Embed Size (px)
Citation preview

Exosomes bearing HLA-DR1 molecules needdendritic cells to ef®ciently stimulatespeci®c T cells
HeÂleÁne Vincent-Schneider1, Pamela Stumptner-Cuvelette1, Danielle Lankar1,Sabine Pain1, GracËa Raposo2, Philippe Benaroch1 and Christian Bonnerot1
1INSERM U520, Institut Curie, 26 rue d'Ulm, 75248 Paris Cedex 05, France2CNRS, UMR 144, Institut Curie, 12 rue Lhomond, 75005 Paris, France
Keywords: antigen presentation, dendritic cell, exosome, mast cell, MHC
Abstract
Exosomes are small vesicles (60±100 nm) secreted by various cell types upon the fusion ofendosomal compartments with the plasma membrane. Exosomes from antigen-presenting cells(APC), such as B lymphocytes and dendritic cells (DC), bear MHC class II molecules. In addition,the injection of DC-derived exosomes was reported to elicit potent T cell responses in vivo. Here,we analyzed the activation of speci®c T cells by MHC class II-bearing exosomes in vitro. The ratmast cell line, RBL-2H3, was engineered to express human class II molecules uniformly loadedwith an antigenic peptide [HLA-DR1±hemagglutinin (HA)]. These cells secreted exosomes bearingDR1 class II molecules upon stimulation by a calcium ionophore or IgE receptor cross-linking.Exosomes bearing DR1±HA(306±318) complexes activated HA/DR1-speci®c T cells only weakly,whereas the cross-linking of such exosomes to latex beads increased stimulation of speci®c Tcells. By contrast, the incubation of free exosomes with DC resulted in the highly ef®cientstimulation of speci®c T cells. Thus, exosomes bearing MHC class II complexes must be taken upby professional APC for ef®cient T cell activation.
Introduction
The endocytic pathway of antigen-presenting cells (APC) has
several key characteristics accounting for their unique ability to
elicit MHC class II-restricted T cell responses. Endosomes and
lysosomes from these cells are compartments with an import-
ant role in antigen presentation, in which antigenic peptides
associate with MHC class II molecules. Antigen-processing
compartments were ®rst described in human B cell lines as
multilaminar/multivesicular MHC class II compartments (MIIC)
(1,2). Resident endosomal proteins, such as CD63, and MHC
class II molecules accumulate in these compartments. In
addition, the fusion of MIIC with the plasma membrane leads to
the plasma membrane expression of peptide±MHC class II
complexes and to the secretion of small vesicles, called
exosomes, bearing MHC class II±peptide complexes.
Although the secretion of exosomes has been reported in
various hematopoietic cells including reticulocytes, platelets,
B lymphocytes, mast cells and dendritic cells (DC) (3±8), the
function of cell-derived exosomes remains unclear.The accumulation of MHC class II±peptide complexes on
the limiting membranes of exosomes derived from APC, such
as B cells or DC, led to evaluation of the role of exosomes
bearing MHC class II in the T cell response. High concentra-
tions of B-Epstein±Barr virus (EBV)-derived exosomes have
been shown to activate speci®c CD4 T cell clones directly (5).
Dendritic cell-derived exosomes have recently been reported
to accumulate MHC class II and I molecules, along with co-
stimulatory molecules (8,9). In addition, exosomes produced
by DC exposed to tumor-derived antigenic peptides have
been shown to induce a cytotoxic T lymphocyte-mediated
response leading to the regression of established tumors in
mice (8). Thus, exosomes bearing MHC molecules and
antigenic peptides induce potent T cell responses, although
the stimulation of speci®c T cells with peptide-pulsed
exosomes was consistently less ef®cient than the stimulation
of intact cells by incubation with peptides (5,8,10). This
suggests that exosomes do not interact directly with effector
cells. The precise mechanism of this phenomenon still remains
to be understood.Exosomes from platelets and mast cells are secreted upon
cell activation, suggesting that exosomes may be involved in
Correspondence to: C. Bonnerot; E-mail: [email protected]
Transmitting editor: I. Pecht Received 21 November 2001, accepted 22 March 2002
International Immunology, Vol. 14, No. 7, pp. 713±722 ã 2002 The Japanese Society for Immunology

homeostasis or in¯ammatory reactions. Furthermore, bonemarrow-derived mast cells (BMMC) and the mast cell line ratbasophilic leukemia (RBL)-2H3 mostly accumulate MHC classII molecules in small vesicles contained in secretory granules.The exocytosis of mast cell secretory granules may betriggered by antigen-induced aggregation of the high-af®nityreceptors (FceRI) for IgE (11) or exposure to a calciumionophore. The activation of BMMC expressing MHC class IIleads, as expected, to the release of exosomes bearing MHCclass II.
Homogeneous and reliable preparations of exosomes,bearing a well-de®ned MHC class II±peptide complex, are akey tool for investigating exosome function. We explored themechanisms of T cell activation by MHC class II+ exosomes,by expressing HLA-DR1 a and b chains alone or in combin-ation with an Ii chain mutant (IiHA), which carries a sequenceencoding the antigenic hemagglutinin (HA) peptide (12) inplace of the class II invariant chain peptide (CLIP) region (13),in the RBL cell line RBL-2H3. Co-expression of the IiHAconstruct facilitated the endogenous loading of most of theMHC class II molecules present in RBL DR1IiHA cells with theHA peptide. The accumulation of MHC class II in secretorygranules made it possible to induce the release of class II+
exosomes by cross-linking IgE receptors or adding calciumionophores. RBL DR1-derived exosomes loaded with HApeptide and RBL DR1IiHA-derived exosomes contain func-tional MHC class II complexes that stimulate speci®c CD4+ Tcells weakly. Coupling exosomes to carrier beads increasedthe ef®ciency of T cell stimulation. In contrast, exosomesstimulated DR1±HA-speci®c CD4+ T cells very ef®ciently ifthey were ®rst incubated with murine DC. Thus, exosomesmay need to be captured by DC to stimulate speci®c T cellsef®ciently.
Methods
Chemical reagents, antibodies and peptides
All chemicals used in this study were purchased from Sigma(St Louis, MO). RPMI 1640, FCS, PBS, penicillin, streptomycin,sodium pyruvate and L-glutamine were purchased from Gibco(Paisley, UK). We used the following mAb: L243 (22,23) whichrecognizes HLA-DR complexes devoid of the Ii chain, TuÈ36(24) directed against HLA-DR alone or associated with the Iichain, 1B5 (25) directed against HLA-DR a AD1 (26) an anti-rat CD63 mAb (a gift from Dr R. P. Siraganian, NIH, Bethesda,MA), LY1C6 an anti-rat lysosome-associated membraneprotein (Lamp1) mAb (a gift from Dr W. Hunziker, Epalinges,Switzerland) and 1A12 an anti-rat CD81 mAb (27) (a gift fromDr J. P. Kinet, Harvard University, Boston, MA). Secondaryantibodies included horseradish peroxidase-, FITC-, phycoer-ythrin (PE)- or Texas Red-coupled F(ab¢)2 fragments and wereobtained from Jackson ImmunoResearch (JacksonLaboratory, West Grove, PA). The HA306±318 peptide(PKYVKQNTLKLAT) (12) was synthesized by Synt:em (Nime,France).
Cells
RBL-2H3 (28) and RBL-derived cell lines, HOM2, THA1.7, andJurkat T cells were grown in RPMI 1640 supplemented with
10% FCS, 1% penicillin/streptomycin, 0.1% b-mercaptoetha-nol and 2% sodium pyruvate. The B-EBV cell line HOM2 ishomozygous for HLA-DR1 (29). THA1.7 are Jurkat T cellstransfected with constructs encoding the a and b chains of aDR1±HA(306±318)-speci®c TCR (30). Jurkat T cells werekindly provided by Dr O. Acuto (Institut Pasteur, Paris,France). RBL cells expressing I-AbIi and TH30 T cellhybridoma were previously described (14). The murineimmature DC line D1 (H-2B) and BMD8 (H-2D) were kindlyprovided by Dr P. Ricciardi-Castagnoli (Centre of Cellular andMolecular Pharmacology, Milan, Italy), and cultured as previ-ously described (15).
