Embed Size (px)
Citation preview

International Journal of Primatology, Vol. 18, No. 6, 1997
Food Competition Between Wild Orangutansin Large Fig Trees
Sri Suci Utami,1'2 Serge A. Wich,1 Elisabeth H. M. Sterck,1-3 andJan A. R. A. M. van Hooff1
Received March 18, 1996; accepted June 3, 1997
Orangutans are usually solitary. However, occasionally aggregations are formed,especially in large fruiting fig trees. Individuals in these aggregations may experiencescramble or contest competition for food. We investigated the type and strengthof food competition in large figs among wild Sumatran orangutans. Adult malesforaged more efficiently than adult females and subadult males did. Theavailability of ripe fruit is positively related to the number of orangutans visitinga fig tree and their foraging efficiency. The number of orangutans in a fig treedid not affect patch residence time and foraging behavior, though orangutans spentmore time feeding when aggregation size increased in a fig tree. Dominancerelationships could be measured in a number of dyads. Differences in dominancedid not affect foraging behavior. The patch residence time of subordinateindividuals was reduced on days that a dominant individual also visited the fig.In conclusion, orangutans seem to adjust aggregation size to the number ofavailable ripe fruits in a fig tree in such a way that scramble competition wasabsent. Contest competition determined access to large fig trees.
KEY WORDS: orangutan; aggregations; food competition; dominance relationships.
INTRODUCTION
Orangutans (Pongo pygmaeus) live a semisolitary life, in contrast tomany other diurnal primate species. Aggregations occur only occasionally
'Ethology & Socioecology, Utrecht University, P.O. Box 80086, 3508 TB Utrecht, TheNetherlands.
2Facultas Biologi, Universitas Nasional, Jl. Sawo Manila—Pasar Minggu, Jakarta, Indonesia.3To whom correspondence should be addressed.
909
0164-0291/97/1200-0909$12.50/0 © 1997 Plenum Publishing Corporation

910 Utami, Wich, Sterck, and van Hooff
and usually in large food patches or in a sexual context (MacKinnon, 1974;Rijksen, 1978; Galdikas, 1978; Sugardjito et al, 1987; Mitani et al., 1991).Other large-bodied frugivorous primates, i.e., chimpanzees (Pan spp.) andspider monkeys (Ateles spp.), also forage solitarily, or in subgroups of vari-able size and composition, and live in a so-called fission-fusion society(Nishida and Hiraiwa-Hasegawa, 1987; Robinson and Janson, 1987). Thefactors that regulate aggregation formation may be similar for orangutansand fission-fusion species.
The absence of permanent groups in fission-fusion species has beenrelated to the combination of a relatively low predation risk (van Schaik,1983,1989; Dunbar, 1988) and potentially strong food competition betweenfemales (van Schaik, 1989). In orangutans, a similar combination of featuresis present. Their large body size and arboreal lifestyle result in reducedpredation risk (MacKinnon, 1974; Rijksen, 1978; Sugardjito, 1986). Aggre-gations are formed in large fig trees with large fruit crops or when generalfood availability is high (Sugardjito et al, 1987).
Orangutans form three types of aggregations: travel bands, in whichindividuals feed together within a patch, leave together, stay together forsome time, and sometimes visit the next fruit patch together; temporaryaggregations, wherein individuals feed together within a fruit patch andleave separately (Sugardjito et al, 1987); and consorts, in which a receptivefemale and an adult or subadult male travel together for a period (Schur-mann and van Hooff, 1986). Travel bands are comparable with the passivetravel associations and other social associations of Mitani et al. (1991).Temporary aggregations are comparable to their passive food associationand aggressive and avoidance social association. Their mating social asso-ciation is comparable to consorts.
The two food related aggregation types are related to different foodavailabilities. Travel bands occur more frequently when there is an abun-dance of fruiting trees, and food competition is normally absent. Travelbands may be formed only as a side effect of foraging rules (te Boekhorstand Hogeweg 1994). Temporary aggregations occur more frequently whengeneral food availability is low and are usually formed inside large fig trees.This observation led to the prediction that orangutans may be forced toform temporary aggregations in fig trees, in spite of food competition (Sug-ardjito et al, 1987). Indeed, aggression was more common in temporaryaggregations than in travel bands (Sugardjito, 1986).
The formation of food-related aggregations by orangutans resem-bles the formation of aggregations in fission-fusion species. As has beensuggested for orangutans (Sugardjito et al, 1987), the sizes of subgroupsfor common chimpanzees (Pan troglodytes) and black-handed spider mon-keys (Ateles geoffroyi) are related to the size, density, and distribution of

