Embed Size (px)
Citation preview

Life Sciences 77 (2005) 3117–3126
www.elsevier.com/locate/lifescie
Extracellular inosine modulates ERK 1/2 and p38 phosphorylation
in cultured Sertoli cells: Possible participation in
TNF-alpha modulation of ERK 1/2
Luiz F. Souza a, Ana P. Horn a, Daniel P. Gelain a, Fernanda R. Jardim a,
Guido Lenz b, Elena A. Bernard a,TaDepartamento de Bioquımica, Instituto de Ciencias Basicas da Saude, Universidade Federal do Rio Grande do Sul,
(ICBS-UFRGS), Rua Ramiro Barcelos, 2600 anexo, CEP 90035-003, Porto Alegre, RS, BrazilbDepartamento de Biofısica, Instituto de Biociencias, Universidade Federal do Rio Grande do Sul, Porto Alegre, RS, Brazil
Received 13 December 2004; accepted 25 May 2005
Abstract
Extracellular ATP and adenosine modulation of MAPKs is well described in different cells types, but few studies
have addressed the effects of extracellular inosine on these kinases. Previous results showed that hydrogen peroxide
and TNF-a increase extracellular inosine concentration in cultured Sertoli cells and this nucleoside protects Sertoli
cells against hydrogen peroxide induced damage and participates in TNF-a induced nitric oxide production. In view
of the fact that MAPKs are key mediators of the cellular response to a large variety of stimuli, we investigated the
effect of extracellular inosine on the phosphorylation of ERK 1/2 and p38 MAPKs in cultured Sertoli cells. The
involvement of this nucleoside in the activation of ERK 1/2 by TNF-a was also investigated. Inosine and the selective
A1 adenosine receptor agonist R-PIA increases the phosphorylation of ERK 1/2 and p38, and this was blocked by the
selective A1 adenosine receptors antagonists, CPT and DPCPX. These antagonists also inhibited TNF-a increase in
the phosphorylation of ERK 1/2. TNF-a also rapidly augmented extracellular inosine concentration in cultured
Sertoli cells. These results show that extracellular inosine modulates ERK 1/2 and p38 in cultured Sertoli cells,
possible troughA1 adenosine receptor activation. This nucleoside also participates in TNF-a modulation of ERK 1/2.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Extracellular inosine; TNF-a; MAPKs; ERK; p38; Sertoli cells
0024-3205/$ -
doi:10.1016/j.l
T Correspond
E-mail add
see front matter D 2005 Elsevier Inc. All rights reserved.
fs.2005.05.049
ing author. Fax: +55 51 3316 5535/5540.
ress: [email protected] (E.A. Bernard).

L.F. Souza et al. / Life Sciences 77 (2005) 3117–31263118
Introduction
Extracellular purines are important regulators of many cellular functions. They activate P1 and P2
cell surface receptors. P1 receptors are mainly responsive to adenosine and P2 receptors to
extracellular nucleotides. P1 or adenosine receptors are divided into four subtypes: A1, A2A, A2B and
A3, which are all G protein-coupled receptors (Fredholm et al., 2001). P2 receptors are divided into
two distinct families: the P2X ligand-gated ionotropic channel family and the P2Y metabotropic G
protein-coupled family (Dubyak and El-Moatassim, 1993; Burnstock and Williams, 2000).
Extracellular inosine, the degradation product of adenosine deamination, was initially considered an
inactive metabolite of purine catabolism; however, recent publications have suggested its role in the
regulation of various processes. In mast cells, inosine stimulates degranulation via A3 adenosine
receptors, leading to increased vasopermeability in mice (Jin et al., 1997; Tilley et al., 2000). A
protective role has been assigned to inosine in many cell and animal models. In endotoxemia (Hasko
et al., 2000) and acute lung injury (Liaudet et al., 2002), inosine reduces the production of
inflammatory cytokines and increases the production of anti-inflammatory cytokines by macrophages,
improving the survival of mice. Extracellular inosine also prevents glial cell death during glucose
deprivation (Jurkowitz et al., 1998; Haun et al., 1996) and improves renal function in ischemia
(Fernando et al., 1976; De Rougemont et al., 1982). In addition, inosine protects RAW 264.7
macrophages from cell death induced by reactive oxygen species, by inhibiting poly (ADP-ribose)
polimerase activity (Virag and Szabo, 2001).
Sertoli cells are the somatic cells of seminiferous tubules, responsible for energy supply, mechanical
support and hormonal control of spermatogenesis. These cells express distinct purinoreceptors: A1
adenosine receptors, that are related to the inhibition of follicle stimulating hormone (FSH) increased
cAMP concentration (Monaco and Conti, 1986; Rivkees, 1994; Monaco et al., 1998); P2Y2 ATP
receptors, which regulate phosphatidyl-inositol turnover and calcium mobilization, inhibit cAMP
accumulation (Filippini et al., 1994) and increase c-glutamyl-transpeptidase and transferrin secretion
(Meroni et al., 1998); and ionotropic P2X(2,3 and 7) ATP receptors that are differentially present at
different stages of the seminiferous epithelium cycle (Glass et al., 2001). Previous works of our group
reported that Sertoli cells present different ectonucleotidases activities responsible for extracellular
purine metabolism (Casali et al., 2001) and that these cells secrete extracellular nucleotides and their
metabolites, including inosine (Gelain et al., 2003). This molecule has been shown to be involved in
TNF-a modulation of nitric oxide production (unpublished results) and protection against lipoperoxida-
tion and death induced by H2O2 (Gelain et al., 2004).