DR expression in RBL cells
The cDNAs used for stable transfection in RBL-2H3 cells wereinserted downstream from a Sra promoter (31) in expressionvectors carrying resistance genes for hygromycin B (NTH2),neomycin (NTNeo) and zeocin (NTZeo). The HLA-DR1 a chaincDNA (32) was subcloned into the NTNeo vector, the HLA-DR1 b chain cDNA (33) into the NTHygro vector and the IiHAcDNA (13) into the NTZeo vector. The resulting a, b and IiHAconstructs were then used to transfect the RBL-2H3 cell line aspreviously described (34). Brie¯y, cells were electroporated at260 V, 975 mF with 50 mg of linearized plasmid, using a BioRad(Marnes la Coquette, France) Gene Pulser II. Two days aftertransfection, the cells were transferred to selection medium.Surviving cells were cloned by limiting dilution. Clones wereanalyzed for surface MHC class II expression by cyto¯uoro-metry with a FACSCalibur ¯ow cytometer (Becton Dickinson,Mountain View, CA).
Flow cytometry analysis
Cells were washed in 3% FCS/0.1% NaN3 in PBS (FACSbuffer) prior to staining as previously described (14). Flowcytometry analysis was performed with a FACSCaliburmachine using CellQuest software (Becton Dickinson). Asexosomes are too small for FACScan analysis, we bound theexosomes to surfactant-free white aldehyde/sulfate latexbeads (3.8 mm diameter) and concentrated at 1.5 3 104
beads/ml (Interfacial; Dynamics, Portland, OR) (35). Weincubated 10 ml beads (corresponding to 1.5 3 105 beads)with 30 mg exosomes for 10±15 min and then for 1 h at roomtemperature in a ®nal volume of 1 ml PBS. Glycine (100 mM) inPBS was added to the beads, which were then incubated for30 min at room temperature to saturate the remaining bindingsites. Beads were washed twice with FACS buffer, andanalyzed with a FACSCalibur and CellQuest software.
Antigen-presentation assays
APC (5 3 104) were incubated with or without the HA peptide 5mM in the presence of 5 3 104 THA1.7 or Jurkat T cells/well for24 h. Each experiment was performed in duplicate. Weremoved 50 ml of supernatant and froze it at ±80°C for 1 h. Wemeasured IL-2 release from the T cell hybridoma by monitor-ing [3H]thymidine incorporation in the IL-2-dependent CTL-L2cell line. Cells were harvested after an additional 6 hincubation in the presence of 0.25 mCi [3H]thymidine/well.Each point corresponds to the mean of duplicate samples,which differed by <5%.
714 T cell stimulation by exosomes

For presentation assays with exosomes and exosomescross-linked to beads, 5 3 104 THA1.7 or Jurkat T cells/wellwere incubated with the corresponding exosome preparationand incubated with or without the HA peptide (0±10 or 50 mM).For exosome antigen-presentation assays in the presence ofD1 DC, exosomes derived from RBL DR1 or RBL DR1IiHAwere incubated with 5 3 104 THA1.7 cells in the presence orabsence of the HA peptide, with or without 5 3 104 D1 cells.For exosome antigen-presentation assays in the presence ofBMD8 DC, exosomes derived from RBL I-Ab Ii were incubatedwith 5 3 104 TH30 cells in the presence of the I-E a 52±68peptide (10 mM) with 5 3 104 BMD8 cells. After 24 h, IL-2release was assayed as described above (36).
Exosome puri®cation
Exosomes were prepared from supernatants of RBL-2H3 cellsand derived cell lines, generally degranulated by incubationwith a calcium ionophore (1 mM ionomycin or A23187) for 30min at 37°C (37) or by cross-linking of FceRI as previouslydescribed (38). Brie¯y, cells were incubated with anti-DNP IgEfor 20 h at 37°C and degranulation was then triggered byadding 10 mg/ml DNP-BSA and incubating for various periodsof time. Exosomes were isolated as previously described (5).Brie¯y, cell supernatants were successively centrifuged at 300g for 10 min, 1200 g for 20 min, 3000 g for 30 min and ®nally70,000 g for 1 h. The exosome pellet was washed in PBS,centrifuged at 70,000 g for 1 h and resuspended in PBS andprotein concentration determined by Bradford assay (BioRad).
For the immunoisolation of exosomes, 5 ml latex beads(corresponding to 0.75 3 105 beads) were incubated over-night with 15 mg of puri®ed anti-CD63 mAb at 4°C. Bindingsites were saturated by incubation in 100 mM glycine in PBSfor 30 min at room temperature and in the beads were thenwashed in FACS buffer. We incubated 10 ml beads with 30 mgpuri®ed exosomes (or FCS as negative control) and carriedout labeling for FACS analysis as described above forexosomes directly coupled to beads.
Sucrose gradient
RBL DR1 exosomes (100 mg) were resuspended in 2 ml 2.5 Msucrose, 20 mM HEPES/NaOH, pH 7.2. A linear sucrosegradient (2.0±2.5 M sucrose, 20 mM HEPES/NaOH, pH 7.2)was layered on top of the exosome suspension in a SW41tube (Beckman Instruments, Gagny, France). Gradients werecentrifuged for 15 h at 100,000 g and a 1-ml fraction wascollected from the top of the tube. Densities were evaluatedusing a refractometer. Membranes were collected from thecollected fractions after centrifugation at 70,000 g for 1 h at4°C in TLA 100.4 tubes (Beckman Instruments). Exosomepellets were solubilized in reducing Laemmli buffer andheated at 95°C for 5 min before analysis by SDS±PAGE andWestern blotting.
Western blotting
Cells were lysed in 0.5% Triton, 300 mM NaCl, 50 mM Tris, pH7.4 in PBS supplemented with 10 mg/ml each of leupeptin,chemostatin, aprotinin, pepstatin, N-ethyl maleinide and 1 mMPMSF. Cell or exosome lysates were diluted in reducingLaemmli buffer and heated at 95°C for 5 min before analysison SDS±12% polyacrylamide gels. Proteins were transferred
to a PVDF membranes (Millipore, Bedford, MA). The mem-branes were incubated in blocking solution, then with IB5 [ananti-DR mAb (25)] and ®nally with horseradish peroxidase-labeled donkey anti-mouse IgG secondary antibody (JacksonLaboratory). Antibody binding was detected by chemi-luminescence, using an ECL kit (Boehringer Mannheim,Mannheim, Germany).