Food Competition in Orangutans 911
food resources (Chapman et al, 1995). In these species, there are positivecorrelations between patch quality and subgroup size [chimpanzees:(Ghiglieri, 1984; Isabirye-Basuta, 1988; White and Wrangham, 1988);black spider monkeys, Ateles paniscus: (Symington, 1988); muriquis,Brachyteles arachnoides: (Strier 1989)]. However, positive relationshipsbetween patch quality and feeding aggregation size are far from uniquefor fission-fusion species and also occur in group living primates, e.g.,mantled howler monkeys [Alouatta palliata (Leighton and Leighton,1982)]. For orangutans, the relationship among measures of patch quality,aggregation size, and the ensuing effect on food competition have notbeen quantified.
In addition, foraging behavior may differ between age-sex classes ow-ing to the marked sexual dimorphism in orangutans. Adult males (50-90kg) are about twice as heavy as adult females (30-50 kg) and subadultmales (30-50 kg) (Rijksen, 1978; Rodman, 1984; Rodman and Mitani,1987). These differences in body size may be reflected in foraging behavior.In orangutans, there is a positive relation between body size and feedingbout length (Wheatly, 1982).
We measured food competition in large fig trees with large fruit cropsin which organgutans formed temporary aggregations. Two types of foodcompetition can occur within groups: scramble and contest (van Schaik,1989). Scramble competition will result in smaller benefits for individualsin larger groups. It can manifest itself in two ways. First, the foraging ef-ficiency of all individuals can decrease when more individuals use a patchsimultaneously. Second, the patch residence time may be smaller when thefeeding aggregration size in the patch is larger, e.g., among chimpanzees(Isabirye-Basuta, 1988).
Contest competition leads to larger benefits for dominant individuals.It can manifest itself in three ways. First, some individuals may systemati-cally replace other individuals, leading to unidirectional dominancerelationships. Second, subordinate individuals may have a relatively low for-aging efficiency when dominant individuals are present. Third, subordinateindividuals were expected to have a shorter patch residence time whendominant individuals also visited the fig tree on the same day.
METHODS
Study Area
We conducted the study at the Ketambe Research Station (3°41'N,97°39'E), Gunung Leuser National Park, Northern Sumatra, Indonesia. The
912 Utami, Wich, Sterck, and van Hooff
area consists mainly of undisturbed primary lowland rain forest (Rijksen,1978; van Schaik and Mirmanto, 1985). The number of fruiting fig treespeaks around April and around October (Sugardjito et al, 1987).
Subjects
The orangutans of Ketambe have been studied since 1971 (Rijksen,1978; Schurmann, 1982; Schurmann and van Hooff, 1986; Sugardjito, 1986;Mitrasetia, 1995). Some individuals have been recognized since 1971. Oth-ers were born during the studies, or are immigrants. Most individuals werehabituated.
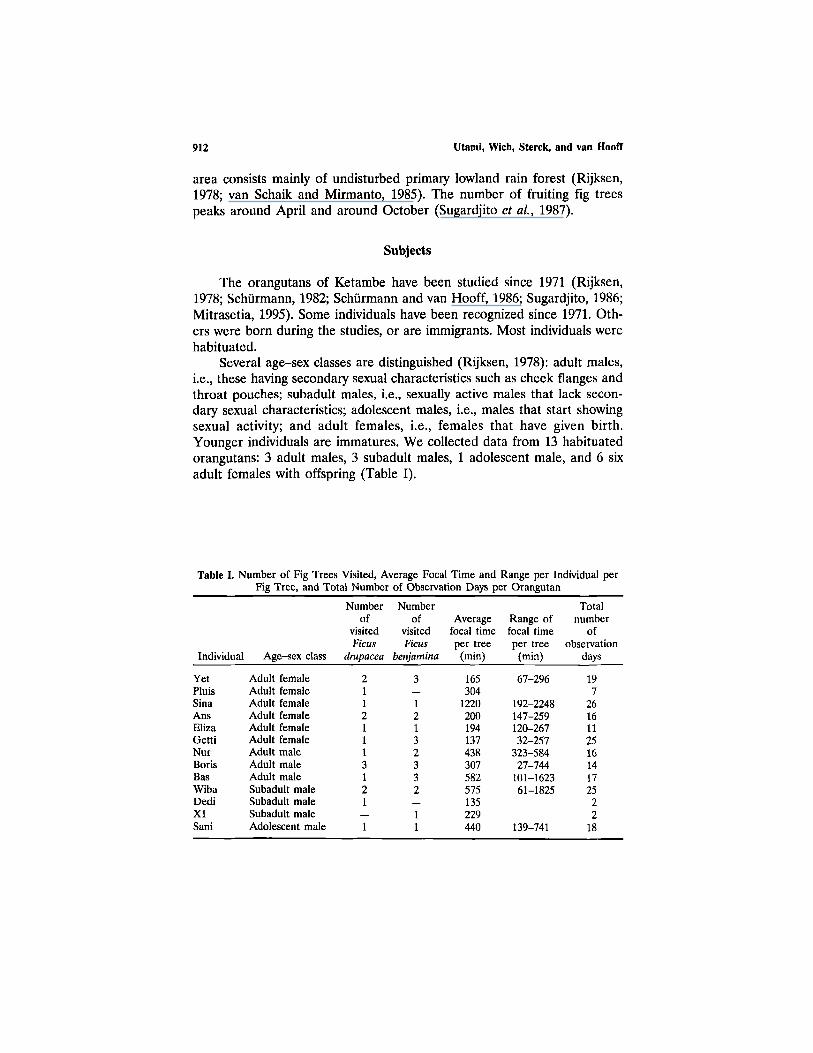
Several age-sex classes are distinguished (Rijksen, 1978): adult males,i.e., these having secondary sexual characteristics such as cheek flanges andthroat pouches; subadult males, i.e., sexually active males that lack secon-dary sexual characteristics; adolescent males, i.e., males that start showingsexual activity; and adult females, i.e., females that have given birth.Younger individuals are immatures. We collected data from 13 habituatedorangutans: 3 adult males, 3 subadult males, 1 adolescent male, and 6 sixadult females with offspring (Table I).
Table I. Number of Fig Trees Visited, Average Focal Time and Range per Individual perFig Tree, and Total Number of Observation Days per Orangutan
Individual
YetPluisSinaAnsElizaGettiNurBorisBasWibaDediX1Sani
Age-sex class
Adult femaleAdult femaleAdult femaleAdult femaleAdult femaleAdult femaleAdult maleAdult maleAdult maleSubadult maleSubadult maleSubadult maleAdolescent male
Numberof
visitedFicus
drupacea
21121113121
1
Numberof
visitedFicus
benjamina
3
12132332
11
Averagefocal timeper tree
(min)
165304
1220200194137438307582575135229440
Range offocal timeper tree
(min)
67-296
192-2248147-259120-26732-257
323-58427-744
101-162361-1825
139-741
Totalnumber
ofobservation
days
197
2616112516141725
22
18

