Embed Size (px)
Citation preview

Accepted by P. Novis: 30 Aug. 2011; published: 14 Oct. 2011 1
PHYTOTAXAISSN 1179-3155 (print edition)
ISSN 1179-3163 (online edition)Copyright © 2011 Magnolia Press
Phytotaxa 30: 1–41 (2011) www.mapress.com/phytotaxa/ Article
Fifteen new diatom (Bacillariophyta) species from Lake Ohrid, Macedonia
ZLATKO LEVKOV1 & DAVID M. WILLIAMS2
1 Institute of Biology, Faculty of Natural Sciences, Gazi Baba bb, Skopje, R. Macedonia. E-mail: [email protected] Department of Botany, the Natural History Museum, Cromwell Road, London, United Kingdom. Email: [email protected]
Abstract
During recent observations of the diatom flora of Lake Ohrid, fifteen new species were found, each possessing uniquemorphological features: Odontidium minutum, Staurosirella pulchella, Staurosirella lata, Fragilaria micra, Naviculasubviridula, Prestauroneis tumida, Placoneis pseudabundans, Placoneis subelegans, Cymbopleura tumida,Gomphosphenia tenuis, Gomphonema mihoi, Gomphonema subaequale, Gomphonema perolivaceolacuum,Gomphonema linearoides and Gomphonema densistriatum. We provide detailed descriptions for all fifteen species basedon light and electron microscopy observations. Noteworthy is the high diversity recorded in the Gomphonema olivaceumspecies complex, which comprises eight species in total, three of which are described as new. We also provide a key foridentification of species in the G. olivaceum complex in Lake Ohrid and a compilation of published data detailing theinfraspecific diversity of this complex.
Introduction
The diatom flora of Lake Ohrid has been under investigation for more than a century (Krstic et al. 2006).There has been intensive study on diatoms during the last 15 years and more than 80 new species have beendescribed (Krammer 1997, 2003, Lange-Bertalot 2001, Levkov et al. 2006a, 2006b, 2007, Levkov &Williams 2006). For example, a revision of the genus Amphora sensu stricto resulted in 23 species recordedfrom Lakes Ohrid and Prespa, of which 16 were new to science (Levkov 2009).
Recent observations on the diatom flora of Lake Ohrid, using samples collected from St. Naum springsand Lake Ohrid, revealed 15 new species, which are described here. The descriptions are based on light andelectron microscopy observations. Of note is the high diversity recorded for the Gomphonema olivaceumspecies complex, which comprises in total eight species, three of which are formally described as new. Wealso provide a key for the identification of taxa within the G. olivaceum species complex and have compiledpublished data detailing the infraspecific diversity of this complex. Further comments on the taxonomy ofseveral genera and species complexes are given.
Material & methods
Monthly samples were collected from St. Naum springs and Lake Ohrid from May 2002 to June 2003 and ona number of other occasions prior to 2008. Samples were prepared by acid digestion with K2MnO4/HCl, and
permanent slides were mounted in Naphrax®. Photomicrographs were taken with a Nikon E–800, a digitalNikon Coolpix 4500, and a digital Axiocam MRc camera mounted on a Zeiss Axioplan microscope.
For SEM analyses, cleaned material was dried onto aluminum stubs and coated with gold/palladium usinga sputter coater. SEM micrographs were taken using a Cambridge Instrument S4 Steroscan electronmicroscope operated at 5 kV.

Slides are deposited in the Natural History Museum, London (BM), the Macedonian National Diatom Collection (MKNDC) at the Institute of Biology, Skopje, Macedonia.
Descriptions derived from the light microscope are indicated with an "LM" after the account; descriptions derived from the scanning electron microscope are indicted with an "SEM" after the account.
The classification largely follows Round et al. (1990) with additional modifications proposed by Medlin & Kaczmarska (2004).
New species descriptions
Descriptions of the new species follow, and illustrations of them are given in Figs 1–258. For the most part the figures follow the sequence of descriptions: in each taxon the light micrographs (LM) are first followed by the scanning electron micrographs (SEM). This sequence was disrupted a little for the first few taxa so that light micrographs could be grouped together where possible.
Division Bacillariophyta Subdivision Bacillariophytina Medlin & Kaczmarska (2004)Class Bacillariophyceae Haeckel emend. Medlin & Kaczmarska (2004)Order Fragilariales Silva (1962) sensu emend.Family Fragilariaceae Greville (1833)Genus Odontidium Kützing (1844)
Odontidium minutum Levkov & D.M.Williams, sp. nov. (Figs 1–9; 25–30)Frustula aspectu cinguli rectangulata. Valvae ellipticae (nec lanceolatae) apicubus rotundatae (nec cuneatae), 12–16
µm longae, 9–11 µm latae. Costae transapicales 2–5 in 10 µm, 4–6 striae inter costas. Striae simplices, paralellae in media parte, transientes radiantes vel paulo curvatae sub apicibus 24–30 in 10 µm facile discernandae microscopio photonico. Cingulum simplex et a 4–8 zonis apertis compositum.
Type:—MACEDONIA: Lake Ohrid, St. Naum springs, Brown filaments, collection date: 31 January 2003 (Accession No. MKNDC 000407). Slide BM 101466 (holotype). Slide MKNDC 000548 (isotype).
Frustules rectangular in girdle view. Valves elliptical, with rounded poles, length 12–16 µm, width 9–11 µm. Transapical ribs nearly all appear primary, 2–5 in 10 µm, c. 4–8 rows of striae between pairs of ribs. Striae simple, parallel towards middle, becoming radiate and slightly curved at valve apices, 24–30 in 10 µm, indistinct. Cingulum simple, composed of 4–8 bands (LM).
External valve surface rippled, with numerous small spines (spinules) along valve face margin, each occurring on a virga, irregularly positioned, pointing out from valve in variable directions (Figs 25, 27, 28). Outer valve surface covered with small protrusions (nodules); internally, with uniseriate striae; transapical ribs primary, with few secondary ribs (Figs 29, 30). Sternum broad, ill-defined, extending from each pole into apical pore field (Fig. 27). One rimoportula per valve, polar, oriented within or adjacent to a stria, replacing 3–5 vimines, a simple slit externally (Figs 27, 28), internally with protruding lips (Figs 29, 30). Apical pore field simple, occurring at each pole, composed of round pores. APF pores are more closely packed than areolae density (Figs 27, 28). Girdle composed of c. 6–10 open bands, all similar, with single series of poroids on pars interior (Figs 25, 26), multiple sets of poroids on pars exterior (Figs 25, 26). Bands become progressively narrower towards epivalve (Figs 25, 26) and covered with small nodules (SEM).
Observations:—Odontidium minutum can be characterized by its small valve size and shape, retained during its life cycle. It resembles some smaller specimens of Odontidium mesodon but can be differentiated from it by the valve shape (lanceolate to elliptic–lanceolate) and its smaller size. Additionally, the striae in O. mesodon are coarser and clearly visible with LM (Williams 1985). Odontidium hiemalis has longer valves
LEVKOV & WILLIAMS2 • Phytotaxa 30 © 2011 Magnolia Press

with a clearer linear outline, coarse striae and a distinct sternum (Williams 1985), features not present in O. minutum. Diatoma vulgaris var. brevis Grunow was considered a synonym of D. vulgaris by Williams (1985) while Krammer & Lange-Bertalot (1991) considered it as an unnamed variant. Patrick & Reimer (1966) recognized Diatoma vulgaris var. brevis as a separate entity characterized by lanceolate shaped valves, larger size (L= 24–50 µm; B= 11–13 µm), coarser and distantly spaced striae (about 16 in 10 µm) and a distinct, linear sternum. Odontidium minutum can be easily differentiated from D. vulgaris var. brevis by its valve shape, size, number and density of ribs (6–8 in 10 µm in the latter), and presence of a distinct sternum.
Odontidium can be separated from Diatoma by the structure of the sternum—wide and diffuse in Odontidium—its complex girdle structure and the coarser structure of the striae and ribs of the valves (Van Heurck 1896). The general features of Odontidium are present in these specimens, hence the generic placement of this taxon.
FIGURES 1–24. LM micrographs. Figs 1–9. Odontidium minutum. Figs 10–17. Staurosirella pulchella. Figs 18–24.
Navicula subviridula. Scale bar = 10 μm.
Phytotaxa 30 © 2011 Magnolia Press • 3FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

FIGURES 25–30: SEM micrographs of Odontidium minutum (type material). Figs 25, 26. SEM external view of frus-tule and girdle bands. Fig. 27. SEM external valve view. Broad lanceolate sternum, apical pore fields (APF) on the valve apices and a single rimoportula present. Fig. 28. Detail of the valve end showing APF and external opening of rimopor-tula. Fig. 29. Internal valve view. Fig. 30. Detail of valve end showing internal opening of rimoportula, striae and APF. Scale bars in Figs 25, 27, 29 = 5 µm, Fig. 26 = 10 µm; Figs 28, 30 = 2 µm.
Genus Staurosirella Williams & Round (1987)
Staurosirella pulchella Levkov & D.M.Williams, sp. nov. (Figs 10–17, 40, 41)Synonym:—Fragilaria pinnata Ehrenberg sensu Jurilj 1954, partim fig. 12iValvae isopolares, lineares ad lineares–lanceolatae apicibus anguste rotundatis. Longitudo valvae 19–43 µm, latitudo
valvae 7–10.5 µm. Area axialis late lanceolatae, ad apices gradatim atenuata. Striae transapicales crassae, distante positae inter se 4.5–5.5 in 10 µm, paralellaе in media parte, radiаntes ad apices.
Type:— MACEDONIA: Lake Ohrid, Biological station, transect, sand 34 m depth, collection date: 3 April 2007 (Accession No. MKNDC 001234). Slide BM 101467 (holotype). Slide MKNDC 001234 (isotype).
LEVKOV & WILLIAMS4 • Phytotaxa 30 © 2011 Magnolia Press

Valves isopolar, linear to linear–lanceolate with narrowly rounded ends. Valve length 19–43 µm, valve width 7.0–10.5 µm. Axial area broad–lanceolate, tapering towards the valve ends. Striae coarse, distantly spaced, 4.5–5.5 in 10 µm, parallel in the middle, becoming radiate towards the valve ends (LM).
Externally, valve striae with robust virgae joining large central sternum; vimines thin, parallel, appear un–occluded; striae raised above surface (Figs 40, 41). Virgae at mantle junction with spines, 2–4, not well pronounced (Figs 40, 41). Apical pores fields probably at each pole, set off from valve features, almost as ocellulimbus (Fig. 41). Girdle unknown (SEM).
Distribution and ecology:—Staurosirella pulchella was observed in the sediment of Lake Ohrid at depths from 20–35 m. According to Jurilj (1954), S. pulchella was observed in the zone inhabited by Charaspp. (10–15 m depth), again in the sediment (mud). So far, there are few records of this species in Lake Ohrid; it was not found in Lake Prespa (Levkov et al. 2007).
Observations:—Staurosirella pulchella is comparable to several Staurosirella (Fragilaria sensu lato) taxa. Fragilaria clevei Pantocsek (1902: figs 9: 216, 217) has similar valve outline, but it differs from S. pulchella in respect of shape of the axial area (narrow, linear in F. clevei) and striae density (6–7 in 10 µm). Hustedt (1931: 161) synonymized F. clevei with F. pinnata var. intercedens (Grunow) Hustedt [= F. mutabilisvar. intercedens Grunow in Van Heurck (1880: fig. 45: 13 = Staurosirella pinnata var. intercedens (Grunow) Hamilton]. Fragilaria pinnata var. intercedens is characterized by linear valves with parallel sides and narrow, linear axial area. Fragilaria mutabilis var. robusta Skvortzow & Meyer (1928: fig. 1: 8) was treated as a synonym of F. pinnata var. intercedens (Hustedt 1931: 161). However, F. mutabilis var. robusta has broadly lanceolate and much wider valves (B=13.6 µm), opposite to linear, symmetrical valves in F. mutabilis var. intercedens. On the other hand F. mutabilis var. baicalensis Skvortzow & Meyer (1928: fig. 1: 9) has asymmetrical to oval and smaller valves (L= 25.5–28.0 µm) with very narrow, linear axial area. Staurosirella lapponica (Grunow) Williams & Round (1987: fig. 21, 26, 27) (= F. lapponica Grunow) is characterized by linear valves with parallel sides 12–40 µm long and 4–6 µm wide. Staurosirella leptostauron var. dubia(Grunow) Edlund (1994: fig. 31) (= F. harrisoni var. dubia Grunow) has elliptic-lanceolate valves with broadly lanceolate axial area, and 6–7 striae in 10 µm, opposite to linear-lanceolate valves with linear to narrow lanceolate axial area and 4–5 striae in S. pulchella.
Staurosirella lata Levkov, D.M.Williams & Cvetkoska, sp. nov. (Figs 31–39; 42–45)Synonym:—Staurosirella leptostauron (Ehrenberg) D.M.Williams & Round sensu Levkov et al. 2007: fig. 19: 26Cellulae solitariae. Frustula aspectu cinguli rectangulata. Valvae late tumidae in media parte cruciateformis interdum
minime asymmetricae. Apices valvarum late rotundatae. Longitudo valvae 35–45 µm, latitudo valvae 26–30 µm. Area axialis late lanceolatae, attenuata versus apices. Striae transapicales crassae, radiantes, 4–5 in 10 µm.
Type:—MACEDONIA: Lake Ohrid, St. Naum springs, sublacustrine spring, mud, 1.5 m depth, collection date: 26 September 2002. Accession No. MKNDC 000132. Slide BM 101468 (holotype). Slide MKNDC 000254. (isotype).
Cells solitary, frustule rectangular in girdle view. Valves strongly swollen at middle, cruciform, occasionally asymmetrical. Valve ends broadly rounded. Valve length 35–45 µm; valve width 26–30 µm. Axial area broad and distinct, tapering towards poles. Striae coarse, radiate, 4–5 in 10 µm (LM).
Valves cruciform, occasionally asymmetrical (Figs 42–44). Valve poles broadly rounded (Figs 42–45). Sternum broad, distinct, tapering towards poles (Figs 42, 43, 45). Externally, valve striae with robust virgae joining large central sternum; vimines thin, parallel, appear un-occluded; striae raised above surface (Figs 42–45). Only very faint indication of spines at mantle junction, mostly at poles (Fig. 43). Apical pores fields probably at each pole, but differing: one pole with narrow, parallel columns (Fig. 45), the alternate pole almost invisible (Figs 43–44). Few bands, valvocopula open, large, copulae open; bands diminish in size away from valve (Fig. 44) (SEM).
Phytotaxa 30 © 2011 Magnolia Press • 5FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Distribution and ecology:—Staurosirella lata was frequently observed in the epipsammic communities near the sublacustrine springs in St. Naum. These sublacustrine springs are calcareous–rich and appear as white unstable (bubbling) plaques “inserted” in dark brown organic sediment. Here the diatom community is dominated by Cocconeis pseudothumensis Reichardt (1982: figs. 1: 1–5) and C. neothumensis Krammer (1990: figs 21–39), while S. lata is usually rare. So far it has only been found in such habitats in St. Naum springs—it has not been observed in the sediment (organic or inorganic) of Lake Ohrid.
FIGURES 31–39: LM micrographs of Staurosirella lata (type material). Scale bar = 10 μm.
Observations:—Staurosirella lata resembles S. leptostauron (Ehrenberg) Williams & Round (1987), which is characterized with strongly inflated valves at the centre, and valve dimensions as 15–30 µm long and 10–16 µm wide, and 6–11 striae in 10 µm. The main differential character between S. leptostauron and S. latais the valve size (L= 35–45 µm; B= 26–30 in the latter) and striae density (4–5 in 10 µm in the latter).
Odontidium harrisonii Roper appears similar to Staurosirella lata, the former now being known as a synonym of F. leptostauron, a proposal made by Hustedt (1931: 154). Hustedt’s view was followed by later authors (e.g. Patrick & Reimer 1966, Krammer & Lange-Bertalot 1991). However, the illustration of O.
LEVKOV & WILLIAMS6 • Phytotaxa 30 © 2011 Magnolia Press

