Embed Size (px)
Citation preview

©
New Phytologist
(2001)
152
: 291–299
www.newphytologist.com
291
Research
Blackwell Science Ltd
Flooding induces a suite of adaptive plastic responses
in the grass
Paspalum dilatatum
P. Insausti
1
, A. A. Grimoldi
1
, E. J. Chaneton
1
and V. Vasellati
2
1
IFEVA – CONICET, Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453, CPA 1417 DSE, Buenos Aires, Argentina;
2
Cátedra de Botánica Agrícola, Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453, CPA 1417 DSE, Buenos Aires,
Argentina
Summary
• We studied individual responses to flooding in the perennial grass
Paspalumdilatatum
, a widespread species in the Flooding Pampa of Argentina, using plantsestablished in species-diverse grassland microcosms.• Flooding effects were evaluated on root and leaf sheath anatomy and shootmorphological traits. Leaf water status and CO
2
exchange rates were monitored inflooded and unflooded plants under changing, natural and controlled atmosphericconditions.• Root porosity and leaf sheath aerenchyma increased with flooding. Leaf extensionrates and tiller height were also higher in flooded plants, which resulted in a largefraction of the shoot architecture emerging above the water surface. Floodingenhanced stomatal conductance, leaf water potential and net photosynthesis, espe-cially under conditions leading to high air-vapour pressure deficits. Therefore,flooded plants experienced fewer water deficits during periods of high atmosphericevaporative demand.•
P. dilatatum
showed tight regulation of water and carbon relations under severesoil-oxygen deficiency, even in the presence of natural competitors. The suite ofadaptive responses documented here might help to explain the observed increase inabundance of this species during extensive floods.
Key words:
aerenchyma, flooding tolerance, leaf growth, Pampa grasslands,
Paspalum dilatatum
, photosynthesis, tiller architecture, water relations.
©
New Phytologist
(2001)
152
: 291–299
Author for correspondence:
P. Insausti
Tel:
+
54 011 4524 8071
Fax:
+
54 011 4514 8730
Email: [email protected]
Received:
9 March 2001
Accepted:
17 July 2001
Introduction
The potential for a plant species to persist within a multi-species community depends to a large extent on its ability tocope with prevailing abiotic constraints in the presence ofneighbours (Weiher & Keddy, 1995; Bazzaz, 1996). Floodingacts as a powerful constraint on plant productivity and speciescomposition in many natural habitats (Pennings & Callaway,1992; Weiher & Keddy, 1995; Trebino
et al.
, 1996; Gough &Grace, 1998). Whereas severe flooding typically causes areduction in the abundance of flood-sensitive plant species, itcan also promote biomass growth in flood-tolerant species(Insausti
et al.
, 1999). In general, endurance of prolonged soilwaterlogging may allow resistant plants to exploit resourcesthat otherwise would be shared with nontolerant competitors
(Crawford
et al.
, 1989). Thus, in order to predict plant spe-cies dynamics in intermittently flooded habitats, a primaryunderstanding of individual-based adaptations to tolerate oravoid waterlogging stress is needed (Osmond
et al.
, 1987;Crawford
et al.
, 1989; Blom
et al.
, 1990).Plants respond to flooding through changes at molecular,
cell, tissue, organ and individual levels of organization. Inflood-intolerant plant species, the lack of soil oxygen triggersa chain process that often results in severe physiological injury,as plant cells require oxygen for their normal metabolism(Jackson & Drew, 1984). In flood-tolerant plants, resistanceto soil-oxygen deficiency can be achieved through variousmechanisms, including anatomical, morphological, physio-logical and life-history adaptations (Blom
et al.
, 1990; Pezeshki,1994; Blom & Voesenek, 1996). Most flood-tolerant species
NPH_257.fm Page 291 Thursday, September 27, 2001 5:19 PM

www.newphytologist.com
©
New Phytologist
(2001)
152
: 291–299
Research292
are capable of developing root aerenchyma in order toavoid tissue anoxia under waterlogging conditions ( Justin &Armstrong, 1987; Blom
et al.
, 1990). The elongation of stemsand leaves is another plastic response commonly observed inflood-tolerant plants (Laan & Blom, 1990; Blom & Voesenek,1996). Shoot extension and aerenchyma formation are bothimportant for restoring the plant’s contact with the atmos-phere, and hence allowing internal oxygen transport to sub-merged organs during flooding periods (Blom & Voesenek,1996).
In addition to these anatomical and morphological changes,flooding often disrupts plant water relations and gas exchangeprocesses. Flood-intolerant species often show leaf dehydra-tion in waterlogged soils, because oxygen deficiency reducesthe cell-membrane permeability of the roots (Pezeshki, 1994;Tyerman
et al.
, 1999). Many plant species also experience rapiddeclines in stomatal conductance when flooded (Kozlowski &Pallardy, 1984; Else
et al.