Food Competition in Orangutans 913
Data Collection
We recorded the behavior of orangutans inside large strangling figtrees with large fruit crops. We monitored 60 fig trees on a monthly basisfor fruit availability and ripeness and additional fig trees on a less regularbasis. In this way, we located fig trees that would start to fruit.
We estimated the size of a fig tree by the height of the crown andthe crown diameter, all in 5-m classes. We also estimated the total fruitavailability and availability of ripe fruits. We counted the fruits on onebranch and multiplied that number by the number of branches in the tree.This resulted in eight fruit availability classes: 0, 1-10, 10-100, 100-500,500-1000, 1000-5000, 5000-10,000, and > 10,000 items. We used the mid-points of these classes for our calculations. The midpoint of the class of> 10,000 items is 15,000.
When fruit started to ripen in a large fig tree, the observer (S.W.)spent 4-23 days from dawn to dusk underneath it. We obtained data withtwo fig species: Ficus drupacea and F. benjamina. We monitored one treeat a time.
We collected data from December 1993 until June 1994. We spent atotal of 70 days observing orangutans inside the two fig species: 51 daysfor F. drupacea and 19 days for F. benjamina.
We recorded the visits of orangutans to the fig trees. An individualcould visit a fig tree several times on one day. We recorded the times atwhich an orangutan entered or left the fig tree. Patch residence time (PRT)is the time spent in the monitored fig tree on a given day by one individual.Total patch residence time (tPRT) is the total number of minutes spentby all orangutans that visited a fig tree on a given day.
We recorded the behavior of an orangutan—focal—via the focal-ani-mal sampling technique (Martin and Bateson, 1986) after it entered a figtree. A cassette recorder facilitated continuous recording of behavior.
When one individual was in a fig tree, we recorded its behavior aslong as it was visible. When several individuals were simultaneously presentinside a fig tree, we alternated observations between individuals and focalsevery 10 min.
We analyzed orangutan behavior in four categories: feeding, social,travel, and resting. A feeding individual was eating or moving between fooditems or both. Feeding started when an individual picked a fig. Feedingended when it stopped chewing, or when its face could not be seen andno fruit was taken by an orangutan for approximately 1 min. Social behaviorincludes playing, chasing and physical contact with another individual.Travel is all movements while an individual was not feeding or involved in

914 Utami, Wich, Sterck, and van Hooff
social behavior. Resting occurred when it did not feed or travel and wasnot involved in social behavior.
We recorded feeding behavior in more detail whenever the orangutanwas sufficiently visible. We noted the consumption of food items picked bymouth or hand whenever possible. During feeding movements of <5 sec,we recorded changes of posture and turns of the body >45° as movements.
In addition, whenever possible, we conducted scans every 10 min torecord the activity—feeding, social, travel, or resting—of every individualin the fig tree. In practice, social and travel were virtually absent, and there-fore the scan data consisted only of two behaviors, which we used tomeasure the activity budget.
For one female (Sina) we excluded one focal, for which the intakerate was three times higher than in comparable focals. Accordingly, we ex-cluded this datum as an outlier.
Displacements
Orangutans usually remain at a large distance from conspecifics.Therefore, we defined displacement as the fast departure out of a foodtree by an individual when another individual approached within <50 m ofthe tree. The dyadic dominance of individuals is based on displacementdata from the years 1993 and 1994. We determined the dominance hier-archies of males and females and treated them separately.
Definitions
Foraging behavior encompasses intake rate, movements, and foragingefficiency. The intake rate (g/min) is calculated as the ingested wet weightdivided by the time spend feeding. We calculated the ingested wet weightfrom the picking rate and the wet weight of F. drupacea [21.75 g per item(van Schaik and van Noordwijk, 1986)] and of F. benjamina [6 g per item(unpublished data)]. The foraging efficiency (g/movement) is calculated bydividing the intake rate by the number of movements.
The maximum aggregation size is as the largest number of adult,subadult and adolescent individuals that were simultaneously present insidethe fig tree on a given day. The aggregation size is the number of neighbors:the number of adult, subadult, and adolescent individuals inside the figtree minus 1, namely, the focal individual. We also calculated the totalnumber of individuals that visited a fig tree per day.

Food Competition in Orangutans 915
Data Preparation
We analyzed only focals in which an orangutan was feeding and dis-carded focals lasting <1 min. We also discarded movements that lasted>5 sec since their effect on behavior could not be determined becauseusually an individual would move out of sight, which renders comparisonof focals impossible.
In 1 of 789 focals we observed no movements. We could not use it tocalculate foraging efficiency because this would entail a division by zero.
Statistics
The data contain repeated measures on the same individuals. Thisleads to the pooling fallacy (Machlis et al, 1985). Therefore, we lumpedthe data per individual as far as possible in all further analyses.
We used the lumped data points as input for General Linear Models(GLM) in the SAS package. We treated the different behaviors as depend-ent variables and entered them separately into GLM. We treated total fruitavailability, ripe fruit availability, fig species, and age-sex class as inde-pendent variables.
We calculated the differences in behavior between the age-sex classesand between the two fig species with the least square means procedure ofGLM (SAS, 1988; Searl et al, 1980). Wherever feasible we performed non-parametric tests with SPSS. All tests are two-tailed, unless otherwisementioned, and considered significant at p < 0.05.
We tested the linearity of a dominance hierarchy based on displace-ment behavior via a randomization test which takes the number of unknowndominance relationships into account (de Vries, 1995); this is implementedin MATMAN (de Vries et al., 1993).
RESULTS
Orangutan Foraging Behavior
We recorded foraging behavior of orangutans in seven fig trees, fourtrees of Ficus drupacea and three of Ficus benjamina . No orangutan visitedall fig trees (Table I). Both species are large strangling fig trees that reachto >40 m and have crown-diameters of >25 m. The crown volume of F.drupacea varies between 12,500 and 48,000 m3 (n = 4) and that of F. ben-jamina between 12,500 and 15,600 m3 (n = 3).