Immunoelectron microscopy
RBL DR1 cells (degranulated or not) were ®xed by incubationin a mixture of 2% paraformaldehyde in phosphate buffer 0.2M, pH 7.4 and 0.125% glutaraldehyde for 2 h at roomtemperature. Fixed cells were processed for ultrathin section-ing and immunolabeling as previously described (6). Cellswere washed with phosphate buffer 0.2 M, pH 7.4 andphosphate buffer 0.2 M, pH 7.4/50 mM glycine, and were thenembedded in 7.5 % gelatin. Small blocks were in®ltrated with2.3 M sucrose at 4°C for 2 h and then frozen in liquid nitrogen.Ultrathin cryosections prepared with a Leica ultracut FCS(Wien, Austria) were retrieved in a mixture of 2% methylcellu-lose and 2.3 M sucrose (v/v), and indirectly immunogoldlabeled by incubation with the rat monoclonal anti-HLA-DR1antibody L243 followed by a rabbit anti-mouse antibody(Dako, Copenhagen, Denmark). Antibodies were detectedwith Protein A coupled to 10-nm gold particles (purchasedfrom Dr J. W. Slot, Department of Cell Biology, UtrechtUniversity, The Netherlands). The sections were contraststained, embedded in a mixture of methylcellulose and uranylacetate, and viewed under a CM120 Twin Phillips electronmicroscope (Eindhoven, The Netherlands). For electronmicroscopy of the isolated exosomes, the pellets from the70,000 g centrifugation were placed on Formvar carbon-coated electron microscopy grids, ®xed as above, immuno-labeled and stained using the method described for ultrathincryosections. Exosomes from RBL I-AbIi cells were labeled insimilar conditions using M5/114 mAb (14)
Results
RBL cells expressing HLA-DR1 chains together with theIiHA construct are ef®cient APC
To generate mast cell lines expressing MHC class II±peptidecomplexes, we transfected the RBL cell line RBL-2H3 withcDNAs encoding HLA-DR1 a and b chains with or without apreviously described IiHA construct (Fig. 1A). We failed todetect any endogenous MHC class II in our RBL-2H3 cloneusing the anti-rat MHC class II, OX6 (data not shown). Thecore CLIP sequence of this human Ii-derived construct hasbeen replaced by the sequence encoding the antigenicHA306±318 peptide of in¯uenza virus HA (13). After selectionand cloning, one clone for each combination was selected onlevels of surface MHC class II expression. RBL DR1 cells, RBLDR1IiHA cells and HOM2 B-EBV cells displayed similar levelsof surface expression of MHC class II molecules, as deter-mined by cyto¯uorimetry (Fig. 1B).
We then tested the ability of these cells to stimulate Jurkat Tcells expressing or not a speci®c TCR recognizing HLA-DR1±HA306±318, known as THA1.7. The cells were incubated withvarious concentrations of HA peptide and IL-2 secretion by T
T cell stimulation by exosomes 715
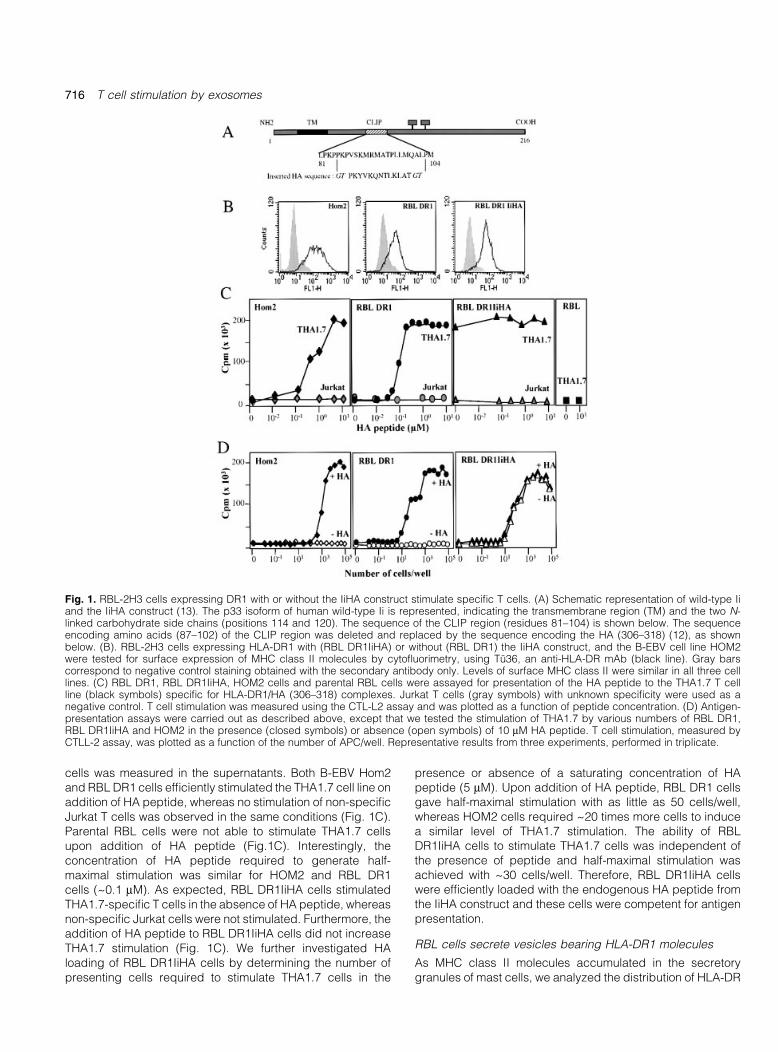
cells was measured in the supernatants. Both B-EBV Hom2and RBL DR1 cells ef®ciently stimulated the THA1.7 cell line onaddition of HA peptide, whereas no stimulation of non-speci®cJurkat T cells was observed in the same conditions (Fig. 1C).Parental RBL cells were not able to stimulate THA1.7 cellsupon addition of HA peptide (Fig.1C). Interestingly, theconcentration of HA peptide required to generate half-maximal stimulation was similar for HOM2 and RBL DR1cells (~0.1 mM). As expected, RBL DR1IiHA cells stimulatedTHA1.7-speci®c T cells in the absence of HA peptide, whereasnon-speci®c Jurkat cells were not stimulated. Furthermore, theaddition of HA peptide to RBL DR1IiHA cells did not increaseTHA1.7 stimulation (Fig. 1C). We further investigated HAloading of RBL DR1IiHA cells by determining the number ofpresenting cells required to stimulate THA1.7 cells in the
presence or absence of a saturating concentration of HApeptide (5 mM). Upon addition of HA peptide, RBL DR1 cellsgave half-maximal stimulation with as little as 50 cells/well,whereas HOM2 cells required ~20 times more cells to inducea similar level of THA1.7 stimulation. The ability of RBLDR1IiHA cells to stimulate THA1.7 cells was independent ofthe presence of peptide and half-maximal stimulation wasachieved with ~30 cells/well. Therefore, RBL DR1IiHA cellswere ef®ciently loaded with the endogenous HA peptide fromthe IiHA construct and these cells were competent for antigenpresentation.
RBL cells secrete vesicles bearing HLA-DR1 molecules
As MHC class II molecules accumulated in the secretorygranules of mast cells, we analyzed the distribution of HLA-DR
Fig. 1. RBL-2H3 cells expressing DR1 with or without the IiHA construct stimulate speci®c T cells. (A) Schematic representation of wild-type Iiand the IiHA construct (13). The p33 isoform of human wild-type Ii is represented, indicating the transmembrane region (TM) and the two N-linked carbohydrate side chains (positions 114 and 120). The sequence of the CLIP region (residues 81±104) is shown below. The sequenceencoding amino acids (87±102) of the CLIP region was deleted and replaced by the sequence encoding the HA (306±318) (12), as shownbelow. (B). RBL-2H3 cells expressing HLA-DR1 with (RBL DR1IiHA) or without (RBL DR1) the IiHA construct, and the B-EBV cell line HOM2were tested for surface expression of MHC class II molecules by cyto¯uorimetry, using TuÈ36, an anti-HLA-DR mAb (black line). Gray barscorrespond to negative control staining obtained with the secondary antibody only. Levels of surface MHC class II were similar in all three celllines. (C) RBL DR1, RBL DR1IiHA, HOM2 cells and parental RBL cells were assayed for presentation of the HA peptide to the THA1.7 T cellline (black symbols) speci®c for HLA-DR1/HA (306±318) complexes. Jurkat T cells (gray symbols) with unknown speci®city were used as anegative control. T cell stimulation was measured using the CTL-L2 assay and was plotted as a function of peptide concentration. (D) Antigen-presentation assays were carried out as described above, except that we tested the stimulation of THA1.7 by various numbers of RBL DR1,RBL DR1IiHA and HOM2 in the presence (closed symbols) or absence (open symbols) of 10 mM HA peptide. T cell stimulation, measured byCTLL-2 assay, was plotted as a function of the number of APC/well. Representative results from three experiments, performed in triplicate.