Mitogen-activated protein kinases (MAPKs) are serine/threonine kinases involved in the regulation
of cell cycle progression, proliferation, differentiation and death. The MAPK family consists of three
main subgroups: the extracellular signal-regulated kinases (ERKs), the stress-activated protein kinases
(SAPKs), also known as c-jun N terminal kinases (JNKs), and the p38 family. The modulation of
MAPKs by extracellular adenosine acting on any of the adenosine receptors (A1, A2A, A2B and A3)
through different mechanisms and in different cells is well described (revised in Schulte and
Fredholm, 2003). Extracellular ATP also modulates MAPKs through P2 receptors (Lenz et al., 2000;
Sellers et al., 2001; Shigemoto-Mogami et al., 2001; Bradford and Soltoff, 2002; Gendron et al.,
2003). However, the action of extracellular inosine on these kinases is not reported. In Sertoli cells,
ERK-dependent signalling is stage-specifically modulated by FSH during primary Sertoli cell
maturation (Crepieux et al., 2001) and transforming growth factor b 3 regulates various Sertoli

L.F. Souza et al. / Life Sciences 77 (2005) 3117–3126 3119
functions via MAPKs. (Lui et al., 2003). Furthermore, TNF-a activates ERK 1/2, p38 and JNK,
regulating interleukine-6 production, via p38, and intercellular adhesion molecule-1 expression, via
JNK (De Cesaris et al., 1998, 1999).
We investigated the modulation by extracellular inosine of ERK 1/2 and p38 MAPKs in cultured
Sertoli cells and studied the possible receptor involved. The role of extracellular inosine in the TNF-amodulation of ERK 1/2 was also investigated.
Materials and methods
Materials and animals
DMEM/F-12 medium was purchased from Gibco-BRL Life Technologies (Rockville, MD, USA).
Anti phospho-ERK 1/2 was purchased from Upstate (USA) and anti-phospho-p38 antibodies from
Promega (USA). TNF-a, R-PIA, CPT, DPCPX, MRS1220, inosine and all other drugs, reagents and
enzymes were purchased from Sigma Chemicals (St. Louis, MO, USA). Pregnant Wistar rats were
housed individually in plexiglass cages. Litters were restricted to eight pups each. The animals were
maintained on a 12 h light/dark cycle at a constant temperature of 23 8C, with free access to commercial
food and water. Male immature rats (18 days old) were killed by ether inhalation.
Isolation and culture of Sertoli cells
Sertoli cells were isolated as previously described (Rocha et al., 1997), following the method of Tung
and Fritz (Tung and Fritz, 1984) with modifications. Testes of immature 18 day-old Wistar rats were
removed, decapsulated and digested enzymatically with trypsin and deoxyribonuclease for 30 min at 34
8C, and then centrifuged at 750 g for 5 min. The pellet was washed with soybean trypsin inhibitor,
centrifuged and incubated with collagenase and hyaluronidase for 30 min at 34 8C followed by a
centrifugation for 10 min at 40 g. After this, the pellet was incubated with hyaluronidase for 30 min at 34
8C and then centrifuged at 40 g for 10 min. After counting, Sertoli cells were plated at a density of
3�105 cells/cm2 in DMEM:F12 (1 :1, low glucose) 1% FBS, supplemented with sodium bicarbonate,
HEPES and gentamicin, and maintained in a humidified atmosphere of 5% CO2 at 34 8C for 72 h. The
medium was changed every 24 h. Sertoli cell cultures were estimated to be 90–95% pure, as assessed by
alkaline phosphatase assay.
Western blots: ERK 1/2 and p38 phosphorylation
After 72 h, cultured Sertoli cells were incubated with or without inosine (0.01–1 mM), R-PIA(100
AM) or TNF-a 500 U/ml (25 ng/ml) for 5, 10 or 15 min. When present, adenosine receptor antagonists
were added 5 min before inosine, R-PIA or TNF-a. At the end of incubation, cultured Sertoli cells were
harvested with a cell scraper, centrifuged and lysed in 1% Triton/PBS. Equal amounts of soluble protein
(50 Ag) were run on SDS-PAGE 12% T, transferred to nitrocellulose and reacted with specific antibodies
for phosphorylated forms of ERK and p38. Peroxidase tagged secondary antibodies (anti immuno-
globulin) were used and the detection was performed using the enhanced chemiluminescence system
(ECL). Immunoblots were quantified by scanning of films followed by densitometric analysis. All

L.F. Souza et al. / Life Sciences 77 (2005) 3117–31263120
results were representative of three independent experiments and only statistically significant results are
reported in text.
