Embed Size (px)
Citation preview

Accepted Manuscript
Field and experimental evidence of a new caiman trypanosome species closelyphylogenetically related to fish trypanosomes and transmitted by leeches
Bruno R. Fermino, Fernando Paiva, Priscilla Soares, Luiz Eduardo R. Tavares,Laerte B. Viola, Robson C. Ferreira, Robinson Botero-Arias, Cátia D. de-Paula, MartaCampaner, Carmen S.A. Takata, Marta M.G. Teixeira, Erney P. Camargo
PII: S2213-2244(15)30019-5
DOI: 10.1016/j.ijppaw.2015.10.005
Reference: IJPPAW 134
To appear in: International Journal for Parasitology: Parasites and Wildlife
Received Date: 28 July 2015
Revised Date: 25 September 2015
Accepted Date: 16 October 2015
Please cite this article as: Fermino, B.R., Paiva, F., Soares, P., Tavares, L.E.R., Viola, L.B., Ferreira,R.C., Botero-Arias, R., de-Paula, C.D., Campaner, M., Takata, C.S.A., Teixeira, M.M.G., Camargo, E.P.,Field and experimental evidence of a new caiman trypanosome species closely phylogenetically relatedto fish trypanosomes and transmitted by leeches, International Journal for Parasitology: Parasites andWildlife (2015), doi: 10.1016/j.ijppaw.2015.10.005.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Blood
Leech

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT1
Field and experimental evidence of a new caiman trypanosome species closely phylogenetically 1
related to fish trypanosomes and transmitted by leeches 2
3
Bruno R. Fermino1, Fernando Paiva2, Priscilla Soares2, Luiz Eduardo R. Tavares2, Laerte B. Viola1, Robson C. 4
Ferreira1, Robinson Botero-Arias3, Cátia D. de-Paula4, Marta Campaner1, Carmen S. A. Takata1, Marta M. G. 5
Teixeira1*, Erney P. Camargo 1 6
7
1. Departamento de Parasitologia, Instituto de Ciências Biomédicas, Universidade de São Paulo, São Paulo, 8
Brasil. 9
2. Centro de Ciências Biológicas e da Saúde, Universidade Federal do Mato Grosso do Sul, Campo Grande, 10
Mato Grosso do Sul, Brasil. 11
3. Caiman Research in Conservation and Management Program, Instituto Mamirauá para o Desenvolvimento 12
Sustentável. 13
Tefé, Amazonas, Brasil. 14
4. Faculdade de Agronomia e Medicina Veterinária, Universidade de Brasília, Brasília, DF, Brasil. 15
16
*Corresponding author: [email protected] 17
18
ABSTRACT 19
Trypanosoma terena and Trypanosoma ralphi are known species of the South American crocodilians Caiman 20
crocodilus, Caiman yacare and Melanosuchus niger and are phylogenetically related to the tsetse-transmitted 21
T. grayi of the African Crocodylus niloticus. These trypanosomes form the Crocodilian clade of the terrestrial 22
clade of the genus Trypanosoma. A PCR-survey for trypanosomes in caiman blood samples and in leeches 23
taken from caimans revealed unknown trypanosome diversity and frequent mixed infections. Phylogenies 24
based on SSU (small subunit) of rRNA and gGAPDH (glycosomal Glyceraldehyde Phosphate Dehydrogenase) 25
gene sequences revealed a new trypanosome species clustering with T. terena and T. ralphi in the crocodilian 26
clade and an additional new species nesting in the distant Aquatic clade of trypanosomes, which is herein 27
named Trypanosoma clandestinus n. sp. This new species was found in Ca. yacare, Ca. crocodilus and M. 28
niger from the Pantanal and Amazonian biomes in Brazil. Large numbers of dividing epimastigotes and unique 29
thin and long trypomastigotes were found in the guts of leeches (Haementeria sp.) removed from the mouths 30
of caimans. The trypanosomes recovered from the leeches had sequences identical to those of T. 31
clandestinus of caiman blood samples. Experimental infestation of young caimans (Ca. yacare) with infected 32
leeches resulted in long-lasting T. clandestinus infections that permitted us to delineate its life cycle. In 33
contrast to T. terena, T. ralphi and T. grayi, which are detectable by hemoculturing, microscopy and standard 34
PCR of caiman blood, T. clandestinus passes undetected by these methods due to very low parasitemia and 35

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT2
could be detected solely by the more sensitive nested PCR method. T. clandestinus n. sp. is the first 1
crocodilian trypanosome known to be transmitted by leeches and positioned in the aquatic clade closest to fish 2
trypanosomes. Our data show that caimans can host trypanosomes of the aquatic or terrestrial clade, 3
sometimes simultaneously. 4
5
Keywords: Crocodilian, Trypanosoma, reptilian parasites, phylogeny, phylogeography, leeches, co-infection, 6
South America. 7
8
1. Introduction 9
10
Flagellates of the genus Trypanosoma (Euglenozoa, Kinetoplastea, Trypanosomatidae) are obligate parasites 11
of all vertebrate classes and are distributed into two major phylogenetic lineages: the Terrestrial clade 12
composed of trypanosomes of mammals, snakes, lizards, crocodilians and birds and the Aquatic clade 13
containing trypanosomes of aquatic leeches and aquatic (fishes) or semi-aquatic (chelonians, anurans and 14
platypus) hosts and, oddly, a trypanosome of chameleon (Stevens et al., 2001; Hamilton et al., 2005, 2007). 15
Trypanosomes infect Neotropical and Afrotropical crocodilians (Telford, 1995, 2009; Fermino et al. 2013). 16
Crocodilians of the family Alligatoridae predominate in the New World and are distributed in the genera 17
Caiman, Paleosuchus and Melanosuchus, generically called caimans. The Afrotropical Crocodylidae 18
originated in Australasia and after transoceanic dispersal (Oaks, 2011) flourished in South America (Scheyer 19
et al. 2013). In the American continent, Alligatoridae and Crocodylidae encountered plenty of opportunities of 20
trypanosome switching in the wide connected and disconnected wetlands shaping the present day 21
hydrographic basins (Fermino et al. 2013). 22
Although there are many reports of the occurrence of trypanosomes in reptilians (revised by Telford, 1995, 23
2009), only three species from crocodilians were molecularly characterized: T. grayi (Hoare 1929, 1931), and 24
T. ralphi and T. terena (Fermino et al., 2013). The reported hosts of these trypanosomes are C. niloticus and 25
O. tetraspis in Africa and the caimans Ca. crocodilus, Ca. yacare and M. niger in South America. Available 26
phylogenetic data show very close phylogenetic relationships between the African T. grayi from C. niloticus 27
and the Brazilian T. ralphi from caimans, as well as between the Brazilian T. terena from caimans and an 28
unnamed African trypanosome from O. tetraspis. These South American and African crocodilian trypanosomes 29
form the major Crocodilian clade comprising the clades Terena, Ralphi and Grayi; each clade harbors a 30
number of closely related genotypes (Fermino et al., 2013). The phylogenetic relationships between the 31
crocodilian trypanosomes concur with the worldwide transoceanic dispersal of Crocodylus during the Miocene 32
that ended in either South America or Africa (Oaks, 2011). 33

