Embed Size (px)
Citation preview

Evolution of the thyrotropin receptor: a G protein coupledreceptor with an intrinsic capacity to dimerize
Vikt�ooria Kaczur,a,b,1 L�aaszl�oo G. Pusk�aas,b,c M�aaria Tak�aacs,d,2 Istv�aan Andr�aas R�aacz,e
Andrea Szendr}ooi,d,3 S�aara T�ooth,d Zsuzsanna Nagy,d Csaba Szalai,f Csaba Bal�aazs,a
Andr�aas Falus,d Bjarne Knudsen,g and Nadir R. Faridb,*
a Ken�eezy Teaching Hospital of Debrecen, Laboratory for Endocrinology and Human Genetics and Third Department of Internal Medicine,Debrecen, Hungary
b Osancor Bioinformatics Ltd, Watford, Herts, UKc Laboratory of Functional Genomics, Biological Research Center of the Hungarian Academy of Sciences, Szeged, Hungary
d Institute of Genetics, Cell, and Immunbiology, Semmelweis University Medical School, Budapest, Hungarye Department of Evolutionary Zoology and Human Biology, University of Debrecen, Debrecen, Hungary
f Central Laboratory, Heim Pal Hospital of Pediatrics, Budapest, Hungaryg Bioinformatics Research Center, Univeristy of Aarhus, Arhus, Denmark
Received 4 February 2003; accepted 10 February 2003
Abstract
The rapidly escalating number of genome sequences has emphasized the basic tenants of the schema of life. By the same token
comparisons according to specialized function or niche within nature expose genomic strategies to optimize the use of resources and
ensure biological success. Increasing complexity may result from diversification, shuffling, and re-arrangement of an otherwise
limited functional genomic complement. To further test the concept of relative structural plasticity of the TSH receptor we se-
quenced the TSHR gene of two Old World monkey speciesMacaca mulatta and Cercopithecus aethiops, evolutionary removed from
Homo sapiens by >20Myr. Both genes encoded a protein of 764 residues. This structure was 99% homologous between the two
species of Old World monkeys while C. aethiops was 97% and M. mulatta was 96% homologous to H. sapiens. TSHR sequence
comparisons were sought for an additional eight mammals as well as four (two Salmon, Tilapia, and Sea Bass) from teleosts. The
amino-acid sequences of the 14 TSH receptors were similar. The most variable sequences were those of the intracellular tail and the
distal cysteine-rich C-terminus flanking region of the ectodomain, whereas the trans-membrane domain was most preserved. Some
sequences were decidedly H. sapiens specific, while others were primate specific or showed the changes expected of evolutionary
descent. Others, however, exhibited ‘‘cross-species polymorphism,’’ sometimes at quite remarkable evolutionary distances. As op-
posed to H. sapiens the sequence differences may have subtle influences on TSHR function or may affect long-range compensation
for radical changes in adducts. The two Old World monkeys share with other lower mammals the absence of a glycosylation site at
113–115. Sea Bass and Tilapia have four glycosylation sites, whereas the two salmon receptors have only three. Changes in some
critical residues raise questions about variation in function: thus S281 is conserved in all mammals and an important determinant of
negative agonist function of TSHR is replaced by R in Sea Bass. Likewise the K183, found at an important transitional region at
LRR 6 conserved in all mammals, is represented by M in fish and may contribute to TSHR lutenization in fish. There is no evidence
that evolutionary changes in primate receptors are more rapid than that in other mammals and the separation times of different
mammals based on silent nucleotide changes of TSHR are closely parallel to archaeological estimates. Results of correlated mu-
tation analysis, referenced to the rhodopsin crystal structure, affirms dimerization of TSHR transmembrane helices. In addition, it
suggests the involvement of critical lipid-facing residues in the helices in receptor dimerization and oligomerization. We highlight the
Molecular Genetics and Metabolism 78 (2003) 275–290
www.elsevier.com/locate/ymgme
*Corresponding author.
E-mail address: [email protected] (N.R. Farid).1 Present address: Department of Biophysics and Cell Biology, Medical University of Debrecen.2 Present address: Faculty of Veterinary Science, Szent Istvan University, Budapest.3 Present address: Division of Virology, Department of Microbiology and Infectious Disease, National Center for Epidemiology Budapest,
Hungary.
1096-7192/03/$ - see front matter � 2003 Elsevier Science (USA). All rights reserved.
doi:10.1016/S1096-7192(03)00036-2

value of evolutionary informatics and set the stage for dissecting out potential subtle differences in TSHR function associated with
structural variations.
� 2003 Elsevier Science (USA). All rights reserved.
Keywords: TSH receptor; Macaca mulatta; Cercopithecus aethiops aethips; Evolution; Mammals; Teleosts; Dimerization
Introduction
The family of leucine-rich repeat containing G pro-
tein-coupled receptors (LGRs) has recently been ex-
panded by the addition of homologs of as yet unknown
function in mammals and orthologs from lower lifeforms (see [1] for review). This knowledge has provided
an unusual opportunity to pose questions about the
structure, function, and evolutionary history of the
LGRs. These receptors have arisen as a result of
the conjunction of two successful protein–protein in-
teraction and signalling modules, namely leucine-rich
repeats (LRR) and (a bared down) prototypic 7 trans-
membrane G protein-coupled signalling unit (which wehave dubbed signalon). These two modular systems are
bridged by the cysteine-rich C-flanking region (CFR).
The CFRs in most LGRs are anchored by double cy-
steines in their N-termini and PCED (Pro–Cys–Glu–
Asp) motif towards their C-termini. The sequences in
between vary in length and composition among LGRs.
The phylogeny of the CFR suggests that the insertion
herein, considered characteristic of TSHR, to have avery deep evolutionary history is not recent innovation
in TSHR. Indeed, we have recently contended that
remnants of such an insertion may be discerned in LHR
[2].
We report on the sequences of TSHRs from two Old
World monkey tribes to try and span the evolutionary
gap between Homo sapiens and canines, the next highest
order of mammals whose sequence is available to date.Arguably mammalian evolution is no more remarkable
than any other branches of life. The transition from
higher apes to humanoids is arresting not only because
of anthropocentric bias but because of the birth of
cognition, speech, and social bonding in H. sapiens. It is
evident that only a limited number of genes are involved
in this evolutionary transition [3]. It has been suggested
that a post-translational modification of some moleculesarising as a result of a human-specific mutation in a
transferase may be a crucial step in this dramatic tran-
sition [3]. While the Cercopithecidae, we have studied do
not stand at the cusp of this transition, we reasoned that
they would provide an important bridge between human
and lower mammals in evolution of the TSHR. At the
other end of the evolutionary scale there are now avail-
able TSHR sequences from four teleosts, allowing us tostudy the evolutionary history and diversification of the
structure to possibly account for new or additional
function for TSHR in alternate life forms. Even within
mammalia the pleomorphism of growth and prolifera-
tive capacity of thyroid cells from different species has
long been the subject of heated debate [4]. Given on the
one hand the central role of TSH signalling to thyrocyte
physiology and on the other the potential of signallingthrough the receptor by a multitude of G-proteins as
well as other mediators [4–6] raise the possibility that
these variations may be intrinsic to species-specific dif-
ferences in TSHR structure between species.
Materials and methods
RNA purification and cDNA synthesis
Total thyroid RNA was extracted using NucleoSpin
RNA II extraction kit (Macherey-Nagel, D€uuren, Ger-many) according to manufacturer�s instructions. For
cDNA synthesis, 10 lg of total RNA was mixed with
1.5 lg of a HPLC-purified anchored oligo(dT) primer
(50-T25(ACG)-30), and a TSHR gene specific reverseprimer (hTSHR gene nucleotides: 2339–2315; 50-TTATTTTGTAAGTTCCCCTACCATT-30) 40U RNasin
(Fermentas, Vilnius, Lithuania) in a total volume of
14 ll, heated to 75 �C for 5min, and cooled on ice. To
this mixture 4 ll of 5� first strand buffer (Promega,
Madison, WI), 1 ll of 10mM dNTP mix, and 1 ll of200U RNaseH ()) point mutant M-MLV reverse
transcriptase (Promega, Madison, WI) were added in20 ll total volume. Samples were incubated at 42 �C for
2 h. One microliter of cDNA synthesis mix was applied
for PCR.
PCR amplifications, cloning, and sequence determination
Amplification was done under the following condi-
tions: initial denaturation at 95 �C for 1min; 32 cycles ofdenaturation at 95 �C for 35 s, annealing at 58 �C for
40 s, and extension at 72 �C for 1min. The reaction mix
contains 50 nM final concentrations of each primers, 1�KlenTaq PCR buffer (Sigma, Mannheim, Germany),
4U KlenTaq LA DNA polymerase mix (Sigma), and
200 lM dNTP. The primers shown in Table 1 were
utilized in various combinations to generate a series of
overlapping fragments that encompassed the full-lengthTSHR cDNA: 1a and 1b to generate a 1079 nt fragment;
5a and 5b to amplify a 1342 nt long; 3a and 3b to am-
276 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