716 T cell stimulation by exosomes
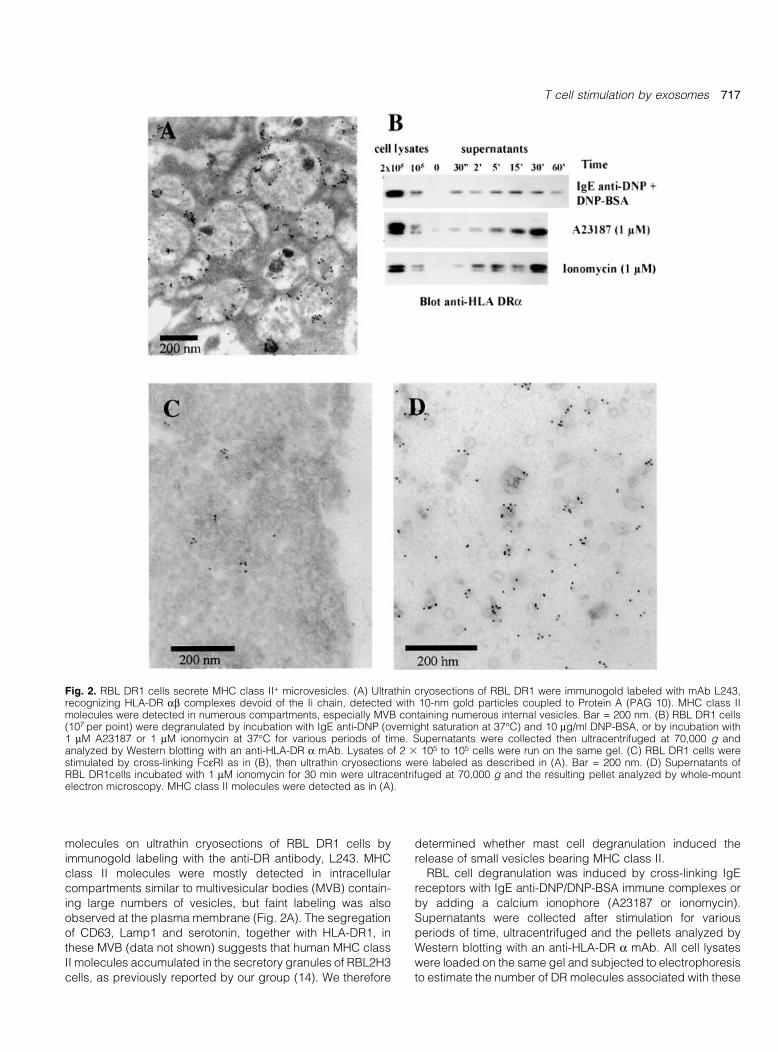
molecules on ultrathin cryosections of RBL DR1 cells by
immunogold labeling with the anti-DR antibody, L243. MHC
class II molecules were mostly detected in intracellular
compartments similar to multivesicular bodies (MVB) contain-
ing large numbers of vesicles, but faint labeling was also
observed at the plasma membrane (Fig. 2A). The segregation
of CD63, Lamp1 and serotonin, together with HLA-DR1, in
these MVB (data not shown) suggests that human MHC class
II molecules accumulated in the secretory granules of RBL2H3
cells, as previously reported by our group (14). We therefore
determined whether mast cell degranulation induced the
release of small vesicles bearing MHC class II.RBL cell degranulation was induced by cross-linking IgE
receptors with IgE anti-DNP/DNP-BSA immune complexes or
by adding a calcium ionophore (A23187 or ionomycin).
Supernatants were collected after stimulation for various
periods of time, ultracentrifuged and the pellets analyzed by
Western blotting with an anti-HLA-DR a mAb. All cell lysates
were loaded on the same gel and subjected to electrophoresis
to estimate the number of DR molecules associated with these
Fig. 2. RBL DR1 cells secrete MHC class II+ microvesicles. (A) Ultrathin cryosections of RBL DR1 were immunogold labeled with mAb L243,recognizing HLA-DR ab complexes devoid of the Ii chain, detected with 10-nm gold particles coupled to Protein A (PAG 10). MHC class IImolecules were detected in numerous compartments, especially MVB containing numerous internal vesicles. Bar = 200 nm. (B) RBL DR1 cells(107 per point) were degranulated by incubation with IgE anti-DNP (overnight saturation at 37°C) and 10 mg/ml DNP-BSA, or by incubation with1 mM A23187 or 1 mM ionomycin at 37°C for various periods of time. Supernatants were collected then ultracentrifuged at 70,000 g andanalyzed by Western blotting with an anti-HLA-DR a mAb. Lysates of 2 3 105 to 105 cells were run on the same gel. (C) RBL DR1 cells werestimulated by cross-linking FceRI as in (B), then ultrathin cryosections were labeled as described in (A). Bar = 200 nm. (D) Supernatants ofRBL DR1cells incubated with 1 mM ionomycin for 30 min were ultracentrifuged at 70,000 g and the resulting pellet analyzed by whole-mountelectron microscopy. MHC class II molecules were detected as in (A).
T cell stimulation by exosomes 717

vesicles. The amount of DR1 a chain present in the super-
natant gradually increased over time, reaching a maximum at30 min (Fig. 2B). The addition of either of the calcium
ionophores induced levels of DR1 secretion twice as high as
those induced by IgE receptor cross-linking. Comparison of
the DR1 a chain signals observed for vesicle preparations
(obtained from 107 cells) with those obtained from cells lysates
(1±2 3 105 cells) indicated that almost 2% of total MHC class II
molecules were secreted upon mast cell degranulation
(Fig. 2B).To con®rm that MHC class II-containing vesicles were
secreted, we prepared ultrathin cryosections of RBL DR1 cells
following IgE receptor cross-linking and analyzed them by
immunogold labeling with the anti-DR antibody, L243.
Exocytosis pro®les with DR1-labeled vesicles were detected
(Fig. 2C). We puri®ed vesicles from the supernatant of
stimulated RBL DR1 cells by ultracentrifugation and analyzed
them directly by whole-mount electron microscopy.
Heterogeneous populations of vesicles 60±80 nm in diameter
were detected (Fig. 2D). Most had single membranes, but
those with double membranes had denser lumina and
frequently displayed positive HLA-DR labeling (Fig. 2D).
Contaminations of vesicles preparations by endoplasmic
reticulum membranes were excluded by the lack of any
endoplasmic reticulum protein (calnexin) in exosomes pellets
(data not shown).Thus, RBL DR1 cells can secrete large numbers of small
vesicles carrying HLA-DR1 molecules, which have morpho-
logical features in common with previously described exo-
somes.
Characterization of the secreted vesicles
To determine whether the vesicles secreted by RBL DR1 cells
were indeed exosomes, vesicles obtained by ultracentrifuga-
tion were subjected to sucrose gradient sedimentation and the
MHC class II content of each fraction was analyzed by western
blotting. Most of the MHC class II molecules were found in
fractions with a sucrose density of 1.16±1.21 g/ml (Fig. 3A).
Thus, the vesicles bearing MHC class II sedimented at a
slightly higher density than that previously reported for
exosomes (1.135 g/ml). As these vesicles originated from
MVB, as described for exosomes, we analyzed their endocy-
tic/lysosomal pathway protein content. The vesicles were too
small for direct analysis by cyto¯uorimetry and most of the
antibodies against rat lysosomal proteins do not work in
Western blotting experiments. We therefore analyzed the
protein composition of the vesicles by coupling the vesicles to
3.8-mm latex beads and immunolabeling with FITC-conjugated
speci®c antibodies. FACScan analysis of the vesicles
attached to the beads revealed that they contained large
amounts of CD63, CD81 and MHC class II, but almost no
Lamp1 (Fig. 3B). A key feature of exosomes is the association
of MHC class II molecules with tetraspanin on the same
vesicle. We investigated whether the vesicles secreted by RBL
cells contained CD63 and DR1, by coating beads with the anti-
CD63 mAb AD1 and incubating them with vesicles prepared
from RBL DR1 cells. We then washed the beads and assessed
the amount of DR1 associated with the beads using a PE-
conjugated anti-DR mAb. Signi®cant speci®c labeling was
observed, suggesting that a population of vesicles carried
both DR and CD63 (Fig. 3C).Upon stimulation, RBL cells secrete vesicles of 60±80 nm in
diameter, which ¯oat on sucrose gradients, and bear CD63,
HLA-DR and CD81 molecules. We therefore concluded that
these vesicles had features similar to those previously
described for exosomes in B cells, DC and mast cells.