harrisonii in Krammer & Lange-Bertalot (1991: fig. 133: 35) is from the type material (Hull, England) and has several differences in valve size and striae density when compared to S. leptostauron. Odontidium harrisonii has a more rhombic outline, smaller and narrower valves (L= 28 µm; B= 20 µm) with 6–7 striae in 10 µm.
Staurosira harrisonii var. fossilis Pantocsek (1892: fig. 5: 77) has smaller valves (L= 17.5 µm; B= 6.5 µm) with lanceolate shape, that are slightly inflated in the middle. Fragilaria harrisonii var. major Héribaud (1903: fig. 10: 12) has a similar valve size (L= 35–40 µm) to S. lata; however, differences can be observed in the valve shape (being wider at the valve ends and having a wider inflation in the mid-valve) and arrangement of the central striae. Fragilaria harrisonii var. sinuata Hustedt (1927a: fig. 7: 19), which possesses coarsely punctate, uniseriate striae, is clearly distinguished from S. lata.
FIGURES 40–45: SEM micrographs of Staurosirella pulchella and S. lata. Figs 40, 41. Staurosirella puchella. Fig. 40. SEM external valve view. Fig. 41. Detail of the valve apex showing the striae structure and APF. Figs 42–45. Staurosirella lata. Figs 42, 44. external view of frustule and girdle bands. Fig. 43. Detail of valve apex showing the striae structure and APF. Fig. 45. Internal view of valve apex showing areolae occlusions and APF. Scale bars in Figs 40, 42, 45 = 5 µm, Fig. 44 = 10 µm; Figs 41, 43 = 2 µm.
Phytotaxa 30 © 2011 Magnolia Press • 7FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Genus Fragilaria Lyngbye (1819)
Fragilaria micra Levkov & D.M.Williams, sp. nov. (Figs 46–63)Cellulae solitariae. Valvae lanceolatae quoad specimina maiora sed rhombico–lanceolatae quoad specimina minora,
apicibus protractis, rostratis vel capitatis (quoad specimina maiora) ad cuneatis (quoad specimina minora). Longitudo valvae 7–18 µm, latitudo valvae 3.5–4.8 µm. Area axialis angusta, linearis, area centralis lata unlateralis. Striae coarse, uniseriatae, omnino radiantes 18–20 in 10 µm. Puncta striarum non aspectabilia microscopio photonico.
Type:—MACEDONIA: Lake Ohrid, Kaneo springs, macrophytes, collection date: 6 August 2004 (accession No. MKNDC 000589). Slide BM 101469 (holotype); Slide MKNDC 000589 (isotype).
FIGURES 46–63: LM and SEM micrographs of Fragilaria micra (type material). Figs 46–58. Diminution series of Fragilaria micra. Figs 59, 60. SEM external valve view. Fig. 61. SEM internal valve view. Internal foramina appear longer than external. Fig. 62. SEM external valve view. Detail of valve end showing small, indistinct opening of rimoportula. External opening simple, slightly larger than adjacent areolae. Fig. 63. SEM internal valve view. Detail of valve end showing internal opening of rimoportula. Scale bars: LM = 10 μm, SEM in Figs 59–61 = 5 µm, Figs 62, 63 = 1
µm.
LEVKOV & WILLIAMS8 • Phytotaxa 30 © 2011 Magnolia Press

Cells solitary, valves lanceolate in larger specimens to rhombic-lanceolate in smaller specimens with protracted, rostrated or capitate (in larger specimens) to cuneate ends (in smaller specimens). Valve length 7–18 µm, valve width 3.5–4.8 µm. Axial area narrow, linear, central area large, unilateral. Striae coarse, uniseriate, radiate throughout, 18–20 in 10 µm; areolae not recognizable in LM (LM).
External valve face (Figs 59, 60, 62) flat without protrusions (marginal spines, spinules); transition from valve face to valve mantle gradual, with valve mantle being fairly low. Sternum narrow, gradually widening towards central area. Central area unilateral, wide, usually possessing depressions representing rudimentary striae. Striae uniseriate, composed of areolae with round to apically elongate foramina. Striae continuing onto valve mantle, not interrupted at the mantle/face junction. Single rimoportula present at one pole. External opening simple, oriented obliquely, slightly larger than areolae. Apical pore fields present on both poles; pore field composed of simple fine porelli arranged in regular rows parallel to the apical axis (Fig. 62). Internally (Figs 61, 63), costa well-developed, wider than striae. Areolae openings apically elongated; no closing plate observed (SEM).
Distribution and ecology:—Specimens of F. micra were frequently observed in the epiphytic communities near the sublacustrine springs in Kaneo, Lake Ohrid. These sublacustrine springs are calcareous–rich. Here the diatom community is dominated by Rhoicosphenia macedonica Levkov & Krstic (in Levkov et al. 2007: figs 30: 1–17), Diatoma ochridana Lange-Bertalot & Rumrich (in Lange-Bertalot et al. 1991: figs 19–25, 45–48), several species of Gomphonema Ehrenberg, Cymbella Agardh and EncyonemaKützing.
Observations:—Two main features easily differentiate F. micra: its small valve size and valve shape (rhombic–lanceolate). Larger specimens of F. micra may be confused with F. capitellata (Grunow) J.B.Petersen. The valves of F. capitellata have narrowly lanceolate to linear–lanceolate outline (not inflated at the mid-valve) and strongly apiculate ends (Tuji & Williams 2008a: figs 30–43). The valves of F. pectinalis(O.F.Müller) Lyngbye are linear with subcapitate to capitate ends and longer (L=28–37) with lower striae density, 14–15 in 10 µm (Tuji & Williams 2008a: figs 14–27). Fragilaria perminuta (Grunow) Lange-Bertalot (in Krammer & Lange-Bertalot 1991) has a comparable valve size (L=9–24 µm, B= 3–3.5 µm) to F. micra, but differs significantly by its valve shape (strictly lanceolate without inflation in the mid-valve). Fragilaria capucina is characterized by linear valves, bilateral central area and two small rimoportulae, one on each pole (Tuji & Williams 2006a: figs 6: H–R).
The valves of F. vaucheriae (Kützing) J.B.Petersen are linear–lanceolate with coarse and distantly spaced striae (9–14 striae 10 µm) and possess a single rimoportula per valve (Tuji & Williams 2006b: figs 21, 22). Linear–lanceolate valves are present in F. parva (Grunow) Tuji & Williams (2008b: figs 13–28). Additionally, F. parva has a wide, bilateral central area, whereas F. micra has a unilateral central area.
Order Naviculales Bessey (1907) sensu emend.Family Naviculaceae Hendey (1937)Genus Navicula Bory (1822)
Navicula subviridula Levkov, sp. nov. (Figs 18–24, 64–68)Valvae lineari–lanceolatae apicibus plus minusve longe protractis et paene acute rotundatis, longae 39–57 µm, latae 9–
10 µm. Fissurae raphis paullo laterales poris centralibus distincte declinatis ad latus primum valvae (opposite inordinationem Voigtii). Area axialis angusta, linearis, area centralis distincte dilatata irregulariter elliptica ad instar, vix asymmetrica. Striae transapicales radiantes valde convergentes ad apices, 8–10 in 10 µm. Lineolae striarum fere crassae 24–28 in 10 µm.
Type:—MACEDONIA. Lake Ohrid, Bay of St. Naum, sand near the shore, collection date: 17 March 2007 (accession No. MKNDC 001123). Slide BM 101470 (holotype). Slide MKNDC 001123 (isotype).
Valves linear-lanceolate with more or less long protracted to acute ends, length 39–57 µm, width 9–10 µm. Raphe slightly lateral, central pores clearly deflected to primary side. Axial area narrow, linear, central
Phytotaxa 30 © 2011 Magnolia Press • 9FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

area large, irregularly elliptical to asymmetric. Striae radiate at middle, becoming strongly convergent towards apices, 8–10 in 10 µm. Lineolae distinct, coarse, 24–28 in 10 µm (LM).
Valve face flat, valve mantle narrow; valve face/mantle transition gradual (Figs 64, 65). Axial area (sternum) very narrow, linear; central area slightly expanded laterally and apically, round. Raphe system central, proximally raphe terminates with expanded central pores oriented towards primary valve side (Fig. 65). Internally, raphe sternum strongly silicified, prominent; helictoglossae prominent (Figs 66, 67). Terminal fissures hooked, finishing at valve face/mantle transition. Striae radiate to slightly arcuate at middle, becoming convergent near apices. Areolae openings apically elongated (lineolae), smaller towards valve midline. Virgae often wider than striae (SEM).
FIGURES 64–68: LM and SEM micrographs of Navicula subviridula. Fig. 64. SEM external valve view. Fig. 65. Detail of mid-valve showing external openings of areolae (lineolae) and proximal raphe ending. Fig. 66. Internal valve view. Fig. 67. Detail of mid–valve showing raphe sternum and internal openings of areolae. Fig. 68. LM micrograph of
holotype specimen. Scale bars: LM = 10 μm, SEM in Figs 64, 66 = 20 µm, Figs 65, 67 = 5 µm.
LEVKOV & WILLIAMS10 • Phytotaxa 30 © 2011 Magnolia Press

Observations:—The main differential character for N. subviridula is the long protracted and acutely rounded valve ends. Navicula subviridula resembles species in the N. viridula group (section Alinea sensuLange-Bertalot 2001). Navicula viridula (Kützing) Kützing (1844: 91) is a large-celled species of Naviculawith linear–lanceolate valves, shortly protracted to obtusely rounded ends and coarsely punctate striae. Navicula viridula var. linearis Hustedt (1937: figs 19: 1, 2) also has larger valves (L= 65–100 µm, B= 15 µm) with cuneate ends, and coarsely punctate and distantly spaced striae (8–9 in 10 µm). Navicula viridula var. rostrata Skvortzow (1938: fig. 1: 17) has a similar valve shape (linear–lanceolate) to N. subviridula, but differences can be noted in the shape of the valve ends: rostrate to subcapitate vs. acute. The valves of Navicula viridula var. rostrata f. jeniseyensis Skvortzow (1971: figs 10: 1, 2) are broader (B= 14.1 µm) than N. subviridula and have rostrate to subrostrate poles.
Navicula viridulacalcis Lange-Bertalot (2001: figs 36: 4–7) has similar valve outline (linear–lanceolate with slightly convex valve sides) to N. subviridula but the valve ends are shortly protracted and broadly rounded. Navicula viridulacalcis ssp. neomundana Lange-Bertalot & Rumrich (in Rumrich et al. 2000: figs 38: 1–4) is also similar to Navicula subviridula having parallel valve sides. However, the differences can be observed in the shape of the valve ends: abruptly cuneate vs. long protracted to acute ends in N. subviridula.
Navicula novaesiberica Lange-Bertalot (1993: figs 61: 1–8) is probably the most similar taxon to N. subviridula. It is characterized by elliptic–lanceolate to linear–lanceolate valves with abruptly wedge-shaped and obtusely rounded ends. The smaller specimens of N. subviridula might be confused with larger specimens of N. novaesiberica (e.g. Lange-Bertalot 2001: fig. 36: 14), but can be differentiated by the shape of valve and valve ends (elliptic lanceolate valves with wedge-shaped and obtusely rounded ends in N. novaesiberica) and valve size (L= 39–57 µm, B= 9–10 µm in N. novaesiberica). The valves of N. novaesiberica from Danube River (Szabó et al. 2007: figs 1–14) differ significantly from N. subviridula in the shape of the valve ends (subprotracted and obtusely rounded). Levkov et al. (2007: figs 63: 6–10) depicted several valves from Lake Prespa, most probably of N. novaesiberica. However, this species significantly differs from N. subviridulawith respect to its valve shape and valve apices.
Family Stauroneidaceae Greville (1833)Genus Prestauroneis K.Bruder in Bruder & Medlin (2008)
Prestauroneis tumida Levkov, sp. nov. (Figs 69–89)Synonym: Parlibellus protractus (Grunow) Witkowski, Lange-Bertalot & Metzeltin sensu Levkov et al. (2007: figs 64:
10–12)?Navicula crucicula Grunow sensu Gallik (1935, fig. 6)
Valvae lineari–lanceolatae quoad specimina maiora sed elliptico–lanceolatae quoad specimina minora, apicibus protractis, vel subprotractis et late rotundatis. Longitudo valvae 22–44 µm, latitudo valvae 9–11 µm. Fissurae raphis filiformis, rectis extremis proximalibus fere rectae poris centralibus inconspicuis. Area axialis fere angusta, linearis. Area centralis vix formata vel parva plus minusve lanceolata ad ellipticam. Striae transapicales paulo radiantes in media parte, tum paulatim transeunt denique modice convergentes 14–17 in 10 µm. Puncta striarum non discernanda microscopio photonico.Chloroplasti duo disciformes contra quoque latera cingulis positi.
Type:—MACEDONIA. Lake Ohrid, Biological Station Transect, mud 12 m depth, collection date: 03.04.2007 (accession No. MKNDC 001231). Slide BM 101471 (holotype), Slide MKNDC 001231 (isotype).
Valves linear-lanceolate in larger specimens, to elliptic–lanceolate in smaller specimens, with protracted to subprotracted and broadly rounded ends, 22–44 µm long and 9–11 µm wide. Raphe filiform, straight, with proximal ends externally straight, central pores inconspicuous. Axial area narrow, linear. Central area slightly variable to barely developed, mostly small lanceolate or elliptic. Striae slightly radiate in central parts becoming gradually subparallel, finally moderately convergent 14–17 in 10 µm. (LM, areolae invisible in LM). Two plate-like chloroplasts, one against each side of the girdle.
External valve face flat, transition of valve face to mantle gradual (Figs 83–85). Raphe branches straight with slightly widened and unilaterally deflected central pores. Distal raphe fissures prolonged, passing on
Phytotaxa 30 © 2011 Magnolia Press • 11FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

