Embed Size (px)
Citation preview

Evolution of grasping among anthropoids
E. POUYDEBAT,* M. LAURIN,� P. GORCE* & V. BELS�*Handibio, Universite du Sud Toulon-Var, La Garde, France
�Comparative Osteohistology, UMR CNRS 7179, Universite Pierre et Marie Curie (Paris 6), Paris, France
�UMR 7179, MNHN, Paris, France
Introduction
Grasping behaviour is a key activity in primates to obtain
food. The hand is used in numerous activities of manip-
ulation and locomotion and is linked to several func-
tional adaptations (Godinot & Beard, 1993; Begun et al.,
1997; Godinot et al., 1997). In particular, the hand is
involved in prehension, such as gripping of static foods
(fruits, leaves) and dynamic foods such as insects or other
prey (frogs, rodents, small antelopes). Some primates
such as chimpanzees (Pan troglodytes) and capuchins
(Cebus apella) use their hands to manipulate tools, to
crack nuts, for example (Boesch & Boesch, 1990; Fraga-
szy et al., 2004), whereas gorillas (Gorilla gorilla) use their
hands to extract food from holes (Pouydebat et al., 2005).
The evolution of primates (humans included) is linked to
the development of those behaviours allowing organisms
to exploit the resources in their environment. A general
model of grasping in primates proposes an evolution from
a ‘power grip’ towards a ‘precision grip’, supposed to
have taken place in hominids; the precision grip has been
suggested to appear with Australopithecus afarensis (Mar-
zke, 1997) or with Homo (Napier, 1956, 1960). The power
grip is defined as a grasp with the palm, and is probably a
very old behaviour, as it occurs in anurans, crocodilians,
squamates and several therian mammals (Gray, 1997;
Iwaniuk & Whishaw, 2000). On the contrary, the
precision grip, in which an object is held between the
distal surfaces of the thumb and the index finger, is
usually viewed as a derived function, linked to tool use
and human morphological autapomorphies (Napier,
1956; Tuttle, 1965; Schultz, 1969; Susman, 1979, 1989;
Marzke et al., 1992; Clark, 1993). The precision grip has
been considered the most important hand function of all
prehensile movements (Napier, 1980).
Our aim is to reconsider this simple model of grasping
evolution in the light of morphometric data from
numerous species of primates and behavioural consider-
ations such as areas of contact between the fingers and
the food grasped by extant primates. Therefore, the
possible presence of a phylogenetic signal in the behavio-
ural and relevant morphometric characters is investi-
gated and the correlation between morphometric and
behavioural characters is also determined. We also
present models that enable inference of behaviours from
morphological characters, which we use to infer behav-
iours in three extinct primates: Proconsul africanus,
Oreopithecus bambolii and Australopithecus afarensis, three
species considered to have divergent grasping abilities.
Proconsul africanus and Australopithecus afarensis are from
Africa, which is probably the cradle of hominoid diver-
sification (Arnason et al., 2000; Folinsbee & Brooks,
Correspondence: Michel Laurin, Comparative Osteohistology, UMR CNRS
7179, Universite Pierre et Marie Curie (Paris 6), Paris, France.
Tel.: (33) 1 44 27 36 92; e-mail: [email protected]
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
1732 J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
Keywords:
behaviour;
grasping;
hominids;
palaeobiology;
phylogeny;
precision grip;
primates;
variance partitioning with PVR.
Abstract
The prevailing hypothesis about grasping in primates stipulates an evolution
from power towards precision grips in hominids. The evolution of grasping is
far more complex, as shown by analysis of new morphometric and behavio-
ural data. The latter concern the modes of food grasping in 11 species (one
platyrrhine, nine catarrhines and humans). We show that precision grip and
thumb-lateral behaviours are linked to carpus and thumb length, whereas
power grasping is linked to second and third digit length. No phylogenetic
signal was found in the behavioural characters when using squared-change
parsimony and phylogenetic eigenvector regression, but such a signal was
found in morphometric characters. Our findings shed new light on previously
proposed models of the evolution of grasping. Inference models suggest that
Australopithecus, Oreopithecus and Proconsul used a precision grip.
doi: 10.1111/j.1420-9101.2008.01582.x

2007). Oreopithecus bambolii was found in Tuscany, Italy.
It is included here because its prehensile behaviour has
been inferred in the literature (Moya-Sola et al., 1999;
Susman, 2004).
Material and methods
Quantification of areas of contact
SubjectsThe data represented in this study are based on a wide
variety of primates observed in captivity (Appendix S1):
nine capuchins (C. apella), nine macaques (Macaca
fuscata), nine baboons (Papio papio), three gibbons
(Hylobates lar), seven orang-utans (Pongo pygmaeus), three
gorillas (G. gorilla) and 14 chimpanzees (P. troglodytes).
We also have observations from nine children, 2–5 years
of age (Homo sapiens), and nine adults (H. sapiens). Data
for three other species were collected from the literature
(Christel, 1993; Christel et al., 1998): black mangabey
(Cercocebus aterrimus), geladas (Theropithecus gelada) and
bonobos (Pan paniscus). These species represent a wide
array of body size, hand morphological traits and
anthropoid taxa. Indeed, capuchins do not possess an
opposable thumb and none of the studied primate species
except humans has morphological traits usually associ-
ated with precision grip.
Protocol of observationsThe observations of grasping of small and large objects
have been made in various groups of animals belonging
to zoological gardens in France. All individuals observed
in any given species belong to a single group and the
hierarchical position of each specimen was established.
The animals were observed without modification of their
social (within their group) or environmental (e.g. logs,
rocks, ropes) context to maintain: (i) all behavioural
interactions between the members of the group; (ii) all
constraints in relation to the environment; and (iii) all
possibility of opportunistic manipulation (Parker &
Gibson, 1977).
All observations of the animals were made for
7 months (Pouydebat, 2004). The duration of observa-
tion for each specimen was standardized following
the usual methods suggested in comparative ethology
(Lehner, 1996). A preliminary analysis was conducted by
‘ad libitum sampling’ (Altmann, 1974) that permits the
individual recognition of all subjects for each species and
the identification of a wide variety of areas of fingers in
contact with the presented objects. During the study,
each individual was observed according to the method of
‘focal animal sampling’ (Altmann, 1974). We filmed the
animals during two sessions of 2 h each for chimpanzees,
baboons, capuchins and macaques and six sessions of 2 h
each for orang-utans, gorillas and gibbons. Every 15 min,
sequences of grasping which lasted 5 min were analysed
to determine the area of the finger in contact with the
object by using frame-by-frame analysis in the labora-
tory. We obtained a minimum of 90 min of observation
of grasping behaviour for each chimpanzee, baboon,
macaque and capuchin, and 180 min for each orang-
utan, gorilla and gibbon.
Frame-by-frame analysis was performed with a Basler
camera (Basler, Ahrensburg, Germany), recording 250
images per second. Each prehension technique was
characterized by contacts between one or several lateral
or ventral areas of a minimum of two digits or the
complete palm. From this analysis, we determined five
categories of object prehension.
