Embed Size (px)
Citation preview

ORIGINAL PAPER
Functional characterization of GDP-mannose pyrophosphorylasefrom Leptospira interrogans serovar Copenhageni
Matıas D. Asencion Diez • Ana Demonte • Jorge Giacomelli • Sergio Garay •
Daniel Rodrıgues • Birgit Hofmann • Hans-Juerguen Hecht • Sergio A. Guerrero •
Alberto A. Iglesias
Received: 4 June 2009 / Revised: 3 December 2009 / Accepted: 7 December 2009
� Springer-Verlag 2009
Abstract Leptospira interrogans synthesizes a range of
mannose-containing glycoconjugates relevant for its viru-
lence. A prerequisite in the synthesis is the availability of
the GDP-mannose, produced from mannose-1-phosphate
and GTP in a reaction catalyzed by GDP-mannose pyro-
phosphorylase. The gene coding for a putative enzyme
in L. interrogans was expressed in Escherichia coli
BL21(DE3). The identity of this enzyme was confirmed by
electrospray-mass spectroscopy, Edman sequencing and
immunological assays. Gel filtration chromatography
showed that the dimeric form of the enzyme is catalyti-
cally active and stable. The recombinant protein was
characterized as a mannose-1-phosphate guanylyltransfer-
ase. S0.5 for the substrates were determined both in GDP-
mannose pyrophosphorolysis: 0.20 mM (GDP-mannose),
0.089 mM (PPi), and 0.47 mM; and in GDP-mannose
synthesis: 0.24 mM (GTP), 0.063 mM (mannose-1-phos-
phate), and 0.45 mM (Mg2?). The enzyme was able to
produce GDP-mannose, IDP-mannose, UDP-mannose and
ADP-glucose. We obtained a structural model of the
enzyme using as a template the crystal structure of man-
nose-1-phosphate guanylyltransferase from Thermus ther-
mophilus HB8. Binding of substrates and cofactor in the
model agree with the pyrophosphorylases reaction mech-
anism. Our studies provide insights into the structure of
a novel molecular target, which could be useful for
detection of leptospirosis and for the development of anti-
leptospiral drugs.
Keywords Leptospira interrogans �Mannose metabolism � Pyrophosphorylase
Introduction
Leptospirosis is considered to be the most widespread
zoonosis in the world, which is reflecting the ability of
pathogenic Leptospira interrogans species to adapt to the
renal tubules of a wide variety of mammalian reservoirs
(Barnett et al. 1999; Haake et al. 1999). Human lepto-
spiral infection frequently results in a hard life-threatening
illness characterized by liver dysfunction, kidney failure,
and pulmonary hemorrhage (Barnett et al. 1999; Haake
Communicated by Sebastian Suerbaum.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00203-009-0534-3) contains supplementarymaterial, which is available to authorized users.
M. D. Asencion Diez � J. Giacomelli � S. A. Guerrero
Laboratorio de Bioquımica Microbiana,
Universidad Nacional del Litoral, Santa Fe, Argentina
A. Demonte � A. A. Iglesias (&)
Laboratorio de Enzimologıa Molecular,
Instituto de Agrobiotecnologıa del Litoral, FBCB,
Universidad Nacional del Litoral,
S3000ZAA Santa Fe, Argentina
e-mail: [email protected]
S. Garay � D. Rodrıgues
Departamento de Fısica, Facultad de Bioquımica y Ciencias
Biologicas, Universidad Nacional del Litoral,
Santa Fe, Argentina
B. Hofmann � H.-J. Hecht
Helmholtz Centre for Infection Research,
Inhoffenstraße 7, 38124 Braunschweig, Germany
Present Address:J. Giacomelli
Laboratorio de Biologıa Vegetal. Facultad de Bioquımica y
Ciencias Biologicas, Universidad Nacional del Litoral,
Santa Fe, Argentina
123
Arch Microbiol
DOI 10.1007/s00203-009-0534-3

et al. 1999). The present treatment for acute cases of
leptospirosis has variable effectiveness (Griffith et al.
2006, 2007; Zunino and Pizarro 2007). Characterization
of novel molecular targets for the design of new inhibitors
will surely help in the therapy of the disease. The car-
bohydrate metabolism is one of the biochemical pathways
to analyze in depth for this bacterium, not only for the
rational design of new therapeutic agents but for a better
knowledge of the metabolic pathway and its relevance on
the Leptospira sp. virulence, pathogenesis and host
immune response (Saavedra-Lira and Perez-Montfort
1996; Opperdoes and Michels 2001; Verlinde et al. 2001;
Gornik et al. 2006).
Different mannose-containing compounds are present in
the structure of the cell wall from bacteria, yeast, and lower
eukaryotic organisms (Preston et al. 1996; Kobayashi et al.