plify a 422 nt long fragment; 3a and 7b to produce a
796 nt long fragment; and 6a and 2b to amplify a 1030 nt
long fragment.
PCR products were purified with PCR purification kit
(Macherey-Nagel, D€uuren, Germany) according to man-ufacturer�s specifications and cloned with blunt-end li-
gation into pBluescript SK-II ()) plasmid (New England
Biolabs, Hertfordshire, England) that was previouslydigested with EcoRV restriction enzyme (Fermentas,
Vilnius, Lithuania) using standard protocols [7]. Each
amplification was performed in duplicate and cloned
separately. Plasmid midi preparations were done on
Qiagen-tip 100 columns (Qiagen, Hilden, Germany).
Both strands of two positive clones from each cloning
step were sequenced by dye-terminator method with
vector-specific primers (T7 and T3 primers) on an au-tomated sequencer (ABI, Applied Biosystems, Foster
City, CA).
Multiple sequence alignments were performed using
CLUSTAL W (version 1.81), secondary protein struc-
ture prediction using GOR4, and sequence similarity
using Blast2.0 all at SWISS-PORT (http://www.ch.
embnet.org). Mammalian TSH receptor sequences were
submitted to evolutionary rate shift analysis. The evo-lutionary rate shift analysis is based on likelihood ratio
testing and assumes that changes occur at a specific
point in evolution and that these changes are indepen-
dent. The changes are not limited to variable versus
invariable sequences but involves shift between any two
rates [8].
Moreover, mutation in mammalian TSHR predicated
on the rates of silent mutations were computed as inKumar and Subramanian [9] and used to estimate evo-
lutionary distances.
We have taken advantage of the correlated mutation
analysis (CMA) available in the GPCR (http//:
www.gpcr.org, [11]) database on 12 of the 14 receptors
reported here, incorporating the Old World monkey
sequences into it. Likewise we incorporated our data
into analysis performed by Gouldson et al. [12] based onfive TSHR sequences. These investigators embedded
their analysis of the TSHR sequences in the framework
of CMA based on a very large number of receptors from
an additional four GPCR families. Moreover, they re-
lated the placement of conserved and correlated residues
on the TM helices to those in the rhodopsin crystal
structure [13]. Where there was discrepancy in the limits
of the TM domains between our schema [14] and that on
the rhodopsin crystal, we opted for the latter.GenBank Accession Nos. of the TSH receptors stud-
ied were as follows: Rattus norvegicus TSHR M34842,
Mus musculus TSHR U2601, Ovis aries TSHR Y13434,
Bus taurus TSHR U15570, Sus scrofa TSHR AF 338249,
Felis catus TSHR AF218264, Canis familiaris TSHR
X17146, Homo sapiens TSHR M3177, Morone saxatilis
(Striped Sea Bass) TSHR AF239761, Oncorhynchus
rhodurus (Salmon) TSHR_A Q9I949, O. rhodurus TSHR_B Q9I948, Orechromis niloticus (Tilapia) Q98TF4,
Cercopithecus aethiops TSHR AY169399, and Macaca
mulatta AY 169400.
Results and discussion
Receptor divergence over 450Myr
With the description of the two Old World monkey
sequences, we now have information on 14 vertebrate
species including 4 from fish and 8 mammalians. Given
the evolutionary separation of these orders of life (400–
450Myr) it is not surprising that they segregate into two
major phylogenetic groups on the basis of protein se-
quence [15, Figs. 1 and 2]. All four fish species belong tothe ray-fin subgroup of bony fish (teleosts). Even within
this subgroup there is a remarkable degree of species
variation. The case of Tilapia is particularly intriguing.
As a Cichlid subspecies it would have been part of a
major species radiation and specialization after the
emergence from a draught bottle-neck and with the
formation of the Central African lakes [16]. Yet this
burst of speciation occurred only 750,000 years ago.The similarity in regional organization and protein
Table 1
The PCR primers used to amplify TSHR cDNAs from C. a. aethiops and M. mulatta
Primer designation TSHR sequence Primer sequence (50 ! 30) Strand
1a 882–991 TCCTTGAGTCCTTGATGTGT Sense
1b 1961–1941 TGAGAGGCTTGTTCAGAATT Antisense
3a 1505–1530 TTTGCAAGCGAGTTATCGGTGTATA Sense
3b 1927–1903 TAGAATGAGATTGGGGCCATGCATA Antisense
5a )47 to 24 TGAGGCGATTTCGGAGGATGGAGA Sense
5b 1295–1274 ATTGCCCAGGAGAGCCAGCAGA Antisense
6a )6 to 16 TGGAAAATGAGGCCGGCGGACT Sense
2b 1024–999 TTGGACTTTTCCTTGTACCCAACAAT Antisense
7b 2302–2274 ACTTACAAAAC(T/C)GTTTGC(G/C)TATACTCTT Antisense
The numbering of nucleotides is based on hTSHR coding sequence from Nagayama et al. [23]. Combinations of various primers were used to
amplify overlapping PCR fragments of the following sizes. 5a/5b: 1342 bp; 3a/3b: 422 bp; 3a/7b: 796 bp; 1a/1b: 1079 bp; and 6a/2b: 1030 bp. Both
strands were sequenced from at least two independent clones; for most three clones were sequenced.
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 277
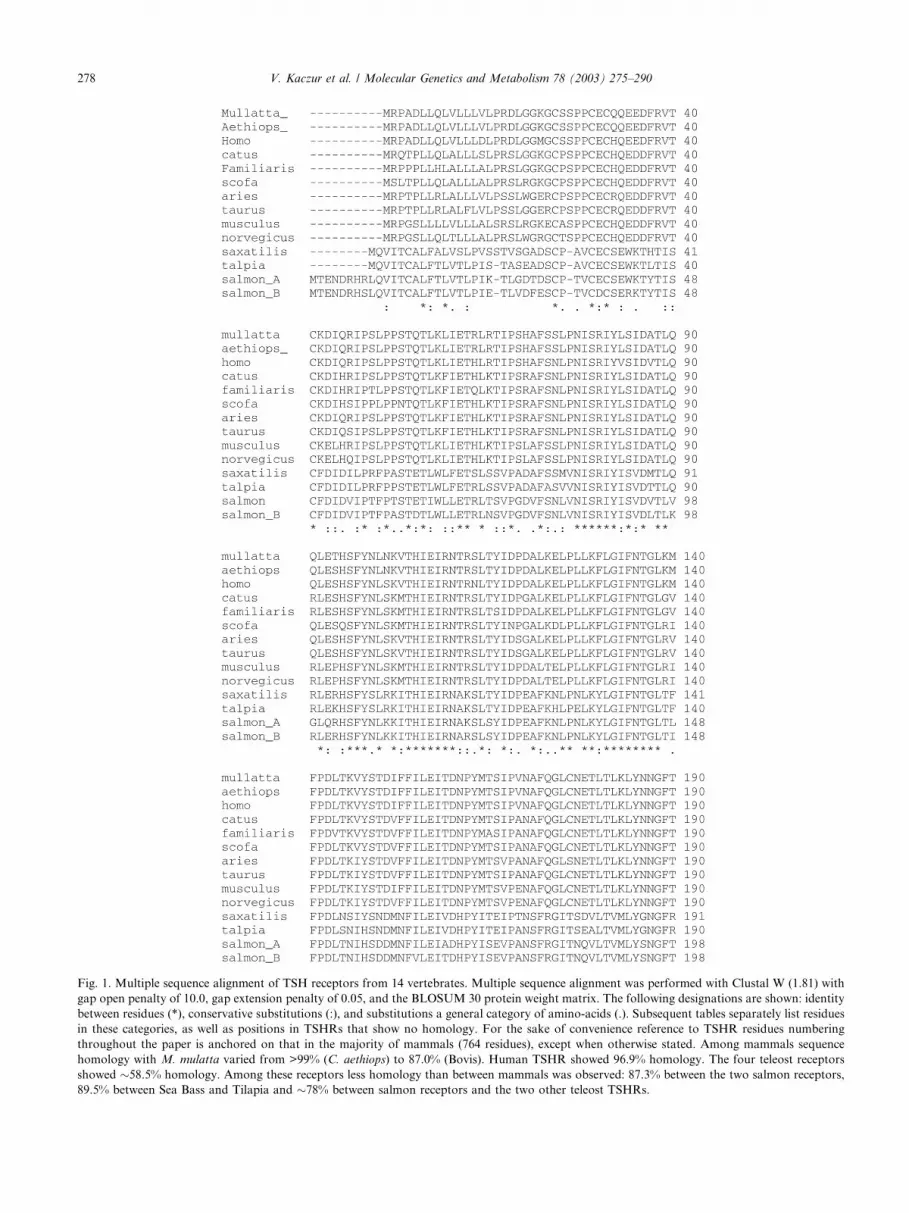
Fig. 1. Multiple sequence alignment of TSH receptors from 14 vertebrates. Multiple sequence alignment was performed with Clustal W (1.81) with
gap open penalty of 10.0, gap extension penalty of 0.05, and the BLOSUM 30 protein weight matrix. The following designations are shown: identity
between residues (*), conservative substitutions (:), and substitutions a general category of amino-acids (.). Subsequent tables separately list residues
in these categories, as well as positions in TSHRs that show no homology. For the sake of convenience reference to TSHR residues numbering
throughout the paper is anchored on that in the majority of mammals (764 residues), except when otherwise stated. Among mammals sequence
homology with M. mulatta varied from >99% (C. aethiops) to 87.0% (Bovis). Human TSHR showed 96.9% homology. The four teleost receptors
showed �58.5% homology. Among these receptors less homology than between mammals was observed: 87.3% between the two salmon receptors,
89.5% between Sea Bass and Tilapia and �78% between salmon receptors and the two other teleost TSHRs.
278 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

