T., Pena-Claros, M., Leano, C., Licona,J.C., Contreras, F., 2003. Tasas de Incremento Diametrico,Mortalidad y Reclutamiento con base en las ParcelasPermanentes Instaladas en Diferentes Regiones de Bolivia.BOLFOR, Santa Cruz, Bolivia, p. 50.
de Graaf, N.R., 2000. Reduced impact logging as part of thedomestication of neotropical rainforest. International ForestryReview 2, 40–44.
de Graaf, N.R., Poels, R.L.H., Van Rompaey, R.S.A.R., 1999. Effect ofsilvicultural treatment on growth and mortality of rainforestin Surinam over long periods. Forest Ecology and Management124, 123–135.
ESRI (Environmental Systems Research Institute), 1999. ArcviewGIS 3.2. Redlands, CA, USA.
Fearnside, P.M., 1997. Protection of mahogany: a catalytic speciesin the destruction of rain forests in the American tropics.Environmental Conservation 24, 303–306.
Fredericksen, T.S., Mostacedo, B., 2000. Regeneration of timberspecies following selection logging in a Bolivian tropical dryforest. Forest Ecology and Management 131, 47–55.
Fredericksen, T.S., Putz, F.E., 2003. Silvicultural intensification fortropical forest conservation. Biodiversity and Conservation 12,1445–1453.
FSC (Forestry Stewardship Council), 2007. Information on certifiedarea available on-line at <http://www.fsc.org> (accessed 1March 2007).
Gentry, A.H., 1992. Flora Neotropica: Bignoniaceae II. Monograph25, The New York Botanical Garden, Bronx, NY, USA.
Gerwing, J.J., 2002. Degradation of forests by logging and fire in theeastern Amazon. Forest Ecology and Management 157, 131–141.
Gourlet-Fleury, S., Favrichon, V., Schmitt, L., Petronelli, P., 2004.Consequences of silvicultural treatments on stand dynamicsat Paracou. In: Gourlet-Fleury, S., Guehl, J.-M., Laroussinie, O.(Eds.), Ecology and Management of a Neotropical Rainforest:Lessons Drawn from Paracou, a Long-term ExperimentalResearch Site in French Guiana. Elsevier, Paris, France, pp. 254–280.
Grogan, J., Barreto, P., Verıssimo, A., 2002. Mahogany in theBrazilian Amazon: Ecology and Perspectives on Management.IMAZON, Belem, PA, Brazil. <http://www.imazon.org.br>.
Grogan, J., Barreto, P., 2005. Big-leaf mahogany on CITES AppendixII: big challenge, big opportunity. Conservation Biology 19, 973–976.
Grogan, J., Galvao, J., 2006. Physiographic and floristic gradientsacross topography in transitional seasonally dry evergreenforests of southeast Para, Brazil. Acta Amazonica 36, 483–496.
Grogan, J., Jennings, S., Landis, R.M., Schulze, M., Baima, A.M.V.,Lopes, J. do C.A., Norghauer, J.M., Oliveira, L.R., Pantoja, F.,Pinto, D., Silva, J.N.M., Vidal, E., Zimmerman, B., 2008. Whatloggers leave behind: impacts on big-leaf mahogany (Swieteniamacrophylla) commercial populations and potential for post-logging recovery in the Brazilian Amazon. Forest Ecology andManagement 255, 269–281.
Grogan, J., Schulze, M., 2008. Estimating the number of trees andforest area necessary to supply internationally traded volumesof big-leaf mahogany (Swietenia macrophylla) in Amazonia.Environmental Conservation 35, 26–35.
Gullison, R.E., Panfil, S.N., Strouse, J.J., Hubbell, S.P., 1996. Ecologyand management of mahogany (Swietenia macrophylla King) in
the Chimanes Forest, Beni, Bolivia. Botanical Journal of theLinnaean Society 122, 9–34.
Hasse, R., Hirooka, R.Y., 1998. Structure, composition and smalllitter dynamics of a semi-deciduous forest in Mato Grosso,Brazil. Flora 193, 141–147.