Size and nature of the objectsFor all primates except humans, the objects were small
and scattered on the ground; the objects involved
spherical cereals and fruits. In humans, the objects were
spherical pearls. It was necessary to standardize the
diameter and the volume in order to calibrate these
parameters according to the length of the hand of the
species studied. In this paper, we always presented
spherical objects to the animals and determined their
diameter. The diameter of the objects was calibrated
according to the length of the hand of the species. As we
knew the length of the hand of the smallest studied
species (76.2 ± 5.3 mm for capuchin) and the diameter
of the smallest object (3.0 ± 0.1 mm) grasped by this
species, we deduced the diameter of objects for other
species as follows (D = diameter, L = length, all units
in mm):
D object for species x¼L hand of the species x�3:0=76:2
For example, to calculate the diameter of objects to be
grasped by chimpanzees, we used the length of the
chimpanzee’s hand (235.0 mm) and that of the smallest
hand (the capuchins’ hand: 76.2 mm) and the diameter
of the smallest object (3.0 mm). In this example, Dxc
corresponds to the determined diameter of the small
object for chimpanzees (c). We calculated the following
value for objects in chimpanzees: Dxc is equal to 9.0 mm
(235.0 mm · 3.0 ⁄ 76.2). We followed the same method
to calculate the diameter of objects for each species
(Appendix S2).
Number of graspsA total of 5549 grasps were recorded for the eight studied
species (Table 1). The percentage of each prehension
category was calculated on the basis of the total number
of grasping observed in each species.
Morphometric dataMorphometric data were obtained from hand skeletons
belonging to the collection of the Museum National
d’Histoire Naturelle (Paris). Our sample consisted of 17
measurements of the hand of 26 taxa (Appendix S3). A
mean of 10 specimens per taxon (males and females) was
measured.
Evolution of grasping among anthropoids 1733
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

Categories of contacts
Each of the six species of primates uses from 5 to 21 of 26
different modes of contacts between areas of digits and
the objects. This large number of contacts can be
classified into five main categories of grasping behaviour
(Table 1). In order to facilitate our comparison with the
previous literature, each category is named (Fig. 1) as
suggested by Napier (1956) and Jones-Engel & Bard
(1996):
Category 1: contact between the distal phalanx of the
thumb, the distal part of the index finger and
the object (precision).
Category 2: contact between the distal phalanx of at
least three fingers and the object (thumb-
distals).
Category 3: contact between the distal part of the thumb,
the lateral side of the middle and proximal
phalanges of the index finger and the object
(thumb-lateral).
Table 1 Use of the grasps from all categories in anthropoid species.
Species N Mean
Category 1
Precision
Category 2
Thumb-distals
Category 3
Thumb-lateral
Category 4
Without thumb
Category 5
Power
Homo sapiens (adults) 600 50.0 ± 0.0 81 ± 3.5 19 ± 2.2 0 ± 0 0 ± 0 0 ± 0
Homo sapiens (child) 450 50.0 ± 0.0 59 ± 4.3 34 ± 1.4 0 ± 0 0 ± 0 7 ± 1.5
Pan troglodytes 804 57.4 ± 4.9 32 ± 2.6 7 ± 0.7 38 ± 2.2 22 ± 3.1 1 ± 0.5
Pan paniscus * * 15 ± 1.7 1 ± 0.2 45 ± 4.0 39 ± 3.2 0 ± 0
Gorilla gorilla 600 200.0 ± 6.7 67 ± 4.4 2 ± 1.1 17 ± 2.0 12 ± 1.9 2 ± 0.3
Pongo pygmaeus 624 89.1 ± 4.7 29 ± 1.3 3 ± 1.3 33 ± 3.1 34 ± 2.0 1 ± 0.4
Hylobates lar 546 182.0 ± 5.3 5 ± 1.2 0 ± 0 95 ± 2.1 0 ± 0 0 ± 0
Papio papio 581 64.6 ± 5.9 71 ± 5.2 3 ± 2.0 24 ± 2.8 0 ± 0 2 ± 0.6
Macaca fuscata 667 74.1 ± 6.2 76 ± 3.9 1 ± 0.3 22 ± 2.4 0 ± 0 1 ± 0.4
Theropithecus gelada * * 77 ± 3.3 2 ± 1.2 18 ± 1.1 0 ± 0 3 ± 1.2
Cercocebus aterrimus * * 70 ± 4.6 0 ± 0 28 ± 1.7 0 ± 0 2 ± 0.6
Cebus apella 677 75.2 ± 7.0 80 ± 4.1 13 ± 2.1 2 ± 0.4 4 ± 1.6 1 ± 0.3
N, Number of grasps of small objects.
The mean number of observed grasps per individual for each species is also given. All other columns represent percentages.
*Data derived from the literature (Christel, 1993; Christel et al., 1998).
(e)
(a)
(c) (d)
(b)
Fig. 1 Definition of contacts (drawings modified from Schultz, 1969). (a) Precision (two digits). (b) Thumb-distal (3, 4 or 5 digits).
(c) Thumb-lateral. (d) Without thumb. (e) Power. The arrows indicate the possible contact between the thumb and the other digits (a–c), and
between the index and middle fingers (d). In the last prehension mode (e), the palm completely covers the object and the ventral surface of the
digits is also implicated in the grasping action. These surfaces are presented in black and the arrows indicate possible modes of contact
between the object and the digit surfaces. In the first example (a), the pad of the thumb and the pad or the extremity of the index can be
simultaneously in contact with the object (photographs: E. Pouydebat).
1734 E. POUYDEBAT ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y
Category 4: contact between one or several fingers,
except the thumb, and the object (without
thumb).
Category 5: contact between the palm, one or several
fingers and the object (power).
Data distribution and transformation
The length of the manus (LM) is used in the analyses
below to remove some of the body size effect on the
remaining characters. For all analyses, this character
(LM) was log-transformed because body size usually
follows a log-normal, rather than normal, distribution.
All other morphometric characters were divided by LM
before all the analyses below (Appendix S3).
Detection of phylogenetic signal
To determine whether or not the phylogeny needed to be
incorporated into the analyses, and whether or not
squared-change parsimony could be used to study char-
acter evolution, we performed two types of tests of
phylogenetic signal. The first one consists of comparing
the squared length of a character over the reference tree
to the squared length of multiple (in this case 10 000)
random trees. For quantitative characters optimized
through squared-change parsimony, branch length data
are critical. Thus, the most appropriate way to create
random trees is to reshuffle the terminal taxa on a tree of
fixed topology and branch lengths (Laurin, 2004; Laurin
et al., 2004). However, squared-change parsimony opti-
mization requires the same assumptions as independent
contrast analysis; thus, we checked if these assumptions
were met using the PDAP (Phenotypic Diversity Analysis
Programs) module for MESQUITEESQUITE (Midford et al., 2003).
This module performs four relevant tests. The first three
regress the absolute value of standardized contrast
against: (i) their expected standard deviation (the square
root of the sum of corrected branch lengths); (ii) the
estimated value of the base node; and (iii) the corrected
height of the base node. The fourth and last test is a
regression of the estimated value of the base node against
the corrected height of the base node. We performed all
these tests (four) for all characters (21) for all trees (five).