Fig. 3. Microvesicles secreted by RBL DR1 correspond toexosomes. (A) Membranes isolated by ultracentrifugation fromsupernatants of RBL DR1 cells were loaded onto a linear sucrosegradient. Membranes were collected from the gradient fractions andanalyzed by Western blotting for HLA-DR expression with the mAb1B5 (upper panel). Results were quanti®ed by analyzing scanned®lms with NIH imaging software. The densities of the variousgradient fractions are indicated at the bottom. The lower panelshows the estimated amount of HLA-DR a chain as a function ofsucrose density. (B) We coupled 30 mg of microvesicles or of BSA to10 ml latex beads and tested for surface expression of CD63 (AD1mAb), Lamp1 (LY1C6 mAb), CD81 (1A12 mAb), and mature HLA-DR1 (L243 mAb) and total HLA-DR1 (TuÈ36 mAb). (C) Latex beadspre-incubated with the anti-CD63 mAb AD1 were used toimmunoisolate microvesicles secreted from RBL DR1 cells. Beadswere further analyzed by ¯ow cytometry for HLA-DR surfaceexpression using PE-coupled L243 and TuÈ36 mAb. In (B) and (C),gray histograms correspond to the same beads incubated with PE-coupled isotypes controls.
718 T cell stimulation by exosomes
RBL exosomes carry functionally competent DR molecules
As we were able to produce homogeneous and reliable
preparations of exosomes bearing MHC class II molecules
from RBL DR1 and RBL DR1IiHA cells, we used these
exosomes to investigate the activation of speci®c T cells
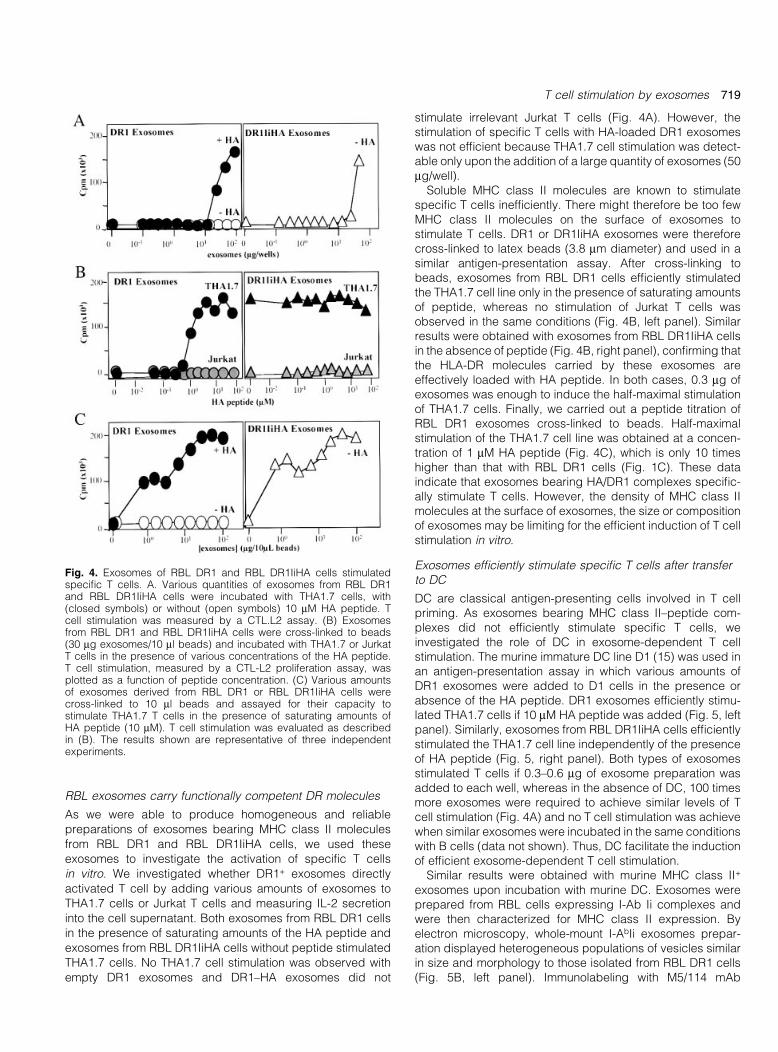
in vitro. We investigated whether DR1+ exosomes directly
activated T cell by adding various amounts of exosomes to
THA1.7 cells or Jurkat T cells and measuring IL-2 secretion
into the cell supernatant. Both exosomes from RBL DR1 cells
in the presence of saturating amounts of the HA peptide and
exosomes from RBL DR1IiHA cells without peptide stimulated
THA1.7 cells. No THA1.7 cell stimulation was observed with
empty DR1 exosomes and DR1±HA exosomes did not
stimulate irrelevant Jurkat T cells (Fig. 4A). However, thestimulation of speci®c T cells with HA-loaded DR1 exosomeswas not ef®cient because THA1.7 cell stimulation was detect-able only upon the addition of a large quantity of exosomes (50mg/well).
Soluble MHC class II molecules are known to stimulatespeci®c T cells inef®ciently. There might therefore be too fewMHC class II molecules on the surface of exosomes tostimulate T cells. DR1 or DR1IiHA exosomes were thereforecross-linked to latex beads (3.8 mm diameter) and used in asimilar antigen-presentation assay. After cross-linking tobeads, exosomes from RBL DR1 cells ef®ciently stimulatedthe THA1.7 cell line only in the presence of saturating amountsof peptide, whereas no stimulation of Jurkat T cells wasobserved in the same conditions (Fig. 4B, left panel). Similarresults were obtained with exosomes from RBL DR1IiHA cellsin the absence of peptide (Fig. 4B, right panel), con®rming thatthe HLA-DR molecules carried by these exosomes areeffectively loaded with HA peptide. In both cases, 0.3 mg ofexosomes was enough to induce the half-maximal stimulationof THA1.7 cells. Finally, we carried out a peptide titration ofRBL DR1 exosomes cross-linked to beads. Half-maximalstimulation of the THA1.7 cell line was obtained at a concen-tration of 1 mM HA peptide (Fig. 4C), which is only 10 timeshigher than that with RBL DR1 cells (Fig. 1C). These dataindicate that exosomes bearing HA/DR1 complexes speci®c-ally stimulate T cells. However, the density of MHC class IImolecules at the surface of exosomes, the size or compositionof exosomes may be limiting for the ef®cient induction of T cellstimulation in vitro.
Exosomes ef®ciently stimulate speci®c T cells after transferto DC
DC are classical antigen-presenting cells involved in T cellpriming. As exosomes bearing MHC class II±peptide com-plexes did not ef®ciently stimulate speci®c T cells, weinvestigated the role of DC in exosome-dependent T cellstimulation. The murine immature DC line D1 (15) was used inan antigen-presentation assay in which various amounts ofDR1 exosomes were added to D1 cells in the presence orabsence of the HA peptide. DR1 exosomes ef®ciently stimu-lated THA1.7 cells if 10 mM HA peptide was added (Fig. 5, leftpanel). Similarly, exosomes from RBL DR1IiHA cells ef®cientlystimulated the THA1.7 cell line independently of the presenceof HA peptide (Fig. 5, right panel). Both types of exosomesstimulated T cells if 0.3±0.6 mg of exosome preparation wasadded to each well, whereas in the absence of DC, 100 timesmore exosomes were required to achieve similar levels of Tcell stimulation (Fig. 4A) and no T cell stimulation was achievewhen similar exosomes were incubated in the same conditionswith B cells (data not shown). Thus, DC facilitate the inductionof ef®cient exosome-dependent T cell stimulation.