, 1995). Since stomata closure causesdiffusional limitation of carbon uptake, intolerant plants mayhave their photosynthetic capacity drastically reduced duringextended floods (Kozlowski & Pallardy, 1984). Furthermore,flood-sensitive species can exhibit nonstomatal inhibitionof photosynthesis, a process thought to be associated withalterations in plant hormone concentrations induced by tissueanoxia (Pezeshki, 1994). By contrast, flood-tolerant speciesmay show little change, or even an increase, in carbon uptakerates during waterlogging periods (Naidoo & Naidoo, 1992).For example, the grass
Sporobolus virginicus
shows lowerphotosynthetic rates in well-drained soils than in floodedones, as a result of increased levels of water stress (Naidoo &Naidoo, 1992). A tight regulation of the plant water status isthus required to maintain photosynthesis, nutrient uptakeand growth in flooded environments (Kozlowski & Pallardy,1984). Ultimately, flooding tolerance will reflect a suite ofstructural and physiological adaptations enabling the plant toremain functional under soil-oxygen deprivation (Jackson &Drew, 1984; Kozlowski & Pallardy, 1984; Blom
et al.
, 1990;Blom & Voesenek, 1996).
In this paper, we investigate the effects of flooding onanatomical, morphological and physiological traits of theperennial grass
Paspalum dilatatum
, a common warm-seasonspecies in periodically flooded grasslands of the ArgentinePampas.
P. dilatatum
exhibits high resistance to waterlogg-ing when grown in isolation (Rubio
et al.
, 1995; Loreti &Oesterheld, 1996) and the capacity to increase in abundancewithin flooded plant communities (Insausti
et al.
, 1999). How-ever, little is known about the integration of morphological andphysiological traits allowing this native grass to thrive undersevere flooding in the presence of neighbours. Moreover,although previous work on
P. dilatatum
revealed patterns ofbiomass allocation and nutrient uptake indicative of floodtolerance (Rubio
et al.
, 1995; Loreti & Oesterheld, 1996;Rubio & Lavado, 1999), there has been no attempt to assesshow plants may adjust water and carbon relations to changing
atmospheric conditions during extended floods. Our aim herewas to evaluate individual responses to flooding compatiblewith the behaviour of
P. dilatatum
in natural plant commu-nities. We hypothesized that a combination of anatomical,morphological and physiological responses minimizes thestress by flooding in this species. Specifically, we predictedthat
P. dilatatum
would make better use of available waterin flooded than in drained soils kept at field capacity. Thus,flooded plants would not develop substantial water deficitsunder conditions of high atmospheric evaporative demand.
Materials and Methods
Plant material and experimental design
Paspalum dilatatum
Poir. is a conspicuous component oftemperate humid grasslands of the Flooding Pampa in easternArgentina. This native species is widely distributed across topo-graphical gradients from upland communities on well-drainedsandy soils, to lowland areas where various plant associationsoccur on annually flooded, fine-textured soils (Burkart
et al.
,1990).
P. dilatatum
is a highly productive C
4
grass with anextended growing season ranging from late spring, throughsummer to early autumn (Sala
et al.
, 1981). We conducted anoutdoor microcosm experiment using plants of
P. dilatatum
naturally established in soil monoliths extracted from a lowlandgrassland site (near Pila, Buenos Aires Province, Argentina).The use of microcosms helped overcome most problemsconcerned with manipulating soil waterlogging in the field,while it also allowed us to monitor individual plant responsesin the presence of natural neighbours (Grimoldi
et al.
, 1999)
.
Twenty plants of
P. dilatatum
of similar size and prostratearchitecture were obtained during spring from a lowlandgrassland site grazed year-round by cattle. Each target plantwas located at the centre of a 30
×
30
×
40 cm soil monolithsupporting intact vegetation. The grassland was unflooded atthe time of monolith extraction. Because of the restrictedgrowth form of
P. dilatatum
, each plant was thought torepresent a different clone within the same local population.Monoliths were transported to an experimental garden, wherethey were placed in plastic containers to manipulate artificialflooding. After a one-month acclimation period, monolithswere randomly assigned to two treatments: control – mono-liths watered daily to field capacity and allowed to drain freelythrough holes drilled in the container; flooding – monolithsinundated with tap water and kept waterlogged to 6-cmheight above soil surface (intact container) until the end of theexperiment. Treatments ran for 45 d from late spring to earlysummer, and were replicated 10 times.
Leaf anatomy and root porosity
Root anatomy and aerenchyma formation in
P. dilatatum
were described elsewhere (Rubio
et al.
, 1995). Therefore, we
NPH_257.fm Page 292 Thursday, September 27, 2001 5:19 PM

©
New Phytologist
(2001)
152
: 291–299
www.newphytologist.com
Research 293
focus here on changes in leaf anatomy as induced by flooding.Segments of leaf sheath from the portion located 1 cm belowthe ligule were taken from each
P. dilatatum
plant at the endof the experiment, and were preserved in a FAA solution forlater microscopical analysis. Leaf sheath sections
c
. 20 µmthick were cut freehand, stained with 50% water-solublesafranine and mounted in gelatine-glycerine. Similar leafsheath segments were embedded in paraffin; longitudinalsections from the keel portion were cut with a rotatory micro-tome and stained with safranine-fast green. Five sections ofleaf sheath were randomly selected from each plant forobservation. Percentage aerenchymatous space (i.e. the ratiobetween the area occupied by aerenchyma and the totalcross-sectional area) in the leaf sheath was measured usingan optical microscope (Zeiss Axioplan, Zeiss, Oberkochen,Germany) connected to an image analyser (Imagenation Px,Imagenation Corp., Beaverton, OR, USA). In addition, the meanlength of parenchymatous cells present along longitudinalsections of leaf sheath was measured under the opticalmicroscope. The number of parenchymatous cells was evaluatedfor the same row used to measure cell length in leaf sheathsections of flooded and unflooded plants. The focal row ofcells occurred between vascular bundles of identical size, andat the same depth in the sheath parenchyma, for the plants ofboth treatments. Microphotographs of this material weretaken through the Zeiss Axioplan microscope. Measurementswere performed on five randomly selected plants from eachtreatment, at the beginning of the experiment and after 45 dof flooding. Gas-filled root porosity was quantified for freshsamples of young roots taken from five plants per treatmentusing the pycnometer method (Sojka, 1988).