916 Utami, Wich, Sterck, and van Hooff
Orangutan foraging behavior is affected by their age-sex class and thefig species. Adult males have a higher intake rate and foraging efficiencythan adult females and subadult males do. Adult males also move less thanthe other age-sex classes do. The behavior of subadult males and adultfemales do not differ significantly (Fig. la). The orangutans behaved dif-ferently in the two fig species. They moved less and had a higher foragingefficiency in F. drupacea than in F. benjamina (Fig. 1b). The intake ratetended to be higher in the latter species (GLM: F = 3.61, p = 0.059).
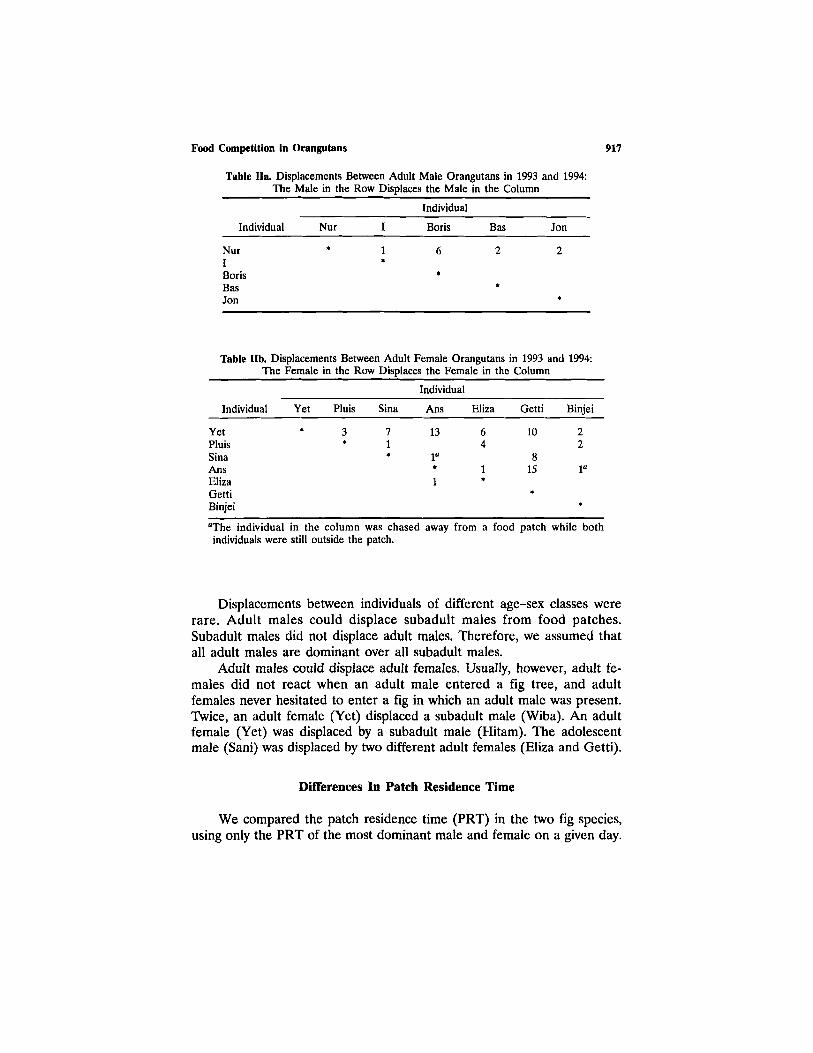
Dominance Relationships
We observed displacements only in large fig trees and other large foodpatches. Few displacements were between adult males. Nur displaced allother adult males and, therefore, was the most dominant adult male in thearea (Table IIa). We did not see displacements among the other adultmales (Table IIa); their dominance relationships are unclear.
Displacements between adult females occurred. They were unidirec-tional, with one exception (Table IIb). The dominance hierarchy tended tobe linear [adjusted Landau's linearity index h' (corrected for the numberof unknown dominance relationships) = 0.68; p < 0.10 (de Vries, 1995)].
Fig. 1. Foraging behavior of orangutans (a) of different age-sex classes and (b) indifferent fig species. The data set is reduced to one data point for each individual ineach fig species. With a Bonferroni correction the significance level is 0.016. The for-aging behavior encompasses movements multiplied by 100 (100*number/min), intake(g/min), and foraging efficiency (g/movement). "0.05 > p > 0.01; "0.01 > p > 0.001;
p < 0.001.
Food Competition in Orangutans 917
Table Ha. Displacements Between Adult Male Orangutans in 1993 and 1994:The Male in the Row Displaces the Male in the Column
Individual
NurIBorisBasJon
Nur
*
I
1*
Individual
Boris
6
*
Bas
2
*
Jon
2
*
Table IIb. Displacements Between Adult Female Orangutans in 1993 and 1994:The Female in the Row Displaces the Female in the Column
Individual
YetPluisSinaAnsElizaGettiBinjei
Yet
*
Pluis
3*
Sina
71*
Individual
Ans
13
1°*
1
Eliza
64
1*
Getti
10
815
*
Binjei
22
1"
*
"The individual in the column was chased away from a food patch while bothindividuals were still outside the patch.
Displacements between individuals of different age-sex classes wererare. Adult males could displace subadult males from food patches.Subadult males did not displace adult males. Therefore, we assumed thatall adult males are dominant over all subadult males.
Adult males could displace adult females. Usually, however, adult fe-males did not react when an adult male entered a fig tree, and adultfemales never hesitated to enter a fig in which an adult male was present.Twice, an adult female (Yet) displaced a subadult male (Wiba). An adultfemale (Yet) was displaced by a subadult male (Hitam). The adolescentmale (Sani) was displaced by two different adult females (Eliza and Getti).
Differences In Patch Residence Time
We compared the patch residence time (PRT) in the two fig species,using only the PRT of the most dominant male and female on a given day.