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT3
Trypanosomes of the three species discovered in crocodilians to date were detected by culturing, standard 1
PCR and microscopic examination of peripheral blood. However, these methods may fail to detect tissue-2
dwelling or scarce blood flagellates and culturing could select against trypanosomes that are fastidious or 3
refractory to cultivation. The alligatorid trypanosomes present varied morphology in the blood and tissues of 4
their hosts that could be the result of the inherent polymorphism of trypanosomes or concomitant infections 5
with more than one species (Viola et al., 2008a; Fermino et al., 2013). The alleged polymorphisms of blood 6
trypanosomes may also result from the occurrence of mixed infections that have been disclosed by molecular 7
analyses of parasites directly from blood samples of anurans, fishes and mammals (Ferreira et al., 2007; Gu et 8
al., 2007; Grybchuk-Ieremenko et al., 2014; Thompson et al., 2013). In addition, molecular studies revealed a 9
number of trypanosomes from wildlife displaying blood forms that are morphologically indistinguishable 10
(Ferreira et al., 2007, 2008; Viola et al., 2008a,b, 2009; Cavazzana et al., 2010; Fermino et al., 2013). These 11
facts have prevented the correct appraisal of the whole assemblage of trypanosomes and the extent of their 12
diversity and host and geographical ranges. 13
Nothing is known about the vectors of trypanosomes of South American caimans. Their close relative T. 14
grayi is transmitted to African crocodiles by tsetse flies. A range of hematophagous flies serve as vectors of 15
trypanosomes of reptiles, including tsetse flies for crocodiles (Hoare, 1929, 1931) and sand flies for lizards 16
(Ayala, 1971; Ayala and McKay, 1971; Christensen and Telford, 1972; Viola et al., 2008b). Leeches are known 17
to transmit trypanosomes of aquatic and semi-aquatic animals including fishes (Letch, 1977; Karlsbaak et al., 18
2005; Khan, 1976; Hayes et al., 2014), anurans (Martin and Desser, 1991; Siddall and Desser, 1992), snakes 19
(Pessoa and Fleury, 1969; Chia and Miller, 1984), turtles (Woo, 1969; Siddall and Desser, 1992) and platypus 20
(Paparini et al., 2014). 21
Trypanosomes of anurans (Ayala, 1971; Martin & Desser, 1991; Siddall & Desser, 1992; Ferreira et al., 22
2008) and serpents (Chia and Miller, 1984; Viola et al., 2008b) are transmitted by leeches in aquatic 23
environments and by insects such as sand flies and culicids in more terrestrial niches. Moreover, a survey 24
performed using nested PCR suggested that terrestrial leeches could transmit trypanosomes of toads and 25
marsupials in Australia (Hamilton et al., 2005). Therefore, flies and leeches could transmit the trypanosomes of 26
the semiaquatic caimans, although no vectors have been identified to date. 27
In the present study, we surveyed for trypanosomes in blood and tissues samples from 122 specimens of 28
South American caimans and the guts of 208 leeches collected from the mouths of caimans. Field surveys, 29
morphological and experimental infections of caimans and phylogenetic analyses were employed to 30
investigate life cycles, species diversity and phylogenetic relationships of crocodilians trypanosomes. 31
32
2. Materials and methods 33
34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT4
2.1. Collection sites, caiman handling and blood sampling 1
Caimans were captured in the Amazonian (AM), Araguaia-Tocantins (TO), Paraguay-Paraná (PP) and Orinoco 2
(OR) river basins. The collection sites are shown in the Figure 1 and Table 1. The capture of animals and all 3
ensuing procedures were conducted as described previously (Fermino et al., 2013) according to the 4
recommendations of IBAMA (the Brazilian Institute for the Environment and Renewable Natural Resources), 5
permit Number 10080-2 and the protocol (108/2013) approved by the Committee on the Ethics of Animal 6
Experimentation of the Institute of Biomedical Sciences, University of São Paulo. 7
Caiman blood samples and liver tissue preserved in ethanol (v/v) were stored in the Blood Sample 8
Collection (BSC) and Tissue Sample Collection (TSC) of the Trypanosomatid Culture Collection of the 9
University of São Paulo (TCC – USP). Total DNA was extracted from blood and tissue samples as previously 10
described using the traditional phenol-chloroform method. DNA samples from all trypanosome-positive 11
crocodilian samples were also preserved in the TCC. 12
13
2.2. Collection and identification of leeches and survey for trypanosomes 14
Leeches were collected from the mouths of the caimans, and identified as Haementeria sp. by COI barcodes 15
(Folmer et al., 1994). Leeches were immediately preserved in ethanol (v/v) or brought to the laboratory and 16
kept for 15-20 days in plain water until they completely digested the ingested blood meal and became almost 17
transparent. Then, the guts of the leeches were examined by microscopy. Leeches positive for trypanosomes 18
were used for smears on glass slides, inoculation into culture tubes and DNA preparations using the phenol-19
chloroform method. 20
21
2.3. PCR amplification and phylogenetic analyses of SSU rRNA and gGAPDH gene sequences 22
The nested-PCR of SSU rRNA sequences (~987 bp) of trypanosomes from crocodilian blood samples was 23
performed as described previously (Noyes et al., 1999). PCR amplification of the V7V8 region of the SSU 24
rRNA and gGAPDH sequences from leech trypanosomes was performed as previously described (Borghesan 25
et al., 2013). The nested PCR amplification of gGAPDH sequences was performed using the first round 26
primers (GAPDH SF) 5’ GTG GCG GTK GTY GAC ATG AAC A3’ and (GAPDH SR) 5’ TTG GAG TCR TAG 27
ATR GAG CT3’, and the second round internal primers (GAP 3F) 5’ GTG AAG GCG CAG CGC AAC 3’ and 28
(GAP 5R) 5’ CCG AGG ATG YCC TTC ATG 3’. The two rounds of amplification (30 cycles each) were 29
performed using PCR reaction mixtures and conditions described previously (Borghesan et al., 2013). 30
Amplified DNA were cloned, and 10-20 clones were sequenced for each gene from each trypanosome 31
sample. Sequencing of cloned amplified DNA allowed for the detection of mixed trypanosome sequences in a 32
single blood sample (Fermino et al., 2013). The SSU rRNA and gGAPDH sequences determined in this study 33
were deposited in GenBank under the accession numbers shown in Table 1. 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT5
For phylogenetic inferences using Parsimony (P), Maximum Likelihood (ML) and Bayesian inference (BI) 1
analyses, sequences were aligned using CLUSTALX (Thompson et al., 1997) and manually refined. Three 2
alignments were created: A1, consisting of the V7V8 region of the SSU rRNA gene (687 characters); A2, 3
containing the gGAPDH sequences (608 characters); and A3, consisting of concatenated SSU rRNA and 4
gGAPDH sequences (1.295 characters) using previous alignments including a large set of taxa for guidance 5
(Hamilton et al., 2004). The parsimony and bootstrap analyses were performed using PAUP* version 4.0b10 6
software (Swofford, 2002) with 500 random sequence addition replicates followed by branch swapping (RAS-7
TBR). The ML analyses were performed using RAxML v.2.2.3 (Stamatakis, 2006) as described previously 8
(Ferreira et al., 2008; Borghesan et al., 2013). The general time reversible (GTR) model of nucleotide 9
substitution with a proportion of invariable sites and gamma distribution was selected for the gene data sets. 10
The GTR model was used in individual analysis of each gene as well as in the combined Bayesian analysis. 11
The analysis was run for 1,000,000 generations with trees sampled every 100 generations using four chains 12
and 25% of the early sample trees discarded as “burn-in”. 13
14
2.4. Experimental infection of caimans 15
Five farm-born Ca. yacare 8-10 months old and weighing ~260 g were kept indoors at 27 °C in large glass 16
tanks of water with a land platform. Blood samples were collected upon the arrival of the animals and for two 17
months at ~15 days intervals to evaluate by nested PCR whether the animals were free of trypanosomes prior 18
to experimental infection. All procedures during the experimental period were performed in accordance with 19
the protocol (#36781-1) approved by the IBAMA and the “Sistema de Autorização e Informação em 20
Biodiversidade" (SISBIO). 21
Approximately 80 leeches were collected from wild Ca. yacare captured in the Miranda river at the PP 22
basin and maintained for 15 days in water at room temperature in the laboratory. Five randomly selected 23
leeches were examined and found to be heavily infected with trypanosomes. The gut contents from these 24
leeches were preserved in ethanol for DNA amplification or smeared onto glass slides for microscopy. 25
For experimental infections, two caimans had 30 leeches each deposited in their mouths and two additional 26
animals were fed with 30 leeches macerated with meat. One caiman received no leeches and was used as the 27
negative control. Starting on the 7th day after infection, ~0.5 ml blood samples were collected every ~15 days 28
for 5 months by caudal venous puncturing. A final blood sample was collected 8 months after the inoculation. 29
Blood samples from each animal were either preserved in ethanol for DNA analysis or immediately inoculated 30
into culture medium as described previously (Fermino et al., 2013) and for the preparation of blood and 31
microhematocrit buffy coat smears on glass slides. The slides were fixed with methanol, stained with Giemsa 32
and examined microscopically. To monitor the experimental infection, DNA preparations from caiman blood 33