LEVKOV & WILLIAMS12 • Phytotaxa 30 © 2011 Magnolia Press
valve mantle and unilaterally curved in opposite way to proximal ends (Fig. 87). Striae uniseriate, regularlyspaced. Usually one central stria; if two, one is shortened. Areolae variable in shape and orientation, usuallyelongated slits in oblique position (Fig. 86). Internally (Figs 88, 89), with distinct pseudosepta at apices.Raphe straight, proximally terminates with teardrop-shaped central pores, and distally with smallhelictoglossae. Interstriae wider than striae. Internal openings of areolae round, occluded by hymenes (SEM).
FIGURES 69–82: LM micrographs of Prestauroneis tumida. Figs 73, 74. LM of live cell with two chloroplasts. Scale
bar = 10 μm.
Observations:—Navicula protracta Grunow sensu Gallik (1935: fig. 5) has the most similar valves toPrestauroneis tumida, characterized by linear–lanceolate valves with shortly protracted and broadly roundedends. However, its valves are slightly wider (B= 12.7 µm) and the margins are parallel at the middle. Thevalves of N. protracta illustrated in Krammer & Lange-Bertalot (1986: figs 7, 8) and Witkowski et al. (2000:fig 109: 9, 10, fig 134: 12, 13) are characterized by a linear outline, abruptly protracted to subcapitate endsand more distantly spaced central striae. Such features, in combination with narrower valves (B= 7–8 µm),enables easy differentiation from P. tumida. The valve identified as N. protracta in Krammer & Lange-Bertalot (1986: fig. 54: 5) is most likely a specimen of N. protracta f. elliptica Hustedt (see below).
In his treatment of N. crucicula (W. Smith) Donkin, Gallik (1935) described the new taxon N. protractavar. elliptica Gallik (1935: 65, fig. 7), which is characterized by narrowly elliptical valves without protractedand broadly rounded ends, providing features to differentiate it from P. tumida. Hustedt (1957: 283) describeda different taxon under the same name (N. protracta f. elliptica) which clearly differs from P. tumida inseveral respects: its valve length (L= 52 µm), the absence of protracted valve ends, and presence of finelypunctate striae (Simonsen 1987: figs 659: 1, 2).
Prestauroneis tumida can be easily differentiated from N. protractoides Hustedt (1957: figs 32, 33) by thelatter's valve size (L= 17–19 µm, B= 4–5 µm), shape (linear with abruptly protracted ends) and striae

morphology (distinctly punctate). Valves of N. crucicula (W.Smith) Donkin (1871: fig. 6: 14) are lanceolate to elliptic–lanceolate with cuneate to obtusely rounded ends. The valve width is much greater than P. tumida(B=16–18 µm) and the striae are more finely punctate. Navicula cruciculoides Brockman (1950: figs 4: 7–10) has lanceolate valves with cuneate to slightly protracted and narrowly rounded ends, and the striae are distinctly punctuate in LM, features not present in P. tumida.
FIGURES 83–89: SEM micrographs of Prestauroneis tumida. Figs 83–85. External view of whole valves. Fig. 86. Detail of mid-valve showing external foramina and proximal raphe ends. Fig. 87. Detail of valve end showing distal raphe ending. Fig. 88. SEM internal valve view. Fig. 89. Detail of mid-valve showing internal openings of areolae and
proximal raphe endings. Scale bars in Fig. 83 = 5 µm; Figs 84, 85, 88 = 10 µm; Figs 86, 77, 89 = 2 µm.
Navicula crucicula var. lanceolata Frenguelli (1945: fig. 8: 32) has a larger valve size (L= 54–57 µm, B= 12–13 µm) and strictly lanceolate valves. A similar valve outline is found in N. pseudocrucicula Kobayasi & Ando (1975: figs 59–62). Both N. crucicula var. lanceolata and N. pseudocrucicula can be easily differentiated from P. tumida by their valve outline and the distinctly punctated striae.
Phytotaxa 30 © 2011 Magnolia Press • 13FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Navicula crucicula var. alaskana Patrick & Freese (1961: fig. 2: 1) has a comparable valve size to P. tumida but there are significant differences in the shape of valve ends (distinctly rostrate and capitate).
Navicula crucicula var. obtusata Cleve (in Cleve & Grunow 1880: fig. 2: 37) has a similar valve size and shape to P. tumida but occurs in brackish water habitats and is characterized by its distinct, lanceolate central area, unilaterally deflected proximal raphe endings and obtusely rounded and not protracted valve ends.
Finally, Prestauroneis was erected to accommodate species from section Microstigmaticae sensu Hustedt (1961–1966) of the genus Navicula, e.g., species related to Navicula integra (Grunow ex Cleve) Fricke. Some species from this group have been earlier transferred to other genera, such as Parlibellus (Cox 1988) and Proschkinia (Karayeva 1978). In their study on marine diatoms, Witkowski et al. (2000) transferred several species of Navicula sensu lato species into Parlibellus, including N. crucicula, N. protracta and N. protractoides. However, these taxa share morphological and ultrastructural features with Prestauroneis and should be transferred there instead. In Prestauroneis, the striae radiate and are more distantly spaced in the mid-valve, separated with thickened costae giving the appearance of a stauros-like structure. However, a typical stauros (as illustrated in Cox 2001) is not present in Prestauroneis (Figs 75–81). On the other hand, Parlibellus has a wide girdle region due to the numerous girdle bands, two deeply-lobed chloroplasts, external raphe fissures that terminate before the valve apices, internal raphe fissures with thickened axial ribs (Cox 1988). Proschkinia has a wide girdle region composed of numerous punctuate bands, and single isolated stigma.
Order Cymbellales D.G.Mann in Round et al. (1990)Family Cymbellaceae Greville (1833)Genus Placoneis Mereschkowsky (1903)
Placoneis pseudabundans Levkov, sp. nov. (Figs 92–95)Valvae late lanceolatae apicibus curte protractis rostratis, 21–28 µm longae, 9–10.5 µm latae. Raphe filiformis, linearis
poris centralibus paulo incrassatis. Area axialis angusta, linearis, area centralis angusta, elliptica striis circum aream centralem regulariter abbreviatis utrimque. Duae stigmata adsunt dense sita ad striam mediam. Striae transapicales paulo radiantes in media parte, tum valde radiantes sub apices 13–15 in 10 µm. Puncta striarum non aspectabilia microscopio photonico.
Type:—MACEDONIA. Lake Ohrid, St. Naum, shore, mud 5 m depth, collection date: 17 March 2007 (accession No. MKNDC 001128). Slide BM 101472 (holotype). Slide MKNDC 001128 (isotype).
Valves broadly lanceolate with short, protracted to rostrate ends, 21–28 µm long, 9.0–10.5 µm wide. Raphe branches filiform and straight, central pores weakly broadened. Axial area narrow, linear; central area narrow, elliptical, formed by regular shortening of central striae. Two stigmata present in central area, located close to central striae. Striae weakly radiate becoming strongly radiate towards valve ends, 13–15 in 10 µm. Areolae not resolved in LM (LM).
Observations:—One of the main features of P. pseudabundans is the presence of two stigmata in the central area. This character is present in several Placoneis species such as Placoneis abundans Metzeltin et al.(2005: figs 73: 1–4), P. clementis (Grunow) Cox (1988: figs 5: 1, 2) and P. clementoides (Hustedt) Cox (Hustedt 1944: figs 8: 19, 20; Simonsen 1987: figs 476: 4–6). Placoneis abundans is characterized by elliptic-lanceolate valves with abruptly protracted and subcapitate ends, while the valves of P. pseudabundans are lanceolate with short, protracted to rostrate ends. Additionally, there are differences in the shape of the central area. The central area in P. pseudabundans is narrow and elliptical, formed by regular shortening of the central striae, while in P. abundans the central area is wider and bordered by alternating short and long striae. Placoneis clementis (Grunow) Cox is a fossil species (Grunow 1882) characterized by broadly lanceolate valves with short, protracted and obtusely rounded ends, and coarsely punctated striae with 18–20 areolae in 10 µm (Figs 90, 91). There are probably two different entities described as Navicula clementoides Hustedt, with valves of both depicted in Hustedt (1944: figs 8: 19, 20) and Simonsen (1987: figs 476: 6–10) with two
LEVKOV & WILLIAMS14 • Phytotaxa 30 © 2011 Magnolia Press

stigmata in the central area. The first type of valve has elliptic-lanceolate outline with abruptly protracted and capitated ends (Hustedt 1944, fig. 20), while the second has strictly lanceolate valves but neither protracted nor cuneate ends (Hustedt 1944, fig. 19). The differences between P. pseudabundans and P. clementoides are found in the shape of the valve ends and size of the central area. Placoneis gracilis Metzeltin et al. (2005: figs 15–17) has a similar valve outline to P. pseudabundans, but lacks stigmata in the central area.
FIGURES 90–107: LM micrographs. Figs 90, 91. Placoneis clementis. Figs 92–95. Placoneis pseudabundans. Fig. 96. Placoneis juriljii. Figs 97–107. Placoneis subelegans. Scale bar = 10 μm. Figs 90, 91, fossil deposit Köpecz, Romania,
Figs 92–107. Lake Ohrid, recent.
In Lake Ohrid there are several Placoneis species groups. In species belonging to the P. placentula group, stigmata are absent, while species similar to P. tumida and P. significans Lange-Bertalot (in Metzeltin et al. 2005) (= N. exigua var. signata Hustedt) have a single, isolated stigma. So far, only two species present in Lake Ohrid have two stigmata in the central area, Placoneis juriljii Miho & Lange-Bertalot (2006: figs 1–11, 27–35) and P. clementis (Grunow) Cox (1987: 155; figs 28–33). Valves of Placoneis juriljii (Fig. 96) are
Phytotaxa 30 © 2011 Magnolia Press • 15FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

broadly lanceolate with short, protracted, broadly rounded to truncate ends, length 40–73 µm and width 16–20 µm; P. clementis (Levkov et al. 2007: figs 89: 8–12, Miho & Lange-Bertalot 2006: fig. 63), while being very similar to P. pseudabundans, can be differentiated by the shape of the valve ends.
According to current understanding, species in Placoneis have highly variable valve morphology and associated ultrastructure: large differences in almost every ultrastructural feature can be noticed in different species. For example, the stigmata vary from none to one, two or four; the striae can be uniseriate, biseriate, or partly biseriate; the external distal raphe endings can be symmetrical or asymmetrical; the internal proximal raphe endings can be deflected, recurved, or hook-like; the external areolae openings can be surrounded by rims, or round, or elongate, or lineola-like; the internal areolae openings can be occluded, either entirely with vola-like occlusions or not occluded at all. These variations suggest that there may be several lineages that might best be named as subgenera. For example, Kociolek & Thomas (2010) recently described the new genus Rexlowea Kociolek & Thomas based on their observations of North American populations of Navicula navicularis Ehrenberg [= Placoneis navicularis (Ehrenberg) Cox (2003: 71)]. The genus Rexlowea is characterized by the presence of septa and pseudosepta, several isolated stigma–like structures around the central area and condensed pores at the poles. However, no information about chloroplast features (number, shape, position) was given and comparison with species of Placoneis, and other naviculoid and cymbelloid genera, is not yet possible.
Placoneis subelegans Levkov, sp. nov. (Figs 97–113)Valvae lanceolatae ad parum elliptico–lanceolatas apicibus parum protractis et acute rotundatis. Longitudo valvae 26–
44 µm, latitudo valvae 12–15 µm. Raphe filiformis, recta vel parum lateralis. Area axialis angusta linearis, area centralis angusta variabilis, aliquuid rhomboidea ad instar. Striae omnino radiantes 8–10 in 10 µm, puncta striarum non facile discernanda microscopio photonico.
Type:—MACEDONIA. Lake Ohrid, Biological Station Transect, mud 24 m depth, collection date: 3 April 2007 (accession No. MKNDC 001233). Slide BM 101473 (holotype). Slide MKNDC 001233 (isotype).
Valves lanceolate to elliptic–lanceolate with weakly protracted and narrowly rounded ends. Valve length 26–44 µm, valve width 12–15 µm. Raphe filiform to weakly lateral towards centre. Axial area narrow linear, central area narrow, variable in shape, approximately rhombic. Striae radiate throughout 8–10 in 10 µm (LM, areolae hardly visible in LM).
Valve face flat without projections (Figs 108–111). Raphe weakly undulate with dilated proximal endings (Figs 108, 110). Distal fissures recurved in same direction—symmetrical (Figs 112, 113). With post-initial cells (Fig. 108), terminal fissures do not continue onto the valve mantle, but are bordered with short striae at valve apex. striae uniseriate, composed of small elliptical to round areolae (SEM).
Observations:—Placoneis subelegans is similar to P. elegans Metzeltin et al. (2005: figs 75: 1–10); the latter has more elliptical valves with obtusely rounded ends and a broader central area. The valves of Navicula dicephala var. cuneata (M.Møller) Foged (1977: fig. 29: 6) appear similar to P. subelegans but differ in the shape of the valve ends (shortly protracted and broadly rounded), valve width (B= 9 µm) and striae density (S= 11–12 in 10 µm). The smaller specimens of P. ignorata (Schimanski) Lange-Bertalot (= Navicula ignorata Schimanski 1978: figs 6: 1–9) also have a similar valve shape to P. subelegans but the valves are narrower (around 8 µm), the central area is much wider, and the striae are coarsely punctate. Placoneis explanata (Hustedt) Mayama (= Navicula explanata Hustedt 1948: figs 7, 8) has abruptly protracted and broadly rounded ends, a broad transapically widened central area and distinctly punctate striae, features that clearly differentiate it from P. subelegans. Navicula elginensis var. cuneata Kobayasi (in Kobayasi & Ando 1975: figs 4: 54, 55) has smaller valves (L= 19.5–20.5 µm; B= 8 µm) with a large, rhombic central area in contrast to P. subelegans, which is characterized with longer and wider valves (L= 26–44 µm; B= 12–15 µm) with a small central area that is variably shaped.
LEVKOV & WILLIAMS16 • Phytotaxa 30 © 2011 Magnolia Press