918 Utami, Wich, Sterck, and van Hooff
Adult males resided on average 176 min in Ficus drupacea (n = 44) and195 min in F. benjamina (n = 18); adult females resided on average 176min in F. drupacea (n = 52) and 149 min in F. benjamina (n = 19); andsubadult males, 206 min in F. drupacea (n = 7) and 128 min in F. benjamina(n = 3). The PRT in F. drupacea and F. benjamina do not differ signifi-cantly for adult males (Mann-Whitney U test: U = 381, p = 0.82), or foradult females (U = 427.5, p = 0.45), or for subadult males (U = 7, p =0.43).
We compared PRT of the different age-sex classes. Because PRT be-tween the fig species did not differ for each age-sex class, we used datapoints per age-sex class together in one data set. PRT of adult males, adultfemales, and subadult males do not differ significantly [Kruskall-Wallis test:X2(ties) = 1.03, p = 0.60].
Effects of Patch Quality
A relationship between the quality of a food tree and its use by orangu-tans was expected. We measured the quality of the food patch as the totalfruit availability (i.e., both ripe and unripe fruit) and as the availability ofripe fruit. These two measures are correlated (Pearsons corr. coeff. = -0.21,p < 0.045, one-tailed).
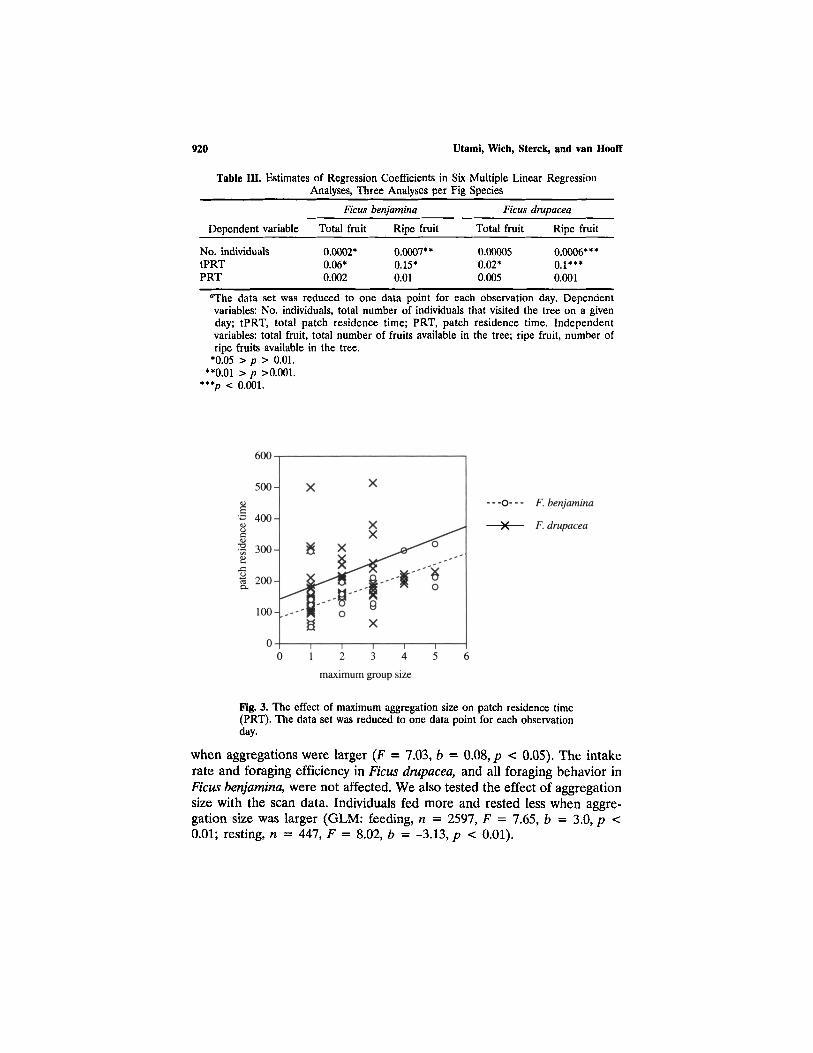
Foraging efficiency is affected by the number of ripe fruits (F = 8.28,b = 0.008, p < .005, one-tailed: the data set was reduced to one data pointfor each individual for each observation day), whereas the intake rate andthe number of movements were not affected. The total number of fruitsdid not affect foraging behavior. Maximum aggregation size is not corre-lated with total number of fruits in a fig tree (Fig, 2a) but is with thenumber of ripe fruits (Fig. 2b). The PRT is not correlated with the numberof ripe fruits (Table III). The number of ripe fruits is a better predictorof the total number of individuals and tPRT than the total number of fruitsis (Table III).
Effects of Aggregation Size
Increasing aggregation size might affect foraging behavior negatively.Yet PRT is positively related to aggregation size in Ficus benjamina (GLMafter correcting for the number of ripe fruits: F = 2.9, b = 30.3, p < 0.05)and Ficus drupacea (GLM after correcting for the number of ripe fruits:F = 2.7, b = 39.5, p = 0.07) (Fig. 3).
We compared foraging behavior in situations with different aggrega-tion sizes in the two fig species. Orangutans moved more in Ficus drupacea

Food Competition in Orangutans 919
920 Utami, Wich, Sterck, and van Hooff
Table III. Estimates of Regression Coefficients in Six Multiple Linear RegressionAnalyses, Three Analyses per Fig Species
Dependent variable
No. individualstPRTPRT
Ficus benjamina
Total fruit
0.0002*0.06*0.002
Ripe fruit
0.0007**0.15*0.01
Ficus drupacea
Total fruit
0.000050.02*0.005
Ripe fruit
0.0006***0.1***0.001
"The data set was reduced to one data point for each observation day. Dependentvariables: No. individuals, total number of individuals that visited the tree on a givenday; tPRT, total patch residence time; PRT, patch residence time. Independentvariables: total fruit, total number of fruits available in the tree; ripe fruit, number ofripe fruits available in the tree.
•0.05 > p > 0.01.**0.01 > p >0.001.
***p < 0.001.
Fig. 3. The effect of maximum aggregation size on patch residence time(PRT). The data set was reduced to one data point for each observationday.
when aggregations were larger (F = 7.03, b = 0.08, p < 0.05). The intakerate and foraging efficiency in Ficus drupacea, and all foraging behavior inFicus benjamina, were not affected. We also tested the effect of aggregationsize with the scan data. Individuals fed more and rested less when aggre-gation size was larger (GLM: feeding, n = 2597, F = 7.65, b = 3.0, p <0.01; resting, n = 447, F = 8.02, b = -3.13, p < 0.01).