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT6
were subjected to nested-PCR of V7V8 SSU rRNA and gGAPDH sequences. The amplified products were 1
cloned and 10-20 clones from each sample were sequenced. 2
3
3. Results 4
5
3.1. New species disclosed by barcoding of caiman and leech trypanosomes using SSU rRNA and 6
gGAPDH sequences 7
Preliminary tests showed that the nested PCR methods targeting both SSU rRNA and gGAPDH 8
sequences could detect trypanosomes in the blood of caimans that tested positive or negative by blood 9
microscopic examination and hemoculturing. Then, we examined 122 caiman blood samples and found 10
trypanosome sequences in 40 caimans, yielding a prevalence of ~33%. 11
The analyses of V7V8 SSU rRNA and gGAPDH trypanosome sequences obtained from blood or tissue 12
samples from 34 caimans revealed that 23 (68%) of the caimans presented sequences from a single species, 13
whereas 11 (32%) presented mixed infections with two to four trypanosome species; T. terena and T. ralphi 14
were detected in ~ 50% of the mixed infections. Sequences corresponding to two new trypanosome species 15
were detected exclusively using the nested-PCR methods. Samples sharing a new sequence formed a clade 16
(Cay03) that nested together with T. terena and T. ralphi in the previously described Crocodilian Clade 17
(Fermino et al., 2013); the other new sequences formed the new Clandestinus clade, which were positioned 18
within the Aquatic clade of Trypanosoma (Table 1, Fig. 1). 19
Leeches have not been reported to be vectors of crocodilian trypanosomes, although they are very 20
common and abundant ectoparasites and vectors of other crocodilian parasites (Glassman et al., 1979; Khan, 21
1980; Leslie et al., 2011). We surveyed for trypanosomes in a sample of 208 leeches randomly collected from 22
the mouths of Ca. yacare and Ca. crocodilus; a total of 120 leeches (~58%) were found to be positive for 23
trypanosomes by microscopy. The number of leeches (TSC codes) from which trypanosomes were sequenced 24
are listed in Table 1. 25
The leeches taken from Ca. yacare from The Pantanal were identified as Haementeria sp. by BLAST 26
analyses of COI sequences (Genbank accession number: leech TSC64Le = KP972452). However, although a 27
single sequence was obtained from 10 randomly selected leeches, the sequences obtained did not completely 28
match any sequence in GenBank, suggesting that they may represent a leech species still lacking a DNA 29
barcode. The leech sequence determined in this study diverged 17% from the closest relative COI sequences 30
from Haementeria officinalis (Genbank accession number: JN850907). 31
Fresh gut samples of leeches removed from Ca. yacare exhibited intense flagellate proliferation. These 32
flagellates always failed to grow when inoculated into the different culture media used to culture a range of 33
trypanosomes including those from the clade Crocodilian (Viola et al., 2008a; Fermino et al., 2013) and 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT7
anurans (Ferreira et al., 2007). Sequences of V7V8 SSU rRNA and gGAPDH identical to those of the 1
Clandestinus clade from caiman blood samples were the only ones detected in all leeches examined (Table 1, 2
Figs 1 and 2). 3
4
3.2. Phylogenetic positioning of caiman and leech trypanosomes within the Aquatic clade 5
In the dendrogram inferred using V7V8 SSU rRNA sequences from trypanosomes of caimans and leeches 6
aligned with sequences of other trypanosome species, the crocodilian trypanosomes of the terrestrial clades 7
Terena, Ralphi and Grayi (Fermino et al., 2013) plus the new clade Cay03 were separated from the 8
Clandestinus clade, which included sequences from trypanosomes allied to fish trypanosomes (Fig. 1). The 9
trypanosomes of the Crocodilian and Clandestinus clades were separated by ~5.0% V7V8 SSU rRNA 10
divergence. 11
Inferred phylogenetic trees using gGAPDH sequences (Fig. 2) and concatenated V7V8 SSU rRNA and 12
gGAPDH sequences (Supplementary Material) strongly supported the distribution of the caiman trypanosomes 13
in the Terrestrial and Aquatic clades of Trypanosoma. The trypanosomes of the terrestrial Crocodilian clade, 14
comprising T. terena, T. grayi and T. ralphi plus the new species provisionally referred to as Cay03, clustered 15
with the trypanosomes from lizards, snakes, birds and mammals. The trypanosomes identified in the leeches 16
always clustered with the caiman trypanosomes of clade Clandestinus, comprising highly homogeneous 17
sequences that were strongly supported within the Aquatic clade (Fig. 2). These results enabled the 18
description of the new species Trypanosoma clandestinus n. sp. in caimans and leeches. 19
The divergence of gGAPDH sequences between the five crocodilian trypanosome species ranged from 20
6.0% to 14%.The largest divergence separated T. grayi from T. clandestinus, and the shortest divergence 21
separated T. ralphi from Cay03. T. clandestinus diverged 11.7%, 12.5% and 11% from T. terena, T. ralphi and 22
Cay03, respectively. The divergence of gGAPDH sequences within each species varied from 0.5% in T. 23
clandestinus to 2.1% in the more heterogeneous clade T. grayi. All gGAPDH sequences obtained from 24
leeches (TSC 4-10 and TSC 62-65) were identical to the sequence of T. clandestinus (Table 1; Figs. 1, 2). 25
Not a single trypanosome sequence of the Crocodilian clade was recovered from leeches. Moreover, T. 26
clandestinus was phylogenetically more related to trypanosomes of fishes and placed closest to trypanosomes 27
from Brazilian freshwater fishes than to any other reptilian trypanosome from the aquatic (T. chelodinae) or 28
terrestrial clade (T. serpentis, T. cascavelli, T. varani and T. sp. Gecko) (Fig. 1). The terrestrial crocodilian 29
clade was sister to the main clade comprising trypanosomes from snakes, birds, lizards and mammals (Fig. 2). 30
31
3.3. Leeches as vectors of Trypanosoma clandestinus n. sp. and long-term infection of caimans 32
Although leeches are known to parasitize crocodilians, they had not been shown to transmit their 33
trypanosomes. In this study, we achieved the experimental infection of caimans with trypanosomes from 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT8
naturally-infected leeches recovered from the mouths of one Ca. yacare captured in the Pantanal. SSU rRNA 1
and gGAPDH sequences of the leech trypanosomes corresponded to the sequence previously determined for 2
the leech TSC64 sample (Table. 1). 3
We employed 30 randomly selected leeches for the experimental infestation of two young caimans. After 4
exposure to the infected leeches, we collected blood samples from the caimans at ~15 day intervals for 5 5
months. Microhematocrit buffy coats show scarce trypanosomes 45 days after infection. Blood samples tested 6
by PCR confirmed the infection of caimans (BSCs 386 and 387) with T. clandestinus (Table 1). The gGAPDH 7
sequences from the infected caiman trypanosome were 99.8-100% similar to those from the leech TSC64. The 8
control caiman and the caimans fed with macerated leeches remained negative by PCR throughout the 9
experiment. Similarly, the ingestion of leeches taken from turtles naturally infected by trypanosomes did not 10
infect turtles (Woo, 1969), suggesting that active leech inoculation of trypanosomes into the bloodstream is 11
required for transmission. 12
13
3.4. Diversity of crocodilian trypanosomes, mixed infections and lack of host-species restriction 14
The surveys using nested-PCR methods for the detection and barcoding of trypanosomes in the blood of 15
four species of caimans disclosed trypanosome diversity and host-species ranges that are greater than those 16
previously reported by hemoculturing. Two new trypanosome species were uncovered increasing to five the 17
number of species (3) previously known (Fermino et al., 2013). Four species placed into the Terrestrial clades 18
(Terena, Ralphi, Gray and Cay03) and one within the Aquatic clade (Clandestinus) of Trypanosoma, each one 19
comprising one trypanosome species and its genotypes (Figs. 1, 3). 20
In addition, results corroborated that diverse host species can successfully be used as hosts by different 21
trypanosome species. The T. clandestinus clade comprises species found in Ca. yacare, Ca. crocodilus and 22
M. niger. The new clade Cay03 was detected in M. niger and Ca. crocodilus. The hosts of T. terena include 23
four of the six species of Brazilian alligatorids: M. niger, Ca. crocodilus, Ca. yacare and P. trigonatus. The list 24
of species hosts for T. ralphi is similar, except for the absence of P. trigonatus (Table 1, Figs. 1, 2). Thus, 25
among all South American alligatorids, trypanosomes remain unreported only in Ca. latirostris and P. 26
palpebrosus, which have not been thoroughly examined yet. 27
Altogether, results from this and previous phylogenetic studies revealed that the species of crocodilian 28
trypanosomes lack host-restriction at species, genus and even family levels. In addition, a crocodilian species 29
may host more than one trypanosome species. Indeed, one-third of the infected caimans harbored two or 30
more trypanosomes. For instance, the Ca. yacare blood sample BSC29 yielded trypanosome sequences of 31
the Ralphi and Clandestinus clades. An extreme example is offered by one M. niger (BSC51) that carried 32
trypanosome sequences from the four Crocodilian subclades (Table 1, Figs. 1, 2). 33
34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT9
1
3.5. The distribution of trypanosome species in caimans of the South American river basins 2
This study shows that T. clandestinus is widespread in the Amazonian, Araguaia/Tocantins and 3
Parana/Paraguay basins, but was not found in the Orinoco basin (Table 1, Fig. 1). Trypanosomes of clade 4
Cay03 occurred in the Amazonian and Orinoco basins, but were not detected in the other basins including the 5
Parana/Paraguay basin, even though large number of caimans were examined (Table 1, Fig. 1). We have 6
previously shown that T. terena and T. ralphi were widespread throughout all of the river basins studied 7
(Fermino et al., 2013). Therefore, the analysis of phylogeographical patterns did not support a strong spatial 8
structure for the crocodilian trypanosomes, because most species occurred throughout the geographic 9
distribution range of their hosts (Fig. 1). 10
11
3.6. Morphology and life cycle of T. clandestinus in caimans and leeches 12
Trypanosomes on blood smears of wild caimans could not be used for description of T. clandestinus 13
trypomastigotes due to the common occurrence of mixed infections with two or more trypanosome species. 14
Our source of blood trypomastigotes were the caimans experimentally infected with T. clandestinus from 15
leeches harboring exclusively this species of trypanosome. Unfortunately, the caimans presented very low 16
parasitemias rarely detected by microhematocrit, although the infection persisted for at least eight months. The 17
few blood trypomastigotes examined (N = 5) in smears from microhematocrit buffy coats exhibited a 18
serpentine body with a pointed posterior extremity, a central rounded nucleus and a posteriorly located small 19
kinetoplast. The flagellum was short and the undulating membrane was convoluted (Fig. 3). The blood forms of 20
T. clandestinus were smaller and thinner than the wider trypomastigotes of T. terena and T. ralphi (Fermino et 21
al., 2013). 22
Leeches of the genus Haementeria are common ecto-parasites of alligatorids. Because the leeches were 23
rather small the examination of the segments of their digestive tract was not attempted and gut contents were 24
wholly mixed for examination. The leeches removed from the caimans were examined after the engorged 25
blood was completely digested, thus ensuring that the flagellates observed ~20 days after leeches collection 26
developed in their guts. 27
As expected for a cyclically transmitted parasite, most leeches were heavily infected with epimastigotes 28
that showed many dividing forms, while some leeches examined after the 20th day presented large numbers of 29
trypomastigotes (Fig. 3). The morphology of the flagellates found in the gut contents of one Haementeria leech 30
(TSC09) removed from a Ca. yacare that had only sequences (14 clones) identical to that of T. clandestinus 31
was selected for the illustration of the trypanosomes found in leeches (defined as type material for species 32
description). The epimastigotes (N = 25) were generally slender, with body lengths varying from 30 to 50 µm 33
(average 42 µm) and widths varying from 2.7 to 4.6 µm (average 3.9 µm). The spherical nuclei and 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT10
kinetoplasts were close together and located near the middle of the body (Fig. 3a-c) and the flagellum length 1
varied from 2.3 to 6.8 µm (average 5.5 µm). The undulating membrane can be narrow or markedly frilled in 2
the widest forms (Fig. 3b). The polymorphism of the trypomastigotes was much more accentuated than that of 3
the epimastigotes. Short trypomastigotes (N = 15) measuring 23 to 42 µm in length (average 38 µm) and 2.2 4
to 2.5 in width (average 2.1 µm) displayed nuclei and kinetoplasts in general displaced towards the anterior 5
end (Fig. 3d, g). Long trypomastigotes, extremely slim at both extremities were abundant. These forms (N = 6
25) measured 60 to 107 µm long (average 77 µm) and 0.7 to 2.5 µm wide (average 1.25 µm) and contained 7
small and elongated nuclei and punctual kinetoplasts located at the center of the body, but not in close 8
proximity to one another. In these thin trypomastigotes the free-flagellum ranged 11 to 20 µm (average 13.3 9
µm) (Fig. 3e, f). 10
Although available data of T. clandestinus in leeches and experimentally infected caimans are not sufficient 11
to explain the entire development of this species in its vertebrate host and vector, overall results allowed us to 12
deduce the following life cycle for T. clandestinus: Haementeria sp. leeches ingest blood containing 13
trypomastigotes upon piercing the caiman buccal mucosa. In the gut of leeches, blood trypomastigotes 14
differentiate into epimastigotes that proliferate and progressively transform into short and long 15
trypomastigotes. Probably, caimans become infected through the bite of leeches carrying trypomastigotes in 16
their proboscis. Although few trypomastigotes were found in the peripheral blood of infected caimans, each 17
leech ingested a considerable volume of caiman blood yielding the high prevalence of T. clandestinus in 18
leeches. 19
20
Taxonomic Summary 21
22
New species description 23
Phylum Euglenozoa Cavalier-Smith 1981; Class Kinetoplastea Honigberg 1963; Order Trypanosomatida (Kent 24
1880) Hollande 1982. Genus Trypanosoma Gruby 1843. Trypanosoma clandestinus Teixeira and Camargo n. 25
sp. 26
Type Material: Hapantotype: trypomastigotes on glass slides of smears of blood sample BSC386 from Ca. 27
yacare, and epi- and trypomastigotes on smears of gut contents from the leech Haementeria sp. (sample TSC 28
64). The slides are deposited at the Trypanosomatid Culture Collection of the University of São Paulo (TCC–29
USP), Brazil. Leeches infected with T. clandestinus are cryopreserved in liquid nitrogen. Paratypes: 30
trypomastigotes on blood films of Ca. yacare blood sample BSC387 stored at TCC-USP. Vertebrate type 31
host: Caiman yacare (Crocodylia, Alligatoridae). Vertebrate additional hosts: Caiman crocodilus and 32
Melanosuchus niger. Invertebrate type host: Haementeria sp. (Hirudinea, Rhynchobdellida), identified by 33
COI sequences deposited in GenBank (KP972452). Habitats: blood of caimans and digestive tube of leeches. 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT11
Type locality: Miranda River (S20o 14’ W56o 22’), Mato Grosso do Sul, the Pantanal, Brazil. Additional 1
localities: AM and AT river basins (Table 1). Morphology: Caiman blood trypomastigotes and leech epi- and 2
trypomastigotes shown in Figure 3. Molecular diagnosis: DNA sequences deposited in GenBank of 3
trypanosomes from Ca. yacare (BSC386), V7V8 SSU rRNA (KP768285) and gGAPDH (KP768260) and from 4
the leech Haementeria sp. (TSC64), V7V8 SSU rRNA (KR107951) and gGAPDH (KP768270). Etymology: 5
clandestinus, Lat. n. m.: hidden, concealed, occult. 6
7
Taxonomical Comments: The lack of host specificity of caiman trypanosomes and the existence of similar 8
blood trypomastigotes shared by different species, intra-species polymorphisms and mixed infections do not 9
permit the assumption that the trypanosomes observed in blood smears belong to a single or multiple species. 10
Therefore, blood trypomastigotes from naturally infected caimans could not be the type material, unless they 11
are proven to belong to a single species. In the absence of derived cultures, it is necessary to verify whether 12
only one trypanosome sequence is present through sensitive PCR and sequencing of a large number of 13
cloned sequences from each blood sample. Alternatively, flagellates from vectors (epi- and trypomastigotes) 14
exhibiting sequences of a single trypanosome species and blood trypomastigotes from animals experimentally 15
infected with molecularly authenticated trypanosomes from the vectors can serve as type material. In this 16
paper, we used the last two procedures to validate the new species because cultures of T. clandestinus are 17
not available. 18
In addition, sequence divergences separating T. terena, T. ralphi and Cay03 were large enough to permit 19
considering Cay03 as a new species within the Terrestrial clade. However, we still do not have a proper type 20
material to taxonomically validate Cay03 as a new species. 21
22
4. Discussion 23
24
Recent molecular studies have uncovered a vast diversity of wildlife parasites within the genus Trypanosoma. 25
However, the molecular phylogenetic studies of trypanosomes of reptiles are still limited to a few species of 26
turtles (Jakes et al., 2001), lizards and snakes (Hamilton et al., 2007; Viola et al., 2008b; 2009) and 27
crocodilians (Viola et al., 2008a; Fermino et al., 2013). In this study, surveys of trypanosomes in blood 28
samples from South American caimans using sensitive nested-PCR methods revealed an unsuspected high 29
prevalence (33%) of infection and common mixed infections by two or more trypanosome species (45%). The 30
phylogenetic positioning of caiman trypanosomes using SSU rRNA and gGAPDH sequences increased our 31
knowledge about the species richness of crocodilian trypanosomes and raised the number of molecularly 32
characterized species from three to five. Two phylogenetically-supported new trypanosome species were 33
identified. One species was designated T. clandestinus n. sp. and molecularly and morphologically 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT12
characterized in the present study. This species was shown to be transmitted by leeches and to cluster within 1
the Aquatic clade of Trypanosoma. Altogether, the information concerning the development and morphological 2
features in leeches and in blood of caimans experimentally infected with this trypanosome species through the 3
bite of leeches, permitted us to outline the probable life cycle of T. clandestinus. 4
Due to the frequent occurrence of mixed infections and the large range of host species, it is impossible to 5
identify a new species of crocodilian trypanosome based on the morphology of blood trypanosomes and host 6
of origin. Phylogenetic analyses are obligatory and, preferably, should be preceded by culturing and 7
morphological characterization of the candidate new species. Before this study, the use of these combined 8
approaches prompted the description of two species of crocodilian trypanosomes (T. terena and T. ralphi) in 9
South American alligatorids (Fermino et al., 2013). Similarly, cultures derived from tsetse flies and C. nyloticus 10
allowed the molecular characterization of T. grayi (Stevens et al., 2001; Hamilton et al., 2009; Fermino et al., 11
2013). 12
In contrast with T. terena, T. ralphi and T. grayi, which are commonly detected by microscopy, 13
hemocultures and conventional PCR, T. clandestinus remains hidden in its crocodilian host and is exclusively 14
detected by highly sensitive nested PCR methods. Contrasting with the very low parasitemia of caiman hosts, 15
T. clandestinus can be easily detected by microscopy and conventional PCR in leeches of the genus 16
Haementeria, which are ectoparasites often found in large clumps in the buccal cavity of crocodilians. The 17
abundant flagellates multiplying in the digestive tract of Haementeria sp. taken from caimans did not grow in 18
culture under any of the conditions we attempted but offered sufficient material for morphological and 19
molecular analyses, and for experimental infections of caimans. T. clandestinus-carrying leeches were used to 20
infect farm-raised Ca. yacare. Blood trypanosomes from the infected caimans yielded V7V8 SSU rRNA and 21
gGAPDH sequences identical to those determined for the trypanosomes multiplying in leeches. Thus, results 22
demonstrated that leeches were capable of transmitting T. clandestinus to Ca. yacare. 23
T. clandestinus is the first crocodilian trypanosome known to be transmitted by leeches and placed into the 24
Aquatic clade. This finding is in agreement with the role of aquatic leeches as vectors of most trypanosomes of 25
fishes, anurans, turtles and the platypus (Martin and Desser, 1991; Siddall & Desser, 1992; Hayes et al., 2014; 26
Paparini et al., 2014). T. clandestinus was the sole trypanosome identified in the gut of Haementeria sp., even 27
in leeches taken from caimans infected with other two or more trypanosome species. This finding support our 28
hypothesis that the crocodilian trypanosomes of the Terrestrial clade are transmitted by hematophagous flies, 29
similarly to some trypanosomes of anurans, lizards and snakes (Ayala, 1971; Ayala and McKay, 1971; Martin 30
and Desser, 1991; Siddall and Desser, 1992; Viola et al., 2008b, 2009) and T. grayi of African crocodiles 31
(Hoare, 1929, 1931). 32
In this study, phylogenetic analyses evidenced, for the first time, that crocodilians of distinct species harbor 33
phylogenetically distant trypanosomes of the two major Trypanosoma clades. The Aquatic clade harbors T. 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT13
clandestinus transmitted by leeches and highly phylogenetically related to fish trypanosomes. The Terrestrial 1
clade harbors T. grayi transmitted by tsetse flies plus T. terena, T. ralphi and Cay03 of unknown vectors. In 2
contrast, all anuran trypanosomes nested in the Aquatic clade even though they are transmitted by leeches or 3
by flies (Ferreira et al., 2007; 2008). It will be interesting to see whether trypanosomes from aquatic snakes 4
transmitted by aquatic leeches (Pessoa and Fleury, 1969; Chia and Miller, 1984) cluster in the Aquatic clade or 5
in the Terrestrial lizard-snake clade of trypanosomes transmitted by sand flies (Viola et al., 2009). To date, a 6
chameleon trypanosome of unknown vector is the only trypanosome from lizard placed in the Aquatic clade 7
(Stevens et al., 2001; Hamilton et al., 2007). 8
Here, we demonstrated that different trypanosome species co-infect different species of crocodilians across 9
South American river basins. The identification of new hosts and new trypanosomes permits us to anticipate 10
that the species richness of crocodilian trypanosomes is much larger than presently known. However, the task 11
of effectively cataloguing the species richness will have to face not only the conceptual issue of species 12
delimitation (Adams, 2001; De Queiroz, 2007), but also the operational problems of new species descriptions. 13
Collecting blood samples and leeches from wild crocodilians in their natural habitats can be very difficult. 14
Moreover, the existence of species such as T. terena and T. ralphi that share morphologically indistinguishable 15
blood forms (Viola et al., 2008a; Fermino et al., 2013) or non-culturable species as T. clandestinus adds 16
difficulties to the appreciation of the trypanosome diversity. T. clandestinus does not belong to the cryptic 17
category of parasites (i.e., two or more species morphologically perceived as a single species), but to a 18
category of parasites that go unnoticed in host blood samples by common methods of detection/identification 19
of trypanosomes. Any inventory of parasite biodiversity and the proposition of well-supported hypotheses 20
about their evolutionary histories depends on the sensitive detection and correct identification of species. The 21
enormous realm of wildlife parasites remains largely underexplored and the occurrence of cryptic, clandestine 22
and misclassified parasite species further complicates the prospect of such an inventory (Criscione et al., 23
2005; Bickford et al., 2007; De León and Nadler, 2010; Detwiler et al., 2010; Nadler and DeLeón, 2011; Poulin 24
et al., 2011b; Bray and Cribb, 2014). Further studies are required to assess whether the specificity of the 25
crocodilian trypanosomes is restricted to closely related crocodilian species or whether they can exploit 26
distantly related hosts of other taxa across their broad geographic range (Poulin et al., 2011a). Caimans are 27
known to be generalist predators that feed on diverse preys including fish, turtles, anurans and other aquatic 28
and terrestrial animals. Animals sharing ecological niches and serving as food are known to share parasites 29
with crocodilians (Junker and Boomker, 2006; Tellez and Nifong, 2014). In this context, it is worth emphasizing 30
that T. clandestinus was placed closest to fish trypanosomes within a major assemblage of parasites 31
transmitted by aquatic leeches. It is known that fishes and anurans play a role of paratenic hosts of 32
Hepatozoon caimani, a highly prevalent caiman hemoparasite transmitted by culicids (Viana et al., 2012; 33
Pereira et al., 2014). Therefore, it is possible that fish and other aquatic vertebrates share trypanosomes with 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT14
caimans, including T. clandestinus, due to host-switching mediated by aquatic leeches and predation by 1
crocodilians. We are currently examining trypanosomes of fishes and anurans that serve as food for the 2
caimans in the Pantanal to investigate the role of ecological fitting favoring host-switching of trypanosomes 3
sharing ecological niches. So far, we have not found trypanosomes identical to T. clandestinus in anurans 4
(Ferreira et al., 2007) sharing ponds with Ca. yacare. However, preliminary phylogenetic analysis revelead 5
partial SSU rRNA sequences closely related to T. clandestinus, but not identical, in blood samples of fishes 6
(BSCs 97, 98, 100) from The Pantanal (Fig. 1). 7
In addition, misclassified trypanosome species may obscure the etiology of infections of wild and farmed 8
caimans. Understanding vectors and parasite species-specific associations have implications for conservation, 9
management and production purposes. Molecular approaches have only recently been used to investigate 10
helminthiasis of endangered species or farmed crocodilians (Zhao et al., 2015; Lott et al., 2015). There are no 11
studies addressing the pathological effects of trypanosome infections in caimans. We did not find any obvious 12
sign of illness in the caimans naturally and chronically infected (8 months) with T. clandestinus. However, the 13
demonstration of trypanosomes in blood and tissue imprints of kidney and lung of wild crocodilians (Lainson, 14
1977; Viola et al., 2008a) deserve further investigations. Trypanosomes in general have been considered non-15
pathogenic in wildlife, although studies are scarce and in general superficial. Recently, studies about 16
trypanosomes infecting endangered Neotropical primates and Australian marsupials started producing relevant 17
data concerning wildlife health and conservation (Maia da Silva et al., 2008; Thompson et al., 2013; Botero et 18
al., 2013; Austen et al., 2015). 19
Results from the present study disclosed an increasing species richness, enlarged host/parasite 20
geographical range, aquatic and terrestrial cycles of transmission and new vectors (leeches) of crocodilian 21
trypanosomes. The diversity and phylogenetic relationships of these trypanosomes likely have been shaped 22
by transoceanic dispersion and ecological fitting with multiple events of host-switching during the long shared 23
evolutionary histories of trypanosomes, crocodilians of different families and leeches. 24
25
Acknowledgements 26
27
We thank Dr. Miguel U. Trefault Rodrigues for the caiman identifications and for providing some archived 28
blood samples of caimans. We also thanks several students and anonymous field workers who helped us in 29
the capture of caimans, fishes and anurans. This work was supported by grants from the Brazilian Agencies 30
CNPq and CAPES. We acknowledge the Brazilian Ministry for Science, Technology and Innovation (MCTI) for 31
field support through the Mamirauá Institute for Sustainable Development (IDSM). Bruno R. Fermino and 32
Priscila Soares are PhD students supported by CAPES and CNPq, respectively. Robson C. Ferreira is a Pos-33
doctoral student sponsored by PNPD-CAPES. 34