1997; Martins et al. 1999; Ning and Elbein 1999; Garami
and Ilg 2001). In many Gram-negative bacteria, mannose
derivative lipopolysaccharides (LPS) are highly toxic and
immunogenic molecules representing major components of
cell membranes (Preston et al. 1996). In Leptospira and
other pathogens (as in Yersinia enterocolitica), LPS con-
stitute the main virulence factor (Zhang et al. 1997; Bulach
et al. 2000a, 2000b; Bharti et al. 2003). In this context,
GDP-D-mannose (GDP-Man) is a key metabolite, as it is
the sugar activated form functioning as the mannosyl group
donor in the synthesis of several mannose-containing
structures (Ginsburg 1964; Ning and Elbein 1999; Ning
and Elbein 2000). GDP-Man is synthesized from mannose-
1-phosphate (Man-1-P) and GTP in a reaction catalyzed by
a specific sugar nucleotide pyrophosphorylase, namely
GDP-Man pyrophosphorylase (EC 2.7.7.22; GDP-ManP-
Pase). In this work, we present the cloning of the gene
encoding a putative GDP-ManPPase from genomic DNA
of Leptospira interrogans serovar Copenhageni, the
expression of the recombinant protein in Escherichia coli
and its chromatographic purification for kinetic and struc-
tural characterization. In addition, we report here a model
of the 3D structure of the enzyme obtained by homology
modeling using as a template the crystal structure of
homologous enzyme from Thermus thermophilus HB8.
Finally, we show that GDP-ManPPase represents a key
immunogenic target for detecting leptospirosis infection in
humans.
Materials and methods
Chemicals
All protein standards, antibiotics, isopropyl-b-thiogalacto-
side (IPTG), and oligonucleotides were of the highest
quality available.
Bacterial strains, growth conditions, and media
Bacterial strains used in this study were E. coli TOP 10 F0
and E. coli BL21(DE3) pLysS (Invitrogen). A protocol
previously described (Veloso et al. 2000) was used to
extract leptospiral DNA from bacteria grown axenically at
the Instituto Nacional de Enfermedades Respiratorias ‘‘Dr.
Emilio Coni’’, Santa Fe, Argentina.
All strains of E. coli were grown at 37�C in Luria–
Bertani broth (LB, per liter 10 g tryptone, 5 g yeast extract
and 10 g NaCl, pH 7.4). Solid media had agar–agar in a
concentration of 18 g/l. Antibiotics were used at the fol-
lowing final concentrations: 100 lg/ml ampicillin, 34 lg/
ml chloramphenicol. Protein expression was induced by
adding IPTG at a final concentration of 0.5 mM.
Cloning of the gene
Amplification of the gmp gene (NCBI acc No. NC005823),
coding for GDP-ManPPase was performed using primers
gmpfo: 50GAATTCATGAACCAAGACAAACCGGT30 and
gmpre: 50AAGCTTCTATTCCGTGTACTTTTGTA30, which
included recognition sites for EcoRI and HindIII, respec-
tively. PCR was performed in 19 Taq PCR Buffer (Invit-
rogen) containing 2 mM MgCl2, 50 ng template DNA,
200 nM of each primer, 0.2 mM dNTPs, 1 U DNA poly-
merase (Invitrogen), in a final volume of 50 ll. The ther-
mocycle program was: 94�C for 5 min, then 30 cycles
including denaturation at 94�C for 1 min, annealing at
60�C for 1 min and elongation at 72�C for 1 min 30 s,
followed by a final elongation step of 10 min. The PCR
product was purified with the QIAquick PCR purification
kit (QIAGEN) and cloned into the pGEM-T Easy cloning
vector (Promega) according to the manufacturer’s instruc-
tions. The gmp identity was confirmed by complete
sequencing. The construction [pGEM-TEasy/gmp] was
digested with EcoRI and HindIII, to release gmp which was
purified and cloned into an EcoRI/HindIII–digested
pRSETB vector. Consequently, 44 amino acids (including
six histidines and an enterokinase cleavage site) were
added to the N-terminal end of the recombinant protein.
The new construction, [pRSETB/gmp] was renamed
pGMP. Competent E. coli BL21(DE3) pLysS cells were
transformed with pGMP yielding the E. coli BL21 (DE3)
pLysS [pGMP] for protein expression.
Overproduction of GDP-ManPPase
E. coli BL21 (DE3) pLysS [pGMP] cells were grown
overnight at 37�C with shaking at 180 rpm in LB broth
supplemented with 100 lg/ml ampicillin and 34 lg/ml
chloramphenicol. The overnight culture was diluted 1/100
Arch Microbiol
123

in fresh media and grown under identical conditions to
exponential phase (OD600 of 0.6). The expression of the
recombinant protein was induced by adding IPTG at a
final concentration of 0.5 mM, followed by incubation at
30�C for 4 h. Cells were harvested by centrifugation,
washed once with ice-cold 50 mM Tris–HCl buffer, pH
8.0, and suspended in an extraction buffer containing
50 mM Tris–HCl, pH 8.0, 300 mM NaCl, and 1 mM
PMSF. After disruption using a Cell Disruptor (Constant
Systems), the crude extract was clarified by centrifugation
during 1 h at 15,0009g and then filtered through a 45-lm
membrane.
Purification of His-tagged GDP-ManPPase
GDP-ManPPase from L. interrogans was expressed in E.
coli with an N-terminal His-tag, which facilitates its further
purification. The enzyme was purified as by affinity chro-
matography, using Talon-Co?2 Superflow resin (Clontech)
according to the protocol supplied by the manufacturer.
The elution fractions containing the recombinant protein
were analyzed electrophoretically by SDS–PAGE (Lae-
mmli 1970) to check for purity. The final purification step
was carried out by gel filtration with a Superdex-200 FPLC
column (Amersham Pharmacia Biotech). The column was
equilibrated with 50 mM Tris–HCl buffer, pH 8.0, and
300 mM NaCl.