No corrections for multiple tests were made, which
makes our procedure more stringent by rejecting trees
which yield artefactual relationships which are signifi-
cant when taken in isolation, but which might no longer
be significant if such corrections were made. Further-
more, it is not clear if such corrections should be made
because these four tests evaluate different statistical
artefacts, and results for one character have no bearing
on other characters. Deviations from the assumptions
were detected in several cases; this prevented analyses of
the relevant characters on a given tree. To maximize the
number of tests that could be performed and to test the
presence of a phylogenetic signal, we produced four
alternative trees (trees 2–5 in Appendix S4) that derive
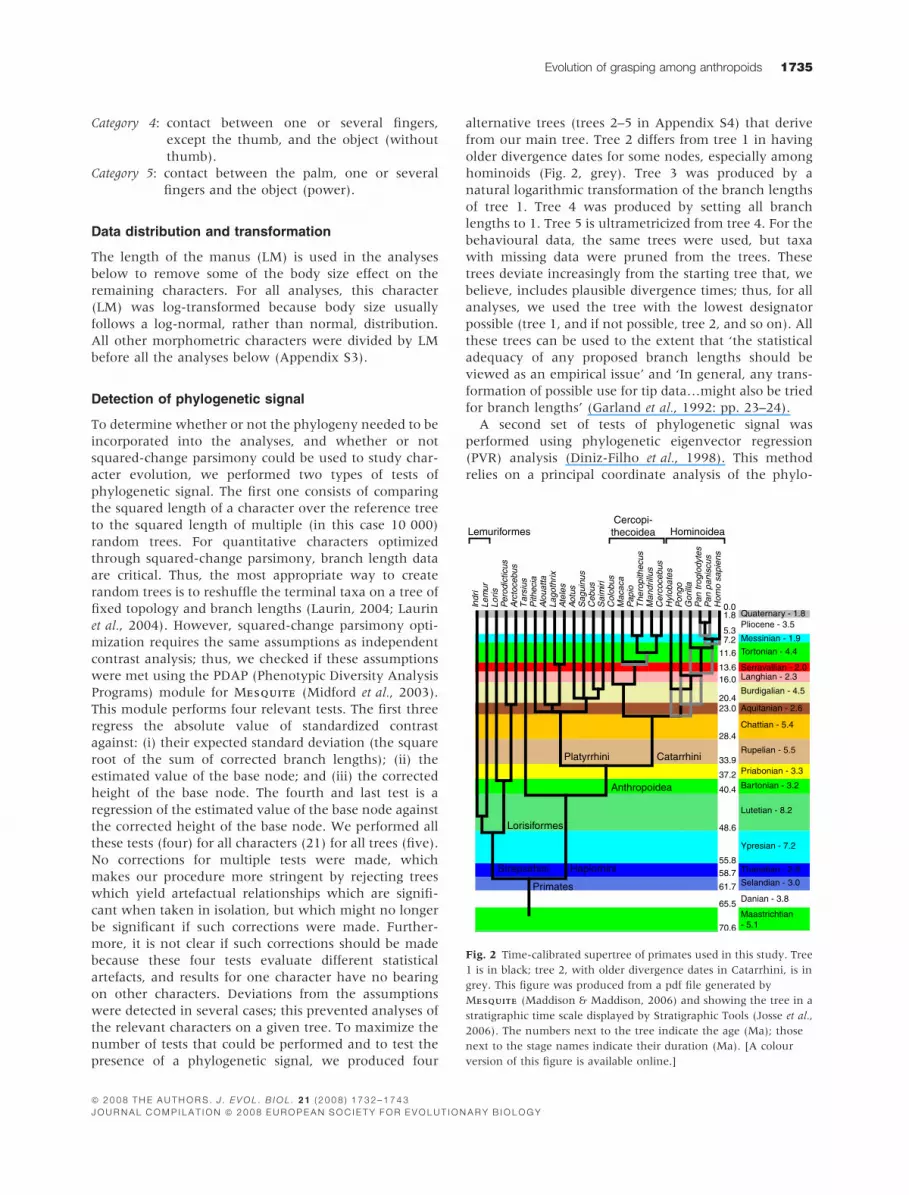
from our main tree. Tree 2 differs from tree 1 in having
older divergence dates for some nodes, especially among
hominoids (Fig. 2, grey). Tree 3 was produced by a
natural logarithmic transformation of the branch lengths
of tree 1. Tree 4 was produced by setting all branch
lengths to 1. Tree 5 is ultrametricized from tree 4. For the
behavioural data, the same trees were used, but taxa
with missing data were pruned from the trees. These
trees deviate increasingly from the starting tree that, we
believe, includes plausible divergence times; thus, for all
analyses, we used the tree with the lowest designator
possible (tree 1, and if not possible, tree 2, and so on). All
these trees can be used to the extent that ‘the statistical
adequacy of any proposed branch lengths should be
viewed as an empirical issue’ and ‘In general, any trans-
formation of possible use for tip data…might also be tried
for branch lengths’ (Garland et al., 1992: pp. 23–24).
A second set of tests of phylogenetic signal was
performed using phylogenetic eigenvector regression
(PVR) analysis (Diniz-Filho et al., 1998). This method
relies on a principal coordinate analysis of the phylo-
Fig. 2 Time-calibrated supertree of primates used in this study. Tree
1 is in black; tree 2, with older divergence dates in Catarrhini, is in
grey. This figure was produced from a pdf file generated by
MESQUITEESQUITE (Maddison & Maddison, 2006) and showing the tree in a
stratigraphic time scale displayed by Stratigraphic Tools (Josse et al.,
2006). The numbers next to the tree indicate the age (Ma); those
next to the stage names indicate their duration (Ma). [A colour
version of this figure is available online.]
Evolution of grasping among anthropoids 1735
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

genetic distance matrix to extract eigenvectors that are
used in a standard linear regression against the character
of interest. The eigenvectors represent the position of the
taxa on the various principal coordinate axes. However,
for n taxa, n – 1 principal coordinate axes are produced,
and they cannot all be used in a linear regression analysis
or there would be no degrees of freedom left. The axes
we used were selected using a broken-stick model
(Diniz-Filho et al., 1998), because it would have taken
too long to test for a significant relationship between all
axes and all characters separately (21 characters and up
to 26 taxa yield 546 tests). Furthermore, none of the
behavioural characters exhibits a phylogenetic signal
according to the squared-change parsimony analysis (see
below), and that method is usually more powerful than
PVR (Cubo et al., 2005). Thus, the broken-stick model
was the only method applicable to all of our data. In all
our analyses of phylogenetic signal, only the first two
axes were used. They represent 60.7% of the phylo-
genetic variance.
The phylogenetic distance matrix was obtained from
tree 1 using the Stratigraphic Tools module (Josse et al.,
2006). The principal coordinate analysis was performed
in PROGICIELROGICIEL R (Casgrain et al., 2004). Linear regressions
were tested for statistical significance using 9999 per-
mutations of the dependent variables (here, the mor-
phometric or behavioural data) in PERMUTEERMUTE! (Casgrain,
2005). A regression is significant (at a 0.05 threshold
value) if fewer than 5% of the data sets have an R2
value at least as large as the original data set (the
original, unpermuted set is included). The advantage of
using permutations to test the significance of the
relationship is that this method requires far fewer
assumptions about the distribution of the data. Thus,
contrary to the other method, this test could be applied
to all characters.