Food Competition in Orangutans 921
Effects of Dominance
Displacements, by definition, affected the use of a particular fig tree.We determined the effect of displacements of adult males on other adultmales and subadult males and of displacements among females. The timeinterval between the displacement and the reentering of a fig tree variedfrom half an hour to several days. For every displacement, the interval be-tween the displacement and the return is in hourly intervals. To calculatethe intervals we used only dawn to dusk hours. When an individual didnot return to the fig it was assigned a time interval larger than the longestobserved interval (35 hr).
The effect of a displacement differed for the age-sex classes (Fig. 4).When an adult male displaced another adult male the latter almost neverreturned to the fig tree and did not reenter it. In contrast, total exclusiondid not occur when adult males displaced subadult males or when adult
Fig. 4. The interval between a displacement and subsequent reentering of the same figtree for adult males with adult males or subadult males and for adult females withadult females. The difference in the effect of displacements between adult males anddisplacements between adult females is significant (Kolmogorov-Smirnov two-sampletest: nl = 5, n2 = 15, K-S Z = 1.936, p < 0.001). The difference in the effect ofdisplacements between adult males and displacements between adult males andsubadult males is also significant (Kolmogorov-Smirnov two-sample test: n1 = 5, n2 =14, K-S Z = 1.919,p < 0.001). The difference in the effect of displacements betweenadult males and subadult males and displacements between adult females is not sig-nificant (Kolmogorov-Smirnov two-sample test: n1 = 5, n2 = 15, K-S Z = 0.794, p =0.55).

922 Utami, Wich, Sterck, and van Hooff
females displaced adult females. The displaced individual usually wouldreenter the food patch, sometimes even while the dominant individual wasstill the patch.
The presence of a dominant individual in a food patch might increasethe feeding costs of a subordinate individual. To test this, we calculatedthe behavior of subordinates in the presence or absence of a dominantindividual. The presence or absence of a dominant individual did not sig-nificantly affect the foraging behavior of subordinate individuals (Table IV).
We calculated PRT of subordinate individuals separately for days onwhich a dominant individual visited the patch as well, and for days on whichno dominant individual visited the patch. We excluded days on which asubordinate individual was the only visitor of a patch. Only for Ficus dru-pacea are sufficient data available. PRT of subordinate individuals wasshortest on days that a dominant individual also visited the fig tree (Wil-coxon matched-pairs test: Z = -2.2014, n = 6, p < 0.05).
DISCUSSION
Orangutans are usually solitary and only occasionally from aggrega-tions. Travel bands form when food is relatively abundant, therefore, theimpact of food competition is weak. In large fig trees, however, organgutansaggregate during periods of presumed food scarcity. Individuals in thesefig trees may experience competition for food (Sugardjito et al, 1987).
First, we determined differences between age-sex classes. Large-bodied adult males probably have higher energetic requirements than otherage-sex classes (Rodman, 1979). Accordingly, adult males moved less, hada higher intake rate, and thus foraged more efficiently than small-bodiedadult females and subadult males did. This may be due to the larger bodysize, and therefore longer reach, of adult males in comparison to adult
Table IV. Effect of Presence of a Dominant IndividualInside a Fig Tree on Foraging Behavior of Subordinate
Orangutans (n = 8)
Foraging behavior
MovementsIntake rateForaging efficiency
Effect of presence ofdominant individual
Z = -1.30, p = 0.19"Z = -1.40, p = 0.16"F = 0.00, p = 0.96fc
"Wilcoxon matched-pair test.''GLM.

Food Competition in Orangutans 923
females and subadult males [cf. gorillas, Gorilla gorilla beringei (Wrangham,personal communication, cited by Fossey and Harcourt, 1977)].
Second, orangutans behaved differently in the two fig species. Theymoved less and had a higher foraging efficiency in Ficus drupacea than inF. benjamina. This was most likely due to the different size of the figs: thelarger F. drupacea can be harvested more efficiently than the smaller fruitof F. benjamina. However, there is no difference in PRT. Therefore, it ismore profitable for an orangutan to feed in a fig of a species with largefruit than one with small fruit.
Competition For Food: Scramble Competition
The aggregation sizes of orangutans are related to the amount of ed-ible food, i.e., the number of ripe fruits, in fig trees. When a fig treecontained more ripe fruit, more orangutans visited the fig per day, andmore individuals feed simultaneously. As a consequence, the tPRT alsoincreased. Hence, orangutan aggregation size is adjusted to the availabilityof ripe fruit. The availability of ripe fruit in a fig tree, however, did notinfluence PRT, intake rate, or number of movements. Moreover, the for-aging efficiency was higher when a larger number of ripe fruits wasavailable. Thus, trees with large fruit crops accommodate more individuals,and they feed more efficiently. This suggests (pace Sugardjito et al, 1987)that scramble competition is absent in large fruiting fig trees.
Scramble competition would result in a positive relationship betweenthe aggregation size and the strength of food competition. Even in fission-fusion species, which are thought to adjust aggregation size to avoid foodcompetition, the formation of aggregations may result in food competition.This can lead to a lower PRT when aggregations are larger [chimpanzees(Isabirye-Basuta, 1988)]. However, in orangutans, aggregation size and PRTare not negatively related. In addition, foraging behavior was usually notaffected and individuals even spent more time feeding when aggregationswere larger. Only the number of movements in one fig species increasedwhen the aggregation size was large. Thus, the aggregation size seemed tomatch food abundance in such a way that scramble competition was absentor low.
Competition For Food: Contest Competition
The formation of an aggregation can result in contest competition forclumped resources. Indeed, displacements were observed in and aroundlarge food trees. Based on these dyadic displacements, rank differences oc-