Fig. 1. (continued)
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 279

Fig. 1. (continued)
280 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

sequences between Tilapia and Striped Sea Bass suggests
forces other than speciation and thus phylogenetic re-
moval were at play in generating and maintaining the
TSHR sequences. By contrast, in Salmon two TSHR
structures have been identified and that these bear se-
quence differences to suggest that they have different
functions or bind different ligands. This situation is as
yet unprecedented in other species studied. Although
TSHR is expressed in a variety of tissues and given the
variation in mediators or putative mediators is surmised
to subserve different functions [2], all those forms are
transcribed from a single TSHR gene. Variants arise as a
Fig. 1. (continued)
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 281

consequence of alternative splicing or foreshortening[17]. Whether the situation in the Salmon is unique and
thus an evolutionary dead-end experiment or an indi-
cation that as yet unidentified TSHR forms exist cannot
at this time be definitively answered. That the degree of
homology between the four fish sequences are much
lower than those between mammals may either reflect
the time of species separation or possibly a more rapid
rate of change than in mammals. Indeed, the salmonidreceptors approach a degree of homology similar to that
between humans and rodents, a separation estimated at
100Myr.
The basic organization of the TSH receptor has been
well preserved over evolution and indeed it harks back
to that of the earliest LGR forms described [1,2]. The
LRR repeats and transmembrane domains are invariant
in length. The region of most variability is the cysteinerich CFR and less so the intracellular tail. Fish receptors
are characterized by insertions in CFR, exaggerated in
the A form of the salmonid receptor whereas single
residue deletions (at separate locations) are found in cat
and bovine receptors. Fish have 8–10 residue insertions
in the intracellular tail and a longer leader peptide in
salmonid TSHR (Fig. 1).
As anticipated the four teleost sequences showgreater structural affinity to each other than they do
with mammals, particularly in the TM domain (Figs. 1
and 2). The two salmon TSHRs sequence differ princi-
pally through and insertion (residues 321–346) in the
CFR of TSHR-A receptor. Salmonids are the only
vertebrates with two TSHR to be identified to date and
both show divergence from the two other bony fish se-quences (Fig. 2).
TSH receptors of Cercopithecidae
The two Old World monkeys TSHRs were >99%
homologous to each other and 97% to that ofH. sapiens.
The two Cercopithecidae represent two distinct pri-
mate�s families and are presumed to have separatedfrom H. sapiens more than 23Myrs ago [15–18]. The
TSHR of the two Cercopithecidae species are identical
except for 17 bp at the DNA level (10 of which result in
silent mutations) and 6 positions at the protein level
(Fig. 1), and only at position 296 is there a replacement
substitution (Tables 1 and 2). The rate of silent muta-
tions allow us to calculate an evolutionary separation
time of approximately 10 million years between the twoOld World monkey families (also see below).
Macaca is an Asian branch of the Old World monkey
tribe Papioni, the majority of whose clades are found in
Africa. Cercopithecus aethiops aethips represent a dis-
tinct Cercopithecidae tribe from Papioni [18–21]. The
evolutionary separation predicated on the evolutionary
clock of TSHR sequences may not be that inconsistent
with the phylogenetic data, although some nuclear geneswithin Cercopithecidae, especially Macaca, yielded in-
congruent results with a consequent anomalous place-
ment within the Papioni [21].
Table 2
Differences in the mature TSHR sequences between humans and Old
World monkeys
Residue Cercopithecidae Homo sapiens
32 Q H
63 R H
74 S N
83 L V
87 A V
101 N S
113 S N
192 I V
235 H Q
331 D G
333 G S
346 A T
389 V I
393 N S
601a Y H
727 E D
759 T M
Only residues in which Homo differ from both Old World monkeys
are considered.a The placement of H in this position was based on the original
hTSHR sequence reported by Nagayama et al. [23]. Histidine in that
position alters the basal activity of the receptor and its coupling to Gq/
11, analysis of other clones by the same workers and the results of
other investigators affirms a conserved Y in that position in all verte-
brates.
Fig. 2. Phylogenetic relationship between the 14 TSH receptor se-
quences considered in this study. Two major families are distinguished:
teleosts and mammals and within the latter primates show up as a
distinct clade. Clustering of other mammals: rodents, ungulates and
carnivores are in keeping with speciation.
282 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