FIGURES 108–113: SEM micrographs of Placoneis subelegans. Fig. 108. External view of initial cell; distal raphe endings not passing on valve mantle. Fig. 109. Detail of mid-valve showing proximal raphe ends, small central area and uniseriate striae. Stigma not present. Figs 110, 111. SEM external view of whole valves. Figs 112, 113. Detail of valve ends showing distal raphe endings. Both raphe fissures symmetrical and doubly curved. Scale bars in Figs 108, 111 = 20 µm; Fig. 109 = 5 µm; Fig. 110 = 10 µm; Figs 112, 113 = 2 µm.
Genus Cymbopleura (Krammer) Krammer (1999)
Cymbopleura tumida Levkov, sp. nov. (Figs 115–121, 126–130)Valvae leviter dorsiventrales, late ellipticae marginibus convexis, dorsali magis convexa, margine ventrali leviter
convexa. Apices curte protracti et acute rotundati. Longitudo 42–56 µm, latitudo 22–27.5 µm. Area axialis modice angusta, ad aream centralem dilatata, area centralis angusta, asymmetrica vel lanceolata. Raphe lateralis ad aream centralem et ad apices sensim filiformis fissuris proximalibus bulbiformibus. Striae transapicales crasse punctatae, parallelae in media parte, 6–7/10 µm, radiatae ad apices 9–10 in 10 µm. Puncta striarum 22–24 in 10 µm.
Phytotaxa 30 © 2011 Magnolia Press • 17FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Type:—MACEDONIA. Lake Ohrid, Bay of St. Naum, sand, small spring, collection date: 8 August 2006 (accession No. MKNDC 000693). Slide MKNDC 000693 (holotype). Slide BM 101474 (isotype).
Valves slightly dorsiventral, broadly elliptical with convex margins, dorsal margin more strongly arched. Valve ends slightly protracted and acutely rounded. Valve length 42–56 µm, valve width 22.0–27.5 µm. Axial area narrow, expanding into an asymmetrical or lanceolate central area. Raphe lateral, becoming filiform near poles. Central pores largely expanded. Striae radiate in middle, 6–7 in 10 µm, becoming strongly radiate at poles, 9–10 in 10 µm. Striae coarsely punctate with 22–24 areolae in 10 µm (LM).
FIGURES 114–125: LM micrographs. Fig. 114. Cymbopleura juriljii. Figs 115–121. Cymbopleura tumida. Figs 122–125. Cymbopleura budayana. Scale bar = 10 µm.
Valve face flat, transition from valve face to valve mantle gradual (Fig. 126). Markings (depressions) in central and axial area are absent (Fig. 127). Raphe slightly lateral, proximally terminates in expanded teardrop-shaped central pores, while distally, the raphe fissures deflect towards dorsal valve side (Figs 127, 128). Apical pore fields absent. Striae uniseriate, composed of round to elliptical areolae. Internally, areolae
LEVKOV & WILLIAMS18 • Phytotaxa 30 © 2011 Magnolia Press

also with round openings. Internally, valve face flat (Figs 129, 130). Interstriae wider than striae. Central nodule is elevated from valve (Fig. 130). Raphe distally terminates with small helictoglossae. Intermissio is absent (SEM).
FIGURES 126–130: SEM micrographs of Cymbopleura tumida. Fig. 126. External valve view. Fig. 127. Detail of mid-valve showing central area, with teardrop–shaped central pores. Fig. 128. Detail of valve end, showing distal raphe fissure and uniseriate striae. Fig. 129. Internal valve view. Fig. 130. Detail of mid-valve showing central nodule. Scale
bars in Figs 126, 129 = 20 µm; Figs 127, 128, 130 = 5 µm.
Observations:—Cymbopleura tumida resembles C. budayana (Pantocsek) Krammer (2003: 19) (here as Figs 122–125) but can be differentiated by valve shape (broadly lanceolate in C. budayana), width of axial and central areas (much wider in C. budayana), areolae density (15–21 in 10 µm in C. budayana), and absence of markings in the axial and central area. Cymbopleura juriljii Levkov & Metzeltin (in Levkov et al.2007: figs 147: 1–4) (here as Fig. 114) has a similar valve outline to C. tumida but is longer (L= 100–120 µm) and wider (B= 34–40 µm). Cymbopleura cuspidata (Kützing) Krammer (2003: 8) has narrower valves (B= 14–17 µm) with densely spaced striae (S= 8–12 in 10 µm in the middle, becoming 12–15 in 10 µm at valve
Phytotaxa 30 © 2011 Magnolia Press • 19FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

apices). Cymbopleura apiculata Krammer (2003: fig 7: 8–10, fig 9: 1–6, fig 10: 1–4, fig 11: 1–3b) has comparable valve size and striae density but has a large central area which is distinctly set off and about ⅓ of valve width.
Family Gomphonemataceae Kützing (1844) Genus Gomphosphenia Lange-Bertalot (1995)
Gomphosphenia tenuis Levkov & D.M.Williams, sp. nov. (Figs 131–139, 161–171)Valvae distinctius lineari-clavatae verticibus distincte obtusius rotundatis quam basipoli. Longitudo valvae 22–42 µm,
latitudo valvae 3.5–6.2 µm. Raphe filiformis poris centralibus distinctis. Area axialis variabilis, modice angusta quoad specimina minora sed lata quoad specimina maiora. Area centralis lata unilateraliter fasciam formans. Striae transapicales breves submarginales, radiantes to subparallelae 14–16 in 10 µm.
Type:—MACEDONIA. Lake Ohrid, Bay of St. Naum, rock scrape, small spring, collection date: 17 March 2007 (accession No. MKNDC 001124). Slide BM 101475 (holotype). Slide MKNDC 001124 (isotype).
Valves distinctly club-shaped, narrowly linear, head poles more obtusely rounded than foot poles. Valve length 22–42 µm, valve width 3.5–6.2 µm. Raphe filiform with distinct central pores. Axial area variable, moderately narrow in smaller specimens, wide in larger specimens. Central area a broad unilateral fascia. Striae short, submarginal, radial to subparallel, 14–16 in 10 µm. (LM).
FIGURES 131–160: LM micrographs. Figs 131–139. Gomphosphenia tenuis. Figs 140–148. Gomphonema mihoi. Figs 149–160. Gomphonema subaequale. Scale bar = 10 µm.
LEVKOV & WILLIAMS20 • Phytotaxa 30 © 2011 Magnolia Press

Valve face flat, transition of valve face to mantle forms a gentle right angle (Fig. 161). Valve mantle fairly high, possessing single row of round to slit-like pores (Fig. 161). Raphe branches straight, not passing onto valve mantle (Figs 162, 163). Raphe proximally and distally expanded, terminating with tear-drop-shaped pores (Figs 165, 166). Central pores slightly unilaterally deflected (Fig. 164). Striae each composed of a single areola of variable shape; usually one central stria absent. Areolae near central area and apices have round foramina (Figs 164, 165); remaining areolae are narrow slits positioned near valve margin. Apical pore field absent on both poles. Internally, pseudoseptum absent at both poles (Figs 167, 169). Raphe branches straight, proximally terminate with anchor-shaped fissures (Figs 168, 170), distally with small helictoglossae (Fig. 171). Central nodule is elevated slightly from rest of valve (Fig. 170). Internal openings of areolae much longer than external, occluded by hymenes (SEM).
FIGURES 161–166: SEM micrographs of Gomphosphenia tenuis. Figs 161–163. External valve view; a single row of areolae present on valve mantle. Fig. 164. Detail of mid-valve, showing proximal raphe endings and striae composed of single areola. Proximal raphe endings widened and slightly deflected. One central striae/areola from one valve side absent. Fig. 165. Detail of head pole. Distal raphe fissure widened, terminates with teardrop-shaped pore. Fig. 166. Detail of foot pole. Raphe terminates with teardrop-shaped pore. APF not present on base pole. Scale bars in Fig. 161 = 5 µm; Figs 162, 163 = 10 µm; Figs 164, 166 = 2 µm; Fig. 165 = 1 µm.
Phytotaxa 30 © 2011 Magnolia Press • 21FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES
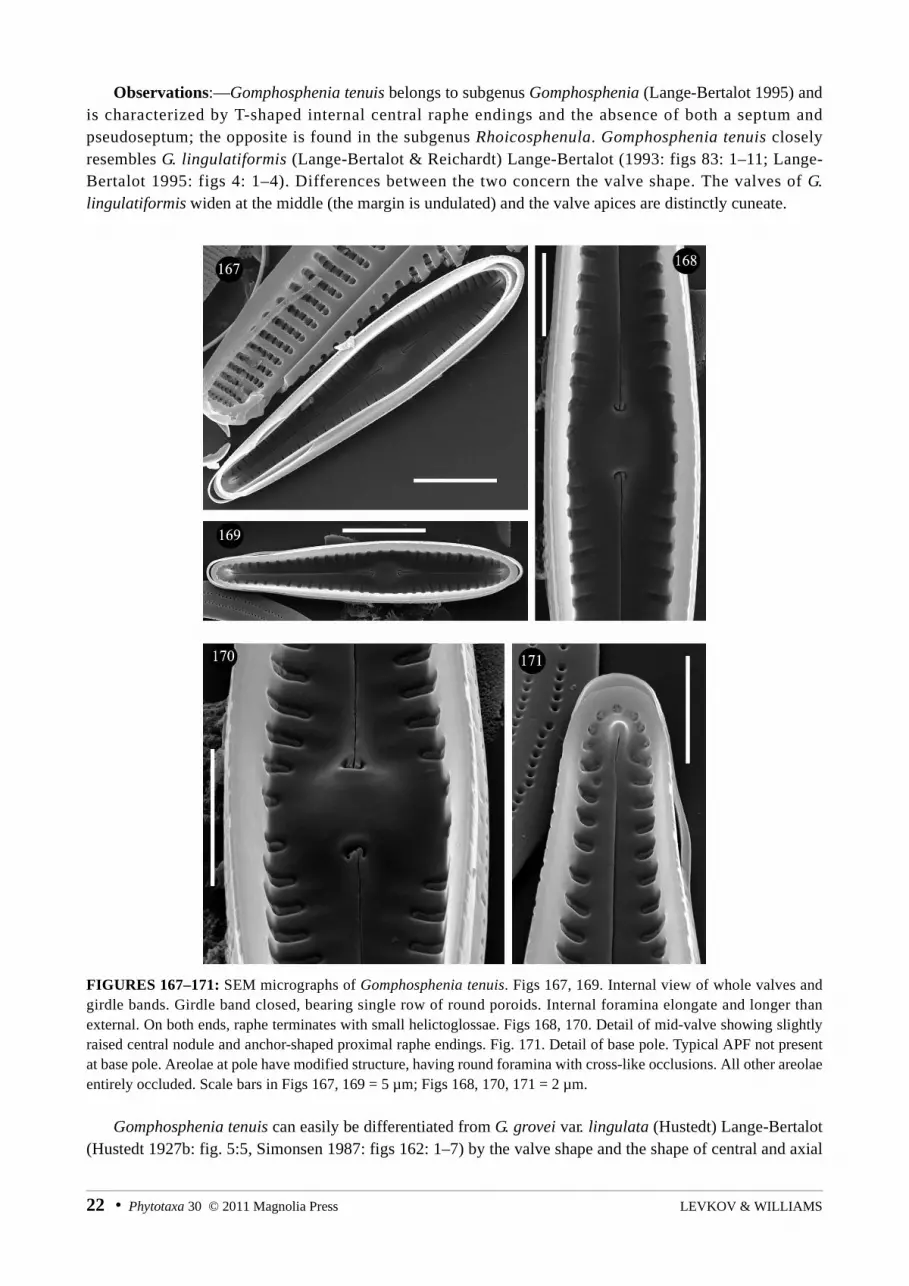
Observations:—Gomphosphenia tenuis belongs to subgenus Gomphosphenia (Lange-Bertalot 1995) and is characterized by T-shaped internal central raphe endings and the absence of both a septum and pseudoseptum; the opposite is found in the subgenus Rhoicosphenula. Gomphosphenia tenuis closely resembles G. lingulatiformis (Lange-Bertalot & Reichardt) Lange-Bertalot (1993: figs 83: 1–11; Lange-Bertalot 1995: figs 4: 1–4). Differences between the two concern the valve shape. The valves of G. lingulatiformis widen at the middle (the margin is undulated) and the valve apices are distinctly cuneate.
FIGURES 167–171: SEM micrographs of Gomphosphenia tenuis. Figs 167, 169. Internal view of whole valves and girdle bands. Girdle band closed, bearing single row of round poroids. Internal foramina elongate and longer than external. On both ends, raphe terminates with small helictoglossae. Figs 168, 170. Detail of mid-valve showing slightly raised central nodule and anchor-shaped proximal raphe endings. Fig. 171. Detail of base pole. Typical APF not present at base pole. Areolae at pole have modified structure, having round foramina with cross-like occlusions. All other areolae entirely occluded. Scale bars in Figs 167, 169 = 5 µm; Figs 168, 170, 171 = 2 µm.
Gomphosphenia tenuis can easily be differentiated from G. grovei var. lingulata (Hustedt) Lange-Bertalot (Hustedt 1927b: fig. 5:5, Simonsen 1987: figs 162: 1–7) by the valve shape and the shape of central and axial
LEVKOV & WILLIAMS22 • Phytotaxa 30 © 2011 Magnolia Press

area. Gomphosphenia tackei (Hustedt) Lange-Bertalot (Hustedt 1942: figs 16–18, Simonsen 1987: figs 459: 7–15) has a similar valve outline to G. tenuis but can be easily differentiated by the narrower valves (B= 2.5–3.5 µm), finely punctate striae and very narrow central and axial areas. Gomphonema tackei var. brevistriatumCamburn (in Camburn et al. 1978: figs 14: 210–214) is even smaller (L= 12–16.5 µm; B= 2–3 µm) and the axial and central areas are indistinct.
Gomphosphenia reicheltii (M.Schmidt) Lange-Bertalot (1995) has distinctly punctate striae and lanceolate axial area in larger specimens and very narrow axial area in smaller specimens. There are probably two different species illustrated as Gomphonema reicheltii M. Schmidt in Schmidt (1899 in Schmidt 1874–1959: figs 214: 19–22); the valves depicted in figs 214: 19, 20 significantly differ in valve shape and shape of the central area from the valves illustrated in figs 214: 21, 22. However, detailed observation of type material is warranted for establishing the proper identity of both populations. Gomphosphenia oahuensis (Hustedt) Lange-Bertalot (in Moser et al. 1998) has isopolar valves with slightly protracted ends, in contrast to the narrowly-clavate valves in G. tenuis. Even larger differences can be observed with SEM in respect to raphe structure (compare with Moser 1998: figs 5: 6–8, figs 53: 1–9).
Gomphosphenia stoermeri Kociolek & E.W.Thomas (in Thomas et al. 2009) has smaller (L=10–20.1 µm) and narrower (B=1.5–3.1 µm) valves with a higher striae density (S= 28–35); the striae are composed of elongated areolae which extend to the raphe slits. These characters make differentiation between G. stoermeriand G. tenuis simple.
Gomphonema brasiliense Grunow (1878: 110) and Gomphonema demerarae (Grunow) Frenguelli (1941: fig. 3: 11) should probably be transferred to Gomphosphenia (Krammer & Lange-Bertalot 1985: figs 41: 1–5); both species significantly differ in the valve shape and size from G. tenuis.
Genus Gomphonema Agardh (1824)
Gomphonema mihoi Levkov, sp. nov. (Figs 140–148, 172–177)Valvae distinctius lineari–clavatae verticibus distincte obtusius rotundatis quam basipoli. Longitudo valvae 18–37 µm,
latitudo valvae 4.5–6.2 µm. Raphe filiformis poris centralibus distinctis. Area axialis angusta linearis, area centralis lata, rectangularis. Stigma solitaria propius ad centrum noduli centralis sita. Striae transapicales radiate vel parallelae sub apices, indistincte punctatae 14–17 in 10 µm.
Type:—MACEDONIA. Lake Ohrid, Bay of Ljubanishta, Chara squeeze from depth 6–7 m; collection date: 2 April 2007 (accession No. MKNDC 001203). Slide BM 101476 (holotype). Slide MKNDC 1203.
Valves distinctly club-shaped, narrowly linear, head poles more obtusely rounded than foot poles. Valve length 18–37 µm, valve width 4.5–6.2 µm. Raphe filiform with distinct central pores. Axial area narrow, linear. Central area broad rectangular. Single isolated stigma present in central area, positioned close to the central pores. Striae slightly radiate to parallel near apices, finely punctate, 14–17 in 10 µm (LM).
Frustules wedge-shaped in girdle view (Fig. 172). Girdle bands with single row of small round poroids. Striae not interrupted on valve mantle, having typical C-like external areolae openings. Valve face smooth (Figs 173, 174). Apical pore field present at base pole, bisected by doubly curved distal raphe ending (Fig. 176). Pore field composed of round porelli arranged in oblique rows. Raphe located in middle of valve; raphe branches straight to weakly undulate. Central pores slightly widened and unilaterally deflected towards the stigma; central area wide (Fig. 174). Single central stria shortened on both valve sides. Striae uniseriate, composed of relatively large, elliptical to oval areolae. Areolae with reniform vela. Internally (Figs 175, 177), pseudoseptum present on the poles. Internal opening of stigma round, with reniform velum similar to areolae. Distal raphe endings terminate with helictoglossae. Proximal raphe endings recurved on elevated central nodule and widened at end (SEM).
Phytotaxa 30 © 2011 Magnolia Press • 23FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