924 Utami, Wich, Sterck, and van Hooff
curred between individuals. In the Ketambe area, one adult male wasclearly dominant over all other adult and subadult males. The females couldbe ordered in a nearly linear dominance hierarchy. Although the test forlinear hierarchy showed only a trend, the displacement data had a highdirectional consistency, and it seems justified to claim that a hierarchy in-deed exists. With the acquisition of more data, this could becomesignificant.
Displacements resulted in the departure from the fig tree of the dis-placed individual. Some displaced individuals returned. The differentage-sex classes differed in the tolerance they showed for individuals of theirown class. Adult males were completely intolerant toward each other, asin other studies (Mitani, 1985; Rodman and Mitani, 1987), and never letanother adult male reenter the patch after a displacement. Because aggre-gations could become rather large, it is unlikely that adult males could nottolerate the food competition other adult males would cause. It is morelikely that sexual competition among adult males led to this intolerance.
Subadult males often reentered the fig tree after a displacement byan adult male, though they usually waited until the adult male had left thefig tree. Adult females also could reenter the fig tree after a displacementby an adult female, sometimes with and sometimes without the dominantfemale still in the fig tree. It seemed that some female-female dyads weremore tolerant than others, though not enough data are available to testthis supposition.
The dominance rank of an individual affected its feeding behavior.Subordinate individuals had a shorter PRT on days that dominant indi-viduals were present. Even so, they did not forage differently in thepresence of a dominant individual. Therefore, contest competition mainlyexpressed itself in the access to fig trees.
In conclusion, like fission-fusion species, orangutans exhibit a positiverelationship between patch quality and the size of temporary aggregations.The nature of food competition in fission-fusion species is unclear, thoughreports suggest that some species experience scramble competition [chim-panzees, negative relationship between aggregation size and foragingefficiency, (Isabirye-Basuta, 1988); black-handed spider monkeys: negativerelationship between aggregation size and feeding time (Chapman, 1988)]or contest competition [black-handed spider monkeys: possible exclusionof low-ranking females from subgroups (Chapman, 1990)]. In these species,social factors also affect aggregation size and composition (Chapman, 1990;Chapman et al, 1995), and in orangutans, aggregation sizes are also af-fected by both food sources and social factors (Mitani et al., 1991).Orangutans in temporary aggregations experience contest, but not scram-ble, competition. Aggregation sizes in large fig trees are adjusted to the

Food Competition in Orangutans 925
availability of food in such a way that scramble competition is absent. Sub-ordinate individuals, however, experienced contest competition. Theiraccess to fig trees was restricted when dominant orangutans were present.Furthermore, male mating competition may limit access of subordinateadult males. Thus, both ecological and social factors affect aggregation sizeand composition in fission-fusion species and orangutans. It is, however,too early to speculate on how these factors affect the type of food com-petition.
ACKNOWLEDGEMENTS
We gratefully acknowledge the cooperation and support of the Indo-nesian Institute of Science (LIPI, Jakarta), the Indonesian NatureConservation Service (PHPA) in Jakarta, Medan and Kutacane (GunungLeuser National Park Office), Universitas National (UNAS, Jakarta), andthe Leuser Development Programme, Medan. Utami was supported byWOTRO, Netherlands Foundation for the Advancement of Tropical Re-search. Wich thanks Romy Steenbeek for helping him in the field and forcomments while setting up this study and the Utrecht University for finan-cially supporting the fieldwork. We thank the Indonesian students Imam,Nahot, and Basrul for help with the data collection. Comments by RubenBlijdorp, Marjolijn Das, Chris Fairgrieve, Zjeffrie Pereboom, Carel vanSchaik, Romy Steenbeek, Han de Vries, and two referees improved earlyversions of the manuscript. Finally, we thank Han de Vries for help withstatistical matters.
REFERENCES
Chapman, C. (1988). Patch use and patch depletion by the spider and howling monkeys ofSanta Rosa National Park, Costa Rica. Behaviour 105: 99-116.
Chapman, C. A. (1990). Association patterns of spider monkeys: The influence of ecologyand sex on social organization. Behav. Ecol. Sociobiol. 26: 409-414.
Chapman, C. A,, Wrangham, R. W., and Chapman, L. J. (1995). Ecological constraints ongroup size: An analysis of spider monkey and chimpanzee subgroups. Behav. Ecol.Sociobiol. 36: 59-70.
de Vries, H. (1995). An improved test of linearity in dominance hierarchies containingunknown or tied relationships. Anim. Behav. 50: 1375-1389.
de Vries, H., Netto, J. W., and Hanegraaf, P. L. H. (1993). MATMAN: A program for theanalysis of sociometric matrices and behavioral transition matrices. Behaviour 125:157-175.
Dunbar, R. I. M. (1988). Primate Social Systems, Crook Helm, Beckenham.Fossey, D., and Harcourt, A. H. (1977). Feeding ecology of free-ranging mountain gorilla
(Gorilla gorilla beringei). In Clutton-Brock, T. H. (ed.), Primate Ecology: Studies of Feeding