As the proline 296 was found in two unrelated indi-
vidual M. mulatta it is unlikely to be a mutation in a
particular individual. The other five residue changes
between the two Cercopithecidae are conservative and
cluster in the intracellular domain: 94 (S in aethiops, T
in mulatta), 635 (M in aethiops, I in mulatta), 731 (G in
aethiops D in mulatta), 736 (E in aethiops, Q in mul-
atta), and 741 (I in aethiops, L in mulatta). With theexception of 296 at which L is invariant in all vertebrate
TSHR, these substitutions may not necessarily affect
TSHR secondary structure or function. Residue 296
occurs in the proximal part of CFR, a domain with a
fair degree of structural flexibility. The replacement of L
with a P in this domain is not without precedent, in that
in Mus a canonical L330 is replaced by P and at 347 P is
characteristic of rodent TSHR, whereas in other verte-brates there are a variety of hydrophobic residues.
As far as the differences between human and Old
World monkey TSHR are concerned only four, includ-
ing that at position 296 were their repeat substitutions,
and only one within the mature receptor (T 760 M).
Interestingly, the substitutions in the leader peptide
(V14D), (K22M) were characteristically H. sapiens.
Some conservative or same group residues are charac-
teristically human.
Other changes in residues may reflect evolution-re-
lated changes within primates. Thus their four charac-teristic changes: 294 (T in mulatta, S in aethiops and
Homo), 645 (I in mulatta and Homo M in aethiops), 731
(D in mulatta G in the two other primates), and 736 (Q
in mulatta E in the two other primates). None of these
residues are the sites of recognized TSHR mutations
associated with change in receptor function. We cannot,
however, exclude the possibility that the changes be-
tween primate sequences may not result in subtlechanges in ligand-binding or receptor signalling [22] gi-
ven the differences in volume, potential side-chain in-
teraction, and hydrophobicity.
Table 3
Replacement substitutions of residues in the mature TSHR of 10 mammalian species
Residue Region Mull Aethio Homo Cat Dog Pig Sheep Cow Mus Rat
22 K K M K K K E E K R
23 G G G G G G R R E G
46 NCR R R R R R S R S R G
70 LLR1 H H H R R R R R L L
116 LLR3 Y Y Y Y S Y Y Y Y Y
139 LLR4 K K K G G R R R R R
169 LLR5 V V V A A A A A E E
235 LLR8 H H Q Y Y Y Y Y S S
296 CFR P L L L L L L L L L
307 CFR Q Q Q R R R R W R R
317 CFR A A A A T A A A I V
323 CFR H H H D D Y Y Y Y Y
329 CFR N N N Y Y D D del D G
330 CFR L L L L L L L La P L
334 CFR I I I H H S S Sa S H
337 CFR Y Y Y Y Y N Y Ya Y Y
346 CFR A A T T T T T Ta S G
347 CFR H H H R D H H Qa P P
360 CFR E E E del E E E Ea E E
426 TM1 L L L La L L L La L P
539 TM4 C C C Ya Y Y Y Ya Y Y
548 TM4 C C C Ca C C C Ca S S
588 TM5 T T T La L L L La L L
595 TM5 V V V Ia I T I Ia V V
617 I2 G G G Ga G G G Ga R R
660 TM7 K K K Ka K K K Ka K G
712 ICT P P P Sa S S S Sa C C
726 ICT H H H Ra R Q P Pa Q Q
738 ICT V V V Da E D D Da T T
741 ICT L L L La L L L La L P
743 ICT E E E Ea E E G Ea G G
749 ICT P P P Pa P H P Pa P P
751 ICT K K K Ka K K Q Qa L L
755 ICT I I I Ia I I T Ta I I
760 ICT T T M Na N K K Ka K T
aBecause of respective deletions (del) in residues 329 of cow and 360 of cat, all residues numerically above that position shown by the asterisk should
have one subtracted. Numbering of residues is based on 764 residues in 8 mammals including 21 in the leader peptide. NCR, N-terminus cysteine-rich
region; LLR, leucine-rich repeats; CFR, C-flanking region; I2, second intracellular loop; TM, transmembrane helix; ICT, intracellular tail.
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 283

A substitution of important functional impact is S113
N, the latter creates the NLT N-terminal glycosylation
signal, which appears to only exist in humans (see [17] for
review). This substitution can result potentially from a
single base-pair substitution (AAT ! AGT).
Variations in mammalian TSHR sequences. Any impli-
cations for function?
With the widening of sequence comparisons in all 10
available mammals separated by in excess of 100 millions
of evolution differences in sequences [9,15] would be ex-
pected. As anticipated these predominate in the leader
peptide, intracellular tail, particularly its terminal end.
Some of these changes are primate-specific, some show
evolutionary gradients, and yet some, interestingly, dis-
play across species polymorphism. Most of the latter fall
in the category of conservative or related residue substi-
tutions. Over the 10 TSHR sequences specific positionmay fall into more than one category outlined above.
Residues 2–5 of the leader peptide are instructive in
that the first four residues are identical in primates,
Table 4
Same class substitutions in mammalian TSHRa
Residue Region Mul Aethio Homo Catb Dog Sheep Cowb Pig Mus Rat
10 Leader V V V A A A A A V T
16 Leader P P P P P P P P S P
25 NCR S S S P P P P P A T
49 NCR S S S S T S S P S S
53 NCR S S S S S S S N S S
74 LLR1 S S N N N N N N S S
87 LLR2 A A V A A A A A A A
101 LLR2 N N S S S S S S S S
113 LLR3 S S N S S S S S S S
119 LLR3 P P P P P S S P P P
120 LLR3 D D D G D G G G D D
176 LLR5 C C C C C S C C C C
215 LLR7 V V V A A V V V A A
217 LLR7 D D D D D D G D D D
222 LLR7 G G G G G G A A G G
243 LLR8 S S S S S S S P S S
308 CFR S S S S S G G S N N
315 CFR V V V V V A A V V V
316 CFR N N N N N S S N N N
319 CFR N N N N N N N N R R
320 CFR S S S G G G G G G G
322 CFR L L L F F F F F I V
331 CFR D D G G G G G G G G
333 CFR G G S S S G G S N N
335 CFR V V V A A A A V V V
348 CFR N N N S S S S S S S
392 CFR D D D G G G G G D D
393 CFR N N S N N S S S N N
437 TM1 L L L I I I V V L F
447 TM2 N N N T T T T T T T
540 TM4 A A A A A V V A T T
548 TM4 C C C C C C C C S S
599 TM5 C C C S S A A S S S
615 I2 P P P T P P P P P P
708 ICT G G G G G G G G G A
713 ICT P P P P P S P P P P
716 ICT S S S S S S S S S N
718 ICT D D D G G G G G G G
725 ICT T T T T T P P T P P
744 ICT N N N N N N N N N S
752 ICT G G G S G D D D G G
762 ICT T T T T T T T P T T
763 ICT V V V V V V V V A A
aThese residues as shown as (.) in the multiple alignment printout.bAs previously subtract 1 for cat residues 360 and above and 329 and above for cow. 447 is the first residue of TM2. As an example of what
constitutes this grouping all the residues at position 10 are small; however V is aliphatic, T is polar, and A is non-polar and T is more soluble than the
other two. At position 120 both G and D are small and D but not G is charged. We suggest that these differences in the characteristics of alternative
residues may well influence receptor structure and function.
284 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