FIGURES 172–177: SEM micrographs of Gomphonema mihoi. Fig. 172. Frustule in girdle view. Fig. 173. External view of whole valve. Fig. 174. Detail of central area, showing proximal raphe endings, stigma and areolae. Fig. 175. Internal view of whole valve. Fig. 176. Detail of base pole showing APF and distal raphe fissures. Fig. 177. Internal valve view, detail of central area, showing internal openings of areolae and stigma. Scale bars in Fig. 172 = 10 µm; Figs 173, 175 = 5 µm; Figs 174, 176, 177 = 2 µm.
Observations:—There are several small-celled species present in Lake Ohrid. Gomphonema balcanicumLevkov & Krstic (in Levkov et al. 2007: figs 169: 11–23, figs 170: 1–8) can be easily differentiated by its very narrow central area and the shape of its foramina. Gomphonema sancti-naumii Metzeltin & Levkov (in Levkov et al. 2007: figs 171: 1–15) has biseriate striae composed of small round areolae. Gomphonema pseudotenellum Lange-Bertalot (in Krammer & Lange-Bertalot 1985: figs 35: 7–12) has narrower valves (B= 3–4 µm) with cuneate apices. Gomphonema mihoi might also be confused with several other species so far not recorded from Lake Ohrid. Gomphonema bohemicum Reichelt & Fricke (in Schmidt 1874–1959: figs 235: 18–25) has a wide unilateral central area which extends to the valve margin (Reichardt 1999: figs 61: 1–22). Gomphonema angustatum (Kützing) Rabenhorst (1864: 283) has lanceolate to rhombic–lanceolate valves
LEVKOV & WILLIAMS24 • Phytotaxa 30 © 2011 Magnolia Press

with acutely rounded to cuneate ends; compared to G. mihoi, it has more distantly spaced striae (10–14 in 10 µm) composed of densely positioned small C-like areolae (Reichardt 1999: figs 23: 1–10, 13–16, figs 24: 1–37). The valve mantle of Gomphonema clavatulum Reichardt (1999: figs 25: 1–23, 27, 28) has 2–3 rows of relatively coarse punctae in contrast to the single row of areolae present in G. mihoi. The central area in G. clavatulum is narrow, made by a slight shortening of central striae and can hardly be differentiated from the axial area (opposite to broad, clearly differentiated central area). Similarly, the differences between G. mihoiand G. paludosum Reichardt (1999: figs 26: 1–22, figs 27: 1–9) are the shape of the central area, areolae morphology and striae density (9–14, usually 10–12 in 10 µm).
Species with similar shaped valves to G. mihoi are from the G. pumilum Reichardt & Lange-Bertalot complex (Reichardt 1997). In general, differences are found in the striae density. The interstriae of the valves in the G. pumilum complex are wide (even wider than the striae) in contrast to the very narrow interstriae in G. mihoi. Additionally, differences in areolae morphology can be found. The areolae in G. pumilum are occluded with a single reniform vola giving the impression of C-shaped foramina. The external opening of the stigma in G. pumilum is surrounded by a rim, while internally the stigma has a simple opening (compare Figs 172–174 with Reichardt 1997: figs 1: 1–7).
Gomphonema subaequale Levkov, sp. nov. (Figs 149–160, 178–184)Frustula aspectu cingulorum cuneata singulis punctis crassis in limbis valve claviformes. Vertices obtuse rotundati vel
late-rotundati, poli basales distincte protracti vel subcapitati. Longitudo valvae 14–21 µm, latitudo valvae 4.5–6.0 µm. Area axialis angustissima, linearis. Area centralis parva variabiliter dilatata ad instar, lanceolata vel transverse elliptica ad rhombico-ellipticam. Stigma solitaria distincte adest. Raphe modice lateralis fissura externa undulata poris centralibus parum incrassatis punctiformibus. Striae transapicales radiantes in media parte subparallelae ad leviter convergentes sub apices, 17–20 in 10 µm. Puncta striarum facile discernanda microscopio photonico.
Type:—MACEDONIA. Lake Ohrid, Kaneo springs, macrophytes, collection date: 6 August 2004 (accession No. MKNDC 000589). Slide BM 101477 (holotype). Slide MKNDC 000589 (isotype).
Frustules in girdle view distinctly cuneate with single row of punctae on girdle bands. Valves clavate with obtusely rounded head pole, distinctly protracted and subcapitate foot pole. Valve length 14–21 µm, valve width 4.5–6.0 µm. Axial area very narrow linear, central area small, variable in shape, lanceolate to rhombic-lanceolate or transversally widened. Single isolated stigma present in central area. Raphe moderately lateral and undulate, central pores small, barely expanded. Striae radiate in valve middle, becoming subparallel or slightly convergent towards valve ends, 17–20 in 10 µm. Areolae hardly visible with LM. (LM).
Valvocopulae bear a single row of poroids, coarser around pole (Fig. 179). Apical pore field situated mainly on mantle, bisected by raphe fissure, composed of small porelli arranged in oblique rows (Figs 178, 183). Porelli smaller than striae areolae, separated from them by narrow hyaline area (Fig. 183). Raphe lateral and undulate (Figs 178, 180). Externally, at both poles, raphe expands and continues as terminal fissures deflected opposite from stigma (Fig. 181). Internally, distal raphe endings terminate with helictoglossae, while proximal endings terminate on elevated central nodule (Figs 182, 184). Proximal raphe fissures recurved (Fig. 182). External opening of stigma small and round, surrounded by rim. Internally, stigma with tear-drop-shaped opening. Striae uniseriate, composed of C-shaped areolae (Fig. 184). Internally, areolae openings round with small finger-like occlusions (Fig. 182) (SEM).
Observations:—The most conspicuous features of G. subaequale are the shape of the foot pole and the densely spaced striae. It may be confused with smaller specimens of G. balcanicum Levkov & Krstic (in Levkov et al. 2007: figs 169: 11–23) but both taxa can be differentiated by valve shape (narrowly clavate valves with slightly protracted foot pole in G. balcanicum) and striae density (13–16 in 10 µm in G. balcanicum). Representatives of the G. minutum complex have biseriate striae, a feature not present in G. subaequale. Gomphonema irroratum Hustedt (1945: figs 42: 47–49) has larger valves (L= 30–55 µm) with distinctly biseriate striae. G. subaequale can be easily distinguished from the G. pumilum complex by valve shape (linear to linear–lanceolate in G. pumilum) and striae density (11–15 in 10 µm).
Phytotaxa 30 © 2011 Magnolia Press • 25FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

FIGURES 178–184: SEM micrographs of Gomphonema subaequale. Figs 178–180. External valve view. Fig. 179. Tilted specimen showing girdle elements. Each girdle band bears a single row of small, round poroids. Fig. 181. Detail of mid-valve, showing proximal raphe endings, stigma and external foramina. Proximal raphe endings widened and slightly deflected towards stigma. External opening of stigma simple and round. Striae uniseriate, composed of C-like areolae. Fig. 182. Internal view of mid-valve showing proximal raphe endings, stigma and internal openings of areolae. Internal fissures crochet-shaped. Fig. 183. Detail of base pole showing APF and distal raphe ending. Fig. 184. Internal valve view. Raphe terminates with small helictoglossae. Scale bars in Figs 178–180, 184 = 5 µm; Figs 181–183 = 2 µm.
The Gomphonema olivaceum (Hornemann) Brébisson (1838) species complex
Gomphonema olivaceum is considered to be one of the most common and frequently encountered diatom species in freshwater ecosystems. It was recorded in a wide range of habitats and differing water qualities from Macedonia as well being frequently identified in Lake Ohrid as an epiphyte living on reed or other submerged macrophytes (Hustedt 1945; Jurilj 1954, Levkov et al. 2007). During his observations of diatoms
LEVKOV & WILLIAMS26 • Phytotaxa 30 © 2011 Magnolia Press
of Lake Ohrid, Hustedt (1945) recorded the nominal variety G. olivaceum var. olivaceum and added another new variety G. olivaceum var. fonticola Hustedt [= G. fonticolum (Hustedt) Levkov & Krstic]. Jurilj (1954), in his study recorded two species from this complex, considered one to be a new variety, i.e. G. olivaceum var. capitatum Jurilj (1954: fig. 68f). On his drawing of this taxon, a stigma in the central area can be clearly seen, a feature not present in members of the G. olivaceum complex. According to biometric data, Jurilj’s figure is more similar to G. italicum Kützing (1844: fig. 75) rather than G. olivaceum.
Recently, Levkov et al. (2007) described the new species G. subolivaceum Levkov & Nakov, with valves possessing biseriate striae and lacking stigmata in the central area. G. subolivaceum differs from other members of the G. olivaceum complex by its smaller valves and densely spaced striae. Additionally, Levkov et al. (2007: figs 173: 3, 4) illustrated a population from Lake Prespa and St. Naum springs on Lake Ohrid, which possesses narrow, linear valves with low stria density. Comparison with other members of the G. olivaceum complex shows that it differs significantly from members of that complex. Additional observations of this complex in Lake Ohrid show that there are three other taxa which have different morphological features or biometric data than those species already known (see Table 1). Here we provide formal descriptions.
TABLE 1. Compilation of published valve shapes, with biometric and ecological data for infraspecific taxa in the G. olivaceum complex. Taxa labelled with a star (*) do not belong to G. olivaceum species complex. Dimensions given in
µm, striae density in 10 µm.
Taxon Valve shape Head pole Length Width Striae Ecology
G. olivaceum f. baicalensis Skvortzow & Meyer 1928: 27, fig. 2: 95
subclavate narrowly rounded
20 3.4 18 freshwater
G. olivaceum f. curta (Van Heurck) Gutwinski 1909: 509*[G. clavatum var. curta Van Heurck 1880: fig. 23: 12]
clavate broadly rounded
31 10 12 ?
G. olivaceum f. minutulum Mayer 1913: pl. 10, fig. 7*
clavate broadly rounded
15–20 5–7 10–12 freshwater
G. (olivaceum var. ?) angustum Kützing sensu Van Heurck 1880: fig. 25: 25*
lanceolate narrowly rounded
26–35 5.9–6.5 11–12 freshwater
G. olivaceum var. balticum (Cleve) Grunow in Van Heurck 1880: figs 25: 24a-c
lanceolate narrowly rounded
29–41 5.9–8.3 12–13 brackish
G. olivaceum var. balticum f. dubia (Mayer) Cleve-Euler 1955: 192, fig. 1291e
clavate truncate 29 8 14–17 brackish
G. olivaceum var. calcareum (Cleve) Van Heurck 1880: fig. 25:23
clavate rounded 22–47 5–8 12–13 freshwater
G. olivaceum var. calcareum f. inflata Mayer 1913: 238; fig. 10: 11
clavate rounded 31 8 22 freshwater
G. olivaceum var. capitata Jurilj 1954: 154, fig. 68f*
lanceolate rounded 23.5 9 20 freshwater
G. olivaceum var. stauroneiformis f. caspia Grunow 1878: fig. 3: 2
lanceolate rounded 65–150 20 6–7 brackish
G. olivaceum var. densestriata Foged 1982: 53–54, fig. 32: 8
clavate broadly rounded
18 8 16–17 freshwater
...... continued
Phytotaxa 30 © 2011 Magnolia Press • 27FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Gomphonema perolivaceolacuum Levkov, sp. nov. (Figs 185–198, 241–245)Valvae late lanceolate marginibus gradatum attenuata ad apices versus. Vertex leniter protractus et late rotundatus.
Polus basalis subprotractus et anguste rotundatus vel subcapitatus. Longitudo valvae 31–53 µm, latitudo valvae 9.5–11.0 µm. Raphe filiformis poris centralibus distinctis. Area axialis angusta linearis, area centralis variabiliter dilatata. Stigma solitaria abest. Striae transapicales valde radiate in media parte, leviter radiate vel parallelae sub apices, 12–17 in 10 µm.
Type:—MACEDONIA. Lake Ohrid, Bay of St. Naum, shore, mosses, collection date: 17 March 2007 (accession No. MKNDC 001120), Slide BM 101478 (Holotype). Slide MKNDC 001120 (isotype).
Valves broadly lanceolate, margins gradually narrowing towards head pole. Head pole slightly protracted and broadly rounded. Foot pole subprotracted and narrowly rounded to subcapitate. Valve length 31–53 µm,
TAB LE 1 (continued)
Taxon Valve shape Head pole Length Width Striae Ecology
G. olivaceum var. fonticola Hustedt 1945: 942, figs 40: 19–22
clavate truncate 19–50 6–9 9–14 freshwater
G. olivaceum var. fossilis Pantocsek 1889: 57, fig. 12: 204
lanceolate, isopolar
rounded 40 4–7 11–12.5 fossil
G. olivaceum var. genuinum Mayer 1913: 237, fig. 10: 8
clavate rounded 15–26 – 10–11 freshwater
G. olivaceum var. genuinum f. minutulum Mayer 1919: 207, fig. 9: 26
clavate rounded 20 7 11 freshwater
G. olivaceum var. genuinum f. typica Mayer 1928: 124, figs 5: 7–11
clavate rounded 25–35 6–7 10–12 freshwater
G. olivaceum var. inflata (Mayer) Mayer 1919: 207, fig. 9: 25
clavate broadly rounded
55 13 13 freshwater
G. olivaceum var. jesperseniana Foged 1982: 54, fig. 32: 11
clavate rounded 29 6.5 16 freswater
G. olivaceum var. minutissima Hustedt 1930: 378, fig. 720
clavate rounded 8–15 3.5–4.5 16–18 freshwater
G. olivaceum var. olivaceoides (Hustedt) Lange-Bertalot in Lange-Bertalot & Krammer 1989: 159, figs 97: 1–20, 24, 25
narrowly clavate
rounded 18–35 5–6 10 freshwater
G. olivaceum var. olivaceolacuum Lange-Bertalot & Reichardt in Lange-Bertalot 1993: 67; figs. 80: 1–8; 81: 3–4; 82: 1–4
clavate obtuse to truncate
20–50 7–13 12–19 freshwater
G. olivaceum var. pusilla Cleve-Euler 1895: 23; fig. 1: 14
clavate truncate 17 4 19 freshwater
G. olivaceum var. quadripunctata Østrup 1908: 85, fig. 1: 11*
clavate rounded 45 8 14 freshwater
G. olivaceum var. ramosissimum (Naegeli) Rabenhorst 1864: 291
- - 23–35 - - freshwater
G. olivaceum var. salina Grunow 1878: 106 lanceolate rounded 30–43 7–9 - saline
G. olivaceum var. salinarum Pantocsek 1889: 58; figs. 9: 160; 11: 199
lanceolate, isopolar
rounded 38–51 7.5–8 10–12.5 fossil
LEVKOV & WILLIAMS28 • Phytotaxa 30 © 2011 Magnolia Press

