Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Journal of Human Evolution xxx (2013) 1e8
Contents lists available
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
Extreme mobility in the Late Pleistocene? Comparing limb biomechanics amongfossil Homo, varsity athletes and Holocene foragers
Colin N. Shawa,b,*, Jay T. Stock a
a PAVE Research Group, Department of Archaeology and Anthropology, University of Cambridge, Downing Street, Cambridge, Cambridgeshire CB2 3ER, UKbMcDonald Institute for Archaeological Research, Department of Archaeology and Anthropology, University of Cambridge, Cambridge, UK
a r t i c l e i n f o
Article history:Received 31 July 2012Accepted 9 January 2013Available online xxx
Keywords:Peripheral Quantitative ComputedTomography (pQCT)Fossil homininDiaphyseal cross-sectionHumerusTibia
* Corresponding author.E-mail addresses: [email protected], cshaw111@m
0047-2484/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.jhevol.2013.01.004
Please cite this article in press as: Shaw, C.NHomo, varsity athletes and Holocene forage
a b s t r a c t
Descriptions of Pleistocene activity patterns often derive from comparisons of long bone diaphysealrobusticity across contemporaneous fossilized hominins. The purpose of this study is to augment existingunderstanding of Pleistocene hominin mobility patterns by interpreting fossil variation through com-parisons with a) living human athletes with known activity patterns, and b) Holocene foragers wheredescriptions of group-level activity patterns are available. Relative tibial rigidity (midshaft tibial rigidity(J)/midshaft humeral rigidity (J)) was compared amongst Levantine and European Neandertals, Levantineand Upper Palaeolithic Homo sapiens, Holocene foragers and living human athletes and controls. Cross-country runners exhibit significantly (p < 0.05) greater relative tibial rigidity compared with swimmers,and higher values compared with controls. In contrast, swimmers displayed significantly (p < 0.05) lowerrelative tibial rigidity than both runners and controls. While variation exists among all HoloceneH. sapiens, highly terrestrially mobile Later Stone Age (LSA) southern Africans and cross-country runnersdisplay the highest relative tibial rigidity, while maritime Andaman Islanders and swimmers display thelowest, with controls falling between. All fossil hominins displayed relative tibial rigidity that exceeded,or was similar to, the highly terrestrially mobile Later Stone Age southern Africans and modern humancross-country runners. The more extreme skeletal structure of most Neandertals and LevantineH. sapiens, as well as the odd Upper Palaeolithic individual, appears to reflect adaptation to intense and/or highly repetitive lower limb (relative to upper limb) loading. This loading may have been associatedwith bipedal travel, and appears to have been more strenuous than that encountered by even universityvarsity runners, and Holocene foragers with hunting grounds 2000e3000 square miles in size. Skeletalvariation among the athletes and foraging groups is consistent with known or inferred activity profiles,which support the position that the Pleistocene remains reflect adaptation to extremely active andmobile lives.
� 2013 Elsevier Ltd. All rights reserved.
Introduction
The relationship between biomechanically imposed loads andskeletal adaptation is quite complex (Pearson and Lieberman,2004). Nevertheless, experimental research supports the generalconcept of bone functional adaptation, and the use of diaphysealcross-sectional properties to explore patterns of habitual behavioramong groups (Currey, 1984; Lanyon and Rubin, 1984; Martin andBurr, 1989; Rubin et al., 1990; Lanyon, 1992; Ruff et al., 2006). An-alyses of the femoral and tibial diaphyseal structure of PleistoceneHomo have concluded that while morphological variation is appa-rent, there has been little shift in locomotor ranging levels amongst
ac.com (C.N. Shaw).
All rights reserved.
., Stock, J.T., Extreme mobilityrs, Journal of Human Evolutio
the foraging populations from this period (c.f. Trinkaus and Ruff,2012). Interpretations of Pleistocene mobility patterns, however,might be augmented through direct comparisons with more recentforagers and living humans where activity patterns have beenbetter documented.
Early analyses concluded that the Neandertal lower limb wasbetter adapted to extended bouts of heavy physical activity com-pared with those of early modern humans (Trinkaus, 1989). Thisfinding led to the suggestion that Neandertals spent comparativelymore time moving continuously and/or vigorously across thelandscape. This interpretation was later reconsidered (c.f. Trinkaus,1997) following the implementation of methodology that bettercontrolled for the influence of body size on diaphyseal geometricproperties (Ruff, 2000a). Later analyses of Neandertals and Levan-tine anatomically modern Homo sapiens lower limb diaphysealstrength concluded that, once properly standardised, differences
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004

C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e82
between these groups were negligible (Trinkaus and Ruff, 1999a,b).More recently, Trinkaus and Ruff’s (2012) detailed assessment of allknown Pleistocene hominins concluded that despite a significantchange in lower limb diaphyseal morphology throughout thePleistocene, there appears to have been little change in the overallrobustness of the femur or tibia within Homo. When scaledappropriately, femoral midshaft rigidity did not show a significanttrend from the Early to the Late Pleistocene, although there wasa suggestion of a slight reduction with the earlier Upper Palae-olithic. In the tibial diaphysis, there were decreases in relativecortical area, especially between the Early Pleistocene and laterhuman groups. Increased tibial diaphyseal circularity (antero-pos-terior versus medio-lateral) was found between the Early Pleisto-cene and later samples, and then again with the earlier UpperPalaeolithic. The authors suggest that these results reflect theimportance of mobility among all Pleistocene foragers, and thatvariation in femoral diaphysis shape may reflect differential rang-ing patterns among later Upper Palaeolithic and Holocene groups(e.g., Holt, 2003; Holt and Formicola, 2008), but not across Pleis-tocene Homo.
Interpretations of prehistoric hominin activity patterns oftendepend primarily upon comparisons of fossilized skeletal structure(Trinkaus et al., 1994; Churchill et al., 1996; Holt, 2003; Holt andFormicola, 2008; Marchi, 2008; Ruff, 2008a, 2009). In partial con-trast, studies of pre- and protohistoric Holocene skeletal pop-ulations often benefit from a greater breadth of associatedarchaeological and ethnographic evidence for behavior, and largersample sizes with which to interpret within-population morpho-logical variation. This type of contextual information allows formore detailed descriptions of general group-level activity patterns(Larsen et al., 1995; Stock and Pfeiffer, 2001, 2004; Auerbach andRuff, 2006; Stock, 2006; Wanner et al., 2007). A third, and rela-tively unexplored, approach (see Trinkaus et al., 1994; Ruff et al.,1998; Ruff, 2000b) compares fossilized hominins against groupsof living humans with known activity patterns and well docu-mented skeletal morphology (i.e., Shaw and Stock, 2009a,b; Shaw,2010, 2011). To inform previous interpretations and provide a fur-ther test of Pleistocene mobility patterns, this study compares therobusticity and ‘pattern’ of robusticity (tibial J/humeral J) in Pleis-tocene hominins, highly terrestrially mobile and marine-adaptedHolocene foragers, and living human cross-country runners andswimmers.
Two main hypotheses are tested: (1) populations with knownand inferred high terrestrial mobility (runners and Later Stone Age(LSA) southern Africans, respectively) will share greater robusticityof lower limb elements relative to upper; and (2) comparisons ofrelative tibial rigidity will reveal that Pleistocene fossil homininswere adapted to levels of terrestrial mobility that are comparablewith, or even exceed, that of highly trained living cross-countryrunners and highly terrestrially mobile Holocene foragers.