position 2 shows R ! S in pig, 3 P ! Q in cat, P ! L in
pig whereas residue 4 which is A in primates may display
a trend to evolutionary gradient in that it is G in ro-
dents, T in ruminants, pig and cat but P in dog. Position
7 shows an aspect of across species polymorphism in
that it is Q in primates, cat, pig, and rat but H in dog, R
in ruminants, and L in mouse. Position 17 is R in all
TSHRs expect ruminants and position 20 is G except forsheep, rat (W), mouse, and pig (R). Because the leader
peptide is free from functional evolutionary pressures,
variation in its sequence gives us some insight into re-
ceptor diversification, within the limits of its short
length. Interestingly, sequence alignment of only the first
60 residues of all 14 TSHR proteins result in phyloge-
netic separation identical to that using the full-length
sequences (not shown).Not surprisingly, there are relatively fewer radical
residue substitutions in the mature receptor, which we
will now consider (Tables 3–5).
Residue 46 is located in the N-terminus cysteine rich
region in a domain important for TSH binding. The
most frequent residue R is replaced by S in cow and pig
but with a relatively hydrophobic G in rat and would be
expected to have an impact on ligand binding [17].
Likewise it is conceivable that replacement of R with G
in rodent second intracellular loop may influence the
association with trimeric G-protein and thus signalling.
Many of the substitutions in LLR fall in the middle part
of a repeat and may thus be buried or constrained to
influence receptor function. It is also worthy to note that
the TM4 is the least mobile of the seven helices and thusthe changes in two cysteines between species is likely to
have less of an effect than if it had occurred elsewhere
[15,24]. All these conjectures can, however, only be tes-
ted by head to head comparisons of homologous and
heterologous TSHR on a slate of expressed recombinant
receptors in cell systems which allow for desensitization,
internalization, and re-cycling.
The sequence flexibility in mammalian TSHR ofcysteine-rich CFR of the ectodomain and the cytoplas-
mic tail reflects that in vertebrates in general. That CFR
represents a bridge between a protein/protein interaction
module and the signalon and that it is cleaved off, at
least in the human receptor (and possibly its dispens-
ability after ligand binding), may justify the marked
structural flexibility. The case of the intracellular
Table 5
Conservative (:) residue substitutions in selected TSH R regionsa
Residue Region Mull Aeth Homo Catb Dog Sheep Cowb Pig Mus Rat
32 NCR Q Q H H R R R H H H
43 NCR D D D D D D D D E E
44 NCR Q Q Q H H Q Q H H H
415 TM1 K K K K K K K R R K
427 TM1 L L L L L L L L L M
433 TM1 I I I I V I I I I V
465 TM2 M M M M M L L M V V
478 E1 H H H H H Q Q Q H H
479 E1 S S S S S S S S S T
496 TM3 T T T A T T T T T T
521 I2 Y Y Y Y Y Y H Y Y Y
528 I2 R R R R R H R R R R
533 I2 I I I M I I I I I I
557 TM4 L L L L L L L L M M
575 E2 E E E E E E E E D D
585 TM5 F F F L L L L L L L
591 TM5 I I I I I I I I V V
592 TM5 V V V V V I I V V V
596 TM5 I I I I I I I I V I
601c TM5 Y Y H Y Y Y Y Y Y Y
612 I3 Q Q Q Q Q H H Q Q Q
621 I3 K K K K K R R K K K
635 TM6 I M I M M M M M M M
648 E3 I I I L L L L L L L
649 E3 L L L M M M M M M M
657 I3 S S S T T T T T T T
661 TM7 I I I I I I I I I V
684 TM7 A A A T A A A A A A
E is external intervening loop.
415 is first residue of TM1, 621 last residue of I3, 648 first residue of E3, and 657 its last residue and 648 is the last residue of TM7.a These residues are indicated by (:) in the multiple alignment output.b Subtract 1 from all sequence numbering starting with 415.cH at 601 in hTSHR is based on Nagayama et al. [23] and was subsequently shown to be Y. It is left here to emphasize the potential deleterious of
what is considered to be an almost equivalent residue.
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 285

domain is, however, less easy to explain on the basis ofcurrent knowledge. The cytoplasmic tail is likely in-
volved in most post-binding events including signalling,
desensitization, internalization, and re-cycling. The
variability in structure between species may imply
qualitative and possibly quantitative differences between
species in these attributes. It has, for example, been
implied that the cytoplasmic tail encodes sequences for
TSHR internalization and re-cycling but the motif in-volved was not disclosed [25]. It is instructive that the
mouse LH receptor has a motif, absent in rat, pig, and
human receptors, in the terminal tail which determines
receptor re-cycling [26]. It is apparent (results not
shown) that hTSHR, which recycles [25], lacks this motif
and must employ other structural cues.
An eighth helical structure predicted by the resolu-
tion of the rhodopsin crystal [13] follows on TM7. Iteffectively forms a fourth intracellular loop as a con-
served cysteine following on that domain is pamuloly-
ated thus attaching it to the membrane. To our
knowledge this covalent linkage has not been directly
demonstrated for the TSHR. We wish to stress the po-
tential relevance of some conservative substitutions on
receptor function. Thus the hTSHR originally reported
by Nagayama et al. [23] and which we have used here inour tables by way of emphasis, designated residue 601 as
an H whereas in other sequences it was Y, confirmed by
the authors when further clones were sequenced. Ap-
parently Y601 forms an important molecular switch,
determining the specificity of engagement with the G
protein. The replacement of Y601 with H abrogates
receptor basal constitutive activity and its ability to
couple to Gq/11. Tyrosine has a larger volume, is moresoluble, and slightly more acidic than H [27,28].
It is worthy to note that although none of many of
the conservative substitutions are found at the beginning
or the ends of TM and inter-helical loops, (Tables 4 and
5), no example of a radical substitution could be dis-
cerned. These substitutions may, nevertheless, influence
the angle/charge of the ends of the TM helices and may
modify binding and/or signal transduction. It is worthyto note that none of the substitutions found correspond
to described sites of mutations or polymorphisms.
Structural motifs and characteristic residues
There are a number of structural motifs, which are
characteristic of TSHR as opposed to LHR and FSHR
or specific regions or residues thought to be essential orimportant for its specific function. Three signature mo-
tifs have been identified for all 14 TSHR sequences and
include: NISRIY predominantly at the beginning of
LLR2 and which comprise the first glycosylation site of
hTSHR, THIEIRN spanning the first eight residues
of LL3, whereas the third LKNPQE [29] occurs just
distal to the end of the 50 residue ‘‘add on’’ in CFR
(CFLANS). These conserved motifs very likely playimportant roles in the packing of the contact surface for
the ligand (the first two) or signalling (the third). While
experimental evidences have not directly identified these
regions as to be that important [17], some clues are being
provided by LLR modelling [30]. Considerations of the
secondary structure of the LLR region of the ectodo-
main indicates that the first eight residues of each LLR
describe a b coil whose external surfaces provide contactwith ligand [30, Fig. 3]
Cysteines
The cysteines in the extra-cellular domain consist of
two paired sets at the N- and C-terminus of the ecto-
domain, a general orientation typical of LLR receptors,
and an orphan at 176 (primate numbering) [17]. The
latter is the only one not preserved though the 14 se-quences: it is replaced by S in sheep and T in teleosts.
The former two cysteine sets are thought to be essential
for maintaining the secondary structure of the ectodo-
main, there is, however some uncertainty with respect to
the pairing of the C-terminus cysteines [2]. The equiva-
lent of primate cysteines 24, 29, 31, 41, 283, 284, 301,
390, 398, and 408 are strictly conserved in all 14 recep-
tors. Cysteines 539 and 599 appear to be strictly primatespecific.
Glycosylation sites differ among species
The hTSHR boasts six potential glycosylation sites.
All six have now been directly demonstrated to be at-
tached to a glycan [17,31]. The equivalents of those at
77, 198, and 302 are preserved in all 14 TSHRs (Figs. 1
and 3). As we show here the glycan attachment site at113 (–115) is apparently specific for H. sapiens, whereas
that at 177 (NET) appears to be characteristic of
mammals. The motif NISR (77 primate numbering) is
present in all vertebrates and its apparent essential role
to receptor trafficking and insertion into the plasma
membrane, appears to have been related to an experi-
mental artefact [31]. Systematic studies indicated that
glycans in four glycosylation sites, irrespective of loca-tion, were necessary for expression and biological ac-
tivity, although efficiency of both processes increased
with the involvement of more sites [31]. Interestingly Sea
Bass and Tilapia (but not Salmon) have a distinctive
potential glycosylation site (NTLT: 276–278 in the for-
mer and 277–279 in the latter species) at the N-terminus
of CRF. Both salmon TSHR_A and _B have only three
glycosylation sites (85–88 NISR; 206–209 NGTR; 310–313 NTLA).
The four site minimum formulation [31] would pre-
clude the two Salmon TSHRs being expressed or to
exhibit biological activity when, in fact, they are effi-
ciently expressed in COS-7 cells and efficiently stimu-
lated by bovine TSH [32], indicating that more than just
the number of glycan adducts must be involved. We
286 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