Extracellular purine nucleotide assay
To investigate purine nucleotides and their metabolites in the incubation medium, we used the method
described by Cunha et al. (Cunha et al., 1989). Cultured Sertoli cells were gently washed three times to
remove remnants of medium and any dead or dying cells, and then incubated with HBSS (without
phenol-red, 15 mM HEPES) in the presence or absence of TNF-a 500 U/ml (25 ng/ml) for 5 min in 5%
CO2 at 34 8C. After incubation the medium was removed and centrifuged to eliminate debris. Samples
were evaporated in a vacuum centrifuge (�61 8C) and resuspended in 10% of the original volume.
Purines were determined by a reverse-phase HPLC system equipped with a C-18 column
(SupelcosilTM, SupelcoR, 25 cm�4.6 mm) and UV detector. The elution program consisted of: 10
min with 96% buffer A (KH2P04 100 mM, pH 6.5) and 4% buffer B (buffer A plus methanol 30%),
followed by a 5 min linear gradient up to 50% of buffer B, held for 10 min, at a flow rate of 1.25 ml/min
(UV absorption of 245 nm). Standard solutions were used to identify the substances detected.
Protein quantification
All the results were standardized against protein content, determined as described by Lowry et al.
(Lowry et al., 1951).
Results
We have previously described that Sertoli cell cultures secrete inosine, leading to the accumulation of
this purine in the extracellular medium (Gelain et al., 2003); this was induced by TNF-a (unpublished
results) and hydrogen peroxide (Gelain et al., 2004). MAPK cascades are involved in the response to
various stimuli, such as growth factors, cytokines and oxidative stress (Schulte and Fredholm, 2003;
Lander, 1997). To investigate whether extracellular inosine could modulate the activation of ERK 1/2 ,
the phosphorylation of this MAPK in cultured Sertoli cells treated with inosine was studied.
Extracellular inosine increased ERK 1/2 phosphorylation with maximal effect in 5 min, followed by
a slow dephosphorylation (Fig. 1A). The increase in phosphorylation of ERK 1/2 was seen at a
concentration of 0.1 mM inosine and was maximal at 1 mM (data not shown).
Fig. 1. Time dependence of extracellular inosine modulation of ERK 1/2 phosphorylation. After 72 h culture, Sertoli cells were
incubated for the indicated time periods in the presence of extracellular inosine 1 mM. Equal loading of cell lysates (50 Ag) wereresolved on SDS/PAGE and transferred to nytrocellulose before being probed with specific anti phospho-ERK 1/2.
Representative Western blot of three independent experiments is shown.
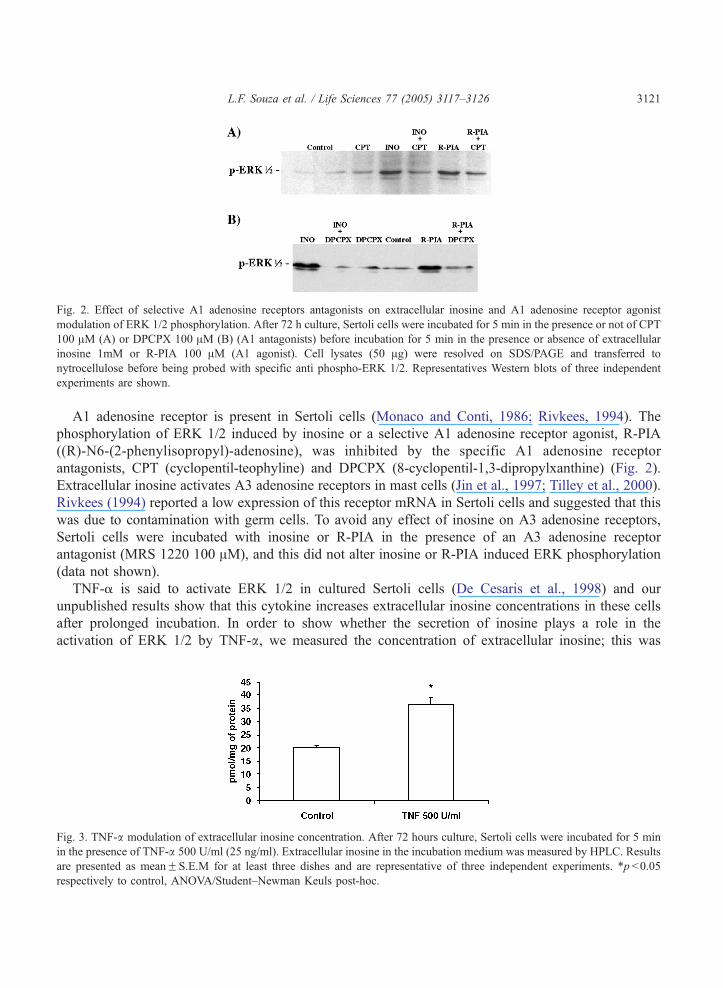
Fig. 2. Effect of selective A1 adenosine receptors antagonists on extracellular inosine and A1 adenosine receptor agonist
modulation of ERK 1/2 phosphorylation. After 72 h culture, Sertoli cells were incubated for 5 min in the presence or not of CPT
100 AM (A) or DPCPX 100 AM (B) (A1 antagonists) before incubation for 5 min in the presence or absence of extracellular
inosine 1mM or R-PIA 100 AM (A1 agonist). Cell lysates (50 Ag) were resolved on SDS/PAGE and transferred to
nytrocellulose before being probed with specific anti phospho-ERK 1/2. Representatives Western blots of three independent
experiments are shown.