Shoot morphology
The leaf extension rate was recorded for all control andflooded plants over a one month period. Measurements of leaflength were taken to the nearest 0.1 cm on the youngestvisible leaf (3rd or 4th, according to day of experiment) ofthree tillers per plant; each leaf was followed until the ligulewas exposed. In addition, we measured the mean leaf bladeand sheath length, and the mean tiller insertion angle andheight (relative to the soil surface) of all
P. dilatatum
plants atthe beginning of the experiment, after 10 days and at the endof the study. These measurements were performed on thethird leaf (with ligule exposed) of 10 tillers per plant.
Stomatal conductance and leaf water potential
Stomatal conductance (g
s
) was measured using a Li-1600 Msteady state porometer (Li-Cor Inc., Lincoln, NE, USA). Thetwo youngest fully expanded leaves of each plant were selectedfor measurement. Leaf water potential (
ψ
l
) was recorded onthe same leaves using a PI-6 Schölander-type pressure chamber(Bio-Control, Buenos Aires, Argentina);
ψ
l
was measured
immediately after completing the g
s
measurements. Both
ψ
l
and g
s
were recorded approximately every second day, simultan-eously with measures of leaf elongation. Air temperature andrh were monitored (Temperature & RH Probe HUMICAPH, Vaisala, Finland) and used to calculate the air vapourpressure deficit (VPD
air
) at the time of each set of g
s
and
ψ
l
measurements. The ambient photosynthetic photon fluxdensity (PPFD, over 400–700 nm) was recorded on each leafsimultaneaously with g
s
measurements using a point quantumsensor (Li-Cor Inc., Lincoln, NE, USA).
Gas exchange measurements
After 45 days under natural atmospheric conditions, eight micro-cosms from each treatment were moved into a temperature-humidity controlled glasshouse. The original treatments weremaintained to examine how flooding affected the capacityof
P. dilatatum
plants to regulate gas exchange rates underdifferent, controlled levels of atmospheric evaporative demand.Leaf stomatal conductance, transpiration rate and net photo-synthesis (P
n
) were recorded in flooded and control plantsat low, medium and high VPD
air
conditions. We modifiedexperimentally the air temperature and humidity within theglasshouse to create a controlled gradient of evaporative demand(range, 0.5–4 kPa), and then measured plant parameters atthree time steps corresponding to increasing levels of VPD
air
.Air vapour pressures were calculated from the rh and temper-ature conditions established in the glasshouse. The experimentwas conducted over three consecutive days; this scheduleallowed the plants to equilibrate to each new VPD
air
levelbefore measurement. Photosynthesis was measured as net CO
2
exchange rate using a Li-6250 portable system (Li-Cor Inc.).For each plant, P
n
measurements were taken on a portionof the youngest fully expanded leaf placed in a 250-cm
3
chamber, with additional lighting from a 1500-µmol m
–2
s
–1
(PPFD) light source. The same leaves were used to record thetranspiration rate, g
s
, and leaf temperature of flooded andunflooded plants at each VPD
air
condition.
Statistical analyses
Anatomical variables were analysed using one-way ANOVA(
n
= 5); percentage data were arcsine square-root transformedbefore analysis (Steel & Torrie, 1988). Differences in shootmorphological traits (leaves and tillers) between control andflooded plants were assessed by two-way ANOVA (
n
= 10),with flooding treatment and measurement date (10 vs 45 d)as main factors. Tiller insertion angle and height data weresquare-root transformed to meet ANOVA assumptions. Effectsof flooding on daily leaf elongation rates and physiologicalparameters measured outdoors (
n
= 10) were evaluated throughrepeated-measures ANOVA. The Greenhouse-Geyser correc-tion was used to adjust for possible violations of the sphericityassumption (Von Ende, 1993). The relationship between
NPH_257.fm Page 293 Thursday, September 27, 2001 5:19 PM
www.newphytologist.com
©
New Phytologist
(2001)
152
: 291–299
Research294
physiological parameters and atmospheric conditions (VPD
air
)during the course of the outdoor experiment was examined bysimple regression analysis, using the absolute difference inmean daily values of g
s
and
ψ
l
between flooded and controlplants as the response variable. For the glasshouse experiment,differences in physiological parameters of flooded and con-trol plants (
n
= 8) at increasing VPD
air
levels were tested byrepeated-measures ANOVA (Potvin & Lechowicz, 1990; VonEnde, 1993). In these analyses, the flooding treatment was thebetween-subject factor and the level of VPD
air
represented thewithin-subject factor (Potvin & Lechowicz, 1990). All resultsare presented as mean
±
standard error (SE).