Materials and methods
Modern human athletes and controls
A total of 50 modern human males between the ages of 19 and30 participated in this study. Subgroups included varsity-leveldistance runners (15) and swimmers (15), and non-athletic con-trols (20). All were recruited from the University of Cambridge, UK.The mean age of all subjects (runners (23.2 � 3.2), swimmers(21.9� 2.5), controls (21.6� 2.5)) was 22.2 (�2.8) years. All athletesbegan their competitive playing and training career during latechildhood/early adolescence (runners; 13.7 years of age, swim-mers; 10.5 years of age), and had been competing in their respec-tive sport for an average of 10 years (runners; 9.7 years, swimmers;
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
10.3 years). Athletic, lifestyle, and medical history was obtainedthrough a detailed questionnaire (Shaw and Stock, 2009b). A singlescan from each of the arms and shanks of each participant resultedin a total of 200 cross-sectional pQCT (peripheral QuantitativeComputed Tomography) images captured at 50% of limb segmentlength using an XCT-2000 (Stratec Medizintechnik GmbH, Pforz-heim, Germany). Raw cross-sectional pQCT images were importedinto Image J (http://rsb.info.nih.gov/ij/) and analyzed usingMoment Macro (http://www.hopkinsmedicine.org/FAE/mmacro.htm).
Holocene skeletal material
The skeletal remains of two Holocene forager populations wereincluded in this study: 16 LSA southern African males and 15 pro-tohistoric Andaman Islander (AI) males. The methods used to cal-culate cross-sectional diaphyseal properties for these populationshave been described previously (Stock and Pfeiffer, 2001). Briefly,cross-sectional dimensions were calculated using two separatemethods, the first involving CT images taken perpendicular to thediaphysis at the section location, the second, a procedure thatcombined information from casts of the periosteal contour andbiplanar radiographs (Trinkaus and Ruff, 1989).
Detailed descriptions of both populations are available in Stockand Pfeiffer (2001). In summary, the LSA southern Africans datefrom between 11,000 and 2000 BP (Before Present), and aredescribed as highly terrestrially mobile. Subsistence was charac-terized by the hunting of small game, terrestrial foraging, and theintensive exploitation of coastal marine resources (Deacon, 1993).An early model predicted that the area over which a band wouldhunt may have been as large as 2000e3000 square miles (Clarke,1959). Resources were broadly distributed, and exploitation,whether terrestrial or marine, was land-based. The terrain of thesouthern African region is generally rocky, with considerable ver-tical relief. Therefore, the exploitation of diverse terrestrial andmarine resources throughout this landscape would have requiredthe negotiation of rugged terrain.
The Andaman Islander foragers date to the late-1800s and hadmobility patterns that were constrained terrestrially and weregenerally marine based. Dietary staples included various fruits andyams in addition to wild pigs and honey, while marine huntingprovided fish, dugong and sea turtles (Myka, 1993). The Andama-nese frequently used the canoe for transportation and food pro-curement. Swimming was a common activity in both sexes, withchildren learning to swim almost as soon as they could walk, and allAndamanese, regardless of age and sex, spending up to severalhours in the water at a time (Man, 1883).
Pleistocene fossil hominin material
Table 1 lists the hominin fossil material included in this study, aswell as relevant provenance information. Analyses were performedon data from three male and one female anatomically modernLevantine H. sapiens (AMHS; Qafzeh 8, 9, Skhul IV and V ), threemale and two female Homo neanderthalensis (Spy 2, La Chapelle 1,La Ferrassie 1, 2, Tabun C1), five male European Early UpperPalaeolithic (EUP) H. sapiens (Paviland 1, Grotte des Enfants 4, DolniVestonici 13, 14, 16), six male European (Gough’s Cave 1, Neussing 1,Obercassel 1, Romanelli 1, Romito 3, Veyrier 1) and 20 male NorthAfrican (Wadi Halfa 2, 3, 24, 26, 31, 37; Jebel Sahaba 21, 29, 38, 39,40, 42; Afalou 2, 13, 25, 27, 28; El Wad 10259, 10260, 10263) LateUpper Palaeolithic (LUP) H. sapiens. Cross-sectional geometricmeasurements for these fossil hominins, taken at the midshaft ofthe tibia and humerus, were obtained from the relevant literature(see Table 2 for citations).
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004

Table 2Relative tibial rigidity (tibial J:humeral J), and raw tibial and humeral J.
n Mass(kg)
Tibial J:Humeral J
Tibial J(mm4)
Humeral J(mm4)
HoloceneLSA SA 19 52.35
(4.94)4.56 30,685 (6997) 6736 (2405)
Runner 15 68.21(5.94)
2.76 51,427 (10,824) 18,642 (3892)
Control 20 69.68(11.67)
2.63 39,091 (7893) 14,849 (3478)
Andaman Is. 15 48.52 2.36 17,326 (2396) 7342 (2077)
C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e8 3
Comparisons
Cross-sectional geometric properties used in this investigationare limited to polar secondmoments of area (J). J is an estimate of thebonesability to resist diaphyseal torsionand (twice) average bendingforces (Ruff, 2008b) that account for a high proportion of midshaftstrains (BertramandBiewener,1988). As such, J is a relevant indicatorof a bone’s overall mechanical performance (Ruff et al., 1993). Rawmeasures of J taken at the tibial and humeral midshaft were used tocalculate relative tibial rigidity (tibial J/humeral J). When right andleft elements were available, relative tibial J was calculated using theequation ((L þ R tibia)/2)/((Lþ R humerus)/2), if either element wasunavailable, the value for the available side alone was used. Usingestimates of relative tibial rigidity, one can assess differences in thedistribution of robusticity in the upper and lower limbs that may notbe apparentwhen comparing upper or lower limbs in isolation. Highratio values can reflect either high relative tibial robusticity or lowhumeral robusticity. Assessment of this data through bivariate plotsallows one to determinewhich factor is driving the ratio for differentindividuals and populations. Additionally, ratios avoid difficultiesassociated with standardising for differences in body size acrossspecies where mass and stature can be quite large.
Thefirst analysis utilizedunivariate analysis of variance (ANOVA)to compare relative tibial rigidity among modern human athletes,controls, andHolocene foragers. Differences in relative tibial rigiditywere contrastedusing eitherHochberg’sGT2orGamesHowell post-hoc tests, based on the results of Levene’s Test for equality of var-iance. Significance was recognized at p � 0.05. Slope values, calcu-lated using Least Squares Linear Regression, were also comparedamong these populations. The second set of analyses included livinghumanathletes and controls, Holocene foragers and fossil hominins.Restrictions imposed by the small number of individuals that makeup most Pleistocene groups necessitated that non-statistical com-parisons were performed. Ratios were ordered and plotted, andfrom this descriptive comparisons were produced.
Table 1Fossil hominin provenance.