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT15
1
2
3
4
5
References 6
7
Adams, B. J., 2001. The species delimitation uncertainty principle. Journal of Nematology 33, 153-160. 8
Austen, J. M., Reid, S. A., Robinson, D. R., Friend, J. A., Ditcham, W. G., Irwin, P. J., Ryan, U., 2015. 9
Investigation of the morphological diversity of the potentially zoonotic Trypanosoma copemani in quokkas and 10
Gilbert's potoroos. Parasitol. 10, 1-10 [Epub ahead of print] 11
Ayala, S. C., 1971.Trypanosomes in Wild California Sandflies, and Extrinsic Stages of Trypanosoma 12
bufophlebotomi. J. Protozool. 18, 433-436. 13
Ayala, S.C., McKay, JG., 1971.Trypanosoma gerrhonoti n. sp., and Extrinsic Development of Lizard 14
Trypanosomes in California Sandflies. J. Protozool. 18, 430-433. 15
Bickford, D., Lohman, D. J., Sodhi, N. S., Ng, P. K., Meier, R., Winker, K., Ingram, K. K. and Das, I., 2007. 16
Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 22, 148-155. 17
Borghesan, T. C., Ferreira, R. C., Takata, C. S., Campaner, M., Borda, C. C., Paiva, F., Milder, R. V., Teixeira, 18
M. M. G., Camargo, E. P., 2013. Molecular phylogenetic redefinition of Herpetomonas (Kinetoplastea, 19
Trypanosomatidae), a genus of insect parasites associated with flies. Protist 164, 129-152. 20
Botero A., Thompson C. K., Peacock C. S., Clode, P. L., Nicholls, P. K., Wayne, A. F., Lymbery, A. J., 21
Thompson, R. C., 2013. Trypanosomes genetic diversity, polyparasitism and the population decline of the 22
critically endangered Australian marsupial, the brush tailed bettong or woylie (Bettongia penicillata). Int. J. 23
Parasitol. Parasites Wildl. 29; 2:77-89. 24
Bray, R. A., and Cribb, T. H., 2015. Are cryptic species a problem for parasitological biological tagging for 25
stock identification of aquatic organisms? Parasitology 142, 1-9. 26
Chia, N. M. and Miller, J. H., 1984. Morphological and Developmental Studies of the Snake Trypanosome 27
Trypanosoma hydrae Ayala, Atkinson, Vakalis, 1983 in Experimentally Infected Hosts and in Culture. J. 28
Protozool. 31, 352-356. 29