Results
Anatomical responses
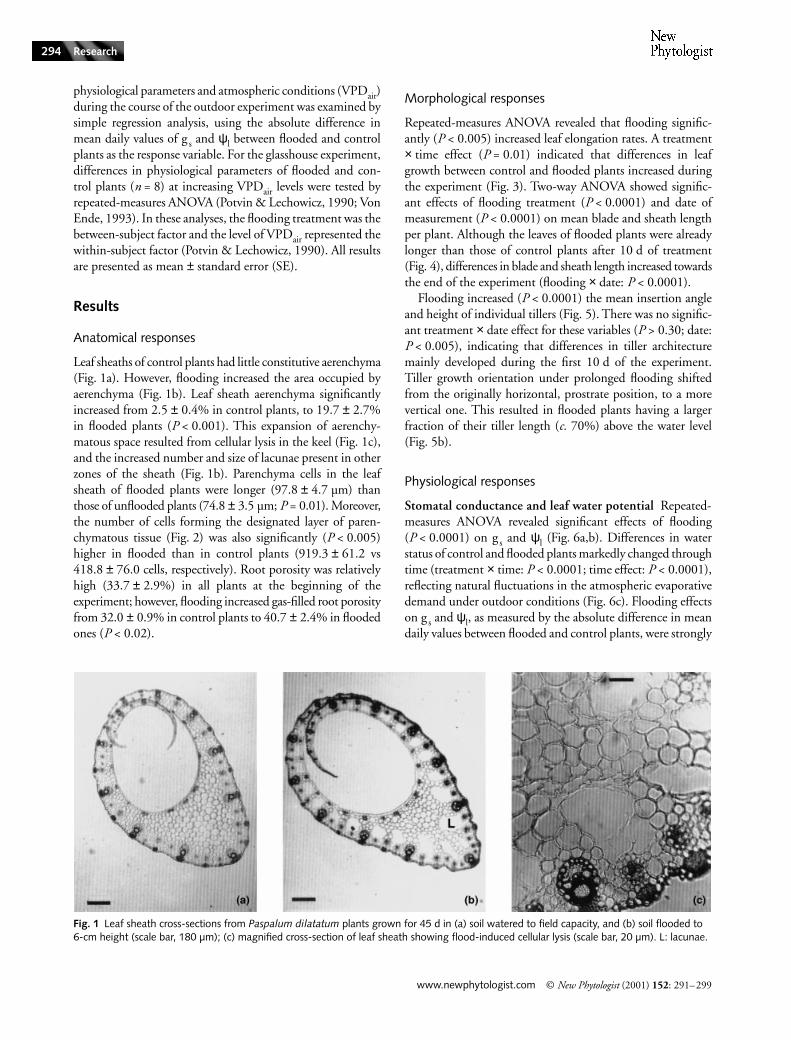
Leaf sheaths of control plants had little constitutive aerenchyma(Fig. 1a). However, flooding increased the area occupied byaerenchyma (Fig. 1b). Leaf sheath aerenchyma significantlyincreased from 2.5
±
0.4% in control plants, to 19.7
±
2.7%in flooded plants (
P
< 0.001). This expansion of aerenchy-matous space resulted from cellular lysis in the keel (Fig. 1c),and the increased number and size of lacunae present in otherzones of the sheath (Fig. 1b). Parenchyma cells in the leafsheath of flooded plants were longer (97.8
±
4.7 µm) thanthose of unflooded plants (74.8
±
3.5 µm;
P
= 0.01). Moreover,the number of cells forming the designated layer of paren-chymatous tissue (Fig. 2) was also significantly (
P
< 0.005)higher in flooded than in control plants (919.3
±
61.2 vs418.8
±
76.0 cells, respectively). Root porosity was relativelyhigh (33.7
±
2.9%) in all plants at the beginning of theexperiment; however, flooding increased gas-filled root porosityfrom 32.0
±
0.9% in control plants to 40.7
±
2.4% in floodedones (
P
< 0.02).
Morphological responses
Repeated-measures ANOVA revealed that flooding signific-antly (
P
< 0.005) increased leaf elongation rates. A treatment
×
time effect (
P
= 0.01) indicated that differences in leafgrowth between control and flooded plants increased duringthe experiment (Fig. 3). Two-way ANOVA showed signific-ant effects of flooding treatment (
P
< 0.0001) and date ofmeasurement (
P
< 0.0001) on mean blade and sheath lengthper plant. Although the leaves of flooded plants were alreadylonger than those of control plants after 10 d of treatment(Fig. 4), differences in blade and sheath length increased towardsthe end of the experiment (flooding
×
date:
P
< 0.0001).Flooding increased (
P
< 0.0001) the mean insertion angleand height of individual tillers (Fig. 5). There was no signific-ant treatment
×
date effect for these variables (
P
> 0.30; date:
P
< 0.005), indicating that differences in tiller architecturemainly developed during the first 10 d of the experiment.Tiller growth orientation under prolonged flooding shiftedfrom the originally horizontal, prostrate position, to a morevertical one. This resulted in flooded plants having a largerfraction of their tiller length (
c.