Similar results were obtained with murine MHC class II+
exosomes upon incubation with murine DC. Exosomes wereprepared from RBL cells expressing I-Ab Ii complexes andwere then characterized for MHC class II expression. Byelectron microscopy, whole-mount I-AbIi exosomes prepar-ation displayed heterogeneous populations of vesicles similarin size and morphology to those isolated from RBL DR1 cells(Fig. 5B, left panel). Immunolabeling with M5/114 mAb
Fig. 4. Exosomes of RBL DR1 and RBL DR1IiHA cells stimulatedspeci®c T cells. A. Various quantities of exosomes from RBL DR1and RBL DR1IiHA cells were incubated with THA1.7 cells, with(closed symbols) or without (open symbols) 10 mM HA peptide. Tcell stimulation was measured by a CTL.L2 assay. (B) Exosomesfrom RBL DR1 and RBL DR1IiHA cells were cross-linked to beads(30 mg exosomes/10 ml beads) and incubated with THA1.7 or JurkatT cells in the presence of various concentrations of the HA peptide.T cell stimulation, measured by a CTL-L2 proliferation assay, wasplotted as a function of peptide concentration. (C) Various amountsof exosomes derived from RBL DR1 or RBL DR1IiHA cells werecross-linked to 10 ml beads and assayed for their capacity tostimulate THA1.7 T cells in the presence of saturating amounts ofHA peptide (10 mM). T cell stimulation was evaluated as describedin (B). The results shown are representative of three independentexperiments.
T cell stimulation by exosomes 719
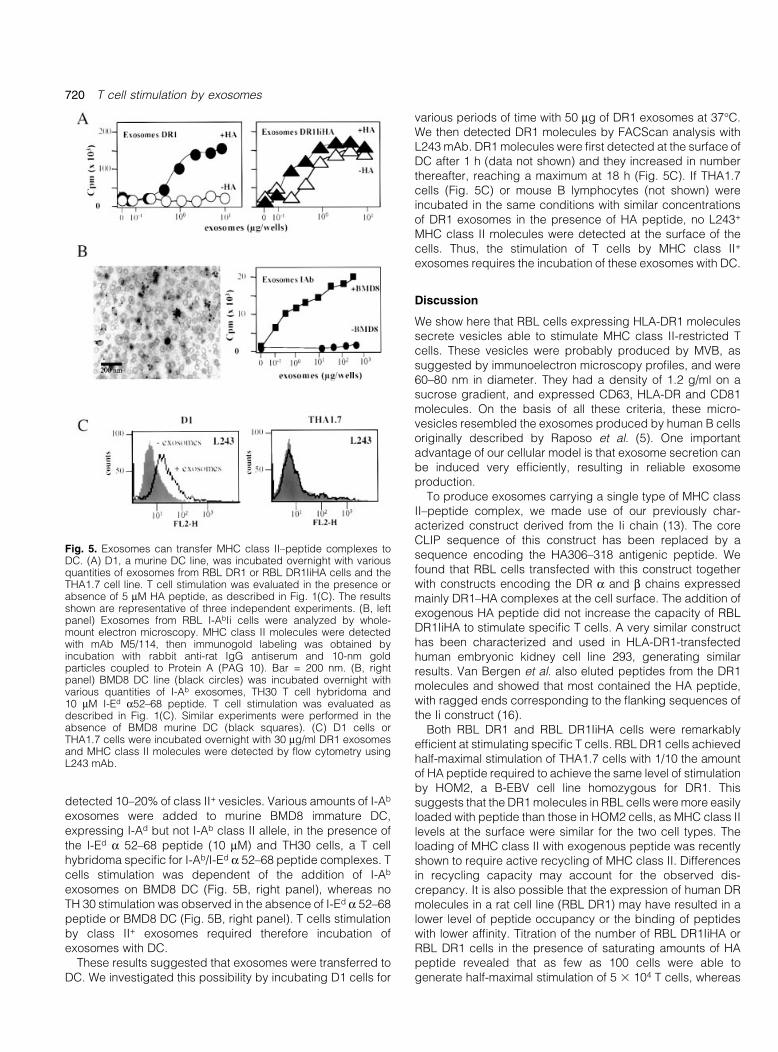
detected 10±20% of class II+ vesicles. Various amounts of I-Ab
exosomes were added to murine BMD8 immature DC,expressing I-Ad but not I-Ab class II allele, in the presence ofthe I-Ed a 52±68 peptide (10 mM) and TH30 cells, a T cellhybridoma speci®c for I-Ab/I-Ed a 52±68 peptide complexes. Tcells stimulation was dependent of the addition of I-Ab
exosomes on BMD8 DC (Fig. 5B, right panel), whereas noTH 30 stimulation was observed in the absence of I-Ed a 52±68peptide or BMD8 DC (Fig. 5B, right panel). T cells stimulationby class II+ exosomes required therefore incubation ofexosomes with DC.
These results suggested that exosomes were transferred toDC. We investigated this possibility by incubating D1 cells for
various periods of time with 50 mg of DR1 exosomes at 37°C.We then detected DR1 molecules by FACScan analysis withL243 mAb. DR1 molecules were ®rst detected at the surface ofDC after 1 h (data not shown) and they increased in numberthereafter, reaching a maximum at 18 h (Fig. 5C). If THA1.7cells (Fig. 5C) or mouse B lymphocytes (not shown) wereincubated in the same conditions with similar concentrationsof DR1 exosomes in the presence of HA peptide, no L243+
MHC class II molecules were detected at the surface of thecells. Thus, the stimulation of T cells by MHC class II+
exosomes requires the incubation of these exosomes with DC.
Discussion
We show here that RBL cells expressing HLA-DR1 moleculessecrete vesicles able to stimulate MHC class II-restricted Tcells. These vesicles were probably produced by MVB, assuggested by immunoelectron microscopy pro®les, and were60±80 nm in diameter. They had a density of 1.2 g/ml on asucrose gradient, and expressed CD63, HLA-DR and CD81molecules. On the basis of all these criteria, these micro-vesicles resembled the exosomes produced by human B cellsoriginally described by Raposo et al. (5). One importantadvantage of our cellular model is that exosome secretion canbe induced very ef®ciently, resulting in reliable exosomeproduction.
To produce exosomes carrying a single type of MHC classII±peptide complex, we made use of our previously char-acterized construct derived from the Ii chain (13). The coreCLIP sequence of this construct has been replaced by asequence encoding the HA306±318 antigenic peptide. Wefound that RBL cells transfected with this construct togetherwith constructs encoding the DR a and b chains expressedmainly DR1±HA complexes at the cell surface. The addition ofexogenous HA peptide did not increase the capacity of RBLDR1IiHA to stimulate speci®c T cells. A very similar constructhas been characterized and used in HLA-DR1-transfectedhuman embryonic kidney cell line 293, generating similarresults. Van Bergen et al. also eluted peptides from the DR1molecules and showed that most contained the HA peptide,with ragged ends corresponding to the ¯anking sequences ofthe Ii construct (16).