L.F. Souza et al. / Life Sciences 77 (2005) 3117–3126 3121
A1 adenosine receptor is present in Sertoli cells (Monaco and Conti, 1986; Rivkees, 1994). The
phosphorylation of ERK 1/2 induced by inosine or a selective A1 adenosine receptor agonist, R-PIA
((R)-N6-(2-phenylisopropyl)-adenosine), was inhibited by the specific A1 adenosine receptor
antagonists, CPT (cyclopentil-teophyline) and DPCPX (8-cyclopentil-1,3-dipropylxanthine) (Fig. 2).
Extracellular inosine activates A3 adenosine receptors in mast cells (Jin et al., 1997; Tilley et al., 2000).
Rivkees (1994) reported a low expression of this receptor mRNA in Sertoli cells and suggested that this
was due to contamination with germ cells. To avoid any effect of inosine on A3 adenosine receptors,
Sertoli cells were incubated with inosine or R-PIA in the presence of an A3 adenosine receptor
antagonist (MRS 1220 100 AM), and this did not alter inosine or R-PIA induced ERK phosphorylation
(data not shown).
TNF-a is said to activate ERK 1/2 in cultured Sertoli cells (De Cesaris et al., 1998) and our
unpublished results show that this cytokine increases extracellular inosine concentrations in these cells
after prolonged incubation. In order to show whether the secretion of inosine plays a role in the
activation of ERK 1/2 by TNF-a, we measured the concentration of extracellular inosine; this was
Fig. 3. TNF-a modulation of extracellular inosine concentration. After 72 hours culture, Sertoli cells were incubated for 5 min
in the presence of TNF-a 500 U/ml (25 ng/ml). Extracellular inosine in the incubation medium was measured by HPLC. Results
are presented as meanFS.E.M for at least three dishes and are representative of three independent experiments. *p b0.05
respectively to control, ANOVA/Student–Newman Keuls post-hoc.
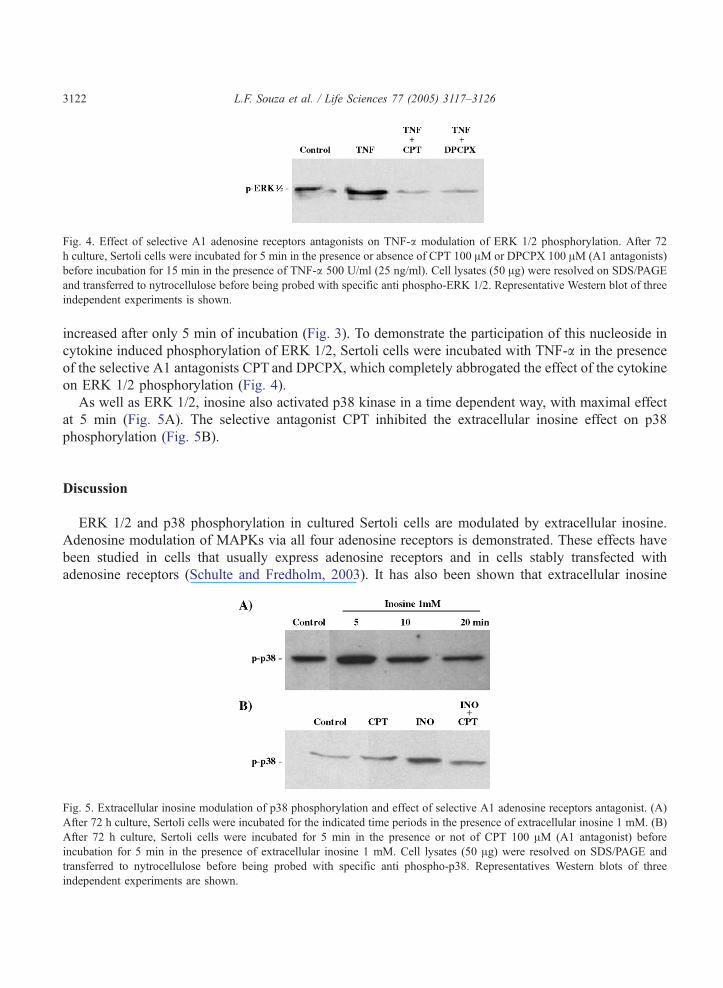
Fig. 4. Effect of selective A1 adenosine receptors antagonists on TNF-a modulation of ERK 1/2 phosphorylation. After 72
h culture, Sertoli cells were incubated for 5 min in the presence or absence of CPT 100 AM or DPCPX 100 AM (A1 antagonists)
before incubation for 15 min in the presence of TNF-a 500 U/ml (25 ng/ml). Cell lysates (50 Ag) were resolved on SDS/PAGE
and transferred to nytrocellulose before being probed with specific anti phospho-ERK 1/2. Representative Western blot of three
independent experiments is shown.