suggest that modification of ternary structure affected bysequences changes in LLRs (and possibly elsewhere)
may modify glycan orientation such that three sites
would be necessary and sufficient for biological function
of the receptor. Glycan adducts are important for
folding and assembly, secretion, and regulation of dis-
position. Both LH and TSH exhibit a unique sulphated
N-linked oligosaccharides, and these are present in all
vertebrate hormones [33].
Residues apparently important for TSH function
As a result of homologous gene segment exchange,
natural mutations as well as site-specific mutagenesis,
certain extracellular domain regions or residues were
deemed important for TSH association and receptor
activation. These include the regions 201–211 (primate
numbering), 222–230, 295–302, and Y385 [17]. The ty-rosine as position 385 or equivalent is preserved in all
vertebrate receptors. The sequences above are conserved
among the 10 mammalian sequences, but diverge in
teleosts, the least divergent being 387–395 equivalentthat follows on the third signature motif (LKNPQE)
and Y385 in CFR. These comparative studies may
suggest that important contact surfaces for ligand in
mammals and fish may have diverged to accommodate
differences in the details of ligand structure and receptor
function (Fig. 2).
Mutation at human 183 (K183R) results in preg-
nancy limited familial hyperthyroidism, related to in-creased sensitivity of TSHR to CG [34]. This residue is
conserved in the LHR and falls within LLR6 in a
transition region in which domain-exchange experi-
ments between the two receptors suggested a switch in
specificity. K183 is preserved in all mammals but its
equivalent position is substituted for M in teleosts.
Systematic mutagenesis of this residue has now been
carried out to [30] allow the surmise that the substitutionin teleosts would impart lutenizing attributes upon
TSHR. Indeed, the K mutation results in one of the
lowest EC50 for TSH and the lowest for GC [30]. This is
Fig. 3. Sequences constituting b strands at the beginning of the 9 leucine rich repeats (LRR) of vertebrate. TSHR The sequences in LLR3 completely
encompass the TSHR-specific motif, whereas the NISRIY motif overlaps the N-terminus of LLR2b strands. The strand have the following consensussequence: LXXLXLXX where X is any residue and L is leucine or equivalent (isoleucine, valine, or unusually methionine or alanine). Threonine is
used in TSHR in 2b strands. Apparently the side-chains of L or equivalent point towards the hyrophobic protein core, whereas the intervening (Xs)
residues are on the protein surface and thus potentially make contact with the ligand. The b strands of the LLR2 and LLR3 are the most conserved.The fifth position (K in mammals M in fish) of LLR 6 (K183 in mammals) is predicted to form a salt bridge in mammals with E (157 in mammals) in
the similar position in LLR5. The model predicts that the change to M would result in the disruption of that interaction and possible alternative
interactions with more proximal residues [30]. It is likely that teleost TSH may acquire additional and/or different biological functions as a con-
sequence.
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 287

without doubt a factor in the ability of fish TSHR (ex-emplified by Sea Bass) to bind and be activated by LH
[29]. By contrast salmon TSHR is reported not to be
expressed in gonads, but on thyroid epithelial cell at a
critical time of development [36].
Apparently K183 forms a salt bridge with E157, a
residue conserved in all LGRs and mutation of the
former to R results in the disruption of the salt bridge
and establishment of an association with a distal D232;change to M would likely result in the establishment of
alternative partners. These mutations clearly cause dif-
ferences in the ternary structure of adjacent LLR and
electrostatic potential to allow GC to be a more potent
ligand for the TSHR (Fig. 3).
S281 is important in receptor silencing
S281 is part of a motif, HSCCF, conserved in all threeglycoprotein hormone receptors and all TSHRs exam-
ined here except for that of Sea Bass (Fig. 1). S281
maintains the receptor in a ‘‘silent’’ constrained state.
Data from natural and introduced mutations suggest
that the degree to which TSHR is constitutively activated
and related to the measure to which the substituted res-
idue modifies the backbone of the protein [35–38]. Dis-
ruption of the local structure introduced by mutatingC283, which is also important for the maintenance of
TSHR ternary structure, releases the suppressive influ-
ence of S281 [37]. Moreover, long range interactions
between the ectodomain and the external inter-helical
loops make for a composite structural module, which can
be disorganized by S281 mutations [38]. In Sea Bass
TSHR R is substituted for S281, a substitution which
might be expected to be associated constitutive activityof the receptor. Careful examination of the data in
Kumar et al. [29], suggests this to be the case
(M. Szkudlinski, personal communications). The exact
physiological consequence of this observation is unclear.
Receptor cleavage
We also note that all three primates have a distinctive
306–307 MQ doublet as opposed to I and R in all othermammals with the exception of Aries which exhibits W
at 307. As this motif is immediately upstream from site I
for TSHR cleavage [17], and given the role that has been
postulated for the 50 residue ‘‘add on’’ sequence [39] in
CFR (CFLANS) cleavage in the pathogenesis of
Graves� disease, it is possible that Old World monkeys
receptors are cleavable. There is, however, no informa-
tion that primates develop spontaneous autoimmunehyperthyroidism. Adamlysins postulated to be involved
in hTSHR cleavage digest membrane anchored struc-
tures at defined distances from the membrane, have
recognition motifs, although there is less certainty as to
whether they have consensus cleavage sequences [40].
The MQ doublet might be important for enzyme
docking or guidance for downstream cleavage.
Mammalian TSHR gene evolution is average
It is apparent, even though remarkable sequence
conservations have been maintained, the separation of
teleosts from mammals (Fig. 2) allow for independent
change in structure to potentially permit new functional
niches for the receptor. We put to the test the possibility
that there may be differences in the evolutionary rate
shifts between primates and lower mammals [8]. Therewas not enough TSHR sequence divergence between
these mammal groups for the rate shift algorithm to
detect differences in evolutionary rates. There is a notion
that mutation rates among mammalian lineages may be
determined by generation time and that some genes
within different mammalian genomes may display sev-
eral fold differences from those in others. Recent work
comprising 8600 genes among 326 mammals showed nodifference in mutation rates according to generation time
and relatively homogeneous mutation rates among dif-
ferent genes [9]. Because of the acknowledged limited
sensitivity of the evolutionary rate shift algorithm, we
estimated the evolutionary separation times of mammals
on the basis of differences in silent mutations in TSHR
and related our results to those of the reputed archae-
ological separation times. The correspondence betweenthe two sets of figures was uncannily close: thus the
separation of C. aethiops fromM. mulatta was estimated
at 11.5Myr (10), between Homo and mulatta at 25Myr
(23), and between mouse and Homo at 103Myr (109);
the figures in brackets giving the archaeological sepa-
ration times. It is of interest that hominoids show a
higher rate of evolution compared to rodents, when a
separate and distant mammalian lineage is used as anoutgroup [9]. In essence, the evolutionary rate of
mammalian TSHR, including those in primates, is av-
erage for the totality of mammalian genomes.
TSHR is a receptor that dimerizes
Dimerization of TSHR was entertained for some time
on the basis of indirect evidence [41,42], and more re-cently based on direct experimental demonstration [43].
There is also evidence for high order oligomers [42].
Apparently homodimerization occurs after TSH cleav-
age and involves covalent bond formation between the
residual ectodomain stumps [44]. This covalent interac-
tion between receptors likely enhances signalling, thus
compensating for the low abundance of TSHR on thy-
roid cells [44]. There is a large body of evidence tosuggest that activation of many GPCRs involve dimer
formation (see [45,46] for review). Rhodopsin dimer-
ization on membranes was very recently demonstrated
by atomic-force microscopy [47].
The correlated mutation analysis (CMA) [10] we per-
formed affirms dimerization of TSHR transmembrane
helices. We have confirmed with a much larger number
288 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290