Heinsdijk, D., Bastos, A., 1963. Inventarios florestais na Amazonia.Boletim do Servico Florestal 6, 1–100.
Heuberger, K., Fredericksen, T.S., Toledo, M., Urquieta, W.,Ramirez, F., 2002. Mechanical cleaning and prescribed burningfor recruiting commercial tree regeneration in a Bolivian dryforest. New Forests 24, 183–194.
Holdsworth, A.R., Uhl, C., 1997. Fire in eastern Amazonian loggedrain forest and the potential for fire reduction. EcologicalApplications 7, 713–725.
Holmes, T.P., Blate, G.M., Zweede, J.C., Pereira Jr., R., Barreto, P.,Boltz, F., Bauch, R., 2002. Financial and ecological indicators ofreduced impact logging performance in the eastern Amazon.Forest Ecology and Management 163, 93–110.
IBGE (Instituto Brasileiro de Geografia e Estatıstica), 2003. BaseCartografica Integrada Digital do Brasil ao Milionesimo. Rio deJaneiro, RJ, Brazil. <www.ibge.gov.br> (accessed on 3:11:05).
Jennings, S.B., Brown, N.D., Boshier, D.B., Whitmore, T.C., Lopes,J.C.A., 2001. Ecology provides a pragmatic solution for themaintenance of genetic diversity in sustainably managedtropical forests. Forest Ecology and Management 154, 1–10.
Johns, J.S., Barreto, P., Uhl, C., 1996. Logging damage duringplanned and unplanned logging operations in the easternAmazon. Forest Ecology and Management 89, 59–77.
Justiniano, M.J., Fredericksen, T.S., Nash, D., 2000. Ecologia ySilvicultura de Especies Menos Conocidas: Tabijos o Lapchos.BOLFOR, Santa Cruz, Bolivia.
Keller, M., Asner, G.P., Silva, J.N.M., Palace, M., 2004. Sustainabilityof selective logging of upland forests in the Brazilian Amazon:carbon budgets and remote sensing as tools for evaluation oflogging effects. In: Zarin, D.J., Alavalapati, J.R.R., Putz, F.E.,Schmink, M. (Eds.), Working Forests in the Neotropics:Conservation through Sustainable Management? ColumbiaUniversity Press, New York, NY, USA, pp. 41–63.
Kometter, R.F., Martinez, M., Blundell, A.G., Gullison, R.E.,Steininger, M.K., Rice, R.E., 2004. Impacts of unsustainablemahogany logging in Bolivia and Peru. Ecology and Society 9.<http://www.ecologyandsociety.org/vol9/iss1/art12>.
Laurance, W.F., Cochrane, M.A., Bergen, S., Fearnside, P.M.,Delamonica, P., Barber, C., D’Angelo, S., Fernandes, T., 2001.The future of the Brazilian Amazon. Science 291, 438–439.
Lentini, M., Verıssimo, A., Lima, E., 2003. Fatos Florestais daAmazonia. IMAZON, Belem, PA, Brazil. <http://www.imazon.org.br>.
Lentini, M., Pereira, D., Verıssimo, A., 2005. Fatos Florestais daAmazonia 2005. IMAZON, Belem, PA, Brazil. <http://www.imazon.org.br>.
Lentini, M., 2007. Land Use Allocation under Multiple Objectivesin the Brazilian Amazon: Creating a Tool to Guide Zoning ofPublic Production Forests. MSc. Thesis. University of Florida,Gainesville, FL, USA.
Loureiro, A., Freitas, J., Ramos, K., Freitas, C., 2000. EssenciasMadeireiras da Amazonia: Volume 4. MCT/INPA-CPPF, Manaus,AM, Brazil.
Metafore, 2004. O Mercado de Produtos Madeireiros Tropicais deValor Agregado nos EUA. Global Woods Facts Sheet, Metafore.<http://www.metafore.org>.
MOBOT (Missouri Botanical Garden), 2005. Tropicos <http://mobot.mobot.org/W3T/Search/vast.html> (accessed July2005).