Phylogenetically independent contrasts
We assessed correlations between the behavioural
(dependent) and morphometric (independent) charac-
ters using phylogenetically independent contrasts
(Felsenstein, 1985), whenever the assumptions of
that method were met on at least one of our trees. These
tests were performed using the PDAP module for
MESQUITEESQUITE (Midford et al., 2003). As for the test a
phylogenetic signal using squared-change parsimony,
we performed this test on the tree with the lowest
designator (tree 1 if possible; if not, tree 2, etc.) that gave
adequate contrast standardization for both characters
analysed.
Variance partitioning with phylogenetic eigenvectorregression
Correlations between the behavioural (dependent) and
morphometric (independent) characters were also tested
using variance partitioning with a PVR analysis (Diniz-
Filho et al., 1998; Desdevises et al., 2003). This method
incorporates the phylogeny into the analysis in the form
of principal coordinate axes, as explained in the section
on phylogenetic signal detection (above). We performed
some exploratory analyses (results not shown) in
PERMUTEERMUTE (Casgrain, 2005) to choose the characters to
analyse through PVR. Correlation between these char-
acters was then tested through variance partitioning with
PVR, using principal coordinate axes that were signi-
ficantly correlated with the dependent (behavioural)
characters. These axes were selected by performing a
simple regression of all axes against the relevant
behavioural characters, because in this case only three
characters are involved.
Linear regression models
Simple or multiple linear regressions were used to
produce inference models of the behavioural characters.
These models are based on the morphometric characters
that are significantly correlated with the behavioural
characters according to the variance partitioning analysis
with PVR (Appendix S5) or according to linear regres-
sions, but they were constructed without incorporation
of the phylogeny. This is unavoidable because principal
coordinate axes have no absolute meaning (they differ
when a taxon is added or removed, or if the topology or
branch length is changed), so incorporating them into
predictive models would preclude their use in palaeobi-
ological inference, which is self-defeating. These models
could not be used on some of the extinct taxa included in
our study because the relevant morphometric characters
are not known. To maximize the number of inferences
which could be drawn about these taxa, we built
additional inference models for several combinations of
these taxa and available osteological characters (Appen-
dix S6). For this purpose, we used a forward selection
procedure in PERMUTEERMUTE to select the characters, among
those that were significantly correlated with each other
according to the variance partitioning or the indepen-
dent contrast analyses. In two cases, to produce the
models, we had to extend character selection to other
characters (because extinct taxa are incompletely
known); in such cases, we used a forward selection
procedure in PERMUTEERMUTE (with p to enter of 0.1) to build
the model.
Results
Taxon-specific distribution of categories
The platyrrhine and all catarrhines were able to modulate
their grasping behaviour for small food items. However,
some clusters of species emerge for all categories of
contact. These groups can be compared on the basis of
the mean percentage (Table 1).
1736 E. POUYDEBAT ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

From these data, five main conclusions can be drawn.
First, great apes (except humans) and capuchins use
grasping category 4 (without thumb), whereas humans
and other primates never do. All primates except
humans use the lateral part of their index (category 3,
thumb-lateral), whereas capuchins hardly ever do and
humans never do. Secondly, adult humans and capu-
chins are similar as regards a majority of grasping
categories. Thirdly, human children do not show clear
similarities with other primate species for any of the
grasping categories. Fourthly, cercopithecids (i.e. maca-
ques and baboons), capuchins and humans use the
precision grip most often (Table 1). Fifthly, humans
present some unique characteristics in their selection of
grasping categories. Human adults are the only primates
to use exclusively the tips of the fingers (precision and
thumb distals). Human children and capuchins are more
similar behaviourally to human adults than to other
primates in our sample, because they mainly use the tips
of their fingers (93%). One key result from our data is
that all species were able to grasp small objects with the
precision grip corresponding to the contact between the
tips of thumb and index finger. This applies even to
capuchins, although their thumb is only pseudo-oppos-
able.
Detection of phylogenetic signal
Globally, about 40% of the characters display a phylo-
genetic signal (Appendix S4). This proportion holds
when using both tests, but is affected by taxonomic
sampling; when only the 11 taxa for which behavioural
data are available are tested, this proportion decreases to
20%. This may explain why no phylogenetic signal was
found in the behavioural data using random taxon
reshuffling and squared-change parsimony or PVR
analysis using the first two principal coordinate axes
selected by the broken-stick model (Appendix S4).
However, when each axis was regressed separately
against the behavioural characters, axis 1 had a signif-
icant effect for precision, without thumb and power
grasp frequency.
Character correlation assessed using phylogeneticindependent contrasts
Only three of the behavioural characters were analysed
using independent contrasts because for the others, the
assumptions of that method were not met (Appendix S7).
Some of the morphometric characters are clearly corre-
lated with behavioural data. The behavioural character
‘precision’, involving the contact of the distal phalanges
of the thumb and the index, and the character ‘thumb-
lateral’, involving contact of the distal phalanx of the
thumb and the lateral side of the index, are correlated
with the length of the carpus and the first ray (the thumb
and its metacarpal). Power grasping is linked with second
and third digit length.
Character correlation assessed using variancepartitioning with PVR
The forward selection test in PERMUTEERMUTE (with a p to enter
of 0.1) resulted in only one or two morphometric
characters being selected for each behavioural character
(results not shown), which implies that grasping behav-
iour can be inferred fairly precisely using few morpho-
metric data. No principal coordinate axis was selected. An
additional (non-phylogenetic) test using simple linear
regressions confirms that all the selected characters are
correlated with behavioural characters and explain more
than 50% of the variance in the behavioural characters
(results not shown). In addition, variance partitioning
analyses of the characters which were correlated with at
least one of the phylogenetic principal coordinate axes
indicate that most of the explained variance is genuinely
explained by morphometry rather than by covariation
with the phylogeny (Table 2).
Linear regression models of inference
Inference models obtained through linear regressions
include one or two morphometric characters (Appendix
S5). Inferences were obtained from these models for
extant species in which morphometric data (but no
Table 2 Variance partitioning with PVR showing the correlation between behavioural (dependent) and the morphometric (independent)
characters that were selected by the forward selection procedure using linear regressions with permutations (Appendix 5) and the phylogenetic
principal coordinate axes (we included those that were correlated with behavioural characters based on several simple regressions with
permutations).
Dependent
character
Morphometric characters Phylogenetic axes
Identity
Portion of variance
explained (probability)
Variance explained by covariation
of morphometry and phylogeny Identity
Portion of variance
explained (probability)
Unexplained
variance
Precision 2, 4 0.4364 (0.103) 0.3125 1 0.0528 (0.509) 0.1984
Without thumb 5 0.3544 (0.017) 0.3682 1 0.0268 (0.307) 0.2506
Power 15 0.3404 (0.061) 0.3132 1 0.0540 (0.488) 0.2923
Evolution of grasping among anthropoids 1737
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

behavioural data) are available (Appendix S8), and for
three extinct species (Table 3).
Discussion
Compilation of a time-calibrated tree
For many of the analyses performed below, a phylogeny
incorporating an estimate of branch lengths is needed.