TSHR sequences the earlier finding [12] that 21/24 TMmutations are predicted to fall on the external surfaces of
the relevant helices. The distribution on the external as-
pects of the helices also support the notion of contact or
domain-swapped dimers occur between the external as-
pects of helices 5 and 6 [12]. Other helices [2,3,4] may be
involved in high-order oligomerization [12]. We empha-
size that the CMA analysis was performed with reference
to the rhodopsin crystal [13], that the CMA analysisembeds the TSHR helix structure within a large number
of GPCRs, with their correspondingmutations and lastly
CMA is upheld by an independent bioinformatics ap-
proach, evolutionary trace analysis [48].
In a different but related vein, we identified 24 TM
residues whose mutations modified receptor function: 5
inactivating and 19 activating [11,24]. The former were
limited to TM1 through 4. Except for two (420 in TM1and 597 in TM5), all these residues were either strictly
conserved or conservatively substituted in all 14 TSHR
sequences; all 24 were absolutely conserved in mammals.
In teleosts V420 was replaced by S and V597 by I. TM6
exhibited a stretch of five residues (629–633), each of
which when mutated result in a constitutively activated
receptor [24]. The characteristics of this region reflect,
on the one hand, the importance of the wild-type con-formation of this mid-region (�1.5 turn of the a-helix)and on the other a short-range domain effect in helical
orientation transduced through to its cytoplasmic end
and il3 to affect engagement with the trimeric G-protein.
Conclusion
The availability of TSHR sequences including 2 we
now describe from lower primates have allowed us new
insights into TSHR evolution, conservation of essential
function as well as the potential of diversification, pos-
sibly to fill niches of function consummate with survival
and success in a particular environment. A case is made
for the structural determinants for receptor dimerization
as an essential pathway to TSHR signalling.
Acknowledgments
The authors gratefully acknowledge the conscientious
technical assistance of Mrs. Krisztina Farkas and Ce-
cilia Ker�eekgy�aart�oo-Zalka (Institute of Genetics, Cell-andImmunbiology, National Center for Epidemiology,Budapest, Hungary). Blood collection and biopsies from
C. aethiops and M. mulatta were performed by Dr.
L�aaszl�oo Nemes (Chief: Prof. Dr. Gy€oorgy Berencsi) (Na-tional Center for Epidemiology, Budapest, Hungary)
and Dr. Peter Kottra (Chief: Prof. Dr. J�aanos Urb�aan)(Institute for Psychology of Hungarian Academy, Bu-
dapest). The work was supported in part by a special
grant from Osancor Biotech Inc. and from the Hun-garian Ministry of Health OKTA Grant T037184.
References
[1] S.Y. Hsu, A.J.W. Hsueh, Discovering new hormones, receptors
and signalling mediators in the genomic era, Mol. Endocrinol. 14
(2000) 594–604.
[2] V. Kaczur, I.A. Racz, A. Szendroi, M. Tackas, N.R. Farid, The
cysteine-rich C-flanking region of the thyrotropin receptor has
very ancient phylogenetic origins: implications from sequence
analysis, J. Endocr. Genet. 3 (2002) 43–56.
[3] J.G. Hacia, Genomes of the apes, Trends Genet. 17 (2001) 637–
645.
[4] T. Kimura, A. Van Keymeullen, J. Goldstein, A. Fusco, J.E.
Dumont, P.P. Roger, Regulation of thyroid cell proliferation by
TSH and other factors: a critical evaluation of in vitro models,
Endocr. Rev. 22 (2001) 631–656.
[5] K.L. Laugwitz, A. Allgeir, S. Offermanns, K. Spicher, J. Van
Sande, J.E. Dumont, G. Schultz, The human thyrotropin recep-
tor: a heptahelical receptor capable of stimulating members of all
four G protein families, Proc. Natl. Acad. Sci. USA 93 (1996)
116–120.
[6] A. Allgier, K.L. Laugwitz, J. Van Sande, G. Schultze, J.E.
Dumont, Multiple G-protein coupling of the dog thyrotropin
receptor, Mol. Cell. Endocrinol. 127 (1997) 81–90.
[7] J. Sambrook, E.F. Fritsch, T. Maniatis, Molecular Cloning: A
Laboratory Manual, second ed., Cold Spring Harbor Laboratory
Press, Cold Spring Harbor, New York, 1989.
[8] B. Knudsen, M.M. Miyamoto, A likelihood ratio test for
evolutionary rate shifts and functional divergence among proteins,
Proc. Natl. Acad. Sci. USA 98 (2001) 14512–14517.
[9] S. Kumar, S. Subramanian, Mutation rates in mammalian
genomes, Proc. Natl. Acad. Sci. USA 99 (2002) 803–808.
[10] G. Vriend, WHAT IF: A molecular modelling and drug design
program, J. Mol. Graph 8 (1990) 52–56.
[11] http://www.gpcr.org.
[12] P.R. Gouldson, M.K. Dean, C.R. Snell, R.P. Bywater, G.
Gkoutos, C.A. Reynolds, Lipid-facing correlated mutations and
dimerization in G-protein coupled receptors, Protein Eng. 14
(2001) 759–767.
[13] K. Palczewski, T. Kumasaka, T. Hori, C.A. Behnke, H. Moto-
shima, B.A. Fox, I. le Trong, D.C. Teller, T. Okada, R.E.
Stenkamp, M. Yamamoto, M. Miyano, Crystal structure of
rhodopsin: A G protein-coupled receptor, Science 289 (2000) 739–
745.
[14] Z. Lin, A. Shenker, R. Pearlstein, A model of the lutropin/
choriogonadotropin receptor: insights into the structural and
functional effects of constitutively activating mutations, Protein
Eng. 10 (1997) 501–510.
[15] http://tolweb.org/tree/phylogeney.html.
[16] G.F. Turner, O. Seehausen, M.E. Knight, C.J. Allender, R.L.
Robinson, How many species of cichlid fishes are there in the
African lakes? Mol. Evol. 10 (2001) 793–806.
[17] B. Rapoport, G.D. Chazenbalk, J.C. Jaume, S.M. McLachlan,
The thyrotropin (TSH)-releasing hormone receptor: interaction
with TSH and autoantibodies, Endocr. Rev. 19 (1998) 673–
716.
[18] S. Tavare�, C.M. Marshall, O. Will, C. Soligo, R.D. Martin, Using
the fossil record to estimate the age of the last common ancestor of
extant primates, Nature 416 (2002) 726–729.
[19] S.L. Page, C.H. Chui, M. Goodman, Molecular phylogeney of
Old World monkeys (Cercopithecidae) as inferred from c-globulin DNA sequences, Mol. Phylogenet. Evol. 13 (1999)
335–348.
V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290 289