Both RBL DR1 and RBL DR1IiHA cells were remarkablyef®cient at stimulating speci®c T cells. RBL DR1 cells achievedhalf-maximal stimulation of THA1.7 cells with 1/10 the amountof HA peptide required to achieve the same level of stimulationby HOM2, a B-EBV cell line homozygous for DR1. Thissuggests that the DR1 molecules in RBL cells were more easilyloaded with peptide than those in HOM2 cells, as MHC class IIlevels at the surface were similar for the two cell types. Theloading of MHC class II with exogenous peptide was recentlyshown to require active recycling of MHC class II. Differencesin recycling capacity may account for the observed dis-crepancy. It is also possible that the expression of human DRmolecules in a rat cell line (RBL DR1) may have resulted in alower level of peptide occupancy or the binding of peptideswith lower af®nity. Titration of the number of RBL DR1IiHA orRBL DR1 cells in the presence of saturating amounts of HApeptide revealed that as few as 100 cells were able togenerate half-maximal stimulation of 5 3 104 T cells, whereas
Fig. 5. Exosomes can transfer MHC class II±peptide complexes toDC. (A) D1, a murine DC line, was incubated overnight with variousquantities of exosomes from RBL DR1 or RBL DR1IiHA cells and theTHA1.7 cell line. T cell stimulation was evaluated in the presence orabsence of 5 mM HA peptide, as described in Fig. 1(C). The resultsshown are representative of three independent experiments. (B, leftpanel) Exosomes from RBL I-AbIi cells were analyzed by whole-mount electron microscopy. MHC class II molecules were detectedwith mAb M5/114, then immunogold labeling was obtained byincubation with rabbit anti-rat IgG antiserum and 10-nm goldparticles coupled to Protein A (PAG 10). Bar = 200 nm. (B, rightpanel) BMD8 DC line (black circles) was incubated overnight withvarious quantities of I-Ab exosomes, TH30 T cell hybridoma and10 mM I-Ed a52±68 peptide. T cell stimulation was evaluated asdescribed in Fig. 1(C). Similar experiments were performed in theabsence of BMD8 murine DC (black squares). (C) D1 cells orTHA1.7 cells were incubated overnight with 30 mg/ml DR1 exosomesand MHC class II molecules were detected by ¯ow cytometry usingL243 mAb.
720 T cell stimulation by exosomes

1000 HOM2 cells were required to generate a similarresponse. This provides support for the idea that RBL DR1cells are easily loaded with exogenous peptide. RBL DR1IiHAand RBL DR1 generated similar maximal responses in thepresence of HA peptide, supporting the notion that, in bothsituations, the majority of the DR1 molecules were loaded withHA peptide. We previously showed that murine MHC class IImolecules (I-Ab) may be expressed as functional molecules inRBL cells (14). Therefore, RBL cells are a useful model forexpressing MHC class II complexes loaded with a givenpeptide, supplied exogenously or introduced via an Ii chain-derived construct.
We found that MHC class II molecules were present onexosomes secreted by RBL DR1 cells exposed to appropriatestimuli. We studied their capacity to stimulate T cells andobserved that the threshold at which T cells respondedincreased by a factor of 2 log, for a given quantity ofexosomes, if the exosomes were chemically cross-linked tobeads. This suggests that there are too few MHC class IImolecules per exosome to create an immunological synapseon the target T cell. Indeed, it has been established that aminimum of ~10 MHC class II molecules are needed foref®cient reorganization of the TCR at the T cell surface, whichin turn leads to a signal transduction cascade activating the Tcell (17,18).
Interestingly, immature mouse DC, like beads, increasedthe capacity of exosomes to stimulate T cells by a factor of 2log. Other cell types, such as B cells and mast cells, had nosuch effect (not shown). The transfer of vesicles from APC to Tcells following speci®c interaction has been reported before(19). It is unclear whether exosomes play a physiological rolein vivo. However, immunization with exosomes proved to bevery ef®cient in a mouse in vivo tumor model (8). Thesigni®cant enhancement by DC of the stimulatory capacity ofMHC class II molecules carried by exosomes in vitro suggeststhat DC may have a similar function in vivo. They may capturecirculating exosomes and present them to T cells. Indeed,follicular DC (FDC) have been demonstrated to bind MHCclass II+ microvesicles, which closely resemble exosomes inprotein content and morphology, speci®cally to their surface(20). Such a role would also be compatible with the reportedability of FDC to trap circulating viral particles, similar in size toexosomes, at their surface (21). It is unclear whether DC fusewith exosomes and process them, and the possibility of suchpresentation in vivo for the maintenance of peripheral toler-ance and induction of speci®c immune responses is alsounknown. However, our model should provide a useful tool foraddressing these important issues in future research.
Acknowledgements
We would like to thank D. Tenza for technical assistance with electronmicroscopy, S. Morchoisne for the subcloning of THA1.7 cells and P.Veron for advice on FACScan analysis with latex beads. We would alsolike to thank Dr C. Hivroz and N. Blanchard for advice concerningJurkat T cells. Special thanks are addressed to Dr C. Thery and Dr. S.Amigorena for stimulating discussion and helpful comments onexosomes. This work was supported by grants from Institut Nationalde la Sante et de la Recherche MeÂdicale, from Institut Curie, fromAPCells and from the comite de Paris de la Ligue Nationale contre leCancer. H. V.-S. holds a fellowship from the MinisteÁre des UniversiteÂs
et de la Recherche. P. S.-C. was supported by a grant from theAgence Nationale de la Recherche contre le SIDA.
Abbreviations
APC antigen-presenting cellBMMC bone marrow-derived mast cellCLIP class II invariant chain peptideDC dendritic cellEBV Epstein±Barr virusFDC follicular dendritic cellHA hemagglutininLamp lysosome-associated membrane proteinMIIC MHC class II compartmentMVB multivesicular bodyPE phycoerythrinRBL rat basophilic leukemia
References
1 Peters, P. J., Neefjes, J. J., Oorschot, V., Ploegh, H. L. and Geuze,H. J. 1991. Segregation of MHC class II molecules from MHCclass I molecules in the Golgi complex for transport to lysosomalcompartments. Nature 349:669.
2 Peters, P. J., Raposo, G., Neefjes, J. J., Oorschot, V.,Leijendekker, R. L., Geuze, H. J. and Ploegh, H. L. 1995. Majorhistocompatibility complex class II compartments in human Blymphoblastoid cells are distinct from early endosomes. J. Exp.Med. 182:325.
3 Denzer, K., Kleijmeer, M. J., Heijnen, H. F., Stoorvogel, W. andGeuze, H. J. 2000. Exosome: from internal vesicle of themultivesicular body to intercellular signaling device. J. Cell Sci.113:3365.
4 Johnstone, R. M., Adam, M., Hammond, J. R., Orr, L. and Turbide,C. 1987. Vesicle formation during reticulocyte maturation.Association of plasma membrane activities with releasedvesicles (exosomes). J. Biol. Chem. 262:9412.
5 Raposo, G., Nijman, H. W., Stoorvogel, W., Leijendekker, R.,Harding, C. V., Melief, C. J. M. and Geuze, H. J. 1996. Blymphocytes secrete antigen-presenting vesicles. J. Exp. Med.183:1.
6 Raposo, G., Tenza, D., Mecheri, S., Perronet, R., Bonnerot, C. andDesaymard, C. 1997. Accumulation of MHC class II molecules inmast cell secretory granules and their release upon degranulation.Mol. Biol. Cell 8:2631.
7 Raposo, G., Vidal, M. and Geuze, H. J. 1997. Secretory lysosomesand the production of exosomes. In Kuchler, A. R. K. and Holland,B., eds, Unusual and Leaderless Secretory Pathways: FromBacteria to Man, p. 161. Landes, Austin, TX.
8 Zitvogel, L., Regnault, A., Lozier, A., Wolfers, J., Flament, C.,Tenza, D., Ricciardi-Castagnoli, P., Raposo, G. and Amigorena, S.1998. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat. Med. 4:594.
9 Thery, C., Regnault, A., Garin, J., Wolfers, J., Zitvogel, L.,Ricciardi-Castagnoli, P., Raposo, G. and Amigorena, S. 1999.Molecular characterization of dendritic cell-derived exosomes.Selective accumulation of the heat shock protein hc73. J. Cell Biol.147:599.
10 Wolfers, J., Lozier, A., Raposo, G., Regnault, A., Thery, C.,Masurier, C., Flament, C., Pouzieux, S., Faure, F., Tursz, T.,Angevin, E., Amigorena, S. and Zitvogel, L. 2001. Tumor-derivedexosomes are a source of shared tumor rejection antigens for CTLcross-priming. Nat. Med. 7:297.