valve width 9.5–11.0 µm. Raphe filiform with distinct central pores. Axial area narrow, linear. Central area variable in width and shape. Striae strongly radiate at valve middle becoming slightly radiate to parallel near apices, 12–17 in 10 µm (LM).
FIGURES 185–209: LM micrographs. Figs 185–198. Gomphonema perolivaceolacuum. Figs 199–209. Gomphonema linearoides. Scale bar = 10 μm.
Valve face flat, transition to valve mantle gradual (Figs 241–242). Raphe centrally located (Figs 241, 243). Proximal raphe endings widened into teardrop-shaped central pores (Fig. 242). Distal raphe endings at head pole slightly expanded at helictoglossa, continuing as slightly curved terminal fissure passing onto the valve mantle (Fig. 245). Striae biseriate, composed of small round areolae. Apical pore field present on base pole, bisected by distal raphe endings (Fig. 244) (SEM).
Observations:—Gomphonema perolivaceolacuum may be confused with G. olivaceolacuum (Lange-Bertalot & Reichardt) Lange-Bertalot & Reichardt (= G. olivaceum var. olivaceolacuum Lange-Bertalot & Reichardt in Lange-Bertalot (1993: figs 80: 1–8, figs 81: 3–4, figs 82: 1–4). However, the latter has valves that
Phytotaxa 30 © 2011 Magnolia Press • 29FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

have broadly rounded to truncate ends and has a different ecology. Gomphonema olivaceolacuum is observed in meso– to eutrophic lakes, while G. perolivaceolacuum inhabits oligotrophic waters. Gomphonema olivaceum var. calcareum (Cleve) Van Heurck (1880: fig. 25: 23) is narrower (B= 5–8 µm) and has a different shape at the head pole (narrowly rounded). Gomphonema olivaceum var. balticum (Cleve) Grunow (in Van Heurck 1880: fig. 25: 24) has lanceolate valves with narrowly rounded ends and occurs in brackish waters or freshwater with high conductivity.
Gomphonema linearoides Levkov, sp. nov. (Figs 199–209, 246–250)Valvae distinctius lineari-clavatae verticibus distincte obtusius rotundatis quam basipoli. Longitudo valvae 24–35 µm,
latitudo valvae 5.1–6.5 µm. Raphe filiformis poris centralibus distinctis. Area axialis angusta linearis, area centralis lata, rectangularis. Stigma solitaria abest. Striae transapicales valde radiate in media parte, leviter radiate vel parallelae sub apices, 7–11 in 10 µm.
Type:—MACEDONIA. Lake Ohrid, St. Naum springs, above ground springs, macrophytes, collection date: 25 April 2003 (accession No. MKNDC 000632). Slide BM 101479 (holotype). Slide MKNDC 000632 (isotype).
Valves distinctly club-shaped, narrowly linear, head poles more obtusely rounded than foot poles. Valve length 24–35 µm, valve width 5.1–6.5 µm. Raphe filiform with distinct central pores. Axial area narrow, linear. Central area broad panduriform. Striae strongly radiate in valve middle, becoming slightly radiate to parallel near apices, 7–10 in 11 µm (LM).
Raphe on post-initial cells not passing to valve mantle (Fig. 246). Distal raphe endings at head pole are modified. Interstriae of vegetative cells broader than striae (Figs 248, 249). Striae biseriate, composed of two rows of alternating, round areolae (Figs 247, 248). Internally, proximal raphe endings unilaterally hooked (Figs 249, 250) (SEM).
Observations:—Two main characters are important for the differentiation of G. linearoides from other members of the G. olivaceum complex: its linear valves and low striae density. It can be distinguished from G. fonticolum (Figs 231–240, 251, 252) by the shape of the head pole (truncate in G. fonticolum), shape of the base pole (subcapitate in G. fonticolum) and striae density (9–14 in 10 µm in G. fonticolum). Gomphonemaolivaceum var. staurophora Pantocsek (1889: 56, fig. 12: 206) has a low striae density (11.0–12.5 in 10 µm)and similar valve outline but significantly differs with respect to the shape of the central area extending to the valve margins. Gomphonema olivaceum var. minutulum Mayer (1919: 207, fig. 9: 26) has smaller, clavate
valves (L= 15–20 µm) which makes differentiating the pair simple.
Gomphonema densistriatum Levkov, sp. nov. (Figs 210–221, 253–258)Valvae lineari–lanceolatae vel anguste clavatae verticibus late rotundatis polis basalibus angustius rotundatis vel
subprotractis. Longitudo valvae 34–44 µm, latitudo valvae 7.2–8.5 µm. Raphe filiformis poris centralibus distinctis. Area axialis angusta linearis, area centralis variabiliter dilatata ad instar rhombica vel transverse elliptica. Stigma solitaria abest. Striae transapicales valde radiate in media parte, leviter radiate vel parallelae sub apices, 15–18 in 10 µm.
Type:—MACEDONIA. Lake Ohrid, Bay of St. Naum rock scrape, shore, collection date: 17 March 2007 (accession No. MKNDC 001122). Slide BM 101480 (holotype). Slide MKNDC 001122 (isotype).
Valves linear-lanceolate to narrowly-clavate, with broadly rounded head poles, narrowly rounded to subprotracted foot poles. Valve length 34–44 µm, valve width 7.2–8.5 µm. Raphe filiform with distinct central pores. Axial area narrow, linear. Central area variable in shape and size, broadly elliptical to rhombic. Striae strongly radiate in valve middle, becoming slightly radiate to parallel near apices, 15–18 in 10 µm. (LM).
Valve face flat, transition to valve mantle gradual (Figs 253–255). Raphe centrally located (Figs 253, 255). Proximal raphe endings widened into teardrop-shaped central pores (Fig. 254). Distal raphe endings at
LEVKOV & WILLIAMS30 • Phytotaxa 30 © 2011 Magnolia Press

head pole continuing as slightly curved terminal fissures passing on valve mantle (Fig. 255). Striae biseriate, composed of small round areolae. Apical pore field present on base pole, bisected by distal raphe endings (Fig. 256). Several isolated pores at central area, without stigmata (Figs 254, 255). Internally (Figs 257, 258), proximal raphe endings unilaterally hooked (Fig. 258). Distally, raphe terminates with prominent helictoglossae (Fig. 257). Stigmata are absent in the central area (SEM).
FIGURES 210–240: LM micrographs. Figs 210–221. Gomphonema densistriatum. Figs 222–230. Gomphonema subolivaceum. Figs 230–240. Gomphonema fonticolum. Scale bar = 10 μm.
Observations:—The main differential characters of G. densistriatum are the valve shape (linear–lanceolate) and the high striae density. Comparable striae density is found in the valves of G. perolivaceoides(S= 12–17 µm) but the latter has broader (B= 9.5–11 µm) and more broadly clavate valves. G. olivaceolacuumcan be differentiated by valve shape (broadly clavate) and the shape of the head pole (obtusely rounded to truncate).
Phytotaxa 30 © 2011 Magnolia Press • 31FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

FIGURES 241–245: SEM micrographs of Gomphonema perolivaceolacuum. Figs 241, 243. External view of whole valves. Fig. 242. Detail of mid-valve showing small central area, teardrop-shaped central pores and biseriate striae. Fig. 244. Detail of base pole showing distinct APF and straight distal raphe fissure. Fig. 245. Detail of head pole, showing slightly deflected distal raphe fissure. Scale bars in Figs 241, 243 = 10 µm; Figs 242, 244, 245 = 2 µm.
Key for identification of members of the Gomphonema olivaceum complex and related taxa from Lake Ohrid
1. Valves broader than 12 µm, marginal lamina present (visible in girdle view) ........................................................... 2-. Valves narrower than 12 µm, marginal lamina absent ................................................................................................ 32. Central area panduriform, striae 7.5–9 in 10 µm ................... Gomphoneis ohridana Levkov (in Levkov et al. 2007)-. Central area round, striae 9–12 in 10 µm ......................................................................................................................
........................................................... Gomphoneis prespanenis Levkov, Metzeltin & Krstic (in Levkov et al. 2007)3. Four stigmoids present in central area ...................... Gomphonema perolivaceoides Levkov (in Levkov et al. 2007)-. Stigmoids absent in central area ................................................................................................................................. 44. Valves linear, narrower than 7 µm, length/breadth ration 5–7 ................................ Gomphonema linearoides Levkov-. Valves always broader than 7 µm ............................................................................................................................... 5
LEVKOV & WILLIAMS32 • Phytotaxa 30 © 2011 Magnolia Press

5. Valves shorter than 25 µm, length/bread ratio around 2 ....................................................................................................................................................................... Gomphonema subolivaceum Levkov & Nakov (in Levkov et al. 2007)
-. Valves always longer than 25 µm, length/bread ratio higher than 4 ........................................................................... 66. Valves with truncate head pole, maximal width at head pole ........................................................................................
............................................................ Gomphonema fonticolum (Hustedt) Levkov & Krstic (in Levkov et al. 2007)-. Valves with broadly rounded head pole, maximal width near mid–valve .................................................................. 77. Valves wider than 9 µm ............................................................................. Gomphonema perolivaceolacuum Levkov-. Valves narrower than 9 µm ................................................................................. Gomphonema densistriatum Levkov
FIGURES 246–252: SEM micrographs. Figs 246–250. Gomphonema linearoides. Fig. 246. External view of post-initial cell. Distal raphe endings modified and not passing on valve mantle. Mid-valve widened. Fig. 247. External view of whole valve. Fig. 248. Detail of mid-valve showing small central area, widened proximal raphe endings and biseriate striae composed of round to elliptical areolae. Fig. 249. Internal view of whole valve. Fig. 250. Detail of mid–valve, showing pinhole-shaped proximal raphe fissures and biseriate striae. Figs 251, 252. Gomphonema fonticolum. Fig. 251. External view of whole valve. Fig. 252. Internal valve view. Scale bars in Fig. 246 = 20 µm; Figs 247, 249, 251 = 10 µm; Figs 248, 250 = 2 µm, Fig. 252 = 5 µm.
Phytotaxa 30 © 2011 Magnolia Press • 33FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

FIGURES 253–258: SEM micrographs of Gomphonema densistriatum. Figs 253, 255. External view of whole valves. Fig. 254. Detail of mid-valve showing small central area, teardrop-shaped central pores and biseriate striae. Fig. 256. Detail of base pole showing distinct APF and straight distal raphe fissure. Fig. 257. Internal valve view. Fig. 258. Detail
of mid–valve. Scale bars in Figs 253, 255, 257 = 10 µm; Figs 254, 256, 258 = 5 µm.
Discussion
Lake Ohrid is recognized as having high biological diversity and extensive endemism (Stankovic 1960). According to Albrech & Wilke (2008) Lake Ohrid is characterized by the highest index of endemic biodiversity among the ancient lakes. Many authors have recognized the relict character of Ohrid flora and fauna (Albrech & Wilke 2008).
There are few hypotheses concerning the origin of the lake and its biota. According to Radoman (1985) some species complexes are similar to those found in fossil species from the Paratethys (Lake Pannon). Korniushin (2004) also suggests that for some bivalve molluscs, there is a species similarity with the
LEVKOV & WILLIAMS34 • Phytotaxa 30 © 2011 Magnolia Press