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT16
Criscione, C. D., Poulin, R. and Blouin, M. S., 2005. Molecular ecology of parasites: elucidating ecological and 1
microevolutionary processes. Mol. Ecol. 14, 2247-2257. 2
Cristensen, H. A.,Telford Jr, S. R., 1972. Trypanosoma thecadactyli sp. n. from forest geckoes in Panama, 3
and its development in the sandfly Lutzomya trinidadensis (Diptera Psychodidae). J. Euk. Microbiol. 19, 403 -4
406. 5
De León, G. P. P. and Nadler, S. A., 2010. What we don't recognize can hurt us: A plea for awareness about 6
cryptic species. J. Parasitol., 96, 453-464. 7
De Queiroz, K., 2007. Species concepts and species delimitation. Syst. Biol. 56, 879-886. 8
Desser, S.S., McIver, S.B., Jez, D., 1975. Observations on the role of simulids and culicids in the transmission 9
of avian and anuran trypanosomes. Int. J. Parasitol 5, 507-509. 10
Fermino, B. R., Viola, L. B., Paiva, F., Garcia, H. A., de Paula, C. D., Botero-Arias, R., Takata, C. S., 11
Campaner, M., Hamilton, P. B., Camargo, E. P. and Teixeira, M. M. G., 2013. The phylogeography of 12
trypanosomes from South American alligatorids and African crocodilids is consistent with the geological history 13
of South American river basins and the transoceanic dispersal of Crocodylus at the Miocene. Parasit. Vectors 14
6, 313. 15
Ferreira, R. C., Campaner, M., Viola, L. B., Takata, C. S., Takeda, G. F. and Teixeira, M. M. G., 2007. 16
Morphological and molecular diversity and phylogenetic relationships among anuran trypanosomes from the 17
Amazonia, Atlantic Forest and Pantanal biomes in Brazil. Parasitol. 134, 1623-1638. 18
Ferreira, R. C., De Souza, A. A., Freitas, R. A., Campaner, M., Takata, C. S., Barrett, T. V., Shaw, J. J. and 19
Teixeira, M. M. G., 2008. A phylogenetic lineage of closely related trypanosomes (Trypanosomatidae, 20
Kinetoplastida) of anurans and sand flies (Psychodidae, Diptera) sharing the same ecotopes in brazilian 21
amazonia. J Euk. Microbiol. 55, 427-435. 22
Folmer, O., Black, M., Hoeh, W., Lutz, R. and Vrijenhoek, R., 1994. DNA primers for amplification of 23
mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mel. Marine Biol. Biot. 3, 24
294-299. 25
Glassman, A. B., Holbrook, T. W. and Bennett, C. E., 1979. Correlation of leech infestation and eosinophilia in 26
alligators. J. Parasitol. 65, 323-324. 27