Sex Date(ca. years BP)
Provenance
NeandertalSpy 2 M 36,000a Spy, BelgiumLa Chapelle 1 M 47e56,000e Corrèze, FranceLa Ferrassie 1 M 72,000d Dordogne, FranceLa Ferrassie 2 F 72,000d Dordogne, FranceTabun C1 F 122,000b Western IsraelAMHSQafzeh 8 M 100,000c Western IsraelQafzeh 9 F 100,000c Western IsraelSkhul IV M 100,000c Western IsraelSkhul V M 100,000c Western IsraelUpper PalaeolithicEUP-Europe M 24e29,000f Wales, Italy, Czech
RepublicLUP-Europe M 9100e18,000g France, Italy,
GermanyLUP-N. Africa M EpiPalaeolithic e Natufianh Sudan, Israel,
Algeria
a Semal et al. (2009).b Grün and Stringer (2000).c McDermott et al. (1993).d Delporte (1984).e Verneau (1906).f Raynal (1990), Svoboda (1995), Trinkaus and Svoboda (2006), Jacobi and
Higham (2008).g Davies (1904), Bonnet (1913e14), Seligman and Parsons (1914), Verworn et al.
(1919), Pittard and Sauter (1945), Narr (1977), Mallegni and Fabbri (1995), Alciatiet al. (2005).
h Boule et al. (1934), McCown (1939), Anderson (1968), Greene and Armelagos(1972), Chamla (1978).
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
Results
Modern human athletes, controls, and Holocene foragers
Table 2 displays raw tibial J and humeral J values, and relativetibial rigidity (tibial J/humeral J) calculations, for each group. TheLSA southern Africans and runners display the highest relativetibial rigidity (4.56 and 2.76, respectively), while Andaman Is-landers and swimmers display the lowest (2.36 and 1.84, respec-tively). The relative tibial rigidity displayed by controls (2.63) fallsin the middle of all other groups. Differences in relative tibial ri-gidity between these groups are presented in Table 3, and also Fig.1.The results indicate a separation of groups at the extremes; thegroups with the highest (LSA southern Africans) and lowest(swimmers) relative tibial rigidity are both significantly differentfrom all other groups (in all cases p < 0.05). No significant differ-ences were found among the groups (runners, controls and Anda-man Islanders) that fall between these extremes. Table 4 providesslope characteristics for the relationship between raw tibial J andhumeral J for all groups. The slope of the line that describes the LSAsouthern Africans is significantly steeper (falling outside of the 95%confidence interval, therefore p < 0.05), compared with all other
(4.81)Swimmer 15 74.27
(7.72)1.84 41,771 (7474) 22,658 (5779)
NeandertalsLa Ferrassie 2 1 66.4a 5.03 30,631a 6084b,R
La Ferrassie 1 1 84.9a 4.63 62,285a 13,447b, R&L
Tabun C1 1 63.3a 3.88 27,216a 7015c, R&L
Spy 2 1 85.5a 3.81 52,606a 13,795b, R&L
La Chapelle 1 1 81.1a 3.33 58,502a 17,578b, R&L
AMHSSkhul V 1 68.8a 6.55 86,569a 13,215c, R&L
Skhul IV 1 69.0a 5.07 63,838a 12,590c, L
Qafzeh 9 1 63.3a 4.25 69,506a 16,346c, R
Qafzeh 8 1 e 2.81 65,702a 23,400c, R
Upper PalaeolithicEUP-Europe 5 75.76
(5.02)a3.28 47,807 (13,692)a,e 15,312 (4341)d,e,f,g
LUP-Europe 6 68.18(8.07)e
2.91 48,279 (13,876)e 16,853 (5747)e,d
LUP-N. Africa 20 66.30(5.68)e,i
3.03 51,956 (11,483)e,h 18,152 (4709)e,h
Data is presented as: mean (standard deviation).J: polar secondmoment of area, Andaman Is.: Andaman Islander, LSA SA: Later StoneAge southern African, AMHS; anatomically modern Homo sapiens.Fossil specimen humeral J calculated from skeletal elements from R (right-side), L(left-side), R&L (average of right & left).Cross-sectional geometric property data for fossil hominins obtained from:
a Trinkaus and Ruff (2012).b Trinkaus et al. (1994).c Trinkaus and Churchill (1999).d Churchill (1994).e Laura Shackelford (Personal communication).f Trinkaus (2000).g Sladek et al. (2000).h Shackelford (2007).i Mass estimates for Jebel Sahaba 21 and 42 were not available for inclusion.
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004

Table 3P-values for ANOVA comparisons of relative tibia torsional rigidity (tibial J:humeralJ) among modern human athletes and controls, and Holocene foragers.
Groups Runners Swimmers Controls LSA SA Andaman Is.
Runners *.024 1.000 *<.001 1.000Swimmers *.031 *�.001 *.007Controls *�.001 1.000LSA SA *�.001Andaman Is.
*Significant difference based upon ANOVA with Hochberg’s GT2 or Games Howellpost-hoc test, p � 0.05.LSA SA: Later Stone Age southern Africans, Andaman Is.: Andaman Islanders.
Table 4Slope characteristics for tibial J versus humeral J among athletes, controls and Ho-locene foragers.
Slope Std. error 95% confidence interval
Lower limit Upper limit
Runners 1.515a 0.647 0.221 2.809Swimmers 1.110 0.184 0.742 1.478Controls 1.178 0.457 0.264 2.092Andaman Is. 1.313 0.192 0.929 1.697LSA SA 2.973b 0.353 2.267 3.679
a Slope for runners is significantly steeper compared against swimmers only.b Slope for LSA SA is significantly steeper compared with all other groups.
C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e84
groups. Additionally, the slope of the line that describes runners issignificantly steeper than that which describes swimmers.
Pleistocene fossil hominins
Compared with all other groups, the relative tibial rigidity of theNeandertals and Anatomically Modern H. sapiens (AMHS) are mostsimilar to the LSA southern Africans, while in general the UpperPalaeolithic H. sapiens are more similar to the runners (Table 2). AllNeandertals and AMHS display higher relative tibial rigidity com-pared with runners, while four of 11 of these individuals (Skhul IVand V and La Ferrassie 1 and 2) display relative tibial rigidity (4.63e6.55) that is higher than the LSA southern Africans (4.56). Addi-tionally, unlike all other Upper Palaeolithic individuals, Paviland 1,a European-EUP H. sapiens, displays relative tibial rigidity that isalso exceptionally high (6.10). When Paviland 1 is removed fromthe calculation of relative tibial rigidity for the EUP-EuropeH. sapiens the average decreases from 3.28 to 2.73. Similarly,three of the 20 LUP-North African specimens also display levels ofrelative tibial rigidity that exceed or are comparable with the LSAsouthern African sample: El Wad 10260 (5.54), El Wad 10252 (4.40)and Jebel Sahaba 21 (4.90). Fig. 2, a bivariate plot of raw tibial ri-gidity against raw humeral rigidity, provides a visualization of allHolocene H. sapiens, Neandertals, AMHS, and the four highlyasymmetric Upper Palaeolithic specimens noted above. This plotallows one to evaluate whether tibial or humeral properties aredriving relative tibial rigidity. The higher slope values (Table 4) thatdescribe the LSA southern African sample reflect a greater increase
Figure 1. Relative tibial strength (tibial J/humeral J) for modern human athletes andcontrols, and Holocene foragers. Boxes represent the 25the75th percentile range, withwhiskers extending to the maximum and minimum values within 1.5 box lengths.Outliers are indicated with an o.