Thus, we compiled a time-calibrated tree (branch lengths
reflect estimated evolutionary time). The topology fol-
lows Goodman et al. (2005). Divergence time estimates
are much more contentious because the affinities of
several primate species based on fragmentary material
are poorly constrained (Ross et al., 1998), and because
divergence time estimates based on molecular data are
often considerably older than the minimal times of
divergence based on fossils (see Marjanovic & Laurin,
2007; for a review). Estimates of divergence times based
on molecular data also often differ substantially between
studies; for instance, Arnason et al. (2000) estimated that
the divergence between strepsirhines and haplorhines at
about 90 Ma (in the Turonian, in the Early Upper
Cretaceous) and that the anthropoid radiation started at
70 Ma, whereas Yoder & Yang (2004) estimated these
events to have occurred at about 80 and 50 Ma, respec-
tively. Yoder & Yang (2004) furthermore estimate the
divergence between Lorisiformes and Lemuriformes at
70–75 Ma, but Roos et al. (2004) estimate it at about
60 Ma, although the oldest fossil in that clade dates from
the Priabonian (Stucky & McKenna, 1993: p. 757), no
more than 37.2 Ma, according to the geological time
scale of Gradstein et al. (2004). Given the poor fossil
record of (crown group) strepsirhines (Stucky &
McKenna, 1993: p. 757), the paleontological date here
is likely to seriously underestimate the actual divergence
dates within Strepsirhini. Considerable differences be-
tween molecular dates are not unexpected because
several methods can be used to obtain molecular dates.
The choice of the calibration date also influences the
results (Brochu, 2004a, b; Poux & Douzery, 2004;
Marjanovic & Laurin, 2007), and there are many other
pitfalls in such analyses (Shaul & Graur, 2002; Graur &
Martin, 2004; Marjanovic & Laurin, 2007). Furthermore,
evolutionary rates are quite variable in primates (Gross-
man et al., 2004; Steiper et al., 2004), which makes
molecular dating more difficult.
For all clades, we have adopted a compromise that uses
minimal divergence ages from the fossil record in the
taxa where this record is reasonably abundant, and
molecular ages for taxa such as Strepsirhini, for which
the fossil record is poor. When using molecular data, we
have tried to use studies that obtained ages compatible
with the fossil record. We assembled the tree in
MESQUITEESQUITE (Maddison & Maddison, 2006) using the
Stratigraphic Tools module (Josse et al., 2006). To facil-
itate comparisons with palaeontological literature, when-
ever molecular dates fall close to geological stage
boundaries, we used the age of the boundary itself
(Fig. 2); this also facilitates tree manipulation in Strati-
graphic Tools (Josse et al., 2006). More information about
individual divergence ages can be found in Appendix S9.
Ontogenetic and taxonomic distribution ofgrasping behaviours
The data presented above show that precision grasping
can be used by all arboreal and terrestrial primate species
in our study. Indeed, the precision grip was recently
reported in capuchins (Spinozzi et al., 2004). However,
according to that report, this grasping technique was less
frequently used by immature individuals. In our study,
all individuals were observed in their social groups, and
we did not observe any obvious effect of ontogenetic age
for this behaviour (data not shown).
Comparisons between great apes and human children
refute the idea that they have similar sensorimotor
organization (Parker & Gibson, 1977). In our study, this
similarity is not great. Categories of grasping used for
small objects differ strongly between human children and
great apes. Great apes use precision grips and the distal
phalanges of their digits less often than children. In
addition, they use the lateral side of their index and the
grip without thumb, contrary to children. These differ-
ences between great apes and children may be explained
by the neural and morphological variability existing
between humans and the other species, regardless of
their age. Finally, the comparison between great apes and
human adults does not show strong similarities. Human
adults use precision grips with small objects and the distal
phalanges of their digits to grasp large objects much more
often.
Relationship between morphometry and graspingbehaviour
The results of this study refute some well-established
ideas about the relationship between morphometry and
Table 3 Inferred behaviour frequency (%) of extinct primate taxa
based on linear regression inference models (Appendix 5).
Taxon Precision
Thumb-
distals
Thumb-
lateral
Without
thumb Power
Australopithecus 58 2 35 )9 (0) 1
Oreopithecus 46 26 18 5 2
Proconsul 76 7 28 13 1
As frequency of a behaviour cannot exceed the interval 0–100%,
values outside this interval represent modelling errors and should be
interpreted as implying values close to the nearest bound (given in
parentheses). Note that as these are inferences, the sums of
percentages on a given line do not necessarily add up to 100% (the
difference between the total and 100% represents modelling errors).
1738 E. POUYDEBAT ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

prehensile behaviour. Our data show that great apes
use precision grips less often than cercopithecoids when
handling small objects and that the contrary pattern is
observed when handling large objects (Pouydebat et al.,
2006a). Furthermore, Pongo and Pan use the grip
without the thumb, contrary to cercopithecoids. These
differences are partly explained by morphometrical data
such as the shorter thumb of great apes (Marzke et al.,
1992), as shown by our analyses (Appendix S7,
precision and characters 3 and 5; Table 2, without
thumb and character 5). Therefore, the length of the
thumb, including the first metacarpal, is an important
morphometrical character in the behaviour of precision
grasp, as previously suggested (Napier, 1956; Schultz,
1969; Susman, 1989). These morphometrical parame-
ters are also negatively correlated with the behavioural
character ‘thumb-lateral’ (Appendix S7), involving the
contact of the distal phalanx of the thumb and the
lateral side of the index. Therefore, the length of the
first ray does not reflect precision grasping only.
Other morphometrical data are specifically correlated
with a single behavioural character. For instance,
the power grasping behaviour, involving the palm of
the hand during the grasp, is strongly correlated with the
lengths of the digits one to three (Appendix S7, power vs.
characters 7–9, 12, 15). The length of digits 2 and 3 is
correlated only with power-grasping (among the behav-
iours studied here); short index and third digits seem to
favour power-grasping.
A few species show prehensile patterns which could
not have been inferred from their hand morphology. For
instance, gorillas show a high percentage of use of the
precision grip in spite of their short thumb (Pouydebat
et al., 2006a). This reflects the fact that the length of the
thumb does not explain all the variance in use of the
precision grip. A quantitative, statistical approach is
required because of the complexity of the relationship
between morphometry and prehensile behaviour.
Finally, we wonder why carpus length is correlated
with precision and thumb-lateral grips. It would be
interesting to test if this can be explained by soft
anatomy, such as muscle or tendon morphology, or by
locomotory behaviour (such as arboreality).
Unexpected similarities between capuchin andhuman prehensile behaviours
Our study reveals similarities in prehensile behaviours
between capuchins and humans. Capuchins use preci-
sion and thumb-distal grips as often as human adults.
Similar to humans, capuchin monkeys almost exclusively
used the distal phalanges of their digits to grasp small
objects. These results can be compared with those of
Spinozzi et al. (2004), who observed a wide variety of
grasping patterns in capuchins. These include various
forms of precision and power grips. Contrary to our
results, Spinozzi et al. (2004) reported that capuchins use
precision and power grips with the same frequency to
grasp small food items. We found that they opted more
often for precision grips (almost 80%). This difference
between our results could be due to the population and
individual variability or the protocol of observation
which was not the same. Capuchins display a wide
variety of prehensile abilities that confirm their capacity,
apparently atypical among New World monkeys, to use
their hands dexterously during extractive foraging and
object manipulation (Fragaszy et al., 1991; Fragaszy &
Boinski, 1995; Christel & Fragaszy, 2000; Pouydebat
et al., 2006b), although they do not possess the true
opposable thumb typical of catarrhine primates.
Inferences about the grasping behaviour of extinctprimates
The inference of grasping behaviour from morphological
analyses of the hands of fossils is a complex problem.