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT17
Grybchuk-Ieremenko, A., Losev, A., Kostygov, A. Y., Lukeš, J. and Yurchenko, V., 2014. High prevalence of 1
trypanosome co-infections in freshwater fishes. Folia Parasit. 61, 495-504. 2
Gu, Z., Wang, J., Li, M., Zhang, J., Ke, X. and Gong, X., 2007. Morphological and genetic differences of 3
Trypanosoma in some Chinese freshwater fishes: difficulties of species identification. Parasitol. Res. 101, 723-4
730. 5
Hamilton, P. B., Gibson, W. and Stevens, J. R., 2007. Patterns of co-evolution between trypanosomes and 6
their hosts deduced from ribosomal RNA and protein-coding gene phylogenies. Mol. Phylog. Evol. 44, 15-25. 7
Hamilton, P. B., Stevens, J. R., Gidley, J., Holz, P. and Gibson, W. C., 2005. A new lineage of trypanosomes 8
from Australian vertebrates and terrestrial bloodsucking leeches (Haemadipsidae). Int. J. Parasitol. 35, 431-9
443. 10
Hamilton, P.B., Adams, E.R., Njiokou, F., Gibson, W.C., Cuny, G., Herder, S., 2009. Phylogenetic analysis 11
reveals the presence of the Trypanosoma cruzi clade in African terrestrial mammals. Infect Genet Evol 9, 81-12
86. 13
Hayes, P. M., Lawton, S. P., Smit, N. J., Gibson, W. C. and Davies, A. J., 2014. Morphological and molecular 14
characterization of a marine fish trypanosome from South Africa, including its development in a leech vector. 15
Parasit Vectors 7, 50. 16
Hoare, C. A., 1929. Studies on Trypanosoma grayi II. Experimental transmission to the crocodile. Trans. R. 17
Soc. Trop. Med. Hyg. 23, 449-484. 18
Hoare, C. A., 1931. Studies on Trypanosoma grayi III, Life cycle in the tsetse fly and in the crocodile. Trans. R. 19
Soc.Trop. Med. Hyg. 23, 449-484. 20
Jakes, K. A., O'Donoghue, P. J., Adlard, R. D., 2001. Phylogenetic relationships of Trypanosoma chelodina 21
and Trypanosoma binneyi from Australian tortoises and platypuses inferred from small subunit rRNA analyses. 22
Parasitol., 123, 483-7. 23
Junker, K., Boomker, J., 2006. Check-list of the pentastomid parasites of crocodilians and freshwater 24
chelonians. 2006. Onderstepoort J. Vet. Res. 73, 27-36. 25
Karlsbaak, E., Haugen, E. and Nylund, A., 2005. Morphology and aspects of growth of a trypanosome 26
transmitted by the marine leech Johanssonia arctica (Piscicolidae) from Northern Norway. Folia Parasitol. 52, 27
209-215. 28

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT18
Khan, R., 1976. The life cycle of Trypanosoma murmanensis Nikitin. Can. J. Zool., 54, 1840-1849. 1
Khan, R., A., Forrester, D. J., Goodwin, T. M. and Ross, C. A., 1980. A haemogregarine from the American 2
alligator (Alligator mississippiensis). J. Parasitol. 66, 324-332. 3
Maia da Silva, F., Naiff, R. D., Marcili, A., Gordo, M., D'Affonseca Neto, J. A., Naiff, M. F., Franco, A. M., 4
Campaner, M., Valente, V., Valente, S. A., Camargo, E. P., Teixeira, M. M. G., Miles, M. A., 2008, Infection 5
rates and genotypes of Trypanosoma rangeli and T. cruzi infecting free-ranging Saguinus bicolor 6
(Callitrichidae), a critically endangered primate of the Amazon Rainforest. Acta Trop. 107, 68-73. 7
Lainson, R., 1977. Trypanosoma cecili n. sp. a parasite of the South American cayman, Caiman crocodilus 8
crocodilus (Linnaeus, 1758) (Crocodilia: alligatoridae). Proc. Natl. Acad. Sci. USA, 3, 87-93. 9
Letch, C. A., 1977. Leeches (Hemiclepsis marginata) as vector of fish trypanosomes. Trans. Roy. Soc. Trop. 10
Med. Hyg. 71, 380-38l. 11
Leslie, A. J., Lovely, C. J. and Pittman, J. M., 2011. A preliminary disease survey in the wild Nile crocodile 12
(Crocodylus niloticus) population in the Okavango Delta, Botswana. J. S. Afr. Vet. Assoc. 82, 155-159. 13
Lott, M. J., Hose, G. C., Isberg, S. R., Power, M. L., 2015. Genetics and infection dynamics of Paratrichosoma 14
sp in farmed saltwater crocodiles (Crocodylus porosus). Parasitol Res. 114:727-35. 15
Martin, D. S. and Desser, S. S., 1991. Development of Trypanosoma fallisi in the leech, Desserobdella picta, in 16
toads (Bufo americanus), and in vitro. A light and electron microscopic study. Parasitol. Res. 77, 18-26. 17
Nadler, S. A. and GP, D. E. L., 2011. Integrating molecular and morphological approaches for characterizing 18
parasite cryptic species: implications for parasitology. Parasitology 138, 1688-1709. 19
Noyes, H. A., Stevens, J. R., Teixeira, M. M. G, Phelan, J. and Holz, P., 1999. A nested PCR for the ssrRNA 20
gene detects Trypanosoma binneyi in the platypus and Trypanosoma sp. in wombats and kangaroos in 21
Australia. Int. J. Parasitol, 29, 331-339. 22
Oaks, J. R., 2011. A time-calibrated species tree of Crocodylia reveals a recent radiation of the true crocodiles. 23
Evolution 65, 3285-3297. 24
Paparini, A., Macgregor, J., Irwin, P. J., Warren, K. and Ryan, U. M., 2014. Novel genotypes of Trypanosoma 25
binneyi from wild platypuses (Ornithorhynchus anatinus) and identification of a leech as a potential vector. 26
Exp. Parasitol. 145C, 42-50. 27

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT19
Pereira, G. R., Soares, P., Gomes, M. Q., Viana, L. A., Manso, P. P., Machado, M. P., Paiva, F., Lourenço-de-1
Oliveira, R., 2014. Are fish paratenic natural host of the caiman haemoparasite Hepatozoon caimani? 2
Parasitol. Res. 113:39-45. 3
Pessoa, S. B. and Fleury G.C., 1969. Duas novas espécies de tripanosomas de cobras do Brasil. Rev. Brasil. 4
Biol. 29, 81-86. 5
Poulin, R., Krasnov, B.R., Mouillot, D., 2011a. Host specificity in phylogenetic and geographic space. Trends 6
Parasitol., 27, 355-361 7
Poulin, R., Krasnov, B., Muillot, D. and Thieltges, D., 2011b. The comparative ecology and biogeography of 8
parasites. Phil. Trans. R. Soc. B. 366, 2379-2390. 9
Scheyer, T. M., Aguilera, O. A., Delfino, M., Fortier, D. C., Carlini, A. A., Sanchez, R., Carrillo-Briceno, J. D., 10
Quiroz, L. and Sanchez-Villagra, M. R., 2013. Crocodylian diversity peak and extinction in the late Cenozoic of 11
the northern Neotropics. Nat. Commun. 4, 1907. 12
Siddall, M. E. and Desser, S. S., 1992. Alternative leech vectors for frog and turtle trypanosomes. J. Parasitol. 13
78, 562-563. 14
Stamatakis, A., 2006. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of 15
taxa and mixed models. Bioinformatics 22, 2688-2690. 16
Stevens, J. R., Noyes, H. A., Schofield, C. J. and Gibson, W., 2001. The molecular evolution of 17
Trypanosomatidae. Adv. Parasitol. 48, 1-56. 18
Swofford, D. L., 2002. Phylogenetic analysis using parsimony (* and other methods). Version 4. Sunderland, 19
MA: Sinauer Associates. 20
Telford Jr, S. R., 1995. Kinetoplastida hemoflagellates of reptiles. In Parasitic Protozoa (ed. Kreier J. P.). Vol. 21
10, pp. 16 – 223. Academic Press, New York. 22
Telford Jr., S.R., 2009. Hemoparasites of Reptilia, Color Atlas and Text., CRC Press, Taylor & Francis Group, 23
Boca Raton, Florida, USA. 24
Tellez, M. and Nifong, J., 2014. Gastric nematode diversity between stuarine and inland freshwater 25
populations of the American alligator (Alligator mississipiensis daudin, 1812), and the prediction of 26
intermediary hosts. Int. J. Parasitol. 3, 227-235. 27