L.F. Souza et al. / Life Sciences 77 (2005) 3117–31263122
increased after only 5 min of incubation (Fig. 3). To demonstrate the participation of this nucleoside in
cytokine induced phosphorylation of ERK 1/2, Sertoli cells were incubated with TNF-a in the presence
of the selective A1 antagonists CPT and DPCPX, which completely abbrogated the effect of the cytokine
on ERK 1/2 phosphorylation (Fig. 4).
As well as ERK 1/2, inosine also activated p38 kinase in a time dependent way, with maximal effect
at 5 min (Fig. 5A). The selective antagonist CPT inhibited the extracellular inosine effect on p38
phosphorylation (Fig. 5B).
Discussion
ERK 1/2 and p38 phosphorylation in cultured Sertoli cells are modulated by extracellular inosine.
Adenosine modulation of MAPKs via all four adenosine receptors is demonstrated. These effects have
been studied in cells that usually express adenosine receptors and in cells stably transfected with
adenosine receptors (Schulte and Fredholm, 2003). It has also been shown that extracellular inosine
Fig. 5. Extracellular inosine modulation of p38 phosphorylation and effect of selective A1 adenosine receptors antagonist. (A)
After 72 h culture, Sertoli cells were incubated for the indicated time periods in the presence of extracellular inosine 1 mM. (B)
After 72 h culture, Sertoli cells were incubated for 5 min in the presence or not of CPT 100 AM (A1 antagonist) before
incubation for 5 min in the presence of extracellular inosine 1 mM. Cell lysates (50 Ag) were resolved on SDS/PAGE and
transferred to nytrocellulose before being probed with specific anti phospho-p38. Representatives Western blots of three
independent experiments are shown.

L.F. Souza et al. / Life Sciences 77 (2005) 3117–3126 3123
activates A3 adenosine receptors in mast cells (Jin et al., 1997; Tilley et al., 2000); however, in Sertoli
cells, inosine activated ERK 1/2 phosphorylation was not prevented by an A3 adenosine receptor
antagonist (MRS1220). The inhibititory effect of selective A1 antagonists, CPT and DPCPX, on
extracellular inosine modulation of ERK 1/2 and p38 phosphorylation, in addition to the activation of
these kinases by the A1 selective agonist R-PIA, suggests that extracellular inosine could be activating
the A1 adenosine receptor in Sertoli cells. Taking into account that in mast cells the binding of
extracellular inosine to this receptor is negligible (Jin et al., 1997), this could suggest that the A1
adenosine receptor expressed by Sertoli cells has a different affinity for extracellular inosine than the
receptor present on mast cells.
We have previously demonstrated the increase of extracellular inosine by hydrogen peroxide and
shown that this nucleoside protects Sertoli cells against oxidative stress-induced damage (Gelain et al.,
2004). The activation of ERK 1/2 is involved in the protection against oxidative induced cell death in
cAMP potentiation of noradrenaline neuroprotection on dopaminergic neurons (Troadec et al., 2002),
estrogen neuroprotection (Mize et al., 2003) and calcitonin gene-related peptide protection in cultured
smooth muscle cells (Schaeffer et al., 2003). In Sertoli cells, the inosine-stimulated phosphorylation of
ERK 1/2, described in this paper, could be involved in the protection against hydrogen peroxide
previously cited. Oxidative stress modulates MAPKs (Lander, 1997), suggesting that extracellular
inosine could participate in hydrogen peroxide modulation of Sertoli cell functions.
In the testis, TNF-a is produced, under physiological conditions, mainly by round spermatids (De et
al., 1993), and exerts a regulatory function on spermatogenesis. However, in inflammatory responses,
interstitial macrophages can account for considerable production of this cytokine (Xiong and Hales,
1993; Moore and Hutson, 1994). Sertoli cells express both TNF-a receptors (Mauduit et al., 1996; De
Cesaris et al., 1999) and this cytokine modulates diverse functions of these cells related to the
maintenance of spermatogenesis, including lactate production (Nehar et al., 1997) and interleukin-6
(Riccioli et al., 1995), inhibin (Magueresse-Battistoni et al., 1995), transferrin (Sigillo et al., 1999) and
insulin-like growth factor binding protein 3 expression (Besset et al., 1996). TNF-a also modulates
various FSH actions (Mauduit et al., 1993), the Fas-ligand system (Pentikainen et al., 2001) and tight
junction dynamics (Siu et al., 2003) in Sertoli cells.
Our unpublished results show that prolonged TNF-a treatment increases extracellular inosine
content in cultured Sertoli cells. It was also shown that this increase is related to increased nitric oxide
production. In the current article, we show that the TNF-a-induced increase in extracellular inosine
content is very rapid, indicating that this nucleoside is acting as a mediator in ERK 1/2 modulation.