[20] A.C. Van der Kuyl, C.L. Kuikek, J.T. Dekker, J. Goudsmit,
Phlyogeney of African monkeys based on mitochondrial 12S
rRNA sequences, J. Mol. Evol. 40 (1995) 173–180.
[21] E.E. Harris, T.R. Disotell, Nuclear trees and the phylogenetic
relationships of the Mangabeys (Primates: Paionini), Mol. Biol.
Evol. 15 (1998) 892–900.
[22] J.M. Scott, Genetic diversity and disease: opportunities and
challenge, Proc. Natl. Acad. USA 98 (2001) 14754–14756.
[23] Y. Nagayama, K.D. Kaufman, P. Seto, B. Rapoport, Molecular
cloning, sequencing and functional expression of the cDNA for
the human thyrotropin receptor, Biochem. Biophys. Res. Com-
mun. 165 (1989) 1184–1190.
[24] N.R. Farid, V. Kaczur, C. Balazs, The human thyrotropin
receptor is highly mutable: a review of gain-of-function mutations,
Euro. J. Endocrinol. 143 (2000) 25–30.
[25] C. Bratti_Elbaz, N. Ghunea, O. Lahuna, H. Loosfelt, C.
Pichon, E. Milgrom, Internalization and recycling pathways of
the thyrotropin receptor, Mol. Endocrinol. 13 (1999) 1751–
1765.
[26] Kishi, X. Liu, T. Hirakawa, D. Reczek, A. Bretscher, M. Ascoli,
Identification of two distinct structural motifs when added to the
C-terminal tail of the rat LR receptor, redirect the internalised
hormone–receptor complex from degradation to a recycling
pathway, Mol. Endocrinol. 15 (2001) 1624–1635.
[27] H. Biebermann, T. Schoneberg, A. Schultz, G. Krause, A. Guters,
G. Schultz, T. Gudermann, A conserved tyrosine residue (Y601)
in transmembrane domain 5 of the human thyrotropin receptor
serves as a molecular switch to determine G-protein coupling,
FASEB J. 12 (1998) 1461–1470.
[28] O. Karamanoglu Arseven, W.P. Wilkes, J.L. Jameson, P. Kopp,
Naturally occurring variants of the human thyrotropin (TSH)
receptor reveal that tyrosine 601 (Y601) plays a pivotal role in
constitutive basal activity and coupling to Gsa and Gq/11,
Thyroid 10 (1999) 1–3.
[29] R.S. Kumar, K. Ijiri, P. Swanson, A. Dittman, D. Alok, Y. Zoher,
J.M. Trant, Cloning and functional expression of a thyrotropin
receptor from the gonads of a vertebrate (bony fish): potential
thyroid-dependent role for thyrotropin in reproduction, Mol. Cell
Endocrinol. 167 (2000) 1–9.
[30] G. Smits, C. Govaerts, I. Nugourgh, L. Pardo, G. Vassart, S.
Costagliola, Lysine 183 and glutamic acid 157 of the TSH
receptor: two interacting residues with a key role in determining
specificity towards TSH and human CG, Mol. Endocrinol. 16
(2002) 722–735.
[31] Y. Nagayama, E. Nishihara, H. Namaba, S. Yamashita, M. Niwa,
Identification of the sites of aspargine-linked glycosylation of the
humanthyrotropin receptor and studies on their role on receptor
function and expression, J. Pharm. Exp. Therapeut. 295 (2000)
404–409.
[32] Y. Oba, T. Hirai, Y. Yoshiura, T. Kobayashi, Y. Nagahama,
Cloning, functional characterization, and functional expression of
thyrotropin receptors in the thyroid of amago salmon (Onorhyn-
chus rhodurus, Biochem. Biophys. Res. Commun. 276 (2000) 258–
263.
[33] Y. Mi, S.D. Shapiro, J.U. Baenziger, Regulation of lutropin
circulatory half-life by the mannose/N-acetylgalosamine-4-SO4
receptor is critical for implantation in vivo, J. Clin. Invest. 109
(2002) 269–276.
[34] P. Rodien, C. Bremont, M.L. Sanson, J. Parma, J. Van Sande, S.
Costagliola, J.P. Luton, G. Vassart, L. Duprez, Familial gesta-
tional hyperthyroidism caused by a mutant thyrotropin receptor
hypersensitive to human chorionic gonadotropin, N. Eng. J. Med.
339 (1998) 1823–1826.
[35] P. Kopp, S. Muirhead, N. Jourdian, W.X. Gu, J.L. Jameson, C.
Rodd, Congenital hyperthyroidism caused by a solitary toxic
adenoma harboring a novel somatic mutation (serine 281! iso-
leucine) in the extracellular domain of the thyrotropin receptor, J.
Clin. Invest. 100 (1997) 1634–1639.
[36] L. Duprez, J. Parma, S. Costagliola, J. Hermans, J. Van Sande,
J.E. Dumont, G. Vassart, Constitutive activation of the TSH
receptor by spontaneous mutations. Affecting the N-terminal
extracellular domain, FEBS Lett. 409 (1997) 469–474.
[37] S.C. Ho, J. Van Sande, A. Lefot, G. Vassart, S. Costagliola,
Effects of mutations involving the highly conserved S281 HCG
motif in the extracellular domain of the thyrotropin (TSH)
receptor in TSH binding and constitutive activity, Endocrinology
142 (2001) 2760–2767.
[38] V. Vlaeminck-Guillem, S.-C. Ho, P. Rodien, G. Vassart, S.
Costagliola, Activation of the cAMP pathway by the TSH
receptor involves switching of the ectodomain from a tethered
inverse agonist to an agonist, Mol. Endocrinol. 16 (2002) 736–746.
[39] G.D. Chazenbalk, P. Pichuria, C.-R. Chen, F. Latofe, A.P.
Johnstone, S.M. McLachlan, B. Rapoport, Thyroid stimulating
autoantibodies in Graves� disease preferentially recognize the freeA subunit, not the thyrotropin holoreceptor, J. Clin. Invest. 110
(2002) 209–217.
[40] R.A. Black, J.M. White, ADAMS: focus on the protease domain,
Curr. Opin. Cell Biol. 10 (1998) 654–659.
[41] M.N. Islam, N.R. Farid, Structure of the porcine thyrotropin
receptor: a 200 kiloDalton glycoprotein heterocomplex, Experi-
entia 41 (1985) 18–23.
[42] P.N. Graves, H. Vlase, Y. Bobvnikova, T.F. Davies, Multimeric
complex formation by the thyrotropin receptor in solubilized
thyroid membranes, Endocrinology 137 (1997) 3915–3920.
[43] R. Latif, P. Graves, T.F. Davies, Oligomerization of the human
thyrotropin receptor: florescent protein-tagged hTSHR reveals
post-translational complexes, J. Biol. Chem. 276 (2001) 45217–
45224.
[44] M.N. Islam, C.L. Hew, V. Kaczur, N.R. Farid, Cleavage of the
thyrotropin receptor, J. Endocrinol. Invest. 24 (2001) 833–835.
[45] P.R. Gouldson, C. Higgs, R.E. Smith, M.K. Dean, G.V. Gkoutos,
C.A. Reynolds, Dimerization and domain swapping in G-protein-
coupled receptors: a computational study, Neuropsychopharma-
cology 23 (2000) S60–S77.
[46] H.E. Hamm, How activated receptors couple to G proteins, Proc.
Natl. Acad. Sci. USA 98 (2001) 4819–4821.
[47] D. Fotiadis, Y. Liang, Filipek, D.A. Saperstein, A. Engel, K.
Palczewski, Rhodopsin dimers in native disc membranes, Nature
421 (2003) 127–128.
[48] M.K. Dean, C. Higgs, R.E. Smith, R.B. Bywater, C.R. Snell, P.D.
Scott, G.J. Upton, T.J. Howe, C.A. Reynolds, Dimerization of G-
coupled receptors, J. Med. Chem. 44 (2001) 4595–4616.
290 V. Kaczur et al. / Molecular Genetics and Metabolism 78 (2003) 275–290





