70%) above the water level(Fig. 5b).
Physiological responses
Stomatal conductance and leaf water potential
Repeated-measures ANOVA revealed significant effects of flooding(
P
< 0.0001) on g
s
and
ψ
l
(Fig. 6a,b). Differences in waterstatus of control and flooded plants markedly changed throughtime (treatment × time: P < 0.0001; time effect: P < 0.0001),reflecting natural fluctuations in the atmospheric evaporativedemand under outdoor conditions (Fig. 6c). Flooding effectson g s and ψl, as measured by the absolute difference in meandaily values between flooded and control plants, were strongly
Fig. 1 Leaf sheath cross-sections from Paspalum dilatatum plants grown for 45 d in (a) soil watered to field capacity, and (b) soil flooded to 6-cm height (scale bar, 180 µm); (c) magnified cross-section of leaf sheath showing flood-induced cellular lysis (scale bar, 20 µm). L: lacunae.
NPH_257.fm Page 294 Thursday, September 27, 2001 5:19 PM

© New Phytologist (2001) 152: 291–299 www.newphytologist.com
Research 295
correlated with natural VPDair levels (r = 0.917 and 0.935,both P < 0.0001, n = 21, for g s and ψl, respectively). Onaverage, flooded plants maintained higher values of g s and ψlthan control plants, although this mainly reflected differencesoccurring during periods of high VPDair when control plantssuffered substantial declines in both physiological parameters(Fig. 6).
Gas exchange rates Repeated-measures ANOVA showed highlysignificant effects of flooding (P < 0.0001) and VPDair (P <0.0001) on physiological variables recorded in the glass-house experiment (Fig. 7). Moreover, significant flooding–VPDair interactions (all P < 0.0001) indicated that differencesin gas exchange rates between flooded and control plantsbecame greater with higher levels of water vapour atmospheric
Fig. 2 Longitudinal leaf sheath sections from Paspalum dilatatum plants grown for 45 d under (a) control and (b) flooded conditions. Scale bar, 20 µm. Asterisks indicate the rows of parenchymatous cells taken for measurements of mean cell number and length.
Fig. 3 Cumulative leaf extension in Paspalum dilatatum plants growing under control (open squares) and flooded (closed squares) conditions. Measures were taken for three tillers per plant. Each value shows the mean of 10 plants.
Fig. 4 Length of leaf blade (a) and sheath (b) in Paspalum dilatatum plants growing under control (open columns) and flooded (closed columns) conditions. Measures are for the last fully expanded leaf of 10 tillers per plant. Results show the mean + 1 SE of 10 plants.
Fig. 5 Tiller insertion angle (a) and height (b) relative to soil surface in Paspalum dilatatum growing under control (open columns) and flooded (closed columns) conditions. The dashed line shows the water depth in the flooded treatment. Measures were taken for 10 tillers per plant. Results show the mean + 1 SE of 10 plants.
NPH_257.fm Page 295 Thursday, September 27, 2001 5:19 PM

www.newphytologist.com © New Phytologist (2001) 152: 291–299
Research296
demand. Plants growing in flooded microcosms showedhigher g s, transpiration and Pn than those watered to fieldcapacity, when exposed to high VPDair conditions (Fig. 7a–c).Flooded and control plants did not differ in any of thesevariables under low VPDair levels. These responses wereaccompanied by a significant increase in leaf temperatureacross the VPDair gradient (Fig. 7d). However, notice that athigh levels of VPDair, mean leaf temperature for floodedplants remained 6°C below that recorded for unfloodedcontrols (flooding × VPDair interaction: P < 0.0001).
Discussion
Two processes are critically associated with a plant speciesability to tolerate flooding: internal oxygen transport toprevent anoxia in submerged plant organs, and maintenance
Fig. 6 Stomatal conductance (a) and leaf water potential (b) of Paspalum dilatatum plants growing in control (open squares) and flooded (closed squares) soils under natural atmospheric conditions. (c) Air vapour pressure deficit (VPDa) during the time of experiment. All measurements were taken on clear days under similar ambient photosynthetic photon flux density (PPFD) conditions (c. 1500 µmol m–2 s–1, over 400–700 nm). Results represent the mean ± 1 SE of 10 plants.
Fig. 7 Changes in net photosynthesis (a), stomatal conductance (b), transpiration rate (c) and leaf temperature (d) of Paspalum dilatatum in response to increasing levels of air vapour pressure deficit within a controlled environment glasshouse, for plants grown under control (open squares) and flooded (closed squares) conditions. Results represent the mean ± 1 SE of eight plants.
NPH_257.fm Page 296 Thursday, September 27, 2001 5:19 PM

© New Phytologist (2001) 152: 291–299 www.newphytologist.com
Research 297
of root function to avoid inhibition of water and nutrientsuptake. In periodically flooded habitats, tolerant plant speciesperform these tasks through various adaptive responses,which allow them not only to survive waterlogging periods(Blom et al., 1990; Naidoo & Naidoo, 1992; Pezeshki, 1994),but also to take competitive advantage over flood-sensitiveneighbours (Crawford et al., 1989; Weiher & Keddy, 1995).The present study shows that P. dilatatum has well-developedmechanisms to avoid stress by severe flooding. Individualplant responses to flooding in this species involved changesin leaf anatomy and shoot architecture, and the abilityto maintain basic physiological processes over a range ofatmospheric conditions. This high degree of functional plasticityallows P. dilatatum to grow during flooding periods, even whensurrounded by natural competitors.