Some authors opposed the hand of extant apes to the
hand of humans and argued that extant apes are unable
to grasp objects with a precision grip or pad-to-pad
gripping. However, many extant ape species use preci-
sion gripping (Christel, 1993; Pouydebat, 2004) without
meeting all the morphological criteria usually considered
to be linked with precision gripping. We have shown that
precision gripping can be performed by hands showing a
greater morphological diversity than previously thought.
Our sampling of behavioural characters in anthropoids
largely restricts our discussion of their possible evolution
within this clade, as suggested by the extant phylogenetic
bracket principle (Witmer, 1995), which was extended
into the context of continuous characters by Laurin et al.
(2004). The absence of a phylogenetic signal in the
behavioural characters (Appendix S4, characters 17–21)
precludes tracing their history over the tree using
optimization procedures because the results would not
be reliable (Laurin, 2004). This limitation may reflect the
relatively low number of taxa for which behavioural data
are available as analyses of our data with the same
taxonomic sampling show a similar absence of a phylo-
genetic signal in most morphometric characters (Appen-
dix S4, central columns). Thus, it might be possible to
reconstruct the evolution of these behavioural characters
by sampling the same clade more densely.
Another possibility is to use the linear regression
inference models that we have produced (Appendix S5)
to infer the behaviour of extinct primate species (Table 3):
A. afarensis, O. bambolii and P. africanus (Fig. 3). Thus, we
can infer that the Plio-Pleistocene hominin A. afarensis
(3–4 Ma old) from Hadar (Ethiopia) exhibited frequent
precision behaviour (Table 3). The linear regression
inference for thumb-distals behaviour in A. afarensis
suggests the infrequent occurrence of this behaviour.
Results about precision behaviour are close to those
obtained by Marzke (1997) and Panger et al. (2002); on
the contrary, several authors suggested that A. afarensis
Evolution of grasping among anthropoids 1739
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

use power grasping most often (Bush et al., 1982; Stern &
Susman, 1983; Susman, 1991, 1994), which we infer to
have occurred infrequently. Australopithecus may share
with humans the absence of without-thumb behaviour,
which may be a synapomorphy of Australopithecus and
Homo among hominoids (although cercopithecoids show
a convergent similarity).
The Miocene ape Oreopithecus may have exhibited
slightly less precision behaviour than Australopithecus, but
like the latter, it resorted to that behaviour much more
often than the power grip, which was rarely displayed.
This result is congruent with the assessment of Moya-
Sola et al. (1999). On the other hand, Susman (2004)
suggested that O. bambolii emphasized the power grip
over the precision grip. Similar to most extant hominoids
and Proconsul, but unlike Homo and Australopithecus,
Oreopithecus may have displayed the without-thumb
behaviour (Table 3).
The linear regression inference models suggest that the
Early Miocene (Burdigalian) stem-hominoid P. africanus
(16–18 Ma) from Kenya used the precision grip fre-
quently (Table 3) and thumb-distals grip more rarely.
P. africanus resorted to thumb-lateral grip relatively
infrequently and used the without-thumb grip even less
frequently. These results about P. africanus are very
different from those published in the literature, which
suggest that P. africanus did not use a precision grip
(Napier & Davis, 1959; McHenry, 1983; Walker & Pick-
ford, 1983; Begun et al., 1994). The widespread distribu-
tion of the precision grip in the primate species sampled in
this study support our palaeobiological inferences.
The evolution of grasping behaviour
The classical model proposes a late appearance of
precision grasping, often considered unique to hominids.
Napier (1956, 1960) suggested that grasping objects with
precision requires opposability of the thumb and favour-
able relative lengths of digits I and II. Furthermore, some
authors suggest that precision grasping is linked with
brain organization and the development of cognitive
processes (Napier, 1960; Jones-Engels & Bard, 1996).
Under such hypotheses, hominoid fossils presenting
morphological characters associated with precision grasp-
ing have been argued to be able to use tools (Susman,
1989; Marzke, 1997). The data presented in this paper
refute this evolutionary scenario because species with
highly different hand morphologies and brain structure
use a precision grip. The evolution of grasping abilities in
platyrrhines and catarrhines is much more complex than
a simple trend from power to precision grasping. Indeed,
several species (i.e. P. troglodytes, P. pygmaeus, M. fuscata,
P. papio and C. apella) use the tips of their thumb and
index finger for grasping small objects, demonstrating
that morphological criteria previously used for deducing
grasping ability are not reliable (Susman, 1998). Fur-
thermore, in our quantitative analysis based on the
percentages of use of five simplified categories of grasp-
ing, capuchins are similar to humans (mainly adult
humans) even though they possess a pseudo-opposable
thumb (rather than a truly opposable thumb, capable of
adduction and rotation of its carpo-metacarpus joint).
This functional similarity of two species which diverged
about 34 Ma (Fig. 2) is surprising.
Our findings are in concordant with the evolutionary
model according to which a primitive power grasp was
subsequently modified into a derived precision grasp
(Susman, 1979), although this transition occurred before
the appearance of hominoids. Our data suggest that both
behaviours were already present in the first anthropoids.
It would be interesting to obtain some data on Tarsius and
strepsirhines to determine when precision grasping
appeared.
All our results suggest that grasping has evolved in a
more complex manner than previously realized. Our
observations show that precision grip is far more wide-
spread than previously thought. This is coherent with the
findings that skilled forelimb movements are also present
in other mammals (Ivanco et al., 1996; Whishaw et al.,
1998) and even in amphibians (Gray, 1997). These
movements may not be homologous and represent
convergent evolution of motor patterns that superficially
resemble reaching (Bracha et al., 1990). However, the
similarities in reaching among different mammalian taxa
suggest that the movements are homologous within
mammals. It would be interesting to apply comparative
methods to a far greater range of taxa to assess broader-
scale evolutionary patterns of various grip patterns.
Acknowledgments
We would like to thank the Foundation Singer-Polignac
for their financial support, P. Piras for helping us with
Fig. 3 Time-calibrated supertree of extant and extinct hominoids
represented in this study. The numbers next to the tree indicate the
age (Ma); those next to the stage names indicate their duration (Ma).
For more information, see Fig. 2. [A colour version of this figure is
available online.]
1740 E. POUYDEBAT ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

some statistical analyses, D. Marjanovic for numerous
stylistic corrections, and all the staff of the Zoo of Beauval
and of the Monkey Valley (France).
References
Altmann, G. 1974. Observational study of behavior. Sampl.
Methods Behav. 49: 227–267.
Arnason, U., Gullberg, A., Burguete, A. S. & Janke, A. 2000.
Molecular estimates of primate divergences and new hypo-
theses for primate dispersal and the origin of modern humans.
Hereditas 133: 217–228.
Begun, D. R., Teaford, M. F. & Walker, A. 1994. Comparative
and functional anatomy of Proconsul phalanges from the
Kaswanga Primate Site, Rusinga Island, Kenya. J. Hum. Evol.
26: 89–165.
Begun, D. R., Ward, C. V. & Rose, M. D. 1997. Function, Phylogeny
and Fossils: Miocene Hominoid Origins and Adaptations. Plenum
Press, New York.
Boesch, C. & Boesch, H. 1990. Tool use and tool making in wild
chimpanzees. Folia Primatol. 54: 86–99.