Protein detection and quantification
Protein concentration was determined by the method of
Bradford (1976) using BSA as standard. To reveal SDS–
PAGE gels, proteins were stained with Coomassie brilliant
blue.
Proteolytic cleavage
To remove the N-terminal His-Tag, the recombinant
GDP-ManPPase was treated with enterokinase (Novagen).
The cleavage was performed in 300 ll using the buffer
provided with the protease, 0.5 mg GDP-ManPPase, and
6 U enterokinase. The reaction was stopped by adding
1 mM PMSF. Complete cleavage was confirmed by SDS–
PAGE, with the difference between both forms of the
enzyme being in 4,600 Da. After cleavage, GDP-ManP-
Pase was purified by gel filtration by the procedure
described below.
Enzyme activity assay
GDP-ManPPase activity was determined at 37�C both, in
the GDP-Man pyrophosphorolysis (assay A) and synthesis
(assay B) directions.
Assay A
Pyrophosphorolysis of GDP-Man was followed by the
formation of [32P]GTP from [32P]PPi, as previously
described for other pyrophosphorylases (Ghosh and Preiss
1966). The reaction mixture contained 50 mM MOPS
buffer (pH 7.5), 5 mM MgCl2, 1.5 mM GDP-mannose,
1 mM [32P]PPi (ca. 3,000 cpm/nmol), 10 mM NaF, 0.2%
(w/v) bovine serum albumin, and enzyme in a total volume
of 150 ll. The reaction was started by the addition of
[32P]PPi, and after 10 min of incubation at 37�C it was
terminated by the addition of 1 ml of cold 5% (W/V) tri-
chloroacetic acid. The [32P]GTP formed was measured as
described by Ghosh and Preiss (1966).
Assay B
Synthesis of GDP-Man was assayed by following the for-
mation of Pi (after hydrolysis of PPi by inorganic pyro-
phosphatase) by a highly sensitive colorimetric method
previously described (Fusari et al. 2006). The reaction
mixture contained 50 mM MOPS buffer (pH 7.5), 5 mM of
MgCl2, 1 mM of GTP, 0.1 U of inorganic pyrophospha-
tase, 0.2% (w/v) bovine serum albumin and variable
amounts of the enzyme. Assays were initiated by addition
of 1 mM Man-1-P in a total volume of 50 ll. The reaction
mixture was incubated for 10 min at 37�C and terminated
by adding color reagent [Malachite Green, see (Fusari et al.
2006)]. The complex formed with the released Pi was
measured at 630 nm with an ELISA EMax detector
(molecular devices).
Under the conditions described earlier, one unit of
activity (U) is defined as the amount of enzyme catalyzing
the formation of 1 lmol of product (either GTP or PPi, for
assay A or B, respectively) per min. The kinetic constants
were determined by fitting the data with a non-linear least-
squares formula and the Hill equation using the program
OriginTM. Kinetic constants are the mean of at least two
independent sets of data, which were reproducible within
±10%.
Western blotting
Western blotting was performed after standard techniques
(Maniatis et al. 1982). Proteins in the gel were blotted onto
PVDF membranes using a Mini-PROTEANII (Bio-Rad)
apparatus. The membrane was blocked overnight at 4�C,
subsequently incubated with primary antibody at room
temperature for 1 h, and then incubated with a HRP-con-
jugated anti–human secondary antibody for 40 min.
Detection was carried out with 3,30-diaminobenzidine and
hydrogen peroxide (Sigma) in 50 mM Tris–HCl, pH 8.0,
150 mM NaCl. Sera from patients with leptospirosis
Arch Microbiol
123

(obtained from Instituto Nacional de Enfermedades Res-
piratorias ‘‘Dr Emilio Coni’’, Santa Fe, Argentina) were
used as the primary antibody.
Determination of native molecular mass
The native molecular mass of GDP-ManPPase was deter-
mined by gel filtration chromatography in a Superdex-200
column previously equilibrated with buffer 50 mM
Tris–HCl pH 8.0, and 300 mM NaCl. Ribonuclease A
(Mr = 13,700), chymotrypsinogen A (Mr = 25,000),
ovoalbumin (Mr = 43,000), bovine serum albumin
(Mr = 67,000) and aldolase (Mr = 158,000) were used as
molecular mass standards. One milligram of pure protein
was applied to the column. Masses were extrapolated from
the standard semi-log curve log Mr vs Kav (a parameter
defined by the following equation: Kav ¼ Ve�Vo
Vc�Vo; where Ve is
the elution volume, Vo is the void volume and Vc is the
column volume.
Electrospray-MS and Edman sequencing
All samples analyzed by ESI–MS and Edman sequencing
were buffer-changed to 10 mM NH4HCO3-NaOH, pH 8.0
by ultracentrifugation and analyzed at the Mass Spectrom-
etry Center-Biophysical Analysis Research Group in the
Helmholtz Institute for Infection Research (Braunschweig,
Germany) as previously described (Jaeckel et al. 2005).
ES–MS was performed with an ion trap mass spectrometer
(ESQUIRE-LC, Bruker Daltonik, Bremen, Germany).
N-terminal amino acid sequence analysis by Edman deg-
radation was carried out on a 476A protein sequencer
(Applied Biosystems, Weiterstadt, Germany).