This is supported by the results with selective A1 adenosine receptor antagonists CPT and DPCPX,
which inhibited cytokine stimulated ERK1/2 phosphorylation. Miki and Eddy (2002) recently reported
that TNF-R1 exhibits ATPase activity that is involved in the regulation of the aggregation of this
receptor when activated by TNF-a. This ATPase activity could lead to alteration in the intracellular
purine content when TNF-a binds to TNF-R1, which, via equilibrative nucleotide/nucleoside
transporters, could alter the extracellular purine level. No alteration of extracellular adenosine
concentration was observed in TNF-a treated Sertoli cells, which indicates that this nucleoside is not
involved.
In the present work, we have shown that extracellular inosine increases ERK 1/2 and p38
phosphorylation in Sertoli cells, possibly through A1 adenosine receptor activation. TNF-a also rapidly
increases the extracellular inosine concentration and A1 antagonists block TNF-a-induced phosphor-
ylation of ERK 1/2, suggesting that extracellular inosine plays a role in cytokine signalling.

L.F. Souza et al. / Life Sciences 77 (2005) 3117–31263124
Acknowledgements
This work was supported by CAPES, CNPq, FAPERGS and PROPESQ/UFRGS.
References
Besset, V., Magueresse-Battistoni, B., Collette, J., Benahmed, M., 1996. Tumor necrosis factor a stimulates insulin-like growth
factor binding protein 3 expression in cultured porcine Sertoli cells. Endocrinology 137 (1), 296–303.
Bradford, M.D., Soltoff, S.P., 2002. P2X7 receptor activate protein kinase D and p42/p44 mitogen activated protein kinase
(MAPK) downstream of protein kinase C. Biochemical Journal 366 (Pt 3), 745–755.
Burnstock, G., Williams, M., 2000. P2 purinergic receptors: modulation of cell function and therapeutic potential. Journal of
Pharmacology and Experimental Therapeutics 295 (3), 862–869.
Casali, E.A., da Silva, T.R., Gelain, D.P., Kaiser, G.R.R.F., Battastini, A.M.O., Sarkis, J.J.F., Bernard, E.A., 2001.
Ectonucleotidase activities in Sertoli cells from immature rats. Brazilian Journal of Medical and Biological Research 34 (10),
1247–1256.
Crepieux, P., Marion, S., Martinat, N., Fafeur, V., Vern, Y.L., Kerboeuf, D., Guillou, F., Reiter, E., 2001. The ERK-dependent
signalling is stage-specifically modulated by FSH, during primary Sertoli cell maturation. Oncogene 20 (34), 4696–4709.
Cunha, R.A., Sebastiao, A.M., Ribeiro, J.A., 1989. Separation of adenosine�triphosphate and its degradation products in
innervated muscle of the frog by reverse phase high-performance liquid-chromatography. Chromatographia 28 (2), 610–612.
De Cesaris, P., Starace, D., Riccioli, A., Padula, F., Filippini, A., Ziparo, E., 1998. Tumor necrosis factor-a induces interleukin-6
production and integrin ligand expression by distinct transduction pathways. Journal of Biological Chemistry 273 (13),
7566–7571.
De Cesaris, P., Starace, D., Starace, G., Filippini, A., Stefanini, M., Ziparo, E., 1999. Activation of jun N-terminal kinase/stress-
activated protein kinase pathway by tumor necrosis factor a leads to intercellular adhesion molecule-1 expression. Journal of
Biological Chemistry 274 (41), 28978–28982.
De Rougemont, D., Brunner, F.P., Torhorst, J., Winderlich, P.F., Thiel, G., 1982. Superficial nephron obstruction and medullary
congestion after ischemic injury: effect of protective treatments. Nephron 31 (4), 310–320.
De, S.K., Chen, H., Pace, J.L., Hunt, J.S., Terranova, P.F., Enders, G.C., 1993. Expression of tumor necosis factor-a in mouse
spermatogenic cells. Endocrinology 133 (1), 389–396.
Dubyak, G.R., El-Moatassim, C., 1993. Signal transduction via P2-purinergic receptors for extracellular ATP and other
nucleotides. American Journal of Physiology 265 (3 Pt 1), C577–C606.
Fernando, A.R., Armstrong, D.M., Griffths, J.R., Hendry, W.F., O’Donoghue, E.P., Perrett, D., Ward, J.P., Wickham, J.E., 1976.
Enhanced preservation of the ischaemic kidney with inosine. Lancet 1 (7959), 555–557.
Filippini, A., Riccioli, A., De Cesaris, P., Paniccia, R., Teti, A., Stefanini, M., Conti, M., Ziparo, E., 1994. Activation of inositol
phospholipid turnover and calcium signaling in rat Sertoli cells by P2-purinergic receptors: modulation of folliclestimulating
hormone responses. Endocrinology 134 (3), 1537–1545.
Fredholm, B.B., Ijzerman, A.P., Jacobson, K.A., Klotz, K., Linden, J., 2001. International Union of Pharmacology: XXV.
Nomenclature and classification of adenosine receptors. Pharmacological Reviews 53 (4), 527–552.
Gelain, D.P., Souza, L.F., Bernard, E.A., 2003. Extracellular purines from cells of seminiferous tubules. Molecular and Cellular
Biochemistry 245 (1–2), 1–9.