Bracha, V., Zhuravin, I. A. & Burges, J. 1990. The reaching
reaction in the rat: a part of the digging pattern? Behav. Brain
Res. 36: 53–64.
Brochu, C. A. 2004a. Calibration age and quartet divergence
date estimation. Evolution 58: 1375–1382.
Brochu, C. A. 2004b. Patterns of calibration age sensitivity with
quartet dating methods. J. Paleontol. 78: 7–30.
Bush, M., Lovejoy, C., Johanson, D. & Coppens, Y. 1982.
Hominid carpal, metacarpal, and phalangeal bones recovered
from the Hadar Formation: 1974–1977 collections. Am. J. Phys.
Anthrop. 57: 651–677.
Casgrain, P. 2005. Permute! [www document]. URL http://www.
bio.umontreal.ca/Casgrain/en/labo/permute/index.html.
Casgrain, P., Legendre, P. & Vaudor, A. 2004. The R Package for
Multidimensional and Spatial Analysis [www document]. URL
http://www.fas.umontreal.ca/BIOL/Casgrain/en/labo/R/v4/
progress.html.
Christel, M. 1993. Grasping techniques and hand preference in
Hominoidea. In: Hands of Primates (H. Preuschoft & D. Chivers,
eds), pp. 91–108. Springer, Berlin.
Christel, M. I. & Fragaszy, D. 2000. Manual function in Cebus
apella. Digital mobility, preshaping, and endurance in repet-
itive grasping. Int. J. Primatol. 21: 697–719.
Christel, M., Kitzel, S. & Niemitz, C. 1998. How precisely do
bonobos (Pan paniscus) grasp small objects? Int. J. Primatol. 19:
165–194.
Clark, J. D. 1993. Stone artifact assemblages from Members 1–3
Swartkrans Cave. In: Swartkrans: A Cave’s Chronicle of Early
Man (C. K. Brain, ed.), pp. 167–194. Transvaal Museum,
Pretoria.
Cubo, J., Ponton, F., Laurin, M., de Margerie, E. & Castanet, J.
2005. Phylogenetic signal in bone microstructure of saurop-
sids. Syst. Biol. 54: 562–574.
Desdevises, Y., Legendre, P., Azouzi, L. & Morand, S. 2003.
Quantifying phylogenetically structured environmental vari-
ation. Evolution 57: 2467–2652.
Diniz-Filho, J. A. F., de Sant’Ana, C. E. R. & Bini, L. M. 1998. An
eigenvector method for estimating phylogenetic inertia.
Evolution 52: 1247–1262.
Felsenstein, J. 1985. Phylogenies and the comparative method.
Am. Nat. 125: 1–15.
Folinsbee, K. E. & Brooks, D. R. 2007. Miocene hominoid
biogeography: pulses of dispersal and differentiation. J. Bio-
geogr. 34: 383–397.
Fragaszy, D. M. & Boinski, S. 1995. Patterns of individual
diet choice and efficiency of foraging in wedge-capped capuchin
monkeys (Cebus olivaceus). J. Comp. Psychol. 109: 339–348.
Fragaszy, D. M., Baer, J. & Adams-Curtis, L. 1991. Behavioral
development and maternal care in tufted capuchins (Cebus
apella) and squirrel monkeys (Saimiri sciureus) from birth
through seven months. Dev. Psychobiol. 24: 375–93.
Fragaszy, D., Visalberghi, E. & Fedigan, L. 2004. The Complete
Capuchin. The Biology of the Genus Cebus. Cambridge University
Press, Cambridge, UK.
Garland, T. Jr, Harvey, P. H. & Ives, A. R. 1992. Procedures for
the analysis of comparative data using phylogenetically
independent contrasts. Syst. Biol. 41: 18–32.
Godinot, M. & Beard, K. C. 1993. A survey of fossil primate
hands. In: Hands of Primates (H. Preuschoft & D. J. Chivers,
eds), pp. 335–378. Springer-Verlag, Vienna.
Godinot, M., Laneque, L., Verdier, R. & Apriletti, K. 1997.
Quantitative morphology of Eocene primates compared to
living primates: examples of current studies. Geobios 30: 271–
281.
Goodman, M., Grossman, L. I. & Wildman, D. E. 2005. Moving
primate genomics beyond the chimpanzee genome. Trends
Genet. 21: 511–517.
Gradstein, F. M., Ogg, J. G. & Smith, A. G. (eds) 2004. A Geologic
Time Scale 2004. Cambridge University Press, Cambridge.
Graur, D. & Martin, W. 2004. Reading the entrails of chickens:
molecular timescales of evolution and the illusion of precision.
Trends Genet. 20: 80–86.
Gray, L. A. 1997. Evolution of forelimb movement patterns
for prey manipulation in anurans. J. Exp. Zool. 277: 417–424.
Grossman, L. I., Wildman, D. E. & Schmidt, T. R. 2004.
Accelerated evolution of the electron transport chain in
anthropoid primates. Trends Genet. 20: 578–585.
Ivanco, T. L., Pellis, S. M. & Whishaw, I. Q. 1996. Skilled
forelimb movements in prey catching and reaching by rats
(Rattus norvegicus) and opossums (Monodelphis domestica): rela-
tions to anatomical differences in motor systems. Behav. Brain
Res. 79: 163–181.
Iwaniuk, A. N. & Whishaw, I. Q. 2000. On the origin of skilled
forelimb movements. Trends Neurosci. 23: 372–376.
Jones-Engel, L. & Bard, K. A. 1996. Precision grips in young
chimpanzees. Am. J. Primatol. 39: 1–15.
Josse, S., Moreau, T. & Laurin, M. 2006. Stratigraphic Tools for
Mesquite. [Computer program]. URL http://www.mesquite
project.org/packages/stratigraphicTools/.
Laurin, M. 2004. The evolution of body size, Cope’s rule and the
origin of amniotes. Syst. Biol. 53: 594–622.
Laurin, M., Girondot, M. & Loth, M.-M. 2004. The evolution of
long bone microanatomy and lifestyle in lissamphibians.
Paleobiology 30: 589–613.
Lehner, P. N. 1996. Handbook of Ethological Methods. Cambridge
University Press, Cambridge, UK.
Maddison, W. P. & Maddison, D. R. 2006. Mesquite: a modular
system for evolutionary analysis, Version 1.1. [Computer
program]. URL http://www.mesquiteproject.org.
Marjanovic, D. & Laurin, M. 2007. Fossils, molecules, divergence
times, and the origin of lissamphibians. Syst. Biol. 56: 369–388.
Marzke, M. W. 1997. Precision grips, hand morphology and
tools. Am. J. Phys. Anthrop. 102: 91–110.
Evolution of grasping among anthropoids 1741
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

Marzke, M. W., Wullstein, K. L. & Viegas, S. F. 1992. Evolution
of the power grip and its morphological correlates in homi-
nids. Am. J. Phys. Anthrop. 89: 283–298.
McHenry, H. M. 1983. The capitate of Australopithecus afarensis
and A. africanus. Am. J. Phys. Anthrop. 62: 187–198.
Midford, P., Garland, T. J. & Maddison, W. P. 2003. PDAP Package
for Mesquite. [Computer program]. URL http://mesquiteproject.
org/pdap_mesquite/index.html.