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT20
Thompson, C., Botero, A., Wayne, A., Godfrey, S., Lymbery, A. and Thompson, R., 2013. Morphological 1
polymorphism of Trypanosoma copemani and description of the genetically diverse T. vegrandis sp. nov. from 2
the critically endangered Australian potoroid, the brush-tailed bettong (Bettongia penicillata) (Gray,1837). 3
Parasit Vectors 6, 121. 4
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. and Higgins, D. G., 1997. The CLUSTAL_X 5
windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic 6
Acids Res. 25, 4876-4882. 7
Viana, L. A., Soares, P., Silva, J. E., Paiva, F., Coutinho, M. E., 2012. Anurans as paratenic hosts in the 8
transmission of Hepatozoon caimani to caimans Caiman yacare and Caiman latirostris. Parasitol. Res., 110, 9
883-6. 10
Viola, L.B., Almeida, R., Ferreira, R. C., Campaner, M., Takata, C. S. A., Rodrigues, A. C., Paiva, F., 11
Camargo, E. P. and Teixeira, M. M. G., 2008a. Evolutionary history of trypanosomes from South American 12
caiman (Caiman yacare) and African crocodiles inferred by phylogenetic analyses using SSU rDNA and 13
GAPDH genes. Parasitology 136, 55-75. 14
Viola, L. B., Campaner, M., Takata, C. S. A., Ferreira, R. C., Rodrigues, A. C., Freitas, R. A., Duarte, M. R., 15
Grego, K. F., Barrett, T. V., Camargo, E. P. and Teixeira, M. M. G., 2008b. Phylogeny of snake trypanosomes 16
inferred by SSU rDNA sequences, their possible transmission by phlebotomines, and taxonomic appraisal by 17
molecular, cross-infection and morphological analysis. Parasitology 135, 595-605. 18
Viola, L. B., Attias, M., Takata, C. S., Campaner, M., De Souza, W., Camargo, E. P., Teixeira, M. M. G., 19
2009b. Phylogenetic analyses based on small subunit rRNA and glycosomal glyceraldehyde-3-phosphate 20
dehydrogenase genes and ultrastructural characterization of two snake Trypanosomes: Trypanosoma 21
serpentis n. sp. from Pseudoboa nigra and Trypanosoma cascavelli from Crotalus durissus terrificus. J. Euk. 22
Microbiol. 56, 594-602. 23
Woo, P.T. K., 1969. The life cycle of Trypanosoma chrysemydis. Can. J. Zool. 47, 1139-1151. 24
Zhao, J. H., Li L., Guo, Y. N., Zhang, L. P., 2015. Dujardinascaris gigantea sp. n. (Nematoda: Ascaridida) from 25
the critically endangered crocodile Alligator sinensis Fauvel (Reptilia: Crocodylia). Parasitol. Res., 114, 801-26
808. 27
28
Legends to Figures 29
30

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT21
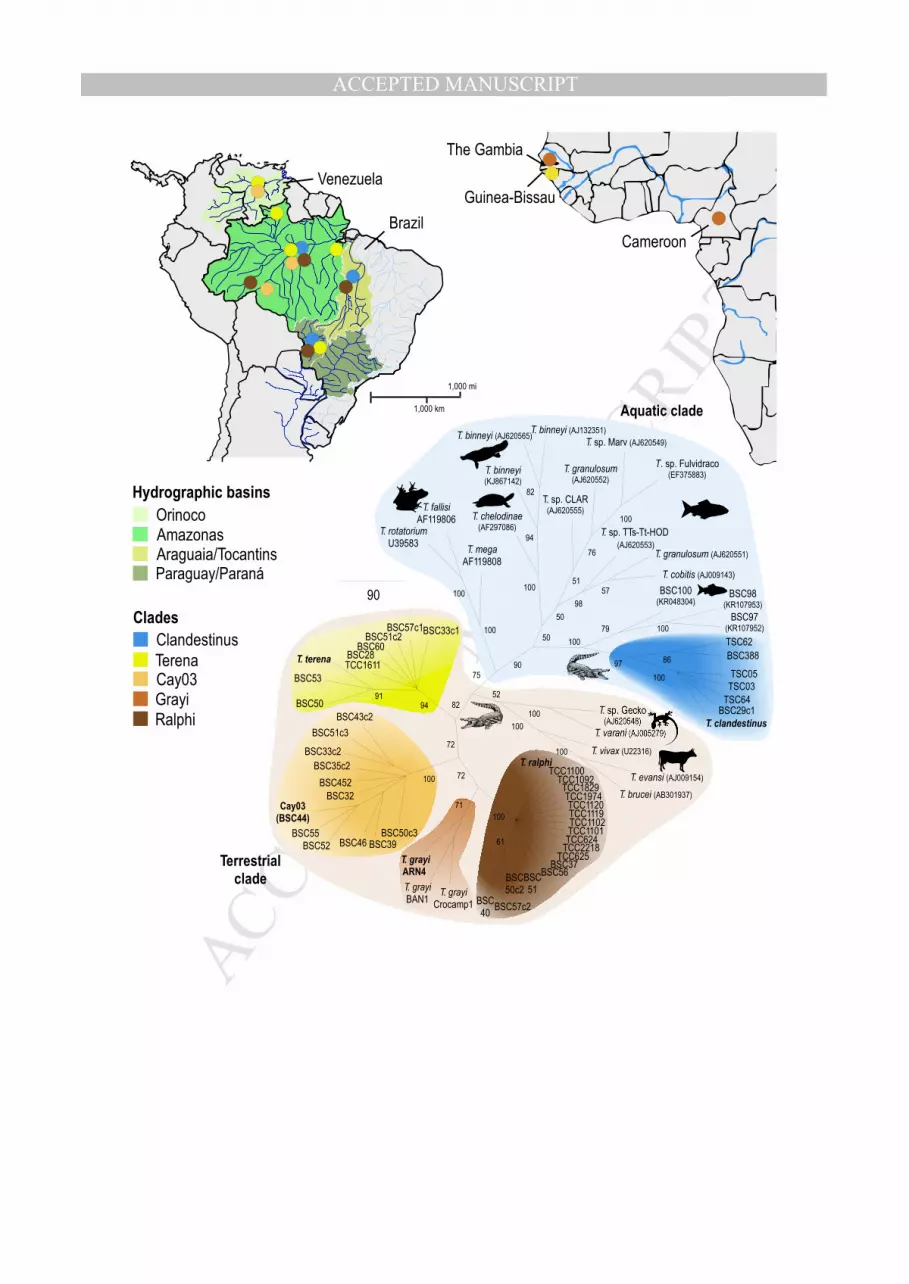
Figure 1. Geographical origin of crocodilian trypanosomes included in the V7V8 SSU rRNA dendrogram 1
inferred to compare the barcode sequences between the new and known trypanosomes from crocodilians and 2
other species of aquatic and semi aquatic hosts. The clade comprising T. clandestinus n. sp. nested into the 3
Aquatic clade closely related to fish trypanosomes whereas sequences of the other new species formed the 4
clade Cay03, which clustered with T. terena, T. grayi and T. ralphi in the Crocodilian Terrestrial clade. The host 5
species and geographic origin and Genbank accession numbers of sequences from the crocodilian 6
trypanosomes are shown in Table 1. Numbers at nodes are bootstrap support values >50% (P/ML) derived 7
from 500 replicates. 8
9
Figure 2. Phylogenetic tree (ML) based on gGAPDH sequences showing the Terrestrial and Aquatic clades of 10
Trypanosoma and the positioning of T. clandestinus. The Crocodilian clade, which is formed by T. terena, T. 11
ralphi, T. gray and Cay03 nests in the Terrestrial Clade whereas the Clandestinus clade comprising T. 12
clandestinus nests in Aquatic clade. Typanosomatid genera other than Trypanosoma were used as outgroups 13
in the phylogenetic trees (608 characters, Ln = -7611.897017). Numbers at nodes are bootstrap support 14
(P/ML) >50% and Bayesan posterior probability > 0.25 derived from 500 replicates. 15
16
Figure 3. Proposed life cycle of T. clandestinus and its developmental and morphological features in caiman 17
blood and leeches. Giemsa-stained blood smears showed blood trypomastigotes of experimentally-infected 18
Ca. yacare, and epi- and trypomastigotes found in the gut of one leech of the genus Haementeria sp. collected 19
in the mouth of a wild Cayman yacare captured in the Pantanal wetland of Brazil. The caiman and the leech 20
trypanosomes were molecularly identified as T. clandestinus. (a-c) epimastigotes; (b) epimastigote dividing by 21
binary fission; (d, g) short trypomastigote; (e,f) long and thin trypomastigotes. Arrow points to the long and thin 22
posterior extremity of very long and slender trypomastigotes. K, kinetoplast; N, nucleus; F, flagellum. 23
24
Supplementary Material 25
26
Figure. Phylogenetic tree (ML) based on concatenated V7V8 SSU rRNA and gGAPDH sequences of T. 27
clandestinus and other trypanosomes of the Aquatic and Terrestrial clades (GenBank accession numbers are 28
in Table 1). Typanosomatid genera other than Trypanosoma were used as outgroups in the phylogenetic trees 29
(1.178 characters, Ln = -17937.481223). Numbers at nodes (P/ML/BI) are bootstrap support values >50% and 30
Bayesian posterior probability > 0.25, derived from 500 replicates. 31