The development of aerenchyma is considered the mostimportant trait for a plant to tolerate soil anaerobiosis duringextended floods (Jackson & Drew, 1984; Blom & Voesenek,1996). Flooded P. dilatatum plants contain substantial amountsof aerenchyma in the root cortex (Rubio et al., 1995) and inleaf tissues (Fig. 1), forming an internal system of inter-connected channels. The increased porosity found in theroots of flooded plants suggests that root tissues were rapidlysupplied with extra-air upon submergence. This is consistentwith the large fractions of constitutive root aerenchyma foundin P. dilatatum clones from various lowland grasslands (Rubioet al., 1995; Loreti & Oesterheld, 1996). Internal oxygensupply to the roots becomes critically important as oxygenconcentrations decrease in the flooded soil (Laan et al., 1990).Aerenchymatous tissues provide the plant with a low-resistance, air diffusion pathway that may enhance the oxygensupply to the roots. The existence of a well-developed systemof air-filled intercellular spaces may also decrease oxygenrequirements by reducing the amount of respiring tissues(Armstrong, 1979; Hsiao & Huang, 1989).
Flooding changed the shoot architecture of P. dilatatum,increasing the amount of leaf and tiller area deployed abovethe water surface (Figs 4, 5). Similar morphological changeshave been reported for other herbaceous species growing inperiodically flooded habitats (Bleecker et al., 1986; Laan &Blom, 1990; Van der Sman et al., 1991; Naidoo & Naidoo,1992; Grimoldi et al., 1999). Several studies suggest thatshoot elongation in submerged plants may be controlled byphytohormones, and in particular by ethylene production, asin flood-tolerant Rumex species (Van der Sman et al., 1991;Banga et al., 1996). Experiments conducted on isolated P.dilatatum plants showed that flooding may increase shootheight without substantial changes in total plant biomass(Rubio et al., 1995). Our study revealed various anatomicaland architectural changes that may account for the remark-able shift in plant morphology induced by flooding in thisgrass species. The leaf sheath of flooded plants had greaternumbers of longer parenchymatous cells, which may explainthe enhanced leaf elongation observed under flooding (Figs 3,
4). Bleecker et al. (1986) showed that shoot submergencein deepwater rice induces rapid internode elongation, aresponse involving an increase in both the number and lengthof internodal cells. We also found that the tillers of floodedplants shifted their insertion angles adopting a more uprightposition (Fig. 5). Such a plastic response has usually beenassociated with changes in red : far-red ratios of incominglight in the presence of neighbours (Gibson et al., 1992), buthas rarely been reported as an adaptation to flooding. Thedevelopment of longer leaves and stems, coupled with thevertical orientation of tillers, restored contact with the atmos-phere in flooded plants, and thus contributed to maintain adiffusion pathway for oxygen transport from air-exposedorgans to submerged roots. The use of atmospheric oxygen tomaintain aerobic respiration in root tissues is essential for aplant’s strategy to survive extended flooding (Laan et al.,1990).
We proposed at the outset that flooding tolerance in P. dila-tatum would be associated with its ability to avoid the stressimposed by soil-oxygen deficiency and to regulate its waterstatus during flooding periods. In flood-intolerant species,persistent root anoxia causes severe limitation in the energyavailable to maintain normal water and mineral uptake rates(Jackson & Drew, 1984). The impaired function of floodedroot systems has inevitable consequences for gas exchangeprocesses at the leaf level (Kozlowski & Pallardy, 1984).We observed that P. dilatatum plants were able to regulatestomatal conductance and leaf water potential throughoutan extended flooding period under variable atmospheric con-ditions (Fig. 6). Flooding had no measurable impact on plantwater status under benign atmospheric conditions (lowVPDair). However, when plants were exposed to the morestressful conditions determined by high VPDair levels, flood-ing exerted a significant positive influence on water andcarbon relations by preventing drastic declines in stomatalconductance, leaf water potential, or photosynthesis (Figs 6,7). The decrease in ψl of unflooded plants at high VPDair con-ditions likely reflected an imbalance between transpirationlosses and water absorption by the roots. In a flooded soil,the root contact surface for water uptake is enlarged as soilmacropores become filled with water (Jackson & Drew, 1984).This might prevent limitation of water absorption due toabnormal root-soil contact in air-filled macropores (Stirzaker& Passioura, 1996). Higher water absorption by flooded rootscould balance increased transpiration losses in periods ofsevere atmospheric water deficit. Therefore, the water balanceof plants growing in waterlogged soil may be less sensitive tochanging atmospheric conditions, provided a mechanismexists to supply internally the extra-oxygen needed to main-tain an active (submerged) root system.