Homology modeling
The 3D structure database RCSB-PDB (Berman et al.
2000) and template identification tools from the Swiss
Model Workspace (Arnold et al. 2006) were used for
sequence search. Sequence alignment was made with
Clustal X 1.81 (Thompson et al. 1994), and its assessment
was evaluated with the server ESPript 2.2 (Gouet et al.
1999). The homology model of L. interrogans GDP-
ManPPase was built with the software Modeller 8v2 (Sali
and Blundell 1993) and refined with 5,000 cycles of
steepest descent algorithm using the crystal structure of
GDP-ManPPase from T. thermophilus HB8 (code 2cu2) as
a template. The evaluation of the final model was done
with the structure assessment tool from the Swiss Model
Workspace, which includes Anolea, Verify3D, and What-
check analysis. The 3D model for the dimeric structure of
leptospiral GDP-ManPPase was built using the symmetry
operations of the crystallographic space group (Guex and
Peitsch 1997) of the specified T. thermophilus HB8 enzyme
template structure (Sugahara 2005). The substrate Man-1-P
was fitted manually in such a way that the hydroxyl group
on the C4 interacts with the oxygen of E174 and one of the
phosphate’s oxygen with the nitrogen of the residue K175
as described before (Jin et al. 2005); whereas the GTP
molecule was positioned on the structure using as a tem-
plate the position of ATP in the structure of potato tuber
ADP-GlcPPase (pdb-code _1_YP3) (Jin et al. 2005), which
was crystallized with the ATP in its binding site. After we
made a structural alignment of our model onto the _1_YP3
using the program STAMP (Russell and Barton 1992), the
GTP molecule was added in the model keeping the orien-
tation of the pentose, the triphosphate moiety and over-
lapping the purine base of GTP with that one of ATP.
Results and discussion
Isolation and heterologous expression of the gmp gene
The identification of a nucleotide sequence (geneID
2770489) encoding a putative GDP-ManPPase (gmp gene)
in the database of the L. interrogans genome project
(Leptospira interrogans serovar Copenhageni Genome
Project (NCBI access NC_005823 and NC_005824)
prompted us to perform the molecular cloning of the gene
full-length. The gene (1,062 bp in length) was amplified
from genomic DNA and its identity was confirmed by
DNA sequencing in both directions. It is predicted that gmp
encodes a protein of 353 amino acids, with a molecular
mass of 40.2 kDa and a calculated pI of 9.35.
We generated the construction pGMP (renamed from
pRSETB/gmp, see details under ‘‘Materials and methods’’),
which was used to transform competent E. coli BL21(DE3)
pLysS cells. After inducing expression and obtaining crude
extracts from the transformed cells, the recombinant protein
was 76-fold purified by affinity chromatography, the
enzyme reaching a specific activity of 3.8 U/mg (Table 1).
The analysis by SDS–PAGE showed in Fig. 1 indicates that
the recombinant enzyme reached a high degree of purifi-
cation and revealed a molecular mass of 45 kDa (Fig. 1a).
This mass corresponds to approximately 40 kDa for GDP-
ManPPase plus an enlargement of ca. 5 kDa caused by the
histidine-tag and extra amino acids, including the entero-
kinase cleavage site. This is reinforced by data obtained
after enterokinase digestion of the recombinant protein
(Fig. 1b). The native molecular mass of the GDP-ManP-
Pase forms (before and after enterokinase treatment) was
determined by gel filtration chromatography to correspond
to a dimeric arrangement in both cases (data not shown).
Interestingly, the purified enzyme was recognized by spe-
cific antibodies present in typified sera pool from patients
Arch Microbiol
123
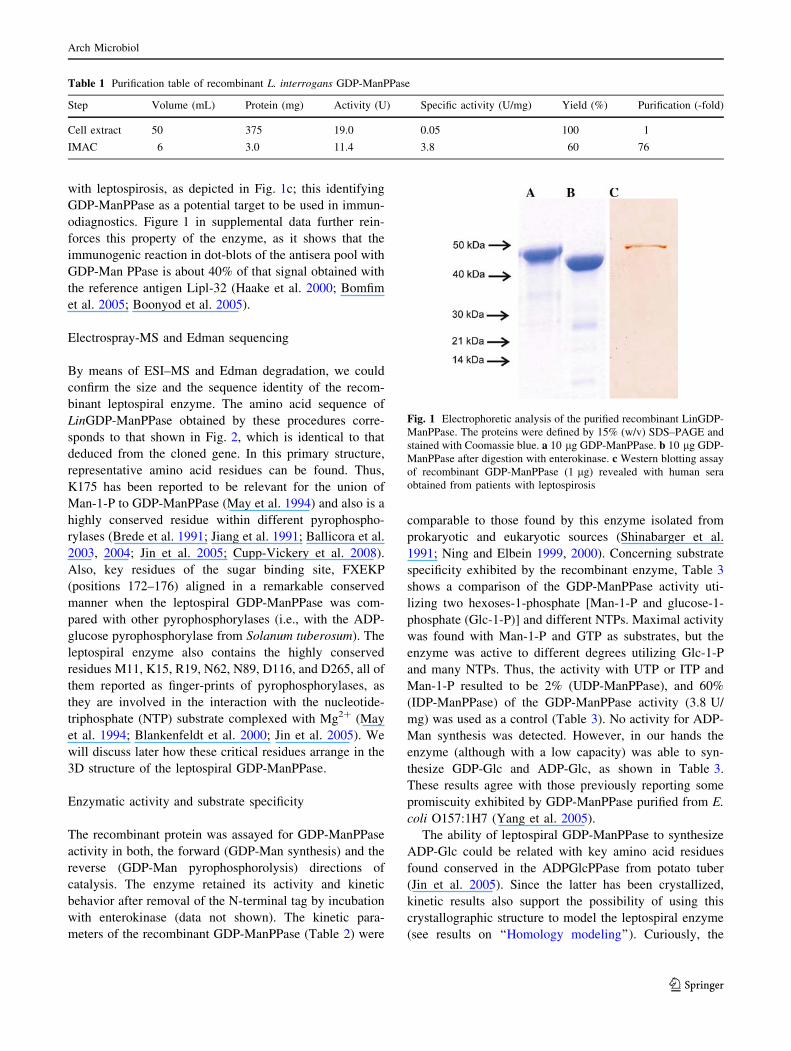
with leptospirosis, as depicted in Fig. 1c; this identifying
GDP-ManPPase as a potential target to be used in immun-
odiagnostics. Figure 1 in supplemental data further rein-
forces this property of the enzyme, as it shows that the
immunogenic reaction in dot-blots of the antisera pool with
GDP-Man PPase is about 40% of that signal obtained with
the reference antigen Lipl-32 (Haake et al. 2000; Bomfim
et al. 2005; Boonyod et al. 2005).