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPTTable 1. Sequences of gGAPDH and V7V8 SSU rRNA trypanosomes from caimans and leeches from South American river basins
Sample# Host River Basin
GPS GenBank Accession Numbers
gGAPDH V7V8 SSU rRNA
Clade Clandestinus
T. clandestinus (BSC386) C. yacare PP 19°57’S 57°01’W KP768260 KP768285
T. clandestinus (TSC64) Haementeria PP 19°57’S 57°01’W KP768270 KR107951
BSC29c1* C. yacare PP 19°57’S 57°01’W KP768252 KP768284
BSC34c1*, BSC35c1*, BSC39, BSC46, BSC49c1*, BSC51c1* M. niger AM 2°19’S 65°70’W KP768253-8 -
BSC64c1* C. crocodilus AT 7°32’S 49°22’W KP768259 -
BSC387 C. yacare PP 19°57’S 57°01’W KP768261 -
BSC388 C. crocodilus AT 7°66’S 49°29W KP768262 KP768286
TSC03 Haementeria PP 19°57’S 57°01’W - KP768287
TSC05 Haementeria PP 19°57’S 57°01’W KP768263 KP768288
TSC06, TSC07, TSC08, TSC09, TSC10 Haementeria PP 19°57’S 57°01’W KP768264-8 -
TSC62 Haementeria AT 7°66’S 49°29W KP768269 KP768289
TSC65 Haementeria PP 19°57’S 57°01’W KP768271 -
Clade Terena
T. terena (TCC621) C. yacare PP 19°57’S 57°01’W EU596252 EU596256
BSC27 O. tetraspis# GB 11°60´N 15°04’W KF546505 -
BSC28 C. crocodilus OR 6°83´N 67°69´W KF546504 KF546518
BSC33c1*, BSC51c2*, BSC57c1* M. niger AM 2°19’S 65°70’W - KP768290-2
BSC50*, BSC53 M. niger AM 2°19’S 65°70’W - KF546519-20
TCC1611 C. yacare PP 19°57’S 57°01’W KF546517 KF546503
BSC60 C. crocodilus AM 7°27’S 64°80W - KP768293
TSC40 P. trigonatus AM 3°80’N 61°73’W KP768272 -
TSC43 C. crocodilus AM 3°20’S 51°86’W KP768273 -
Clade Ralphi
T. ralphi (TCC1838) M. niger AM 7°27’S 64°80W KF546512 KF546527
BSC29* C. yacare PP 19°57’S 57°01’W KF546515 -
BSC37 M. niger AM 2°19’S 65°70’W - KP768294
BSC38 M. niger AM 2°19’S 65°70’W KP768274 -
BSC40 M. niger AM 2°19’S 65°70’W KP768275 KP768295
BSC42, BSC43c1* M. niger AM 2°19’S 65°70’W KP768276-7 -
BSC50c2* M. niger AM 2°19’S 65°70’W - KP768296
BSC51* M. niger AM 2°19’S 65°70’W - KF546524
BSC53 M. niger AM 2°19’S 65°70’W KP768278 -
BSC56 M. niger AM 2°19’S 65°70’W - KF546525
BSC57c2* M. niger AM 2°19’S 65°70’W - KP768297
BSC64c2* C. crocodilus AT 7°32’S 49°22’W KF546516 -
TCC624 C. yacare PP 19°57’S 57°01’W EU596253 EU596257
TCC625, TCC1100 C. yacare PP 19°57’S 57°01’W KF546506 EU596259
TCC1092 C. yacare PP 19°57’S 57°01’W EU596258 EU596254
TCC1101, TCC1102, TCC1119 C. yacare PP 19°57’S 57°01’W KF546507-9 EU596261-3
TCC1120 C. yacare PP 19°57’S 57°01’W KF546510 EU596255
TCC1829 C. crocodilus AM 7°27’S 64°80W KF546511 KF546521
TCC1974 C. yacare PP 19°57’S 57°01’W KF546513 KF546522
TCC2218 C. crocodilus AT 7°32’S 49°22’W KF546514 KF546523
Clade Cay03
BSC32, BSC33c2*, BSC35C2*, BSC39 M. niger AM 2°19’S 65°70’W - KP768298-301
BSC43c2*, BSC44, M. niger AM 2°19’S 65°70’W KP768279-80 KP768302-3
BSC46 M. niger AM 2°19’S 65°70’W - KP768304
BSC49c2* M. niger AM 2°19’S 65°70’W KP768281 -
BSC50c3*, BSC51c4*, BSC52 M. niger AM 2°19’S 65°70’W - KP768305-7
BSC55 M. niger AM 2°19’S 65°70’W KP768282 KP768308
BSC363 C. crocodilus AM 8°80’S 63°95’W KP768283 -
BSC452 C. crocodilus OR 6°83´N 67°69´W - KR107954
# Identification code of the samples deposited at the Trypanosomatid Culture Collection of the University of São Paulo (TCC-USP): TCC, culture
number; BSC, blood sample number; TSC, tissue sample number. *Mixed infections, “c” designates the clone number. South American river basins:
AM, Amazonas; AT, Araguaia-Tocantins; PP, Paraná-Paraguay; OR Orinoco. # GB, Guinea Bissau.
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Venezuela
Brazil
1,000 km
1,000 mi
BSC51c3BSC43c2
BSC33c2
BSC55BSC52 BSC46
Cay03(BSC44)
BSC39
BSC35c2
BSC32BSC452
BSC50c3
90
T. vivax (U22316)
T. brucei (AB301937)
T. evansi (AJ009154)
T. sp. Gecko(AJ620548)
T. varani (AJ005279)
T. sp. CLAR(AJ620555)
T. sp. Fulvidraco(EF375883)
T. sp. Marv (AJ620549)
T. granulosum(AJ620552)
T. granulosum (AJ620551)
T. cobitis (AJ009143)
T. sp. TTs-Tt-HOD(AJ620553)
T. grayiCrocamp1
T. grayiARN4T. grayiBAN1
BSC56BSC51
BSC50c2
BSC37
TCC1829TCC1974TCC1120TCC1119TCC1102TCC1101
TCC1100TCC1092
TCC625TCC624
TCC2218
T. ralphi
BSC57c2BSC40
BSC50
BSC53
T. terena TCC1611BSC28
BSC33c1BSC51c2BSC57c1
BSC60
T. rotatoriumU39583
T. fallisiAF119806
T. megaAF119808
T. binneyi (AJ132351)T. binneyi (AJ620565)
T. binneyi(KJ867142)
T. chelodinae(AF297086)
100
100
100
100
100
100100
100
100
71
9491
72
61
72
52
75
82
90
50
5098
5751
7694
82
Terrestrialclade
BSC29c1T. clandestinus
TSC64TSC03TSC05
TSC62BSC388
BSC100(KR048304)
BSC97(KR107952)
BSC98(KR107953)
100
10079 100
97 86
Aquatic clade
ClandestinusTerena
RalphiGrayiCay03
Clades
OrinocoAmazonasAraguaia/TocantinsParaguay/Paraná
Hydrographic basins
The Gambia
Guinea-Bissau
Cameroon

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Haementeria sp.
Mn Cy Cc
Caiman yacareCaiman crocodilusPaleosuchus trigonatusCrocodylus niloticusOsteolaemus tetraspis
MnCyCcPtCnOt
Host speciesMelanosuchus niger
Glossina spp.
0.1100:100:1
Leishmania major (XM001684852)Angomonas deanei (EU079132)
Phytomonas serpens (EU084894)Herpetomonas megasiliae (DQ092547)
Herpetomonas samuelpessoai (AF047494)83:81:150:87:0.9470:62:-
T. rotatorium (AJ620256)T. mega (AJ620253)
T. fallisi (AJ620254)61:50:0.91T. boissoni (AJ620245)
T. binneyi PB85 (KJ867165)T. binneyi AWW (AJ620266)
T. sp. CLAR (AJ620251)T. sp. Ts-Cc-Sp (AJ620250)T. Ts-Caa-PO (AJ620249)
T. sp. Marv (AJ620248)T. granulosum Portugal (AJ620247)
T. granulosum UK (AJ620246)BSC39BSC51c1
BSC387BSC35c1T. clandestinusBSC46BSC49c1BSC34c1TSC10TSC05TSC08TSC09
BSC388TSC6299:94:1
TSC64c1BSC29c1BSC64TSC65TSC06TSC07
80:84:0.90
64:77:0.84
68:70:0.90
99:96:1
84:76:0.93
T. rangeli (GQ140364)T. conorhini (AJ620267)
T. vespertilionis (AJ620283)T. erneyi (JN040965)
T. dionisii (FN599054)T. cruzi (AJ620269)T. cruzi marinkellei (AJ620270)75:64:0.75
76:55:0.8194:90:1
T. serpentis (FJ236512)T. cascavelli (FJ236511)
T. varani (AJ620261)T. sp. Gecko (AJ620259)
T. avium (AJ620263)T. culicavium (HQ906659)T. corvi (FJ649496)
77:52:0.98
T. vivax (AF053744)T. simiae (AJ620293)
T. evansi (AF053743)T. brucei (AJ620284)100:91:1
99:88:1
51:56:0.73
71:62:0.83
T. terenaBSC28TCC1611
BSC27TSC40TSC43
95:99:1
Cay03BSC49c2BSC363BSC43c2
T. grayi BAN1T. grayi ANR4
T. grayi CroCamp72:63:0.80BSC64TCC1102TCC1119TCC1101BSC29TCC2218BSC42
BSC40TCC624TCC625TCC1092TCC1829TCC1120T. ralphiTCC1974
BSC53BSC38BSC43c1
85:74:1
71:61:0.91
75:60:0.94
99:95:1
AnuransMarine Fishes/Leeches
Freshwater Fishes/Leeches
Mammals
Mammals/Tsetse flies
Lizards/Snakes
Birds
Cy Cc Pt Ot
Mn Cy Cc
Cn
Mn Cc
Platypus/Leeches
Crocodilian Clade
ClandestinusClade
Terrestrial Clade
Aquatic Clade
Leishmania tarentolae (DQ092549)
ClandestinusTerena
RalphiGrayi
Clades
Cay03

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
BloodF
N
K UM
Caiman yacare
g
Trypanosoma clandestinus
Caiman yacare Leech
Leech Haementeria sp.
e
aa N
K
N
K
N
K
NKF
N K
F
f
d
cb
a
N
K

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPTHighlights
• Species richness and geographical range of caiman trypanosomes
• T. clandestinus n. sp. is transmitted by leeches
• T. clandestinus nests in the Aquatic clade closest to fish trypanosomes
• Caimans were infected by the bite of leeches carrying T. clandestinus





