The results of the glasshouse experiment show thatincreased leaf extension rates in flooded plants correspondedwith the higher leaf water potential and photosynthesis ratesmaintained under stressful atmospheric conditions. It is
NPH_257.fm Page 297 Thursday, September 27, 2001 5:19 PM

www.newphytologist.com © New Phytologist (2001) 152: 291–299
Research298
possible, however, that hormonal effects were involved inthe expression of this growth response (Voesenek et al., 1992).Because photosynthesis was always measured under highPPFD levels, differences in Pn between flooded and controlplants may be accounted for by differences in stomatalconductance (Fig. 7). Increased leaf temperatures at higherVPDair would contribute to enhance Pn rates in this C4 grass,as long as the process is not limited by stomata closure. Lowstomatal conductance constrained CO2 uptake in water-stressed, control plants, especially when these were exposed tohigh VPDair conditions. Such a stomatal response couldreflect either a hydraulic effect (threshold ψl exceeded) or adirect influence from the leaf-air gradient in water vapourpressure associated with high leaf temperatures (Lange et al.,1971; Else et al., 1995).
Our results add to previous work suggesting that the plasticresponses that allow P. dilatatum to grow above ground duringprolonged floods may not entail a cost in terms of resourceacquisition below ground (Rubio et al., 1995; Rubio &Lavado, 1999). Furthermore, we demonstrate here that, whengrowing in flooded soil, this grass species has a remarkablecapacity for regulating leaf water and carbon relations underhighly changing atmospheric conditions. It must be noted,however, that in order to provide a more realistic assessmentof adaptive responses to flooding, and unlike previous studiesusing potted plants, we have focused on the behaviour ofP. dilatatum plants that were surrounded by assemblages ofnatural competitors. Visual observations during the experi-ment suggested that total above-ground biomass, and indeedinterspecific competition on target plants, could be greaterin flooded microcosms (Insausti et al., 1999). Thus, whilesome of the observed changes in the shoot architecture ofP. dilatatum may have partly reflected the influence of neigh-bouring vegetation (Gibson et al., 1992; Rubio et al., 1995),it seems unreasonable to attribute the enhanced physiologicaladjustment of flooded plants to a modification in the com-petitive environment.
To summarize, the evidence reported here indicates that asuite of responses occurring at anatomical, morphological andphysiological levels enables P. dilatatum to withstand severeflooding in the presence of neighbours. This has implicationsfor the population dynamics of this grass species in periodic-ally flooded environments. The ability to survive and growduring waterlogging periods provides a competitive advantageover less tolerant species (Crawford et al., 1989; Weiher &Keddy, 1995) and may thus account for the biomass increaseof P. dilatatum observed in flooded plant communities(Insausti et al., 1999). Nevertheless, within a communitycontext, positive flooding effects on vegetative growth andnutrient status of flood-tolerant species (Rubio & Lavado,1999) may also involve the use of soil resources released bymortality of nontolerant neighbours. Such community feed-backs on the abundance of flood-resistant species deservefurther experimental investigation.
Acknowledgements
We thank Edy Ploschuk and Gustavo Striker for their assist-ance with gas exchange measurements and Gabriela Zarlavskyfor technical assistance with the anatomical analysis. We areespecially grateful to the Bordeu family for facilitating ourwork at Estancia ‘Las Chilcas’. The comments from threeanonymous reviewers improved the consistency of the manu-script. This study was funded by grants from the Universityof Buenos Aires and Consejo Nacional de InvestigacionesCientíficas y Técnicas (CONICET) of Argentina.
References
Armstrong W. 1979. Aereation in higher plants. In: Woolhouse HWW, ed. Advances in botanical research, Vol. 7. London, UK: Academic Press Inc, 225–332.
Banga M, Blom CWPM, Voesenek LACJ. 1996. Sensitivity to ethylene: The key factor in submergence-induced shoot elongation of Rumex. Plant, Cell & Environment 19: 1423–1430.
Bazzaz FA. 1996. Plants in changing environments. Cambridge, UK: Cambridge University Press.
Bleecker AB, Schuette JL, Kende H. 1986. Anatomical analysis of growth and development patterns in the internode of deepwater rice. Planta 169: 490–497.
Blom CWPM, Bogemann GM, Laan P, Van der Sman AJM. 1990. Adaptations to flooding in plants from river areas. Aquatic Botany 38: 29–47.
Blom CWPM, Voesenek LACJ. 1996. Flooding: the survival strategies of plants. Trends in Ecology and Evolution 11: 290–295.
Burkart SE, León RJC, Movia CP. 1990. Inventario fitosociológico del pastizal de la Depresión del Salado (Prov. Bs. As.) en un área representativa de sus principales ambientes. Darwiniana 30: 27–69.
Crawford RMM, Studer C, Studer K. 1989. Deprivation indifference as a survival strategy in competition: advantages and disavantages of anoxia tolerance in wetland vegetation. Flora 182: 189–201.
Else MA, Davies WJ, Malone M, Jackson MB. 1995. A negative hydraulic message from oxygen-deficient roots of tomato plants. Influence of soil flooding on leaf water potential, leaf expansion, and synchrony between stomatal conductance and root hydraulic conductivity. Plant Physiology 109: 1017–1024.
Gibson D, Casal JJ, Deregibus VA. 1992. The effects of plant density on shoot and leaf lamina angles in Lolium multiflorum and Paspalum dilatatum. Annals of Botany 70: 69–73.
Gough L, Grace JB. 1998. Effects of flooding, salinity and herbivory on coastal plant communities, Louisiana, United States. Oecologia 117: 527–535.