Electrospray-MS and Edman sequencing
By means of ESI–MS and Edman degradation, we could
confirm the size and the sequence identity of the recom-
binant leptospiral enzyme. The amino acid sequence of
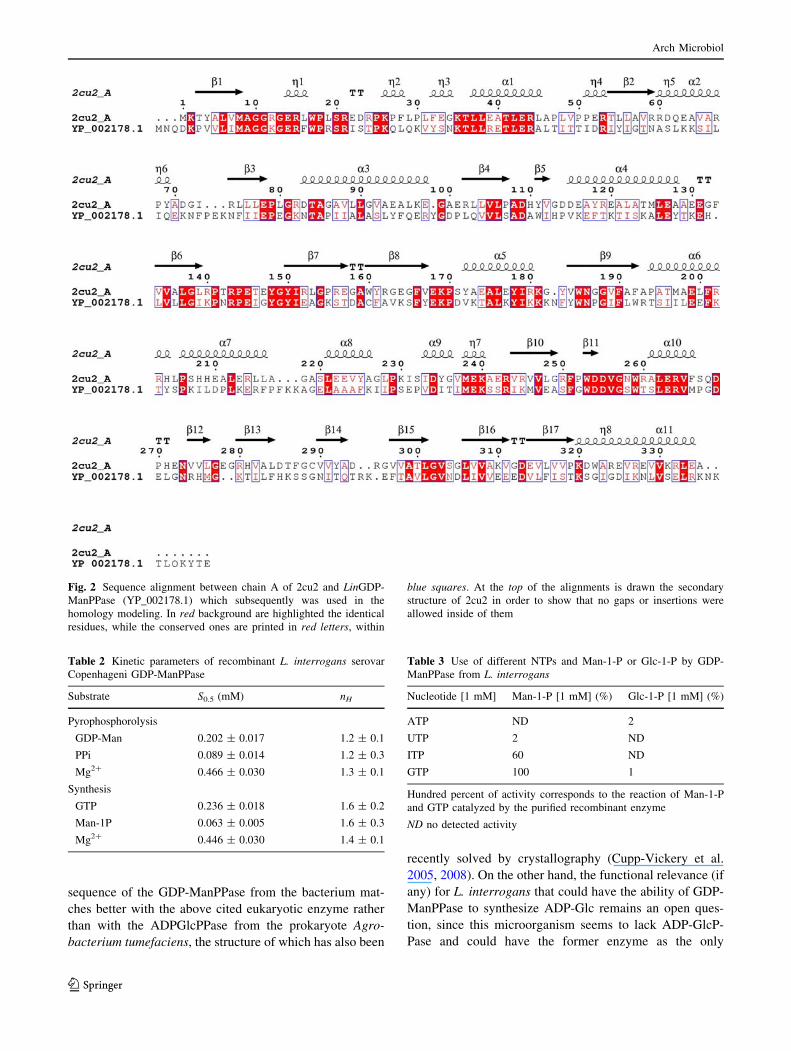
LinGDP-ManPPase obtained by these procedures corre-
sponds to that shown in Fig. 2, which is identical to that
deduced from the cloned gene. In this primary structure,
representative amino acid residues can be found. Thus,
K175 has been reported to be relevant for the union of
Man-1-P to GDP-ManPPase (May et al. 1994) and also is a
highly conserved residue within different pyrophospho-
rylases (Brede et al. 1991; Jiang et al. 1991; Ballicora et al.
2003, 2004; Jin et al. 2005; Cupp-Vickery et al. 2008).