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
in tibial rigidity relative to increases in humeral rigidity. The higherrelative tibial rigidity of the eight Pleistocene fossil homininsidentified above indicate that these individuals have even morehypertrophied lower limbs, relative to their upper limbs, comparedwith all other groups, including the LSA southern Africans. Thehumeral rigidity of these Pleistocene hominins is not out of placeamongst modern humans, falling between the small-bodied Ho-locene foragers and the athletes. Their tibial morphology, in con-trast, is at the high end, indicating that the relative tibial rigidity inthe fossil specimens is unique, driven by high tibial rigidity ratherthan unusually weak humeri (however, see the end of theDiscussion section for more on the potential problems associatedwith interpreting raw J values).
Table 5 provides predictions of tibial J for each of the Pleistocenefossil hominins using equations derived from the relationship ofraw tibial J and raw humeral J constructed using the swimmer,runner, LSA southern African, and Andaman Islander samples. Forvirtually all Neandertals and AMHS, tibial rigidity was most accu-rately predicted using the equation calculated from the LSAsouthern African sample. The exception to this trend is that themost accurate predictions were made for La Ferrassie 2, Qafzeh 8and the Upper Palaeolithic H. sapiens using the equation derivedfrom the runner sample. The most accurate predictions of tibialrigidity for Neandertals and Upper Palaeolithic individuals arerelatively close to the actual measured values. By contrast, pre-dictions for the AMHS appear well below the actual measuredvalue.
Figure 2. Bivariate plot of tibial rigidity (J) versus humeral rigidity (J) for modernhuman athletes, Holocene foragers, and Pleistocene hominins.
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004
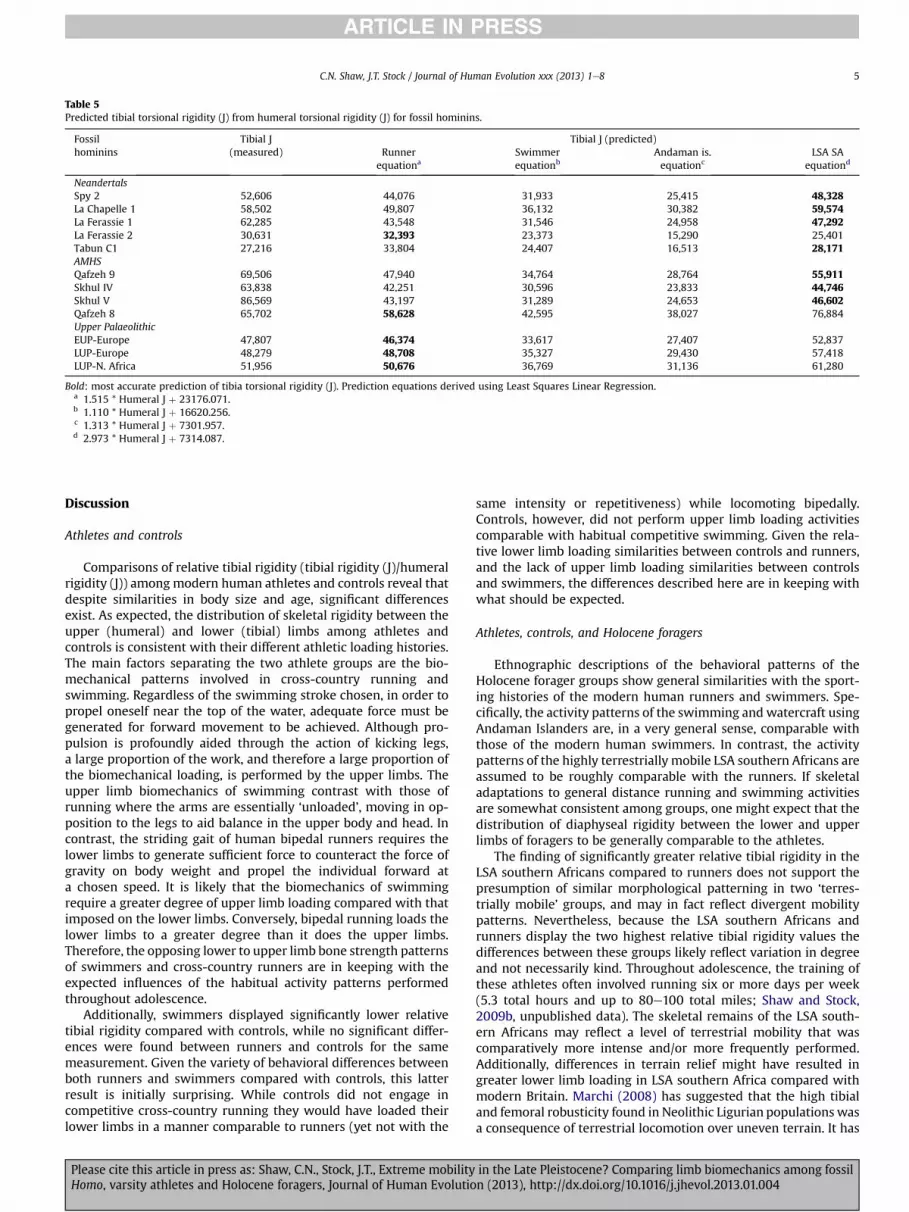
Table 5Predicted tibial torsional rigidity (J) from humeral torsional rigidity (J) for fossil hominins.
Fossilhominins
Tibial J(measured)
Tibial J (predicted)Runner
equationaSwimmerequationb
Andaman is.equationc
LSA SAequationd
NeandertalsSpy 2 52,606 44,076 31,933 25,415 48,328La Chapelle 1 58,502 49,807 36,132 30,382 59,574La Ferassie 1 62,285 43,548 31,546 24,958 47,292La Ferassie 2 30,631 32,393 23,373 15,290 25,401Tabun C1 27,216 33,804 24,407 16,513 28,171AMHSQafzeh 9 69,506 47,940 34,764 28,764 55,911Skhul IV 63,838 42,251 30,596 23,833 44,746Skhul V 86,569 43,197 31,289 24,653 46,602Qafzeh 8 65,702 58,628 42,595 38,027 76,884Upper PalaeolithicEUP-Europe 47,807 46,374 33,617 27,407 52,837LUP-Europe 48,279 48,708 35,327 29,430 57,418LUP-N. Africa 51,956 50,676 36,769 31,136 61,280
Bold: most accurate prediction of tibia torsional rigidity (J). Prediction equations derived using Least Squares Linear Regression.a 1.515 * Humeral J þ 23176.071.b 1.110 * Humeral J þ 16620.256.c 1.313 * Humeral J þ 7301.957.d 2.973 * Humeral J þ 7314.087.
C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e8 5
Discussion
Athletes and controls
Comparisons of relative tibial rigidity (tibial rigidity (J)/humeralrigidity (J)) amongmodern human athletes and controls reveal thatdespite similarities in body size and age, significant differencesexist. As expected, the distribution of skeletal rigidity between theupper (humeral) and lower (tibial) limbs among athletes andcontrols is consistent with their different athletic loading histories.The main factors separating the two athlete groups are the bio-mechanical patterns involved in cross-country running andswimming. Regardless of the swimming stroke chosen, in order topropel oneself near the top of the water, adequate force must begenerated for forward movement to be achieved. Although pro-pulsion is profoundly aided through the action of kicking legs,a large proportion of the work, and therefore a large proportion ofthe biomechanical loading, is performed by the upper limbs. Theupper limb biomechanics of swimming contrast with those ofrunning where the arms are essentially ‘unloaded’, moving in op-position to the legs to aid balance in the upper body and head. Incontrast, the striding gait of human bipedal runners requires thelower limbs to generate sufficient force to counteract the force ofgravity on body weight and propel the individual forward ata chosen speed. It is likely that the biomechanics of swimmingrequire a greater degree of upper limb loading compared with thatimposed on the lower limbs. Conversely, bipedal running loads thelower limbs to a greater degree than it does the upper limbs.Therefore, the opposing lower to upper limb bone strength patternsof swimmers and cross-country runners are in keeping with theexpected influences of the habitual activity patterns performedthroughout adolescence.