926 Utami, Wich, Sterck, and van Hooff
and Ranging Behavior in Lemurs, Monkeys and Apes, Academic Press, London, pp.415-447.
Galdikas, B. M. F. (1978). Orang-utan Adaptation at Tanjung Puting Reserve, Central Borneo,Ph.D. thesis, University of California, Los Angeles.
Ghiglieri, M. P. (1984). Feeding ecology and sociality of chimpanzees in Kibale forest, Uganda.In Rodman, P. S., and Cant, J. (eds.), Adaptations for Foraging in Non-human Primates,Contributions to an Organismal Biology of Prosimians, Monkeys and Apes, ColombiaUniversity Press, New York, pp. 161-194.
Isabirye-Basuta, G. (1988). Food competition among individuals in a free-ranging chimpanzeecommunity in Kibale forest, Uganda. Behaviour 105: 135-147.
Leighton, M., and Leighton, D. R. (1982). The relationship of size and of feeding aggregateto size of food patch: Howler monkeys (Alouatta palliata) feeding in Trichilia cipo fruittrees on Barro Colorado Island. Biotropica 14: 81-90.
Machlis, L., Dodd, P. W., and Fentress, J. C. (1985). The pooling fallacy: problems arisingwhen individuals contribute more than one observation to the data set. Z. Tierpsychol.68: 201-214.
MacKinnon, J. R. (1974). The ecology and behavior of wild orang-utans (Pongo pygmaeus).Anim. Behav. 22: 3-74.
Martin, P. , and Bateson, P. (1986). Measuring Behavior, Cambridge University Press,Cambridge.
Mitani, J. C. (1985). Mating behavior of male orangutans in the Kutai Reserve, EastKalimantan, Indonesia. Anim. Behav. 33: 392-402.
Mitani, J. C, Grether, G. F., Rodman, P. S., and Priatna, D. (1991). Associations among wildorang-utans: sociality, passive aggregations or chance? Anim. Behav. 42: 33-46.
Mitrasetia, T. (1995). Hirarki dominansi orangutan jantan dewasa serta tanggapan orangutanjantan dan betina dewasa terhadap seruan panjang dan jantan dewasa lain di PusatPenelitian Ketambe, Taman Nasional Gunung Leuser, Master's thesis, UniversitasIndonesia, Jakarta.
Nishida, T., and Hiraiwa-Hasegawa, M. (1987). Chimpanzees and bonobos: Cooperativerelationships among males. In Smuts, B. B., Cheney, D. L., Seyfarth, R. M., Wrangham,R. W., and Struhsaker, T. T. (eds.), Primate Societies, University of Chicago Press,Chicago, pp. 165-177.
Rijksen, H. D. (1978). A Field Study on Sumatran Orangutans (Pongo pygmaeus abelli Lesson1827): Ecology, Behavior and Conservation, Veenman, Wageningen.
Robinson, J. G., and Janson, C. H. (1987). Capuchins, squirrel monkeys, and atelines:Socioecological convergence with old world monkeys. In Smuts, B. B., Cheney, D. L.,Seyfarth, R. M., Wrangham, R. W., and Struhsaker, T. T. (eds.), Primate Societies,University of Chicago Press, Chicago, pp. 69-82.
Rodman, P. S. (1979). Individual activity profiles and the solitary nature of orangutans. InHamburg, D. A., and McCown, E. R. (eds.), The Great Apes, Benjamin/Cumings, MenloPark, CA, pp. 234-255.
Rodman, P. S. (1984). Foraging and social systems of orangutans and chimpanzees. InRodman, P. S., and Cant, J. G. H. (eds.), Adaptations for Foraging in Nonhuman Primates,Contributions to an Organismal Biology of Prosimians, Monkeys and Apes, ColumbiaUniversity Press, New York, pp. 54-72.
Rodman, P. S., and Mitani, J. C. (1987). Orangutans: Sexual dimorphism in a solitary species.In Smuts, B. B., Cheney, D. L., Seyfarth, R. M., Wrangham, R. W., and Struhsaker, T.T. (eds.), Primate Societies, University of Chicago Press, Chicago, pp. 146-154.
SAS (1988). SAS/STAT User's Guide. Release 6.03 ed.Schurmann, C. L. (1982). Courtship and mating behavior of wild orangutans in Sumatra. In
Chiarelli, A. B., and Corruccini, R. S. (eds.), Primate Behavior and Sociobiology, Springer,Berlin, pp. 129-135.
Schurmann, C. L., and van Hooff, J. A. R. A. M. (1986). Reproductive strategies of theorang-utan: New data and a reconsideration of existing socio-sexual models. Int. J.Primatol. 1: 265-287.

Food Competition in Orangutans 927
Searl, S. R., Speed, F. M., and Milliken, G. A. (1980). Population marginal means in thelinear model: An alternative to least square means. Am. Stat. 34: 216-221.
Strier, K. B. (1989) Effects of patch size on feeding associations in muriquis (Brachytdesarachnoides). Folia Primatologica 52: 70-77.
Sugardjito, J. (1986). Ecological Constraints on the Behavior of Sumatran Orang-utans (Pongopygmaeus abelii) in the Gunung Leaser National Park, Indonesia, Ph.D. thesis, UtrechtUniversity, Utrecht.
Sugardjito, J., te Boekhorst, I. J. A., and van Hooff, J. A. R. A. M. (1987). Ecologicalconstraints on grouping of wild orangutans (Pongo pygmaeus) in the Gunung LeuserNational Park, Sumatera, Indonesia. Int. J. Primatol. 8: 17-41.
Symington. M. McF. (1988). Food competition and foraging party size in the black spidermonkey (Ateles paniscus chamek). Behaviour 105: 117-133.
te Boekhorst, I. J. A., and Hogeweg, P. (1994). Self-structuring in artificial "chimps" offersnew hypotheses for male grouping in chimpanzees. Behaviour 130: 229-252.
van Schaik, C. P. (1983). Why are diurnal primates living in groups? Behaviour 87: 120-144.van Schaik, C. P. (1989). The ecology of social relationships amongst female primates. In
Standen, V., and Foley, R. A. (eds.), Comparative Socioecology, the Behavioral Ecology ofHumans and Mammals, Blackwell Scientific, Oxford, pp. 195-218.
van Schaik, C. P., and Mirmanto, E. (1985). Spatial variation in the structure and litterfall ofa Sumatran rainforest. Biotropica 17: 196-205.
van Schaik, C. P., and van Noordwijk, M. A. (1986). The hidden costs of sociality: Intra-groupvariation in feeding strategies in Sumatran long-tailed macaques (Macaca fascicularis).Behaviour 99: 296-315.
Wheatly, B. P. (1982). Energetics of foraging in Macaca fascicularis and Pongo pygmaeus anda selective advantage of large body size in the orang-utan. Primates 23: 348-363.
White, F. J., and Wrangham, R. W. (1988). Feeding competition and patch size in thechimpanzee species Pan paniscus and Pan troglodytes. Behaviour 105: 148-164.





