Also, key residues of the sugar binding site, FXEKP
(positions 172–176) aligned in a remarkable conserved
manner when the leptospiral GDP-ManPPase was com-
pared with other pyrophosphorylases (i.e., with the ADP-
glucose pyrophosphorylase from Solanum tuberosum). The
leptospiral enzyme also contains the highly conserved
residues M11, K15, R19, N62, N89, D116, and D265, all of
them reported as finger-prints of pyrophosphorylases, as
they are involved in the interaction with the nucleotide-
triphosphate (NTP) substrate complexed with Mg2? (May
et al. 1994; Blankenfeldt et al. 2000; Jin et al. 2005). We
will discuss later how these critical residues arrange in the
3D structure of the leptospiral GDP-ManPPase.
Enzymatic activity and substrate specificity
The recombinant protein was assayed for GDP-ManPPase
activity in both, the forward (GDP-Man synthesis) and the
reverse (GDP-Man pyrophosphorolysis) directions of
catalysis. The enzyme retained its activity and kinetic
behavior after removal of the N-terminal tag by incubation
with enterokinase (data not shown). The kinetic para-
meters of the recombinant GDP-ManPPase (Table 2) were
comparable to those found by this enzyme isolated from
prokaryotic and eukaryotic sources (Shinabarger et al.
1991; Ning and Elbein 1999, 2000). Concerning substrate
specificity exhibited by the recombinant enzyme, Table 3
shows a comparison of the GDP-ManPPase activity uti-
lizing two hexoses-1-phosphate [Man-1-P and glucose-1-
phosphate (Glc-1-P)] and different NTPs. Maximal activity
was found with Man-1-P and GTP as substrates, but the
enzyme was active to different degrees utilizing Glc-1-P
and many NTPs. Thus, the activity with UTP or ITP and
Man-1-P resulted to be 2% (UDP-ManPPase), and 60%
(IDP-ManPPase) of the GDP-ManPPase activity (3.8 U/
mg) was used as a control (Table 3). No activity for ADP-
Man synthesis was detected. However, in our hands the
enzyme (although with a low capacity) was able to syn-
thesize GDP-Glc and ADP-Glc, as shown in Table 3.
These results agree with those previously reporting some
promiscuity exhibited by GDP-ManPPase purified from E.
coli O157:1H7 (Yang et al. 2005).
The ability of leptospiral GDP-ManPPase to synthesize
ADP-Glc could be related with key amino acid residues
found conserved in the ADPGlcPPase from potato tuber
(Jin et al. 2005). Since the latter has been crystallized,
kinetic results also support the possibility of using this
crystallographic structure to model the leptospiral enzyme
(see results on ‘‘Homology modeling’’). Curiously, the
Table 1 Purification table of recombinant L. interrogans GDP-ManPPase
Step Volume (mL) Protein (mg) Activity (U) Specific activity (U/mg) Yield (%) Purification (-fold)
Cell extract 50 375 19.0 0.05 100 1
IMAC 6 3.0 11.4 3.8 60 76
Fig. 1 Electrophoretic analysis of the purified recombinant LinGDP-
ManPPase. The proteins were defined by 15% (w/v) SDS–PAGE and
stained with Coomassie blue. a 10 lg GDP-ManPPase. b 10 lg GDP-
ManPPase after digestion with enterokinase. c Western blotting assay
of recombinant GDP-ManPPase (1 lg) revealed with human sera
obtained from patients with leptospirosis
Arch Microbiol
123
sequence of the GDP-ManPPase from the bacterium mat-
ches better with the above cited eukaryotic enzyme rather
than with the ADPGlcPPase from the prokaryote Agro-
bacterium tumefaciens, the structure of which has also been
recently solved by crystallography (Cupp-Vickery et al.
2005, 2008). On the other hand, the functional relevance (if
any) for L. interrogans that could have the ability of GDP-
ManPPase to synthesize ADP-Glc remains an open ques-
tion, since this microorganism seems to lack ADP-GlcP-
Pase and could have the former enzyme as the only
Fig. 2 Sequence alignment between chain A of 2cu2 and LinGDP-
ManPPase (YP_002178.1) which subsequently was used in the
homology modeling. In red background are highlighted the identical
residues, while the conserved ones are printed in red letters, within
blue squares. At the top of the alignments is drawn the secondary
structure of 2cu2 in order to show that no gaps or insertions were
allowed inside of them
Table 2 Kinetic parameters of recombinant L. interrogans serovar
Copenhageni GDP-ManPPase
Substrate S0.5 (mM) nH
Pyrophosphorolysis
GDP-Man 0.202 ± 0.017 1.2 ± 0.1
PPi 0.089 ± 0.014 1.2 ± 0.3
Mg2? 0.466 ± 0.030 1.3 ± 0.1
Synthesis
GTP 0.236 ± 0.018 1.6 ± 0.2
Man-1P 0.063 ± 0.005 1.6 ± 0.3
Mg2? 0.446 ± 0.030 1.4 ± 0.1
Table 3 Use of different NTPs and Man-1-P or Glc-1-P by GDP-
ManPPase from L. interrogans
Nucleotide [1 mM] Man-1-P [1 mM] (%) Glc-1-P [1 mM] (%)
ATP ND 2
UTP 2 ND
ITP 60 ND
GTP 100 1
Hundred percent of activity corresponds to the reaction of Man-1-P
and GTP catalyzed by the purified recombinant enzyme
ND no detected activity
Arch Microbiol
123

alternative to produce the sugar-nucleotide if necessary.
However, in the genome database of L. interrogans no
gene coding for enzymes involved in glycogen synthesis is
found, which makes doubtful the use of ADP-Glc by this
microorganism.