Additionally, swimmers displayed significantly lower relativetibial rigidity compared with controls, while no significant differ-ences were found between runners and controls for the samemeasurement. Given the variety of behavioral differences betweenboth runners and swimmers compared with controls, this latterresult is initially surprising. While controls did not engage incompetitive cross-country running they would have loaded theirlower limbs in a manner comparable to runners (yet not with the
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
same intensity or repetitiveness) while locomoting bipedally.Controls, however, did not perform upper limb loading activitiescomparable with habitual competitive swimming. Given the rela-tive lower limb loading similarities between controls and runners,and the lack of upper limb loading similarities between controlsand swimmers, the differences described here are in keeping withwhat should be expected.
Athletes, controls, and Holocene foragers
Ethnographic descriptions of the behavioral patterns of theHolocene forager groups show general similarities with the sport-ing histories of the modern human runners and swimmers. Spe-cifically, the activity patterns of the swimming andwatercraft usingAndaman Islanders are, in a very general sense, comparable withthose of the modern human swimmers. In contrast, the activitypatterns of the highly terrestrially mobile LSA southern Africans areassumed to be roughly comparable with the runners. If skeletaladaptations to general distance running and swimming activitiesare somewhat consistent among groups, one might expect that thedistribution of diaphyseal rigidity between the lower and upperlimbs of foragers to be generally comparable to the athletes.
The finding of significantly greater relative tibial rigidity in theLSA southern Africans compared to runners does not support thepresumption of similar morphological patterning in two ‘terres-trially mobile’ groups, and may in fact reflect divergent mobilitypatterns. Nevertheless, because the LSA southern Africans andrunners display the two highest relative tibial rigidity values thedifferences between these groups likely reflect variation in degreeand not necessarily kind. Throughout adolescence, the training ofthese athletes often involved running six or more days per week(5.3 total hours and up to 80e100 total miles; Shaw and Stock,2009b, unpublished data). The skeletal remains of the LSA south-ern Africans may reflect a level of terrestrial mobility that wascomparatively more intense and/or more frequently performed.Additionally, differences in terrain relief might have resulted ingreater lower limb loading in LSA southern Africa compared withmodern Britain. Marchi (2008) has suggested that the high tibialand femoral robusticity found in Neolithic Ligurian populations wasa consequence of terrestrial locomotion over uneven terrain. It has
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004

C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e86
been shown that tibial strain in humans is significantly higherduring running performed on uneven ground compared withtreadmill running (Milgrom et al., 2003), and also during uphill anddownhill walking and running compared with travel over evensurfaces (Burr et al., 1996). If the age at which runners began seri-ously training (w13 years of age, Shaw and Stock, 2009b) was laterthan when LSA children began undertaking ‘long distance’ bipedaltravel, this could also go some way to explaining differences inrelative tibial rigidity between these groups.
Results comparing swimmers and Andaman Islanders aresimilarly complicated. While the Andamanese have been descri-bed as habitual paddlers and swimmers ethnographic accountssuggest that these people also foraged terrestrially when the op-portunity arose. By contrast, swimmers trained for an average of9.3 hours per week throughout adolescence (Shaw and Stock,2009a). It is therefore not overly surprising that significant dif-ferences were found between these two groups regardless of thefact that they displayed the two lowest relative tibial rigidityvalues.
Athletes, controls, Holocene foragers, and Pleistocene fossil hominins
The relative tibial strength of the Pleistocene fossil hominins isquite high, and in a number of cases far exceeds the upper to lowerlimb asymmetry displayed by all Holocene foragers and athletesincluded in this study. Variation among the fossil hominins and allother groups is evident in Fig. 2, where all Pleistocene individualsfall near or above the line of best fit for the LSA southern Africans,the most asymmetric Holocene population included in this study.La Ferrassie 1 and 2, Skhul IV and V exemplify the exceptionalrelative tibial strength of the Neandertals and Levantine anatomi-cally modern H. sapiens. This patterning was also displayed ina single, atypical, European Early Upper Palaeolithic individual(Paviland 1), and three North African Late Upper Palaeolithic in-dividuals (El Wad 10260 and 10252, and Jebel Sahaba 21). Takentogether, these findings indicate that these individuals are adaptedto quite high levels of lower limb loading (relative to upper limbloading). The hypertrophy of the lower limb may reflect adaptationto very high levels of terrestrial mobility, perhaps considerablygreater than the level of habitual walking or running among highlytrained modern human cross-country runners and highly terres-trially mobile early Holocene foragers. An alternative interpretationis that the Pleistocene hominin skeleton reflects adaptation totravel over highly undulating terrain (Marchi, 2008). However, tosupport this alternative interpretation one would have to arguethat terrain was comparable among the different regions that thesefossil hominins inhabited.
An assessment of whether the more pronounced relative tibialrigidity of the fossil hominins is sex-specific is possible by focusingon the three females in the sample. The relative tibial rigidity ofTabun C1, a female Neandertal, is comparable with that of othermale and female Pleistocene hominins included in the study.However, raw tibial rigidity and raw humeral rigidity for Tabun C1are considerably lower than most other fossil hominins, likelyreflective of a comparatively small body size. The second femaleNeandertal, La Ferrassie 2, displays similarly low raw tibial rigidity,yet particularly low humeral rigidity, similar to the mean for thesmall-bodied LSA foragers. As a result, La Ferrassie 2 displays one ofthe highest relative tibial rigidity scores of all individuals includedin this study. Compared with Tabun C1, the higher relative tibialstrength of La Ferrassie 2 likely indicates variation in the level ofrelative lower limb loading between these twoNeandertals, despitedisplaying comparable estimated body mass (Table 2). Interest-ingly, Qafzeh 9, a female anatomically modern H. sapiens, displaysa level of relative tibial rigidity similar to most other fossil
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
hominins, yet levels of raw tibial and humeral rigidity that exceedsmany of the Pleistocene males.
Lieberman and Shea (1994) proposed mobility pattern variationas one of the behavioral differences between Levantine Neandertalsand early H. sapiens. Using cementum annulation of gazelle teeth,Lieberman (1993, 1998) concluded that early modern humansoccupied sites (Qafzeh Units XVIIeXXIV, Tabun C) on a seasonalbasis while Neandertals occupied sites (Amud B, Kebara IXeXII,Tabun B) on a more continuous basis. Lieberman and Shea (1994;Shea, 1998) concluded that early H. sapiens practiced a circulatingmobility strategy and emphasized intercept hunting while Nean-dertals practiced a radiating strategy and focused on ambushhunting. Other investigators have argued against some of theseassertions (Hovers, 1997; Stutz, 2002). More recent analyses ofMiddle Palaeolithic lithic artifacts have hinted at differences inresidential mobility between these two Middle Palaeolithic pop-ulations (Wallace and Shea, 2006), while zooarchaeological ana-lyses have provided additional evidence of large-game hunting inNeandertals (Niven et al., 2012). Regardless of potential differences,the archaeological and zooarchaeological analyses discussed aboveall concluded that both Levantine Neandertals and AMHS engagedin high levels of terrestrial mobility.