Moya-Sola, S., Kohler, M. & Rook, L. 1999. Evidence of
hominid-like precision grip capability in the hand of the
Miocene ape Oreopithecus. Proc. Natl. Acad. Sci. USA 96: 313–
317.
Napier, J. R. 1956. The prehensile movements of the human
hand. J. Bone Joint Surg. 38B: 902–913.
Napier, J. R. 1960. Studies of the hands of living primates. Proc.
Zool. Soc. London 134: 647–657.
Napier,J.R.1980.Hands.PrincetonUniversityPress,Princeton,NJ.
Napier, J. & Davis, P. 1959. The forelimb skeleton and
associated remains of Proconsul africanus. Fossil Mam. Afr. 16:
1–70.
Panger, M., Brooks, A. & Richmond, B. 2002. Wood, older than
the Oldowan? Rethinking the emergence of hominin tool use.
Evol. Anthrop. 11: 235–245.
Parker, S. T. & Gibson, K. R. 1977. Object manipulation, tool
use, and sensorimotor intelligence as feeding adaptations in
cebus monkeys and great apes. J. Hum. Evol. 6: 623–641.
Poux, C. & Douzery, E. J. P. 2004. Primate phylogeny,
evolutionary rate variations, and divergence times: a contri-
bution from the nuclear gene IRBP. Am. J. Phys. Anthrop. 124:
1–16.
Pouydebat, E. 2004. La prehension chez les Primates: approches
ethologique, biomecanique et morphometrique. Ph.D. Thesis, at the
Museum National d’Histoire Naturelle, Paris, 300 p.
Pouydebat, E., Berge, C., Gorce, P. & Coppens, Y. 2005. Use and
manufacture of tools to extract food by captive Gorilla
gorilla gorilla: experimental approach. Folia Primatol. 76: 180–
183.
Pouydebat, E., Berge, C., Gorce, P. & Coppens, Y. 2006a. La
prehension chez les Primates: precision, outils et perspectives
evolutives. C. R. Palevol. 5: 597–602.
Pouydebat, E., Gorce, P., Coppens, Y. & Bels, V. 2006b. Substrate
optimisation in nut cracking by capuchin monkeys. Am. J.
Primatol. 68: 1017–1024.
Roos, C., Schmitz, J. & Zischler, H. 2004. Primate jumping genes
elucidate strepsirrhine phylogeny. Proc. Natl Acad. Sci. USA
101: 10650–10654.
Ross, C., Williams, B. & Kay, R. F. 1998. Phylogenetic analysis of
anthropoid relationships. J. Hum. Evol. 35: 221–306.
Schultz, A. H. 1969. The Life of Primates. Weidenfeld and
Nicolson, London, 341 p.
Shaul, S. & Graur, D. 2002. Playing chicken (Gallus gallus):
methodological inconsistencies of molecular divergence date
estimates due to secondary calibration points. Gene 300: 59–61.
Spinozzi, G., Truppa, V. & Lagana, T. 2004. Grasping behavior in
tufted capuchin monkeys (Cebus apella): grip types and manual
laterality for picking up a small food item. Am. J. Phys. Anthrop.
125: 30–41.
Steiper, M. E., Young, N. M. & Sukarna, T. Y. 2004. Genomic
data support the hominoid slowdown and an Early Oligocene
estimate for the hominoid-cercopithecoid divergence. Proc.
Natl Acad. Sci. USA 101: 17021–17026.
Stern, J. & Susman, R. 1983. The locomotor anatomy of
Australopithecus afarensis, Am. J. Phys. Anthrop. 60: 279–317.
Stucky, R. K. & McKenna, M. C. 1993. Mammalia. In: The Fossil
Record 2 (J. M. Benton, ed.), pp. 739–771. Chapman & Hall,
London.
Susman, R. L. 1979. Comparative and functional morphology of
hominoid fingers. Am. J. Phys. Anthrop. 50: 215–236.
Susman, R. L. 1989. New hominid fossils from the Swartkrans
formation: postcranial specimens. Am. J. Phys. Anthrop. 79:
451–474.
Susman, R. L. 1991. Who made the Oldowan tools? Fossil
evidence for tool behavior in Plio-Pleistocene hominids.
J. Anthrop. Res. 47: 129–149.
Susman, R. L. 1994. Fossil evidence for early hominid tool use.
Science 265: 1570–1573.
Susman, R. L. 1998. Hand function and tool behavior in early
hominids. J. Hum. Evol. 35: 23–46.
Susman, R. L. 2004. Oreopithecus bambolii: an unlikely case of
hominidlike grip capability in a Miocene ape. J. Hum. Evol. 46:
105–117.
Tuttle, R. H. 1965. A Study of the Chimpanzee Hand with Comments
on Hominoid Evolution. University of California, Berkeley, CA,
526 p.
Walker, A. C. & Pickford, M. 1983. New postcranial fossils of
Proconsul africanus and Proconsul nyanzae. In: New Interpretations
of Ape and Human Ancestry (R. L. Ciochon & R. S. Corruccini,
eds), pp. 325–351. Plenum Press, New York.
Whishaw, I. Q., Sarna, J. R. & Pellis, S. M. 1998. Rodent-typical
and species-specific limb use in eating: evidence for specialized
paw use from a comparative analysis of ten species. Behav.
Brain Res. 96: 79–91.
Witmer, L. M. 1995. The extant phylogenetic bracket and the
importance of reconstructing soft tissues in fossils. In:
Functional Morphology in Vertebrate Paleontology, Vol. 1 (J. J.
Thomason, ed.), pp. 19–33. Cambridge University Press, New
York.
Yoder, A. D. & Yang, Z. 2004. Divergence dates for Malagasy
lemurs estimated from multiple gene loci: geological and
evolutionary context. Mol. Ecol. 13: 757–773.
Received 28 March 2008; revised 18 June 2008; accepted 23 June 2008
Supporting information
Additional supporting information may be found in the
online version of this article:
Appendix S1 Animals observed.
Appendix S2 Diameter of the objects for the studied
species.
Appendix S3 Matrix of morphometric (1–16) and
behavioural (17–21) characters used in this study.
Appendix S4 Phylogenetic signal in the characters.
Appendix S5 General linear regression inference mod-
els for the behavioural characters.
Appendix S6 Specific linear regression inference mod-
els for the behavioural characters.
Appendix S7 Correlation between behavioural (depen-
dent) and morphometric (independent) characters
assessed using independent contrasts.
1742 E. POUYDEBAT ET AL.
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y

Appendix S8 Inferred behavioural character values
for extant primate taxa for which such data are unavailable
but for which relevant morphometric data are known.
Appendix S9 Notes about the divergence dates used to
compile the time-calibrated supertree.
Please note: Wiley-Blackwell are not responsible for
the content or functionality of any supporting materials
supplied by the authors. Any queries (other than missing
material) should be directed to the corresponding author
for the article.
Evolution of grasping among anthropoids 1743
ª 2 0 0 8 T H E A U T H O R S . J . E V O L . B I O L . 2 1 ( 2 0 0 8 ) 1 7 3 2 – 1 7 4 3
J O U R N A L C O M P I L A T I O N ª 2 0 0 8 E U R O P E A N S O C I E T Y F O R E V O L U T I O N A R Y B I O L O G Y






