Homology modeling
With the aim of obtaining an in silico 3D structure of GDP-
ManPPase from L. interrogans, we searched for an accu-
rate template using bioinformatic tools specified under
‘‘Materials and methods’’. After comparing our target
sequence against the ExPDB (template library extracted
from PDB) using the gapped BLAST algorithm (Altschul
et al. 1997), we found two homologous proteins as the
more appropriate to templates for modeling. These two
proteins were: 2cu2, the GDP-ManPPase from T. thermo-
philus Hb8 (Sugahara 2005); and 2qh5, the mannose-6-
phosphate isomerase from Helicobacter pylori. We decided
to employ 2cu2 as a template because it matched several
more residues than 2qh5 with our target enzyme (102
identical residues against 77) and it also had fewer gaps (10
vs. 22), thus minimizing the probability of producing a
poor model (Larsson et al. 2008). Figure 2 shows the
alignment used in the homology modeling, and where the
GDP-ManPPase from T. thermophilus Hb8 (Sugahara
2005) exhibited 29% of identity and 47% of similarity
when compared with the enzyme form L. interrogans. In
selecting the appropriate template, special care was taken
to avoid the presence of breaks in the regions being part of
regular secondary structure motifs.
Utilizing the specified 2cu2 template and considering
the quaternary structure determined in the present study for
the recombinant enzyme, a 3D structural model was
established for the dimeric GDP-ManPPase from
L. interrogans (Fig. 3). As depicted in Fig. 3, the subunits
are arranged in a head-to-tail fashion. This characteristic
form of quaternary fold has also been documented for the
complex ATP-phosphoribosyl transferase from Thermoto-
ga maritima (Vega et al. 2005). Evaluation of the final 3D
structure showed in Fig. 3, utilizing Verify-3D (Bowie
et al. 1991; Luthy et al. 1992), indicated that our model
reached a Z-score value of 118, which is between the best
hope, considering its sequence length (353 residues) and
the lower limit of confidence (72). The calculated ANO-
LEA (Melo and Feytmans 1998) profile showed few
regions with positive values, mainly in loops. In addition,
the Ramachandran plot (Ramachandran et al. 1963) pointed
out that the overall conformation of the backbone was
reliable (less than 2% of the non-glycine residues in non
allowed areas).
Taking an insight look respect to how the substrates fit
in the GDP-ManPPase model, it is observed that the con-
sensus binding site of the sugar moiety exhibited a similar
orientation as that found in ADPGlcPPase from potato
tuber (Jin et al. 2005). This is in agreement with the above
described promiscuity of the leptospiral enzyme to use
different substrates, and is also supported by a previous
report (May et al. 1994) demonstrating that GDP-ManP-
Pase from Pseudomonas aeruginosa contains a catalytic
domain homologous to the Glc-1-P site found in other
sugar-nucleotide pyrophosphorylases. Also, a previous
analysis demonstrated that, even when the similarity
between GDP-ManPPases and ADP-GlcPPases is extre-
mely low (identity 8.8%), an alignment can be done in a
form that predicted structures match helices and sheets
(Ballicora et al. 2003). Using this approach, we obtained
the sequence alignment shown in Fig. 4, where the con-
servation state of each residue in key regions is represented
as histogram graphic.
By applying the structural alignment defined in Fig. 4, it
was possible to superimpose monomeric forms of our 3D
model of the leptospiral GDP-ManPPase (shown by Fig. 3)
onto the _1_YP2 structure of potato tuber ADP-GlcPPase,
as illustrated by Fig. 5. The residues of both structures
were colored according to their sequence similarity, using
BLOSUM 60 substitution matrix. Thick blue cylinders
show the most conserved residues, while thin red cylinders
the least ones (Fig. 5). Reinforcing the models derived
from this approach, the higher bars obtained in the histo-
gram graphic related with sequence alignment (shown in
Fig. 4) correspond to the latter specified thick blue cylin-
ders. The similarity of the binding site (residues in blue
around ATP) explains why we used the ATP coordinates
from _1_YP2 to insert our GTP in the pocket of our model.
Figure 6 is derived from the above detailed superim-
position of structures (Fig. 5), where the relative orienta-
tion of conserved residues FXEKP is compared between
Fig. 3 Cartoon renderization of the proposed dimeric form for the
GDP-ManPPase. b-sheets and a-helices are shown as arrows and
ribbons, respectively. The loops connectors are drawn as cylinders
Arch Microbiol
123

Arch Microbiol
123

the leptospiral GDP-ManPPase (yellow/light) and the
potato tuber ADP-GlcPPase _1_YP2 structure (red/dark).
On the other hand, following the same substrate positioning
strategy, the Man-1-P moiety locates far from residues
F172, Y173 and P176. The length of the Man-1-P (8.5 A)
allows for its interaction with at most two of the four
consecutive residues that have been already mentioned as
the motif of the sugar-P binding site. We found the distance
from OE1 of E174 to the NZ of K175 also appropriate for
interaction with Man-1P in the enzyme active site. We can
assume that other conserved residues are important for the
correct folding of this portion of the molecule. The Man-1-
P moiety could thus locate close to residues D244, I245,
and E249 which are highly conserved between several
pyrophosphorylases (Jin et al. 2005). Also, D244 and E249
are residues that could putatively interact with the cofactor
Mg2?, although at the present not enough experimental
evidence is available to support such a model.