Gelain, D.P., Souza, L.F., Ribeiro, G.R., Zim, M., Jardim, F.R., Moreira, J.C.F., Bernard, E.A., 2004. Extracellular inosine
is modulated by H2O2 and protects Sertoli cells against lipoperoxidation and cellular injury. Free Radical Research 38
(1), 37–47.
Glass, R., Bardinini, M., Robson, T., Burnstock, G., 2001. Expression of nucleotide P2X receptor subtypes during
spermatogenesis in the adult rat testis. Cells Tissues Organs 169 (4), 377–387.
Gendron, F.P., Neary, J.T., Theiss, P.M., Sun, G.Y., Gonzalez, F.A., Weisman, G.A., 2003. Mechanisms of P2X7 receptor-
mediated ERK1/2 phosphorylation in human astrocytoma cells. American Journal of Physiology-Cell Physiology 284 (2),
C571–C581.
Hasko, G., Kuhel, D.G., Nemeth, Z.H., Mabley, J.G., Stachlewitz, R.F., Virag, L., Lohinai, Z., Southan, G.J., Salzman, A.L.,
Szabo, C., 2000. Inosine inhibits inflammatory cytokine production by a posttranscriptional mechanism and protects against
endotoxin-induced shock. Journal of Immunology 164 (2), 1013–1019.

L.F. Souza et al. / Life Sciences 77 (2005) 3117–3126 3125
Haun, S.E., Segelon, J.E., Trapp, V.L., Clotz, M.A., Horrocks, L.A., 1996. Inosine mediates the protective effect of adenosine in
rat astrocyte cultures subjected to combined glucose-oxygen deprivation. Journal of Neurochemistry 67 (5), 2051–2059.
Jin, X., Shepherd, R.K., Duling, B.R., Linden, J., 1997. Inosine binds to A3 adenosine receptors and stimulates mast cell
degranulation. Journal of Clinical Investigation 100 (11), 2849–2857.
Jurkowitz, M.S., Litsky, M.L., Browning, M.J., Hohl, C.M., 1998. Adenosine, inosine and guanosine protect glial cells during
glucose deprivation and mitochondrial inhibition: correlation between protection and ATP preservation. Journal of
Neurochemistry 71 (2), 535–548.
Lander, H.M., 1997. An essential role for free radicals and derived species in signal transduction. FASEB Journal 11 (2),
118–124.
Lenz, G., Gottfried, C., Luo, Z., Avruch, J., Rodnight, R., Nie, W.J., Kang, Y., Neary, J.T., 2000. P(2Y) purinoceptor subtypes
recruit different mek activators in astrocytes. British Journal of Parmacology 129 (5), 927–936.
Liaudet, L., Mabley, J.G., Pacher, P., Virag, L., Soriano, F.G., Marton, A., Hasko, G., Deitch, E.A., Szabo, C., 2002.
Inosine exerts a broad range of antiinflammatory effects in a murine model of acute lung injury. Annals of Surgery 235
(4), 568–578.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurement with folin phenol reagent. Journal of
Biological Chemistry 193 (1), 265–275.
Lui, W.Y., Wong, C.H., Mruk, D.D., Cheng, C.Y., 2003. TGF-beta3 regulates the blood-testis barrier dynamics via the p38
mitogen activated protein (MAP) kinase pathway: an in vivo study. Endocrinology 144 (4), 1139–1142.
Magueresse-Battistoni, B.L., Morera, A.M., Benahmed, M., 1995. In vitro regulation of rat Sertoli cell inhibin
messenger RNA levels by transforming growth factor-b1 and tumor necrosis factor a. Journal of Endocrinology 146
(3), 501–508.
Mauduit, C., Jaspar, J.M., Poncelet, E., Charlet, C., Revol, A., Franchimont, P., Benahmed, M., 1993. Tumor necrosis factor-aantagonizes follicle-stimulating hormone action in cultured Sertoli cells. Endocrinology 133 (1), 69–76.
Mauduit, C., Besset, V., Caussanel, V., Benahmed, M., 1996. Tumor necrosis factor a receptor p55 is under hormonal (follicle-
stimulating hormone) control in testicular Sertoli cells. Biochemical and Biophysical Research Communications 224 (3),
631–637.
Meroni, S.B., Canepa, D.F., Pellizzari, E.H., Schteingart, H.F., Cigorraga, S.B., 1998. Effects of purinergic agonists on
aromatase and gamma-glutamyl transpeptidase activities and on transferrin secretion in cultured Sertoli cells. Journal of
Endocrinology 157 (2), 275–283.
Miki, K., Eddy, E.M., 2002. Tumor necrosis factor receptor 1 is an ATPase regulated by silencer of death domain. Molecular
and Cellular Biology 22 (8), 2536–2543.
Mize, A.L., Shapiro, R.A., Dorsa, D.M., 2003. Estrogen receptor-mediated neuroprotection from oxidative stress requires
activation of the mitogen-activated protein kinase pathway. Endocrinology 144 (1), 306–312.
Monaco, L., Conti, M., 1986. Localization of adenosine receptors in rat testicular cells. Biology of Reproduction 35 (2),
258–266.