A limitation to the present study is the use of raw cross-sectionaldiaphyseal data. Merely constructing ratios of lower to upper limbproperties, as was done here, may not adequately control for theinfluence of body mass on diaphyseal structure. Variation in rela-tive tibial rigidity appears to be driven primarily by differences inraw tibial rigidity and less so by difference in raw humeral rigidity(Table 2). One might then argue that, rather than reflecting adap-tation to differing mobility patterns, variation in relative tibial ri-gidity primarily reflects the influence of body mass on tibialdiaphysis structure. If correct, this assumes that heavier individualsshould display more rigid tibiae, and a tendency towards higherrelative tibial rigidity. However, using data standardised by bodymass and bone length, as might be prescribed, to calculate relativetibial rigidity is problematic. By standardising both the humerusand the tibia by body mass the effect of standardisation is effec-tively nullified following the subsequent calculation of the relativetibial rigidity ratio. Additionally, not standardising tibial and hu-meral rigidity by biomechanical lengths (squared) (Ruff, 2008b)avoids artificially reducing relative tibial rigidity ratios, due to dif-ferences in limb segment lengths between tibiae and humeri,providing a closer match with locomotor expectations (see Ruff,2000a, 2002; Shaw and Ryan, 2012). In support of the approachimplemented in this study, two observations are relevant. First,because humans are bipedal, it is reasonable to expect that theeffect of body mass on the lower limbs relative to the upper limbswill be at least comparable for all individuals. Second, the fossilhominins that display the highest relative tibial rigidity (La Fer-rassie 2 (66.4 kg), Skhul 4 (69.0 kg), Skhul 5 (68.8 kg) Paviland 1(72.4 kg)) have estimated body masses that are similar to, or below,the body mass average for the living human samples (68.2e74.3 kg). By contrast, two of the three Neandertals and AMHSwith the highest estimated body mass (Spy 2 (85.5 kg), La Chapelle1 (81.1 kg)) display relative tibial rigidities far below the average forthe LSA southern Africans, whose estimated body mass is up toone-third less than those particular fossil hominin individuals.These findings do not discount the potential influence that bodymass may be having on relative tibial rigidity, as calculated here.However, the point is made that despite the influence of body masson tibial diaphysis structure, the distribution of diaphyseal rigiditybetween the upper and lower limbs appears to correspond withstated mobility patterns. Additionally, variation between the LSAsouthern Africans and the Pleistocene sample may be related todifference in body size. If the LSA sample was scaled-up to
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004

C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e8 7
a comparable body mass a positive allometric effect may occurcreating a relative tibial rigidity that is more similar to the fossilhominins. Finally, variation described in this study may also havebeen influenced by inter-specific differences in bone sensitivity tobiomechanical loading (Wallace et al., 2012).
Conclusion
Comparisons of relative tibial rigidity among modern humanathletes (cross-country runners, swimmers) and controls, Holoceneforagers (LSA southern Africans, Andaman Islanders) and Pleisto-cene fossil hominins (Neandertals, anatomically modern and UpperPalaeolithic H. sapiens) allow for three general conclusions:
1. As would be expected from their known activity patterns,runners display quite high levels of tibial rigidity relative tohumeral rigidity. When compared with swimmers and con-trols, this appears to be reflective of skeletal adaptation tohabitual training for competitive cross-country running.
2. Marine-adapted Andaman Islanders and highly terrestriallymobile LSA southern Africans display patterns of relative tibialrigidity closest to the athlete groups with more comparablehabitual activity patterns (swimmers and runners, respec-tively). Nevertheless, significant differences between the LSAsouthern Africans and runners, and swimmers and the Anda-man Islanders, limit how accurately one can infer the durationand intensity of specific activity patterns from skeletal remainsalone.
3. Modern human cross-country athletes reported running up to80e100 miles per week, and the total hunting area of the LSAsouthern Africans may have been as large as 2000e3000square miles. The relative tibial strength of the Neandertalsand Anatomically Modern H. sapiens included in this study aremost similar to the LSA southern Africans, while almost half ofthese individuals display even more pronounced relative tibialrigidity. If relative tibial rigidity reflects adaptation to terrestrialmobility, the results presented here suggest that these Nean-dertals, the Skhul and Qafzeh H. sapiens and a few rare UpperPalaeolithic H. sapiens were walking or running to a far greaterdegree than even highly trained modern athletes and highlyterrestrially mobile early Holocene foragers.
Acknowledgements
Grant sponsorship: Royal Anthropological Institute, UK & Cush-ing Anthropological Research Fund (CNS), The Leverhulme Trustand NERC, UK (JTS). The authors wish to thank Eric Trinkaus andLaura Shackelford for very generously providing much of the fossilhominin data analysed in this manuscript, and members of thePAVE Research Group (University of Cambridge) who providedvaluable intellectual input. Finally, this manuscript was greatlyimproved by the efforts of Editor in Chief David Begun and twoanonymous reviewers.
References
Alciati, G., Pesce Delfino, V., Vacca, E., 2005. Catalogue of Italian fossil human re-mains from the Palaeolithic to the Mesolithic. J. Anthropol. Sci. Suppl. 84, 1e184.
Anderson, J., 1968. Late Paleolithic skeletal remains from Nubia. In: Wendorf, F.(Ed.), The Prehistory of Nubia, vol. 2. Southern Methodist University Press,Dallas, pp. 996e1040.
Auerbach, B., Ruff, C., 2006. Limb bone bilateral asymmetry: variability and com-monality among modern humans. J. Hum. Evol. 50, 203e218.
Bertram, J., Biewener, A., 1988. Bone curvature: sacrificing strength for load pre-dictability? J. Theor. Biol. 131, 75e92.
Bonnet, R., 1913e14. Die ersten vollständigen Skelette der Diluvialmenschender Renntierzeit aus Deutschland, gefunden bei Oberkassel. Jahresbericht
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
der Rheinischen Gesellschaft fur wissenschaftliche Forschung, Bonn 3,28e41.
Boule, M., Vallois, H., Verneau, R., 1934. Anthropologie. In: Arambourg, C. (Ed.), LesGrottes Paleolithiques des Beni-Segoual (Algerie). Archives de l’Institut dePaleontologie Humaine. Masson, Paris, pp. 83e239.
Burr, B., Milgrom, C., Fyhrie, D., Forwood, M., Nyska, M., Finestone, A., Hoshaw, S.,Saiag, E., Simkin, A., 1996. In vivo measurement of human tibial strains duringvigorous activity. Bone 18, 405e410.
Chamla, M., 1978. Le peuplement de l’Afrique du Nord de l’Epipaleolithique a lepoque actuelle. L’Anthropologie 82, 385e430.
Churchill, S.E., Weaver, A.H., Niewoehner, W.A., 1996. Late Pleistocene humantechnological and subsistence behavior: functional interpretations of upperlimb morphology. Quaternaria Nova 6, 413e447.
Churchill, S.E., 1994. Human upper body evolution in the Eurasian Later Pleistocene.Ph.D. Dissertation, University of New Mexico.