11 Segal, D. M., Taurog, J. D. and Metzger, H. 1977. Dimericimmunoglobulin E serves as a unit signal for mast celldegranulation. Proc. Natl Acad. Sci. USA 74:2993.
12 Rothbard, J. B., Lechler, R. I., Howland, K., Bal, V., Eckels, D. D.,Sekaly, R., Long, E. O., Taylor, W. R. and Lamb, J. R. 1988.Structural model of HLA-DR1 restricted T cell antigen recognition.Cell 52:515.
13 Stumptner, P. and Benaroch, P. 1997. Interaction of MHC class II
T cell stimulation by exosomes 721

molecules with the invariant chain: role of the invariant chain (81±90) region. EMBO J. 16:5807.
14 Vincent-Schneider, H., Thery, C., Mazzeo, D., Tenza, D., Raposo,G. and Bonnerot, C. 2001. Secretory granules of mast cellsaccumulate mature and immature MHC class II molecules. J. CellSci. 114:323.
15 Winzler, C., Rovere, P., Rescigno, M., Granucci, F., Penna, G.,Adorini, L., Zimmermann, V. S., Davoust, J. and Ricciardi-Castagnoli, P. 1997. Maturation stages of mouse dendritic cellsin growth factor-dependent long-term cultures. J. Exp. Med.185:317.
16 van Bergen, J., Schoenberger, S. P., Verreck, F., Amons, R.,Offringa, R. and Koning, F. 1997. Ef®cient loading of HLA-DR witha T helper epitope by genetic exchange of CLIP. Proc. Natl Acad.Sci. USA 94:7499.
17 Boniface, J. J., Rabinowitz, J. D., Wul®ng, C., Hampl, J., Reich, Z.,Altman, J. D., Kantor, R. M., Beeson, C., McConnell, H. M. andDavis, M. M. 1998. Initiation of signal transduction through the Tcell receptor requires the multivalent engagement of peptide/MHC ligands [Corrected]. Immunity 9:459.
18 Lanzavecchia, A., Lezzi, G. and Viola, A. 1999. From TCRengagement to T cell activation: a kinetic view of T cellbehavior. Cell 96:1.
19 Arnold, P. and, Mannie, M. 1999. Vesicles bearing MHC class IImolecules mediate transfer of antigen from antigen-presentingcells to CD4+ T cells. Eur. J. Immunol. 29:1363.
20 Denzer, K., van Eijk, M., Kleijmeer, M. J., Jakobson, E., de Groot,C. and Heuze, J. G. 2000. Follicular dendritic cells carry MHCclass II-expressing microvesicles at their surface. J. Immunol.165:1259.
21 Geijtenbeek, T. B., Kwon, D. S., Torensma, R., van Vliet, S. J., vanDuijnhoven, G. C., Middel, J., Cornelissen, I. L., Nottet, H. S.,KewalRamani, V. N., Littman, D. R., Figdor, C. G. and van Kooyk,Y. 2000. DC-SIGN, a dendritic cell-speci®c HIV-1-binding proteinthat enhances trans-infection of T cells. Cell 100:587.
22 Shackelford, D. A., Kaufman, J. F., Korman, A. J. and Strominger,J. L. 1982. HLA-DR antigens: structure, separation ofsubpopulations, gene cloning and function. Immunol. Rev.66:133.
23 Shackelford, D. A., Lampson, L. A. and Strominger, J. L. 1983.Separation of three class II antigens from a homozygous human Bcell line. J. Immunol. 130:289.
24 Ziegler, A., Heinig, J., Muller, C., Gotz, H., Thinnes, F. P.,Uchanska-Ziegler, B. and Wernet, P. 1986. Analysis by sequentialimmunoprecipitations of the speci®cities of the monoclonalantibodies TU22, 34, 35, 36, 37, 39, 43, 58 and YD1/63. HLKdirected against human HLA class II antigens. Immunobiology171:77.
25 Adams, T. E., Bodmer, J. G. and Bodmer, W. F. 1983. Productionand characterization of monoclonal antibodies recognizing thealpha-chain subunits of human Ia alloantigens. Immunology50:613.
26 Kitani, S., Berenstein, E., Mergenhagen, S., Tempst, P. andSiraganian, R. P. 1991. A cell surface glycoprotein of rat
basophilic leukemia cells close to the high af®nity IgE receptor(Fc epsilon RI). Similarity to human melanoma differentiationantigen ME491. J. Biol. Chem. 266:1903.
27 Fleming, T. J., Donnadieu, E., Song, C. H., Laethem, F. V., Galli,S. J. and Kinet, J. P. 1997. Negative regulation of Fc epsilon RI-mediated degranulation by CD81. J. Exp. Med. 186:1307.
28 Barsumian, E. L., Isersky, C., Petrino, M. G. and Siraganian, R. P.1981. Ig-E induced histamine release from rat basophilicleukaemia cell lines: isolation of releasing and nonreleasingclones. Eur. J. Immunol. 11:317.
29 Benaroch, P., Yilla, M., Raposo, G., Ito, K., Miwa, K., Geuze, H. J.and Ploegh, H. L. 1995. How MHC class II molecules reach theendocytic pathway. EMBO J. 14:37.
30 Hewitt, C. R., Lamb, J. R., Hayball, J., Hill, M., Owen, M. J. andO'Hehir, R. E. 1992. Major histocompatibility complexindependent clonal T cell anergy by direct interaction ofStaphylococcus aureus enterotoxin B with the T cell antigenreceptor. J. Exp. Med. 175:1493.
31 Takebe, Y., Seiki, M., Fujisawa, J., Hoy, P., Yokota, K., Arai, M.,Yoshida, M. and Arai, N. 1988. SRalpha promoter: an ef®cient andversatile mammalian cDNA expression system composed of thesimian virus 40 early promoter and the R-U5 segment of human Tcell leukemia virus type I long terminal repeat. Mol. Cell. Biol.8:466.
32 Larhammar, D., Gustafsson, K., Claesson, L., Bill, P., Wiman, K.,Schenning, L., Sundelin, J., Widmark, E., Peterson, P. A. andRask, L. 1982. Alpha chain of HLA-DR transplantation antigens isa member of the same protein superfamily as theimmunoglobulins. Cell 30:153.
33 Bell, J. I., Estess, P., St John, T., Saiki, R., Watling, D. L., Erlich, H.A. and McDevitt, H. O. 1985. DNA sequence and characterizationof human class II major histocompatibility complex beta chainsfrom the DR1 haplotype. Proc. Natl Acad. Sci. USA 82:3405.
34 Latour, S., Bonnerot, C., Fridman, W. H. and Daeron, M. 1992.Induction of tumor necrosis factor-alpha production by mast cellsvia Fc gamma R. Role of the Fc gamma RIII gamma subunit. J.Immunol. 149:2155.
35 Thery, C., Boussac, M., Veron, P., Ricciardi-Castagnoli, P.,Raposo, G., Garin, J. and Amigorena, S. 2001. Proteomicanalysis of dendritic cell-derived exosomes: a secretedsubcellular compartment distinct from apoptotic vesicles. J.Immunol. 166:7309.
36 Bonnerot, C., Lankar, D., Hanau, D., Spehner, D., Davoust, J.,Salamero, J. and Fridman, W. H. 1995. Role of B cell receptor Igalpha and Ig beta subunits in MHC class II-restricted antigenpresentation. Immunity 3:335.
37 Stump, R. F., Oliver, J. M., Cragoe, E. J., Jr and Deanin, G. G.1987. The control of mediator release from RBL-2H3 cells: rolesfor Ca2+, Na+, and protein kinase C1. J. Immunol. 139:881.
38 Furuichi, K., Rivera, J. and Isersky, C. 1984. The fate of IgE boundto rat basophilic leukemia cells. III. Relationship between antigen-induced endocytosis and serotonin release. J. Immunol.133:1513.
722 T cell stimulation by exosomes





