Monaco, L., Manno, D.A., Martin, M.W., Conti, M., 1998. Adenosine inhibition of the hormonal response in the Sertoli cell is
reversed by pertussis toxin. Endocrinology 122 (6), 2692–2698.
Moore, C., Hutson, J.C., 1994. Physiological relevance of tumor necrosis factor in mediating macrophage-Leydig cell
interactions. Endocrinology 134 (1), 63–69.
Nehar, D., Mauduit, C., Boussouar, F., Benahmed, M., 1997. Tumor necrosis factor-a-stimulated lactate production is linked
to lactate dehydrogenase A expression and activity increase in porcine cultured Sertoli cells. Endocrinology 138 (5),
1964–1971.
Pentikainen, V., Erkkıla, K., Suomalainen, L., Otala, M., Pentikainen, M.O., Parvinen, M., Dunkel, L., 2001. TNFa down-
regulates the Fas ligand and inhibits germ cell apoptosis in the human testis. Journal of Clinical Endocrinology and
Metabolism 86 (9), 4480–4488.
Riccioli, A., Filippini, A., De Cesaris, P., Barbacci, E., Stefanini, M., Starace, G., Ziparo, E., 1995. Inflammatory mediators
increase surface expression to lymphocytes, and secretion of interleukin 6 in mouse Sertoli cells. Proceedings of the National
Academy of Sciences of the United States of America 92 (13), 5808–5812.
Rivkees, S.A., 1994. Localization and characterization of adenosine receptor expression in rat testis. Endocrinology 135 (6),
2307–2313.
Rocha, A.B., Guma, F.C.R., Casali, E.A., Scherer, G.S., Elena, M.A., Bernard, E.A., 1997. Influence of the biomatrix on the
response of Sertoli cells to FSH. Archives of Physiology and Biochemistry 105 (5), 473–477.

L.F. Souza et al. / Life Sciences 77 (2005) 3117–31263126
Schaeffer, C., Vandroux, D., Thomassin, L., Athias, P., Rochette, L., Connat, J.L., 2003. Calcitonin gene-related peptide partly
protects cultured smooth muscle cells from apoptosis induced by an oxidative stress via activation of ERK1/2 MAPK.
Biochimica et Biophysica Acta 1643 (1–3), 65–73.
Schulte, G., Fredholm, B.B., 2003. Signalling from adenosine receptors to mitogen-activated protein kinases. Cellular
Signalling 15 (9), 813–827.
Sellers, L.A., Simon, J., Lundahl, T.S., Cousens, D.J., Humphrey, P.P.A., Barnard, E.A., 2001. Adenosine nucleotides acting at
the human P2Y1 receptor stimulate mitogen-activated protein kinases and induce apoptosis. Journal of Biological Chemistry
276 (19), 16379–16390.
Shigemoto-Mogami, Y., Koizumi, S., Tsuda, M., Ohsawa, K., Kohsaka, S., Inoue, K., 2001. Mechanisms underlying
extracellular ATP-evoked interleukin-6 release in mouse microglial cell line, MG-5. Journal of Neurochemistry 78 (6),
1339–1349.
Sigillo, F., Guillou, F., Benahmed, M., Magueresse-Battistoni, B.L., 1999. In vitro regulation of rat Sertoli cell transferrin
expression by tumor necrosis factor a and retinoic acid. Molecular and Cellular Endocrinology 148 (1–2), 163–170.
Siu, M.K.Y., Lee, W.M., Cheng, C.Y., 2003. The interplay of collagen IV, tumor necrosis factor-a, gelatinase B (matrix
metalloprotease-9), and tissue inhibitor of metalloproteases-1 in the basal lamina regulates Sertoli cell-tight junction
dynamics in the rat testis. Endocrinology 144 (1), 371–387.
Tilley, S.L., Wagoner, V.A., Salvatore, C.A., Jacobson, M.A., Koller, B.H., 2000. Adenosine and inosine increase cutaneous
vasopermeality by activatyng A3 receptors on mast cells. Journal of Clinical Investigation 105 (3), 361–367.
Troadec, J.D., Marien, M., Mourlevat, S., Debeir, T., Ruberg, M., Colpaert, F., Michel, P.P., 2002. Activation of the mitogen-
activated protein kinase (ERK(1/2)) signaling pathway by cyclic AMP potentiates the neuroprotective effect of the
neurotransmitter noradrenaline on dopaminergic neurons. Molecular Pharmacology 62 (5), 1043–1052.
Tung, P.S., Fritz, I.B., 1984. Extracellular matrix promotes rat Sertoli cell histotypic expression in vitro. Biology of
Reproduction 30 (1), 213–229.
Virag, L., Szabo, C., 2001. Purines inhibit poly(ADP-ribose) polymerase activation and modulate oxidant-induced cell death.
FASEB Journal 15 (1), 99–107.
Xiong, Y., Hales, D.B., 1993. Expression, regulation and production of tumor necrosis factor-a in mouse testicular interstitial
macrophages in vitro. Endocrinology 133 (6), 2568–2573.





