Clarke, J., 1959. The Prehisitory of Southern Africa. Penguin Books, London.Currey, J., 1984. The Mechanical Adaptation of Bones. Princeton University Press,
Princeton.Davies, J., 1904. The discovery of human remains under the stalagmite floor of
Gough’s Cave, Cheddar. Q. J. Geol. Soc. Lond. 60, 335e348.Deacon, H., 1993. Planting an idea: an archaeology of stone age gatherers in South
Africa. S. Afr. Archaeol. Bull. 48, 86e93.Delporte, H., 1984. L’Aurignacien de la Ferrassie. Étud. Quatern. 7, 145e234.Greene, D., Armelagos, G., 1972. The Wadi Halfa Mesolithic Population. Research
Report No. 11. Department of Anthropology, University of Massachusetts,Amherst.
Grün, R., Stringer, C., 2000. Tabun revisited: revised ESR chronology and new ESRand U-series analyses of dental material from Tabun C1. J. Hum. Evol. 39,601e612.
Holt, B.M., Formicola, V., 2008. Hunters of the ice age: the biology of Upper Pale-olithic people. Yearb. Phys. Anthropol. 137, 70e99.
Holt, B.M., 2003. Mobility in Upper Paleolithic and Mesolithic Europe: evidencefrom the lower limb. Am. J. Phys. Anthropol. 122, 200e215.
Hovers, E., 1997. Variability of Levantine Mousterian assemblages and settlementpatterns: implications for understanding the development of human behaviour.Ph.D. Dissertation, Hebrew University.
Jacobi, R.M., Higham, T.F.G., 2008. The ‘Red Lady’ ages gracefully: new ultrafiltrationAMS determinations from Paviland. J. Hum. Evol. 55, 898e907.
Lanyon, L.E., Rubin, C., 1984. Static vs. dynamic loads as an influence on boneremodeling. J. Biomech. 17, 897e905.
Lanyon, L.E., 1992. Control of bone architecture by functional load bearing. J. BoneMiner. Res. 7, S369eS375.
Larsen, C.S., Ruff, C.B., Kelly, R., 1995. Structural analysis of the stillwater postcranialremains: behavioural implications of articular joint pathology and long bonediaphyseal morphology. In: Larsen, C., Kelly, R. (Eds.), Bioarchaeology of theStillwater Marsh: Prehistoric Human Adaptation in the Western Great Basin.University of Florida Press, Gainsville.
Lieberman, D.E., Shea, J.J., 1994. Behavioral differences between archaic and modernhumans in the Levantine Mousterian. Am. Anthropol. 96, 300e332.
Lieberman, D.E., 1993. Mobility and strain: the biology of cementogenesis and itsapplication to the evolution of hunter-gatherer seasonal mobility duringthe Late Quaternary in the Southern Levant. Ph.D. Dissertation, HarvardUniversity.
Lieberman, D.E., 1998. Neandertal and early modern human mobility patterns:comparing archaeological and anatomical evidence. In: Akazawa, T. (Ed.), Ne-andertals and Modern Humans in Western Asia. Plenum Press, New York,pp. 263e276.
Mallegni, F., Fabbri, P.F., 1995. The human skeletal remains from the Upper Palae-olithic burials found in Romito cave (Papasidero, Cosenza, Italy). Bull. Mem. Soc.Anthropol. Paris 7, 99e137.
Man, E., 1883. On the aboriginal inhabitants of the Andaman Islands. J. R. Anthropol.Inst. 12, 69e434.
Marchi, D., 2008. Relationships between lower limb cross-sectional geometry andmobility: the case of a Neolithic sample from Italy. Am. J. Phys. Anthropol. 137,188e200.
Martin, R.B., Burr, D.R., 1989. Structure, Function, and Adaptation of Compact Bone.Raven Press, New York.
McCown, T., 1939. The Natufian crania from Mount Carmel, Palestine, and theirinterrelationships. Ph.D. Dissertation, University of California, Berkeley.
McDermott, F., Grün, R., Stringer, C.B., Hawkesworth, C.J., 1993. Mass-spectrometricU-series dates for Israeli Neanderthal/early modern hominid sites. Nature 363,252e255.
Milgrom, C., Finestone, A., Segev, S., Olin, C., Arndt, T., Ekenman, I., 2003. Areoverground or treadmill runners more likely to sustain tibial stress fracture? Br.J. Sport Med. 37, 160e163.
Myka, F., 1993. Decline of Indigenous Populations: The Case of the Andaman Is-landers. Rawat Publications, Jaipur.
Narr, K., 1977. Das Rätsel von Neuessing Bemerkungen zu dem Skelettfund aus derMittleren Klause. In: Schröter, P. (Ed.), 75 Jahre Anthropologishe Staatssamm-lung München. Selbstverlag der Anthropologishen Stattssammlung, München,pp. 53e56.
Niven, L., Steele, T., Rendu, W., Mallye, J., McPherron, S., Soressi, M., Jaubert, J.,Hublin, J., 2012. Neandertal mobility and large-game hunting: the exploitationof reindeer during the Quina Mousterian at Chez-Pinaud Jonzac (Charente-Maritime, France). J. Hum. Evol. 63, 624e635.
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004

C.N. Shaw, J.T. Stock / Journal of Human Evolution xxx (2013) 1e88
Pearson, O.M., Lieberman, D.E., 2004. The aging of Wolff’s ‘law’: ontogeny andresponses to mechanical loading in cortical bone. Yearb. Phys. Anthropol. 47,63e99.
Pittard, E., Sauter, M.R., 1945. Un squelette magdalénien provenant de la station desGrenouilles (Veyrier, Haute-Savoie). Arch. Suisses Anthropol. Gén. Geneva 11,149e200.
Raynal, J., 1990. Essai de datation directe. In: Raynal, J.P., Pautrat, Y. (Eds.), LaChapelle-aux-Saints et la préhistoire en Corrèze. Association pour la RechercheArchéologique en Limousin, Limousine.
Rubin, C., McLeod, K., Basin, S., 1990. Functional strains and cortical bone adapta-tion: epigenetic assurance of skeletal integrity. J. Biomech. 23 (Suppl. 1), 43e54.
Ruff, C.B., 2000a. Body size, body shape and long bone strength in modern humans.J. Hum. Evol. 38, 269e290.
Ruff, C.B., 2000b. Body mass prediction from skeletal frame size in elite athletes.Am. J. Phys. Anthropol. 112, 507e517.
Ruff, C.B., 2002. Long bone articular and diaphyseal structure in Old World monkeysand apes. I: locomotor effects. Am. J. Phys. Anthropol. 119, 305e342.
Ruff, C.B., 2008a. Femoral/humeral strength in early African Homo erectus. J. Hum.Evol. 54, 383e390.
Ruff, C.B., 2008b. Biomechanical analyses of archaeological human skeletons. In:Katzenberg, M., Saunders, A. (Eds.), Biological Anthropology of the HumanSkeleton. John Wiley and Sons Inc., Hoboken, pp. 183e206.
Ruff, C.B., 2009. Relative limb strength and locomotion in Homo habilis. Am. J. Phys.Anthropol. 138, 90e100.
Ruff, C.B., Trinkaus, E., Walker, A., Larsen, C.S., 1993. Postcranial robusticity in Homo.I: temporal trends and mechanical interpretation. Am. J. Phys. Anthropol. 91,21e53.