Nepstad, D., Verıssimo, A., Alencar, A., Nobre, C., Lima, E.,Lefebvre, P., Schlesinger, P., Potter, C., Moutinho, P., Mendoza,E., Cochrane, M., 1999. Large-scale impoverishment ofAmazonian forests by logging and fire. Nature 398, 505–508.

B I O L O G I C A L C O N S E R V A T I O N 1 4 1 ( 2 0 0 8 ) 2 0 7 1 – 2 0 8 5 2085
Nepstad, D., Alencar, A., Barros, A.C., Lima, E., Mendoza, E.,Ramos, C.A., Lefebvre, P., 2004. Governing the Amazon timberindustry. In: Zarin, D.J., Alavalapati, J.R.R., Putz, F.E., Schmink,M. (Eds.), Working Forests in the Neotropics: Conservationthrough Sustainable Management? Columbia University Press,New York, NY, USA, pp. 388–414.
NYBG (New York Botanical Garden), 2005. Website: <http://sciweb.nybg.org/science2/hcol/netr/index.asp> (accessed July2005).
Oliveira, L.C., 2005. Efeito da exploracao da madeira e dediferentes intensidades de desbastes sobre a dinamica davegetacao de uma area de 136 ha na Floresta Nacional doTapajos. Ph.D. thesis, Escola Superior de Agricultura ‘Luiz deQueiroz’/Universidade de Sao Paulo, Piracicaba, SP, Brazil.
Oliveira, M.V.N., 2000. Sustainable forest management for smallfarmers in Acre State in the Brazilian Amazon. PhD thesis,University of Aberdeen, Aberdeen, UK.
Phillips, P.D., de Azevedo, C.P., Degen, B., Thompson, I.S., Silva,J.N.M., van Gardingen, P.R., 2004. An individual-based spatiallyexplicit simulation model for strategic forest managementplanning in the eastern Amazon. Ecological Modelling 173,335–354.
RadamBrasil, 1974. Projeto Radam, Programa de IntegracaoNacional, Volumes 1–24, Levantamento de Recursos Naturais.Ministerio das Minas e Energia, Departamento Nacional deProducao Mineral, Rio de Janeiro, RJ, Brazil.
Rodan, B., Newton, A., Verıssimo, A., 1992. Mahoganyconservation: status and policy initiatives. EnvironmentalConservation 19, 331–342.
Rolim, S., Couto, H., Jesus, R., Franca, J., 2006. Volumetric modelsfor Tapirape-Aquirı National Forest (Para, Brazil). ActaAmazonica 36, 107–114.
Schulze, M., 2003. Ecology and behavior of nine timber treespecies in Para, Brazil: links between species life history andforest conservation and management. PhD thesis, ThePennsylvania State University, University Park, PA, USA.
Schulze, M., Vidal, E., Grogan, J., Zweede, J., Zarin, D., 2005.Madeiras nobres em perigo: praticas e leis atuais de manejoflorestal nao garantem a exploracao sustentavel. Ciencia Hoje36, 66–69.
Schulze, M., Zweede, J., 2006. Canopy dynamics in unlogged andlogged forest stands in the eastern Amazon. Forest Ecologyand Management 236, 56–64.
Schulze, M., 2008. Management of tree regeneration in logginggaps as a potential component of reduced impact logging inthe eastern Amazon. Forest Ecology and Management 255,866–879.
Schulze, M., Grogan, J., Vidal, E., 2008. Technical challenges tosustainable forest management in concessions on publiclands in the Brazilian Amazon. Journal of Sustainable Forestry26, 61–75.
SECEX (Secretariat of External Commerce), 2005. BrazilianMinistry of Development, Industry and Commerce (MDIC),External Commerce Information Analysis System via <http://aliceweb.desenvolvimento.gov.br/> (accessed 04.7.2005).
Silva, J., Carvalho, M., 1984. Equacoes de volume para uma florestasecundaria no planalto do Tapajos-Belterra, PA. Boletim doPesquisa Florestal Curitiba 8/9, 1–15.