Grimoldi AA, Insausti P, Roitman GG, Soriano A. 1999. Responses to flooding intensity in Leontodon taraxacoides. New Phytologist 141: 110–119.
Hsiao AI, Huang WZ. 1989. Effects of flooding on rooting and sprouting of isolated stem segments and on plant growth of Paspalum distichum L. Weed Research 29: 335–344.
Insausti P, Chaneton EJ, Soriano A. 1999. Flooding reverted grazing effects on plant community structure in mesocosms of lowland grassland. Oikos 84: 266–276.
Jackson MB, Drew M. 1984. Effects of flooding on growth and metabolism of herbaceous plants. In: Kozlowski TT, ed. Flooding and plant growth. London, UK: Academic Press Inc, 47–128.
Justin SHFW, Armstrong W. 1987. The anatomical characteristics of roots and plant responses to soil flooding. New Phytologist 106: 465–495.
NPH_257.fm Page 298 Thursday, September 27, 2001 5:19 PM

© New Phytologist (2001) 152: 291–299 www.newphytologist.com
Research 299
Kozlowski TT, Pallardy SG. 1984. Effect of flooding on water, carbohydrate and mineral relations. In: Kozlowski TT, ed. Flooding and plant growth. London, UK: Academic Press Inc, 165–193.
Laan P, Tosserams M, Blom CWPM, Veen BW. 1990. Internal oxygen transport in Rumex species and its significance for respiration under hipoxic conditions. Plant and Soil 122: 39–46.
Laan P, Blom CWPM. 1990. Growth and survival responses of Rumex species to flooded and submerged conditions: The importance of shoot elongation, underwater photosynthesis and reserve carbohydrates. Journal of Experimental Botany. 41: 775–783.
Lange OL, Lasch R, Schulze ED, Kappen L. 1971. Responses of stomata to changes in humidity. Planta 100: 76–86.
Loreti J, Oesterheld M. 1996. Intraspecific variation in the resistance to flooding and drought in populations of Paspalum dilatatum from different topographic positions. Oecologia 108: 279–284.
Naidoo G, Naidoo S. 1992. Waterlogging responses of Sporobolus virginicus (L.). Kunth. Oecologia 90: 445–450.
Osmond CB, Austin MP, Berry JA, Billings WD, Boyer JS, Dacey JWH, Nobel PS, Smith SD, Winner WE. 1987. Stress physiology and the distribution of plants. Bioscience 37: 38–48.
Pennings SC, Callaway RM. 1992. Salt marsh plant zonation: the relative importance of competition and physical factors. Ecology 73: 661–690.
Pezeshki SR. 1994. Plant response to flooding. In: Wilkinson RE, ed. Plant–environment interactions. New York, USA: Marcel Dekker Inc, 289–321.
Potvin C, Lechowicz MJ. 1990. The statistical analysis of ecophysiological response curves obtained from experiments involving repeated measures. Ecology 71: 1389–1400.
Rubio G, Casasola G, Lavado RS. 1995. Adaptations and biomass production of two grasses in response to waterlogging and soil nutrient enrichment. Oecologia 102: 102–105.
Rubio G, Lavado RS. 1999. Acquisition and allocation of resources in two waterlogging-tolerant grasses. New Phytologist 143: 539–546.
Sala OE, Deregibus VA, Schlichter T, Alippe H. 1981. Productivity dynamics of a native temperate grassland in Argentina. Journal of Range Management 34: 48–51.
Sojka RE. 1988. Measurement of root porosity (volume of root air space). Environmental and Experimental Botany 28: 275–280.
Steel RGD, Torrie JH. 1988. Principles and procedures of statistics. A biometrical approach. New York, USA: McGraw-Hill.
Stirzaker RJ, Passioura JB. 1996. The water relations of the root–soil interface. Plant, Cell & Environment 19: 201–208.
Trebino HJ, Chaneton EJ, León RJC. 1996. Flooding, topography and successional age as determinants of species diversity in old field vegetation. Canadian Journal of Botany 74: 582–588.
Tyerman SD, Bohnert HJ, Maurel C, Steudle E, Smit JAC. 1999. Plant aquaporins: their molecular biology, biophysics and significance for plant water relations. Journal of Experimental Botany 50: 1055–1071.
Van der Sman AJM, Voesenek LACJ, Blom CWPM, Harren FJM, Reuss J. 1991. The role of ethylene in shoot elongation with respect to survival and seed output of flooded Rumex maritimus L. Functional Ecology 5: 304–313.
Voesenek LACJ, Van der Sman AJM, Harren FJM, Blom CWPM. 1992. An amalgamation between hormone physiology and plant ecology: a review on flooding resistance and ethylene. Journal of Plant Growth Regulation 11: 171–188.
Von Ende CN. 1993. Repeated-measures analysis: growth and other time-dependent measures. In: Scheiner SM, Gurevitch J, eds. Design and analysis of ecological experiments. New York, USA: Chapman & Hall, 113–137.
Weiher E, Keddy PA. 1995. The assembly of experimental wetland plant communities. Oikos 73: 323–335.
NPH_257.fm Page 299 Thursday, September 27, 2001 5:19 PM





