Paratethys. Studies on selected species flocks (mostly of molluscs) suggest that only a few lineages originally colonized Lake Ohrid from the Balkans and that the majority of endemic species found today probably started to evolve within the lake during the early Pleistocene. Alternatively, Albrecht et al. (2007) pointed out that there are a number of alternative biogeographical scenarios for the lake’s colonization from Aegean or Anatolian areas during the Messinian Salinity Crisis (see also Wilke et al. 2007, these patterns were also proposed by Banarescu 2004).
Molecular studies on different animal groups show that there are probably two separate groups, one in St. Naum springs and the second in the lake itself (Albrecht et al. 2006), having different origins. Similar results were found by Levkov et al. (2007), pointing out that the relict diatom species occur almost entirely in the lake itself, while the springs (like St. Naum springs) possess a diatom flora comparable to other European waters. For example, Rhoicosphenia is represented by two species which occur in different parts of the lake ecosystem: R. tenuis Levkov & Nakov (2008: fig. 1–14, 30–49) occurs only in St. Naum springs, while R. macedonica Levkov & Krstic (in Levkov et al. 2007) occurs in the shallow waters of the lake itself. Note that, based on morphological features, R. tenuis is assumed to be more closely related to the widespread freshwater species R. abbreviata (Agardh) Lange-Bertalot (1980: figs 1A, 3C–D, 5A), while R. macedonica shares more features with the marine species R. marina M. Schmidt (in Schmidt 1874–1959: figs 213: 29–34). Similar data were shown by Radoman (1983) and Albrecht et al. (2006, 2008), who pointed out that the southern feeder springs harbour their own set of endemic species of molluscs that are absent from the lake proper.
Several studies on diatoms show the similarity of the diatom flora between Lake Ohrid and other ancient lake basins. Hustedt (1945) suggested that the diatom flora of Lake Ohrid resembled that of the volcanic lakes on Java, as well as Köpecz and Bodos (Romania). Maybe the most important studies on this topic are those of Jurilj (1948, 1954). In these two studies, Jurilj found that several of his species were similar, even identical to, fossil species previously described by Pantocsek (1892) from Bodos and Köpecz. Recent studies also show similarities in morphology between populations of certain species of Lake Ohrid with those found in fossil deposits from Romania (Levkov et al. 2007, Levkov 2009). Several species living in Lake Ohrid –Amphora transsilvanica Pantocsek (1892: fig. 1: 12), Stephanodiscus transsilvanicus Pantocsek (1892: fig. 8: 125), Navicula turris Hustedt (in Schmidt 1874–1959: figs 405: 15–18) –have almost identical sets of ultrastructural features as those in populations from Köpecz and Bodos. The morphospecies of Lake Ohrid that appear to have persisted over geological time may have done so as the result of little change to the environment over that time, leading to a slow rate of evolution. This reasoning is commonly invoked to explain the occurrence of so-called “relic” species (e.g. Stankovic 1960).
Knowledge concerning the distribution of endemic diatoms in Lake Ohrid is far from complete. However, four groups of endemic diatoms can be distinguished according to their distribution:
1. Species found in the springs and Lake Ohrid; 2. Species found in both Lakes Prespa and Ohrid; 3. Species found only in the springs; 4. Species found only in certain parts of Lake Ohrid separated by horizontal and vertical barriers such as
the Chara belt.
Those species in Group 4 are represented by relict species. Recent analyses of diatoms from Lake Ohrid in the last glacial-interglacial cycle (~140 ky) show dramatic
changes in diatom composition (Reed et al. 2010). Several species have been recorded in cores but not in recent samples (e.g. Mastogloia spp., Achnanthes spp.). Additionally, changes in valve morphology of the dominant species were noted. For example, Cyclotella ocellata Pantocsek (1902: fig. 318) had 22 different types of valve morphology. So far two hypotheses have been offered as explanations for this large diversity in valve morphology (Cvetkoska 2011): The first hypothesis states that different morphologies are the result of different environmental conditions and that the variation in Lake Ohrid is affected by climate. The second hypothesis states that as different morphologies appear under the same environmental conditions, they represent real, distinct taxa rather than environmentally induced variation.
Phytotaxa 30 © 2011 Magnolia Press • 35FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Based on morphological features, it is suggested that diatom endemism in Lake Ohrid may involve three patterns: 1. Presence of relict species similar to those from Bodos and Köpecz, indicating a previous connectivity of Lake Ohrid with Paratethys (Lake Pannon);2. Presence of endemic species with marine relatives indicating a possible marine invasion, colonization from Aegean or Anatolian areas; 3. Presence of local endemic species indicates intralacustrine speciation. Similar to diatoms, a few studies on molluscs (for details see Radoman 1985) suggested the presence of species flocks in Lake Ohrid.
TABLE 2: List of new species described herein with their supposed origin and distribution.
In Table 2, for each of the 15 new species we have noted their suspected source of origin, distribution and a similar species that lives outside the environs of the lake. By far the most (12) are suspected to be intralacustrine in origin. That is, we suspect they are Lake Ohrid endemics that have evolved in situ. We predict that these species will not be found elsewhere. Two species are suspected to have persisted at this site over geological time and further comparison with a range of fossils is warranted. Finally, one species is considered to be of marine origin.
These patterns of diversity, and the wealth of new diatom taxa in and around Lake Ohrid (many yet to be fully described) suggests that this lake is a unique biological entity, worthy of further study.
Acknowledgements
We thank the Department of Botany, the Natural History Museum, London (BM), for funding a study visit for Dr. Levkov in 2011, and the Alexander von Humboldt Foundation for funding a study visit to the Friedrich Hustedt Centre for Diatom Research (BRM), Bremerhaven, Germany. We are grateful to Dr. Mark Edlund and Dr. Patrick Kociolek for many useful comments on earlier versions of this manuscript.
Species Origin Distribution Similar to
Odontidium minutum Intralacustrine Springs Diatoma mesodon
Staurosirella pulchella Intralacustrine Only certain parts of Lake Ohrid Fragilaria clevei
Staurosirella lata Intralacustrine In springs Staurosirella leptostauron
Fragilaria micra Intralacustrine In the springs and Lake Ohrid Fragilaria capitellata
Navicula subviridula Intralacustrine In the springs and Lake Ohrid Navicula viridula
Prestauroneis tumida Marine Both Lake Prespa and Ohrid Navicula protracta
Placoneis pseudabundans Relict Only certain parts of Lake Ohrid Placoneis clementis
Placoneis subelegans Intralacustrine Only certain parts of Lake Ohrid Placoneis elegans
Cymbopleura tumida Relict Only certain parts of Lake Ohrid Cymbopleura budayana
Gomphosphenia tenuis Intralacustrine In the springs and Lake Ohrid Gomphosphenia lingulatiformis
Gomphonema mihoi Intralacustrine Only certain parts of Lake Ohrid Gomphonema balcanicum
Gomphonema subaquale Intralacustrine In the springs and Lake Ohrid Gomphonema minutum
Gomphonema perolivaceolacuum Intralacustrine In the springs and Lake Ohrid Gomphonema olivaceolacuum
Gomphonema linearoides Intralacustrine In the springs and Lake Ohrid Gomphonema fonticolum
Gomphonema densistriatum Intralacustrine In the springs and Lake Ohrid Gomphonema olivaceum
LEVKOV & WILLIAMS36 • Phytotaxa 30 © 2011 Magnolia Press

References
Agardh, C.A. (1824) Systema Algarum. Adumbravit C.A. Agardh. Lund: Lundae Literis Berlingianis, 312 pp.Albrecht, C., Schultheiß, R., Kevrekidis, T. , Streit, B. & Wilke, T. (2007) Invaders or endemics? Molecular
phylogenetics, biogeography and systematics of Dreissena in the Balkans. Freshwater Biology 52: 1525–1536.Albrecht, C., Trajanovski, S., Kuhn, K., Streit, B. & Wilke, T. (2006) Rapid evolution of an ancient lake species flock:
Freshwater limpets (Gastropoda: Ancylidae) in the Balkan lake Ohrid. Organisms Diversity & Evolution 6: 294–307.
Albrech, C. & Wilke, T. (2008) Ancient Lake Ohrid: biodiversity and evolution. Hydrobiologia 615:103–140.Albrecht, C., Wolff, C., Glöer, P. & Wilke, T. (2008) Concurrent evolution of ancient sister lakes and sister species. The
freshwater gastropod genus Radix in lakes Ohrid and Prespa. Hydrobiologia 615: 157–167.Banarescu, P.M. (2004) Distribution patterns of the aquatic fauna of the Balkan Peninsula. In: Griffith, H. I., et al. (Eds),
Balkan Biodiversity-Pattern and Process in the European Hotspot, Dordrecht, Boston, London: Kluwer Academic Publishers, pp. 243–260.
Bessey, C.E. (1907) A synopsis of plant phyla. Nebraska University Studies 7: 275–373.Bory de Saint-Vincent, J.B.M. (1822) Dictionnaire Classique d'Histoire Naturelle. Paris: Rey & Gravier, libraires-
éditeurs; Baudouin Frères, libraires-éditeurs, 604 pp.Brébisson, A. de (1838) Considérations sur les Diatomées et essai d'une classification des genres et des espèces
appartenant à cette famille. Falaise: Brée l'Ainé, Imprimeur-Libraire. 22 pp.Brockmann, C. (1950) Die Watt-Diatomeen der schleswig-holsteinischen Westküste. Abhandlugen der
Senckenbergischen Naturforschenden Gesellschaft, Frankfurt am Main 478: 1–26.Bruder, K. & Medlin, L.K. (2008) Morphological and molecular investigations of naviculoid diatoms. II. Selected genera
and families. Diatom Research 23: 283–329.Camburn, K.E., Lowe, R.L. & Stoneburner, D.L. (1978) The haptobenthic diatom flora of Long Branch Creek, South
Carolina. Nova Hedwigia 30: 149–279.Cleve, P.T. (1894) Synopsis of the Naviculoid Diatoms, Part I. Kongliga Svenska-Vetenskaps Akademiens Handlingar
26: 1–194.Cleve, P.T. & Grunow, A. (1880) Beiträge zur Kenntniss der Arctischen Diatomeen. Kongliga Svenska-Vetenskaps
Akademiens Handlingar 17: 1–121.Cleve-Euler, A. (1895) On recent freshwater Diatoms from Lule Lappmark in Sweden. Bihang till Kongliga Svenska
Vetenskaps-Akademiens Handlingar 21: 1–44.Cleve-Euler, A. (1955) Die Diatomeen von Schweden und Finnland. Part IV. Biraphideae 2. Kongliga Svenska
Vetenskaps-Akademiens Handligar, ser. 4, 5: 1–232.Cox, E.J. (1987) Placoneis Mereschkowsky: the re-evaluation of a diatom genus originally characterized by its
chloroplast type. Diatom Research 2: 145–157.Cox, E.J. (1988) Taxonomic studies on the diatom genus Navicula Bory. V. The establishment of Parlibellus gen. nov.
for some members of Navicula sect. Microstigmaticae. Diatom Research 3: 9–38.Cox, E.J. (2001) What constitutes a stauros? A morphogenetic perspective. In Jahn, R., Witkowski, A. & Compère, P.
(Eds), Festschrift für H. Lange-Bertalot . Rugell: A.R.G. Gantner Verlag, pp. 303–316.Cox, E.J. (2003) Placoneis Mereschkowsky (Bacillariophyta) revisited: resolution of several typification and
nomenclatural problems, including the generitype. Botanical Journal of the Linnean Society 141: 53–83.Cvetkoska, A. (2011) Lake Ohrid diatoms as palaeoclimate indicators of climate change during the last glacial–
interglacial cycle. Msc thesis, Faculty of Natural Sciences, Skopje, 220 pp. Donkin, A.S. (1871) The Natural History of the British Diatomaceae. Part 2. London: Van Voorst, pp. 25–48. Edlund, M.B. (1994) Additions and confirmations to the algal flora of Itasca State Park. II. Diatoms from Chambers
Creek. Journal of the Minnesota Academy of Science 59: 10–21.Foged, N. (1977) Freshwater diatoms in Ireland. Bibliotheca Phycologica 34: 1– 220.Foged, N. (1982) Diatoms in Bornholm, Denmark. Bibliotheca Phycologica 59: 1–175.Frenguelli, J. (1941) Diatomeas del Río de la Plata. Revista del Museo de la Plata, Nueva Serie, Sección Botánica 3:
213–334.Frenguelli, J. (1945) El Platense y sus diatomeas, Las diatomeas del Platense. Revista del Museo de La Plata (Nueva
Serie), Seccion Paleontologia 3: 77–221.Gallik, O. (1935) A Navicula crucicula (W. Smith) Donkin, valamint nehany rokona elofordulasa a Balatonban es a
Hevizben. Pannonhalmi Szemle 10: 64–66.Greville, R.K. (1833) Diatomaceae. In: William Jackson Hooker (Ed.), The English Flora of Sir J.E. Smith. Volume 5.
London: Longman, Rees, Orme, Brown, Green and Longman, pp. 401–415.Grunow, A. (1878) Algen und Diatomaceen aus dem Kaspischen Meere. In: Schneider, O. (Ed), Naturwissenschafte
Beiträge zur Kenntniss der Kaukasusländer, auf Grund seiner Sammelbeute, Dresden: Verlage der Burdach'schen
Phytotaxa 30 © 2011 Magnolia Press • 37FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Hofbuchhandlung, pp. 98–132.Grunow, A. (1882) Beiträge zur Kenntniss der Fossilen Diatomeen Österreich–Ungarns. In: E. von Mojsisovics und N.
Neumayr (eds), Beiträge zur Paläontologie Österreich–Ungarns und des Orients, 2(4), pp. 136—159, Wien: Alfred Hölder, K.K. hof- und Universitäts Buchhändler.
Gutwinski, R. (1909) Flora Algarum Montium Tetransium. Bulletin de l'Academie des Sciences de Cracovie, Classe des sciences mathematiques et naturelles 1: 415–560.
Hendey, N.I. (1937) The plankton diatoms of the Southern Seas. Discovery Reports 16: 151–364. Héribaud, J. (1903) Les Diatomées Fossiles d'Auvergne (Second Mémoire). Paris: Librairie des Sciences Naturelles, 166
pp. Hustedt, F. (1927a) Fossile bacillariaceen aus dem Loa-Becken in der Atacama-Wüste, Chile. Archiv für Hydrobiologie
18: 224–251.Hustedt, F. (1927b) Bacillariales aus dem Aokikosee in Japan. Archiv für Hydrobiologie 18: 155–172. Hustedt, F. (1930) Bacillariophyta (Diatomeae). In: Pascher, A. (Ed.), Die Süsswasser-Flora Mitteleuropas (Zweite
Auflage), 10, Gustav Fischer, Jena. 466 pp.Hustedt, F. (1931) Die Kieselalgen Deutschlands, Österreichs und der Schweiz unter Berücksichtigung der übrigen
Länder Europas sowie der angrenzenden Meeresgebiete. In: Rabenhorst, L. (ed.), Kryptogamen Flora von Deutschland, Österreich und der Schweiz 7 (Teil 2, Lief. 1), Leipzig: Akademische Verlagsgesellschaft, 176 pp.
Hustedt, F. (1937) Systematische und ökologische Untersuchungen über die Diatomeen-Flora von Java, Bali und Sumatra nach dem Material der Deutschen Limnologischen Sunda-Expedition. “Tropische Binnengewässer, Band VII“. Archiv für Hydrobiologie, Supplement 15: 187–295.
Hustedt, F. (1942) Süßwasser-Diatomeen des indomalayischen Archipels und der Hawaii-Inslen. Internationale Revue der gesamten Hydrobiologie und Hydrographie 42: 1–252.
Hustedt, F. (1944) Neue und wenig bekannte Diatomeen. Bericht der Deutschen Botanischen Gessellschaft 61: 271–290. Hustedt, F. (1945) Diatomeen aus Seen und Quellgebieten der Balkan-Halbinsel. Archiv für Hydrobiologie 40: 867–973.Hustedt, F. (1948) Die Diatomeenflora diluvialer Sedimente bei dem Dorfe Gaj bei Konin im Warthegebiet.
Schweizerische Zeitschrift für Hydrologie 11: 181–209.Hustedt, F. (1957) Die Diatomeenflora des Flußsystems der Weser im Gebiet der Hansestadt Bremen. Abhandlungen des
Naturwissenschaftlichen Verein zu Bremen 34: 181–440.Hustedt, F. (1961–1966) Die Kieselalgen Deutschlands, Österreichs und der Schweiz unter Berücksichtigung der übrigen
Länder Europas sowie der angrenzenden Meeresgebiete. In: Rabenhorst, L. (ed.), Kryptogamen Flora von Deutschland, Österreich und der Schweiz 7 (Teil 3, Lief. 1), Leipzig: Akademische Verlagsgesellschaft, 160 pp.
Jurilj, A. (1948). Nove dijatomeje – Surirellaceae – iz Ohridskog jezera i njihovo filogenetsko znaćenje [New Diatoms – Surirellaceae – of Ochrida Lake in Yugoslavia and their phylogenetic significance]. Prirodoslovnih istraživanja 24: 171–260.
Jurilj, A. (1954) Flora and vegetation of Diatoms from Ochrida Lake in Yugoslavia. Prirodoslovnih istraživanja 26: 99–190.
Karajeva, N.I. (1978) A new suborder of diatoms. Botanicheskii Zhurnal 63: 1747–1750.Kobayasi, H. & Ando, K. (1975) Diatoms from Hozoji-numa, Jizoin-numa and Nakashindennuma ponds in Hanyu City,
Saitama Prefecture. Bulletin of Tokyo Gakugei University, Series 4, Mathematics and Natural Sciences 27: 178–204.Kociolek, J.P. & Thomas, E.W. (2010) Taxonomy and ultrastructure of five naviculoid diatoms (class Bacillariophyceae)
from the Rocky Mountains of Colorado (USA), with the description of a new genus and four new species. Nova Hedwigia 90: 195–214.
Korniushin, A. V. (2004) The bivalve mollusk fauna of ancient lakes in the context of the historical biogeography of the Balkan region. In: Griffith, H. I., et al. (eds.), Balkan Biodiversity–Pattern and Process in the European Hotspot, Dordrecht, Boston, London: Kluwer Academic Publishers, pp. 219–241.
Krammer, K. (1990) Zur identitat von Cocconeis diminuta Pantocsek und Cocconeis thumensis A. Mayer. In: Ricard, M. (ed.) Ouvrage dédié à la Mémoire du Professeur Henry Germain (1903–1989). Königstein: Koeltz Scientific Books, pp. 145–156.
Krammer, K. (1997) Die cymbelloiden Diatomeen – Eine Monographie der weltweit bekannten Taxa. Teil 1. Allgemeines und Encyonema Part. Bibliotheca Diatomologica 36: 1–382.
Krammer, K. (1999) Validierung von Cymbopleura nov. gen. In: Lange–Bertalot, H. (ed.), Iconographia Diatomologica6. Königstein: Koeltz Scientific Books, p. 292.
Krammer, K. (2003) Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. In: Lange-Bertalot, H. (ed.), Diatoms of Europe. Diatoms of the European inland waters and comparable habitats 4, Ruggel: A.R.G. Gantner Verlag, 530 pp.
Krammer, K. & Lange-Bertalot, H. (1985) Naviculaceae Neue und wenig bekannte Taxa, neue Kombinationen und Synonyme sowie Bemerkungen zu einigen Gattungen. Bibliotheca Diatomologica 9: 5–230.
Krammer, K. & Lange-Bertalot, H. (1986) Bacillariophyceae, 1. Teil: Naviculaceae. In: Ettl, H., Gerloff, J., Heying, H. & Mollenhauer, D. (eds.), Süßwasserflora von Mitteleuropa 2/1, Stuttgart: Gustav Fischer, 876 pp.
LEVKOV & WILLIAMS38 • Phytotaxa 30 © 2011 Magnolia Press