Silva, J.N.M., Carvalho, J.O.P., Lopes, J.C.A., Almeida, B.F., Costa,D.H.M., Oliveira, L.C., Vanclay, J.K., Skovsgaard, J.P., 1995.Growth and yield of a tropical rain-forest in the BrazilianAmazon 13 years after logging. Forest Ecology andManagement 71, 267–274.
Silva, J.N.M., Carvalho, J.O.P., Lopes, J.C.A., Oliveira, R.P., Oliveira,L.C., 1996. Growth and yield studies in the Tapajos region,Central Brazilian Amazon. Commonwealth Forestry Review75, 325–329.
Snook, L.K., 1996. Catastrophic disturbance, logging and theecology of mahogany (Swietenia macrophylla King): grounds forlisting a major tropical timber species in CITES. BotanicalJournal of the Linnaean Society 122, 35–46.
Souza, M., Maagliano, M., Camargos, J., 1997. Madeiras TropicaisBrasileiras. IBAMA-DITEC, Brasılia, DF, Brazil.
Stone, S.W., 1998. Using a geographic information system forapplied policy analysis: the case of logging in the EasternAmazon. Ecological Economics 27, 43–61.
Uhl, C., Verıssimo, A., Mattos, M., Brandino, Z., Vieira, I.C.G., 1991.Social, economic, and ecological consequences of selectivelogging in an Amazon frontier: the case of Tailandia. ForestEcology and Management 46, 243–273.
Uhl, C., Barreto, P., Verıssimo, A., Vidal, E., Amaral, A., Barros, A.C.,Souza Jr., C., Johns, J., Gerwing, J., 1997. Natural resourcemanagement in the Brazilian Amazon. BioScience 47, 160–168.
Valle, D., Schulze, M., Vidal, E., Grogan, J., Sales, M., 2006.Identifying bias in stand-level growth and yield estimations: acase study in eastern Brazilian Amazonia. Forest Ecology andManagement 236, 127–135.
Verıssimo, A., Barreto, P., Mattos, M., Tarifa, R., Uhl, C., 1992.Logging impacts and prospects for sustainable forestmanagement in an old Amazonian frontier: the case ofParagominas. Forest Ecology and Management 55, 169–199.
Verıssimo, A., Barreto, P., Tarifa, R., Uhl, C., 1995. Extraction of ahigh-value natural resource in Amazonia: the case ofmahogany. Forest Ecology and Management 72, 39–60.
Verıssimo, A., Souza Jr., C., Stone, S., Uhl, C., 1998. Zoning oftimber extraction in the Brazilian Amazon. ConservationBiology 12, 128–136.
Verıssimo, A., Cochrane, M., Souza Jr., C., Salomao, R., 2002a.Priority areas for establishing national forests in the BrazilianAmazon. Conservation and Ecology 6, 4–14.
Verıssimo, A., Lima, E., Lentini, M., 2002b. Polos Madeireiros doEstado do Para. IMAZON, Belem, PA, Brazil. <http://www.imazon.org.br>.
Verıssimo, A., Barreto, P., 2004. National forests in the BrazilianAmazon: opportunities and challenges. In: Zarin, D.J.,Alavalapati, J.R.R., Putz, F.E., Schmink, M. (Eds.), WorkingForests in the Neotropics: Conservation through SustainableManagement? Columbia University Press, New York, NY, USA,pp. 31–40.
Vidal, E., 2004. Dinamica de florestas manejadas e sob exploracaoconvencional na Amazonia Oriental. PhD thesis, Escola deEngenharia de Sao Carlos/Universidade de Sao Paulo, SaoCarlos, SP, Brazil.




![Adventitious shoot induction from adult tissues of the tropical timber tree yellow Ipé primavera (Tabebuia donnell-smithii rose [bignoniaceae])](https://img.pdfslide.net/doc/110x75/63316a228d2c463a58006148/adventitious-shoot-induction-from-adult-tissues-of-the-tropical-timber-tree-yellow.jpg)
