Ruff, C.B., Trinkaus, E., Holliday, T.W., 1998. Body mass estimation in Olympic ath-letes and Pleistocene Homo. Am. J. Phys. Anthropol. Suppl. 26, 192e193.
Ruff, C.B., Holt, B.M., Trinkaus, E., 2006. Who’s afraid of the big bad Wolff? ‘Wolff’sLaw’ and bone functional adaptation. Am. J. Phys. Anthropol. 129, 484e498.
Seligman, C., Parsons, F., 1914. The Cheddar Man: a skeleton of late Palaeolithic date.J. R. Anthropol. Inst. 44, 241e263.
Semal, P., Rougier, H., Crevecoeur, I., Jungels, C., Flas, D., Hauzeur, A., Maureille, B.,Germonpré, M., Bocherens, H., Pirson, S., Cammaert, L., De Clerck, L.,Hambucken, A., Higham, T., Toussaint, M., van der Plicht, J., 2009. New data onthe late Neandertals: direct dating of the Belgian Spy fossils. Am. J. Phys.Anthropol. 138, 421e428.
Shackelford, L.L., 2007. Regional variation in the postcranial robusticity of late UpperPaleolithic humans. Am. J. Phys. Anthropol. 133, 655e668.
Shaw, C., Ryan, T., 2012. Does skeletal anatomy reflect adaptation to locomotorpatterns? Cortical and trabecular architecture in human and nonhuman an-thropoids. Am. J. Phys. Anthropol. 147, 187e200.
Shaw, C., Stock, J., 2009a. Habitual throwing and swimming correspond with upperlimb diaphyseal strength and shape in modern human athletes. Am. J. Phys.Anthropol. 140, 160e172.
Shaw, C., Stock, J., 2009b. Intensity, repetitiveness, and directionality of habitualadolescent mobility patterns influence the tibial diaphysis morphology ofathletes. Am. J. Phys. Anthropol. 140, 149e159.
Shaw, C., 2010. ‘Putting flesh back onto the bones?’ Can we predict soft tissueproperties from skeletal and fossil remains? J. Hum. Evol. 59, 484e492.
Shaw, C., 2011. Is ‘hand preference’ coded in the hominin skeleton? An in-vivo studyof bilateral morphological variation. J. Hum. Evol. 61, 480e487.
Please cite this article in press as: Shaw, C.N., Stock, J.T., Extreme mobilityHomo, varsity athletes and Holocene foragers, Journal of Human Evolutio
Shea, J.J., 1998. Neandertal and early modern human behavioural variability. Curr.Anthropol. 39, S45eS78.
Sladek, V., Trinkaus, E., Hillson, S.W., Holliday, T.W., 2000. The People of the Pav-lovian: Skeletal Catalogue and Osteometrics of the Gravettian Fossil Hominidsfrom Dolní Vĕstonice and Pavlov. In: Dolní Vĕstonice Studies, vol. 5. Akademievĕd �Ceske republicky, Brno.
Stock, J., Pfeiffer, S., 2001. Linking structural variability in long bone diaphyses tohabitual behaviors: foragers from the southern African Later Stone Age and theAndaman Islands. Am. J. Phys. Anthropol. 115, 337e348.
Stock, J.T., Pfeiffer, S., 2004. Long bone robusticity and subsistence behavior amongLater Stone Age foragers of the forest and fynbos biomes of South Africa.J. Archaeol. Sci. 31, 999e1013.
Stock, J., 2006. Hunter-gatherer postcranial robusticity relative to patterns ofmobility, climatic adaptation, and selection for tissue economy. Am. J. Phys.Anthropol. 131, 194e204.
Stutz, A., 2002. Polarizing microscopy identification of chemical diagenesis inarchaeological cementum. J. Archaeol. Sci. 29, 1327e1347.
Svoboda, J., 1995. L’art gravettien en Moravie: contexte dates et styles. L’An-thropologie 99, 258e272.
Trinkaus, E., Churchill, S.E., 1999. Diaphyseal cross-sectional geometry of NearEastern Middle Palaeolithic humans: the humerus. J. Archaeol. Sci. 26, 173e184.
Trinkaus, E., Ruff, C.B., 1989. Diaphyseal cross-sectional morphology and bio-mechanics of the Fond-de-Foret 1 Femur and the Spy 2 Femur and tibia. Bull.Soc. R. Belge Anthropol. Prehist. 100, 33e42.
Trinkaus, E., Ruff, C.B., 1999a. Diaphyseal cross-sectional geometry of Near EasternMiddle Palaeolithic humans: the femur. J. Archaeol. Sci. 26, 409e424.
Trinkaus, E., Ruff, C.B., 1999b. Diaphyseal cross-sectional geometry of Near EasternMiddle Palaeolithic humans: the tibia. J. Archaeol. Sci. 26, 1289e1300.
Trinkaus, E., Ruff, C.B., 2012. Femoral and tibial diaphyseal cross-sectional geometryin Pleistocene Homo. Paleoanthropology 2012, 13e62.
Trinkaus, E., Svoboda, J., 2006. Early Modern Human Evolution in Central Europe:The People of Dolní V�estonice and Pavlov. Oxford University Press, Oxford.
Trinkaus, E., Churchill, S.E., Ruff, C.B., 1994. Postcranial robusticity in Homo. II:humeral bilateral asymmetry and bone plasticity. Am. J. Phys. Anthropol. 93,1e34.
Trinkaus, E., 1989. The Upper Pleistocene transition. In: Trinkaus, E. (Ed.), The Emer-gence of Modern Humans. Cambridge University Press, Cambridge, pp. 42e46.
Trinkaus, E., 1997. Appendicular robusticity and the paleobiology of modern humanemergence. Proc. Natl. Acad. Sci. 94, 13367e13373.
Trinkaus, E., 2000. The ‘robusticity transition’ revisited. In: Stringer, C.B., Finlayson, C.(Eds.), Gibraltar and the Neanderthals. Oxbow Press, Oxford, pp. 227e236.
Verneau, R., 1906. Les Grottes de Grimaldi (Baouss, Rouss) III: Anthropologie.Imprimerie de Monaco, Monaco.
Verworn, M., Bonnet, R., Steinmann, G., 1919. Der diluviale Menschenfund vonOberkassel bei Bonn. Verlag von J.F. Bergmann, Wiesbaden.
Wallace, I., Shea, J.J., 2006. Mobility patterns and core technologies in the MiddlePaleolithic of the Levant. J. Archaeol. Sci. 33, 1293e1309.
Wallace, I., Tommasini, S., Judex, S., Garland, T., Demes, B., 2012. Genetic variationsand physical activity as determinants of limb bone morphology: an exper-imental approach using a mouse model. Am. J. Phys. Anthropol. 148, 1e12.
Wanner, I., Sierra-Sosa, T., Alt, K., Blos, V., 2007. Lifestyle, occupation, and wholebone morphology of the pre-hispanic Maya coastal population from Xcambo,Yucatan, Mexico. Int. J. Osteoarchaeol. 17, 253e268.
in the Late Pleistocene? Comparing limb biomechanics among fossiln (2013), http://dx.doi.org/10.1016/j.jhevol.2013.01.004





