Krammer, K. & Lange-Bertalot, H. (1991) Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In: Ettl, H., Gerloff, J., Heying, H. & Mollenhauer, D. (eds.): Süßwasserflora von Mitteleuropa 2/3, Jena: Gustav Fisher Verlag, 576 pp.
Krstic, S., Levkov Z. & Nakov, T. (2006) Diatom diversity in Republic of Macedonia – our present knowledge. In: Witkowski, A. (Ed.), Proceedings of the 18th International Diatom Symposium. Bristol: Biopress Limited, pp. 209–220.
Kützing, F.T. (1844) Die Kieselschaligen. Bacillarien oder Diatomeen. Nordhausen: Förstemann, 152 pp. Lange-Bertalot, H. (1980) Ein Beitrag zur Revision der Gattungen Rhoicosphenia Grun., Gomphonema C. Ag.,
Gomphoneis Cl. Botaniska Notiser 133: 585–594.Lange-Bertalot, H. (1993) 85 neue taxa und über 100 weitere neu definierte Taxa ergänzend zur Süsswasserflora von
Mitteleuropa, Vol. 2/1–4. Bibliotheca Diatomologica 27: 1–164. Lange-Bertalot, H. (1995) Gomphosphenia paradoxa nov. spec. et nov. gen. und Vorschlag zur Lösung taxonomischer
Probleme infolge eines veränderten Gattungskonzepts von Gomphonema (Bacillariophyceae). Nova Hedwigia 60: 241–252.
Lange-Bertalot, H. (2001) Navicula sensu stricto, 10 genera separated from Navicula sensu lato, Frustulia. In: Lange-Bertalot, H. (Ed.), Diatoms of Europe. Diatoms of the European inland waters and comparable habitats 2, Ruggel: A.R.G. Gantner Verlag, 526 pp.
Lange-Bertalot, H. & Krammer, K. (1989) Achnanthes eine Monographie der Gattung mit Definition der Gattung Cocconeis und Nachtragen zu den Naviculaceae. Bibliotheca Diatomologica 18: 1–393.
Lange-Bertalot, H., Rumrich, U. & Hofmann, G. 1991. Zur Revision der Gattung Diatoma Bory (Subgenus Diatoma, Bacillariophyceae). Identifikation okologisch wichtiger, aber taxonomisch problematischer Arten. Acta Biologica Benrodis 3: 115–130.
Levkov, Z. (2009) Amphora sensu lato. In: Lange–Bertalot, H. (Ed.): Diatoms of Europe. Diatoms of the European inland waters and comparable habitats 5. Ruggel: A.R.G. Gantner Verlag, 918 pp.
Levkov, Z. & Nakov, T. (2008) Rhoicosphenia tenuis, a new diatom species from Lake Ohrid. Diatom Research 23: 377–388.
Levkov, Z. & Williams, D.M. (2006) Two new species of Diatoma from Lakes Ohrid and Prespa, Macedonia. Diatom Research 21: 281–296.
Levkov Z., Nakov, T. & Metzeltin, D. (2006a) New species and combination from genus Sellaphora Mereschkowsky (Bacillariophyceae) from Macedonia. Diatom Research 21: 297–312.
Levkov, Z., Krstic, S. & Nakov T. (2006b) A new Gyrosigma species from Lakes Prespa and Ohrid, Republic of Macedonia. In: Ognjanova–Rumenova, N. & Manoylov, K. (eds.), Fossil and Recent Phycological Studies. Sofia: Pensoft, pp. 139–145.
Levkov, Z., Krstic, S., Metzeltin, D. & Nakov, T. (2007) Diatoms of Lakes Prespa and Ohrid (Macedonia). Iconographia Diatomologica 16. Rugell, A.R.G. Gantner Verlag, 603 pp.
Lyngbye, H.C. (1819) Tentamen Hydrophytologiae Danicae Continens omnia Hydrophyta Cryptogama Daniae, Holsatiae, Faeroae, Islandiae, Groenlandiae hucusque cognita, Systematice Disposita, Descripta et iconibus illustrata, Adjectis Simul Speciebus Norvegicis. Copenhagen, 248 pp.
Mayer, A. (1913) Die Bacillariaceen der Regensburgen Gewasser. Systematischer Teil. Berichte des naturwissenschaftlichen Vereines zu Regensburg 14: 1–364.
Mayer, A. (1919) Bacillariales von Reichenhall und Umgebung. Kryptogamische Forschungen herausgegeben von der Kryptogamenkommission der Bayerischen Botanischen Gesellschaft zur Erforschung der heimischen Flora 1: 191–216.
Mayer, A. (1928) Die bayerischen Gomphonemen. Denkschriften der Koniglich-Baierischen Botanischen Gesellschaft in Regensburg 17: 83–128.
Medlin, L.K. & Kaczmarska, I. (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245–270.
Mereschkowsky, C. (1903) Uber Placoneis, ein neues Diatomeen-Genus. Beihefte zum Botanischen Centralblatt 15: 1–30.
Metzeltin, D., Lange-Bertalot, H. & García-Rodríguez, F. (2005) Diatoms of Uruguay. Compared with other taxa from South America and elsewhere. Iconographia Diatomologica 15. Rugell, A.R.G. Gantner Verlag, 736 pp.
Miho, A. & Lange-Bertalot, H. (2006) Diversity of the genus Placoneis in Lake Ohrid and other freshwater habitats in Albania. In: Witkowski, A. (Ed.), Proceedings of the 18th International Diatom Symposium. Bristol: Biopress Limited, pp. 301–313.
Moser, G., Lange-Bertalot, H. & Metzeltin, D. (1998) Insel der Endemiten Geobotanisches Phänomen Neukaledonien. Bibliotheca Diatomologica 38: 1– 464.
Østrup, E. (1908) Beiträge zur Kenntnis der Diatomeenflora des Kossogolbeckens in der nordwestlichen Mongolei. Hedwigia 48: 74–100.
Pantocsek, J. (1889) Beiträge zur Kenntnis der Fossilen Bacillarien Ungarns. Teil II. Brackwasser Bacillarien. Anhang:
Phytotaxa 30 © 2011 Magnolia Press • 39FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES

Analyse de marine Depots von Bory, Bremia, Nagy-Kurtos in Ungarn; Ananio und Kusnetzk in Russland. Nagy-Tapolcsány: Buchdrucherei von Julius Platzko, 123 pp.
Pantocsek, J. (1892) Beiträge zur Kenntnis der Fossilen Bacillarien Ungarns. Teil III, Susswasser Bacillarien. Anhang–analysen 15 neuer Depots von Bulgarien, Japan, Mahern, Russland und Ungarn. Nagy-Tapolcsány: Buchdrucherei von Julius Platzko, pl. I—XLII.
Pantocsek, J. (1902) Kieselalgen oder Bacillarien des Balaton. Resultate der Wissenschaftlichen Erforschung des Balatonsees, herausgegeben von der Balatonsee–Commission der Ung. Geographischen Gesellschaft 2: 1–112.
Patrick, R.M. & Freese, L.R. (1961) Diatoms (Bacillariophyceae) from Northern Alaska. Proceedings of the Academy of Natural Sciences of Philadelphia 112: 129–293.
Patrick, R.M. & Reimer, C.W. (1966) The Diatoms of the United States exclusive of Alaska and Hawaii. Volume 1. Fragilariaceae, Eunotoniaceae, Achnanthaceae, Naviculaceae. Philadelphia: Monographs of the Academy of Natural Sciences of Philadelphia vol 13, 688 pp.
Rabenhorst, L. (1864) Flora Europaea Algarum aquae dulcis et submarinae. Sectio I. Algas diatomaceas complectens, cum figuris generum omnium xylographice impressis. Leipzig: Apud Eduardum Kummerum, 359 pp.
Radoman, P. (1983) Hydrobioidea a superfamily of Prosobranchia (Gastropoda), I. Systematics. Belgrade: Serbian Academy of Sciences and Arts, 256 pp.
Radoman, P. (1985) Hydrobioidea a superfamily Prosobranchia (Gastropoda), II. Origin, Zoogeography, Evolution in the Balkans and Asia Minor. Monographs Institute of Zoology 1, Belgrade: Pensoft, 256 pp.
Reed, J. M., Cvetkoska, A, Levkov, Z., Vogel, H &. Wagner, B. (2010) The last glacial-interglacial cycle in Lake Ohrid (Macedonia/Albania): testing diatom response to climate. Biogeosciences 7: 1–26.
Reichardt, E. (1982) Die Diatomeenflora der "Steinernen Rinnen" in Mittelfranken. Berichte der Bayerischen Botanischen Gesellschaft 53: 97–112.
Reichardt, E. (1997) Taxonomische Revision des Artenkomplexes um Gomphonema pumilum (Bacillariophyceae). Nova Hedwigia 65: 99–130.
Reichardt, E. (1999) Zur Revision der Gattung Gomphonema. Die Arten um G. affine/insigne, G. angustatum/micropus, G. acuminatum sowie gomphonemoide Diatomeen aus dem Oberoligozan in Bohmen. Iconographia Diatomologica8: 1–203.
Round, F.E., Crawford, R.M. & Mann, D.G. (1990) The diatoms: Biology and Morphology of the genera. Cambridge: Cambridge University Press, 747 pp.
Rumrich, U., Lange-Bertalot, H. & Rumrich, M. (2000) Diatoms of the Andes. From Venezuela to Patagonia/Tierra del Fuego and two additional contributions. Iconographia Diatomologica 9: 1–673.
Schimanski, H. (1978) Beitrag zur Diatomeenflora des Frankenwaldes. Nova Hedwigia 30: 557–634. Schmidt, A. (1874–1959) Atlas der Diatomaceen-Kunde. Leipzig: Aschersleben, 460 pl.Silva, P.C. (1962) Classification of algae. In: Lewin, R.A. (Ed) Physiology and Biochemistry of Algae. New York &
London: Academic Press, pp. 827–837.Simonsen, R. (1987) Atlas and Catalogue of the Diatom Types of Friedrich Hustedt. Berlin & Stuttgart: J. Cramer, 3
volumes, 525 pp., 772 pls. Skvortzow, B.W. (1938) Diatoms from Argun River, Hsing–An–Pei Province, Manchoukuo. Philippine Journal of
Science 66: 43–72.Skvortzow, B.W. (1971) Diatoms from Yenisei River and its tributaries, middle part of Siberia, western Asia. Philippine
Journal of Science 98: 57–113.Skvortzow, B.W. & Meyer, C.I. (1928) A contribution to the Diatoms of Baikal Lake. Proceedings of the Sungaree River
Biological Station 1: 1–55. Stankovic, S. (1960) The Balkan Lake Ohrid and its Living World. Monographiae Biologicae, Vol. IX. The Hague: Dr. W.
Junk, 357 pp. Szabó, K.E., Kiss, K.T., Tóth, B., Metzeltin, D. & Ács, E. (2007) Navicula novaesiberica Lange-Bertalot; a new record
of the species from the Danube River in Hungary. Diatom Research 22: 497–502.Thomas, E.W., Kociolek, J.P., Lowe, R.L. & Johansen, J.R. (2009). Taxonomy, ultrastructure and distribution of
Gomphonemoid diatoms (Bacillariophyceae) from Great Smoky Mountains National Park (U.S.A.). In: Kociolek, J.P., Theriot, E.C. & Stevenson, R.J. (eds.), Diatom taxonomy, ultrastructure and ecology: modern methods and timeless questions. A tribute to Eugene F. Stoermer. Nova Hedwigia, Beiheft 135: 201–237.
Tuji, A. & Williams, D.M. (2006a) Typification of Conferva pectinalis O. F. Müll. (Bacillariophyceae) and the identity of the type of an alleged synonym, Fragilaria capucina Desm. Taxon 55: 193–197.
Tuji, A. & Williams, D.M. (2006b) Examination of the type material of Synedra rumpens = Fragilaria rumpens, Bacillariophyceae. Phycological Research 54: 99–103.
Tuji, A. & Williams, D.M. (2008a) Examination of types in the Fragilaria pectinalis–capitelata species complex. In: Likhoshway (ed.), Proceedings of 19th International Diatom Symposium. Bristol: Biopress Limited, pp. 125–139.
Tuji, A. & Williams D.M. (2008b) Typification and type examination of Synedra familiaris Kütz. and related taxa. Diatom 24: 25–29.
LEVKOV & WILLIAMS40 • Phytotaxa 30 © 2011 Magnolia Press

Van Heurck, H. (1880) Synopsis des Diatomées de Belgique. Atlas. Antwerp: Ducaju & Cie., 10 pls.Van Heurck, H. (1896) A Treatise on the Diatomaceae. Translated by W.E. Baxter. London: William Wesley & Son, 558
pp.Wilke, T., Albrecht, C., Anistratenko, V.V., Sahin, S.K., & Yildirim, Z. (2007) Testing biogeographical hypotheses in
space and time: Faunal relationships of the putative ancient lake Egirdir in Asia Minor. Journal of Biogeography 34: 1807–1821.
Williams, D.M. (1985) Morphology, taxonomy and interrelationships of the ribbed araphid diatoms from the genera Diatoma and Meridion (Diatomaceae: Bacillariophyta). Bibliotheca Diatomologica 8: 1–228.
Williams, D.M. & Round, F.E. (1987) Revision of the genus Fragilaria. Diatom Research 2: 267–288. Witkowski, A., Lange-Bertalot, H. & Metzeltin, D. (2000) Diatom Flora of Marine Coasts I. Iconographia
Diatomologica 7, 925 pp.
Phytotaxa 30 © 2011 Magnolia Press • 41FIFTEEN NEW DIATOM (BACILLARIOPHYTA) SPECIES





