We constructed a complete model of the enzyme toge-
ther with both of the substrates (GTP and Man-1-P) and the
cofactor (Mg2?), all of them positioned in the respectively
proposed binding site (Fig. 2 of supplemental data). As
shown, residues G13, G14, M11, A12 and I93 (orange
sticks) build up a hydrophobic pocket for the purine ring of
GTP, in agreement with that described in other pyrophos-
phorylases (Blankenfeldt et al. 2000; Zuccotti et al. 2001;
Ballicora et al. 2003; Jin et al. 2005). Residues E84 and
N89 (in red sticks) show the right alignment to make H-
bonds to the purine ring particularly with the O6 (bound to
C6) and amine N (bound to C2) (see Fig. 2 of supplemental
data). In green sticks are represented the D116 and S114
residues, which have their side chains in the appropriate
orientation to make H-bonds to the hydroxyl groups of the
ribose moiety. In the model depicted in Fig. 2 of supple-
mental data, the GTP molecule lays deeper inside the cleft
than the sugar, in a way that the phosphate moiety of Man-
1-P is next to the a-phosphate from GTP. This suggests that
the nucleotide would enter first to the cleft and after some
reordering on the surrounding residues the sugar might fit
in its place for taking part in the reaction. This view is in
agreement with the ordered bi-bi sequential kinetic reaction
mechanism proposed for this kind of enzymes (Klecz-
kowski et al. 1993; Zuccotti et al. 2001); where the GTP
molecule would arrive to its binding site before than Man-
1-P, and then the a-phosphate of the former substrate
would produce a nucleophilic attack on Man-1-P giving
PPi and GDP-Man as products (Melo and Glaser 1965;
Kleczkowski et al. 1993; Blankenfeldt et al. 2000). Addi-
tionally, in the 3D model (see Fig. 2 of supplemental data),
the R19 residue is positioned close to Mg2? and the
phosphates moieties of Man-1-P and GTP. This amino acid
has been described as critical partner of the GTP-Mg2? to
keep the full activity of GDP-ManPPases (May et al.
1994).
Given the 3D model and considering that GDP-ManP-
Pase was recognized by specific antibodies present in
typified sera pool from patients with leptospirosis, we
Fig. 5 Superimposition of _1_YP2 onto our 3D model, after a
structural alignment was applied. The residues of both structures were
colored according to their sequence similarity, using BLOSUM 60
substitution matrix. Thick blue cylinders show the most conserved
residues, while thin red cylinders the least ones
Fig. 6 Overlay of consensus region that binds sugar in GDP-
ManPPase model (yellow) onto _1_YP2 (red, potato tuber ADP-
pyrophosphorylase). It should be noted that the side chains are
oriented in a similar way in both structures
Fig. 4 Sequence alignment to the structural alignment depicted in
Fig. 5. Over the sequences is shown a histogram graphic, where the
higher bars indicate the more conserved residues. In a box is
highlighted the sugar binding site residues. _1_YP2 corresponds to
the potato tuber ADP-pyrophosphorylase amino acid sequence
b
Arch Microbiol
123

evaluated the possible structural regions in the protein
having potentiality as epitopes. Based on the predicted
structure of L. interrogans GDP-ManPPase, we identified
conformational (CE) and sequential (SE) epitopes, as
detailed in Table 4. For the epitope recognition, we
employed an algorithm previously developed (Kulkarni-
Kale et al. 2005), which demonstrated a good performance
reaching up to 75% of accuracy when it was applied to
known 3D structures of antigen–antibody complexes. The
specific regions shown in Table 4 are of interest for future
studies oriented to establish a structural insight for the
potential design of new improved vaccines or/and diag-
nostic tests for the leptospiral infection.
Concluding remarks
This work improves the knowledge about the carbohydrate
metabolism in L. interrogans serovar Copenhageni, spe-
cifically about GDP-Man metabolism, a key precursor in
the synthesis of several mannose-containing glycoproteins
and lypopolysaccharides. The gene encoding a GDP-
ManPPase was cloned from genomic DNA; the protein was
heterologously expressed and purified before its functional
characterization. The purified enzyme was identified as a
dimeric protein, exhibiting properties of GDP-ManPPase
with a relative promiscuity to utilize other hexose-1-Ps and
NTPs. The 3D structure of the enzyme was deducted from
molecular modeling on the basis of similarities with other
GDP-ManPPases and also PPases in general. Of particular
interest was that the purified enzyme was quite specifically
recognized in immunoassays by antibodies present in a sera
pool from patients with leptospirosis, which supports a
relevance of this protein as a useful antigen for the diag-
nosis of the disease. Studies on carbohydrate metabolism in
L. interrogans must be continued in depth in order to
determine its relevance both in the virulence of the path-
ogen, the interaction with the host and the identification of
molecular targets for the rational design of new chemo-
therapeutic agents.
Acknowledgments This work was supported by grants from AN-
PCyT (PICTO004 15-22427; PICTO005 05-13469), CONICET (PIP
112-2008-01-02519), and UNL (CAI ? D 2006, CAI ? D 2009
Orientados & Redes). DER, SAG and AAI are investigator career
members from CONICET. AAI is a Fellow from The John Simon
Guggenheim Memorial Foundation.
Table 4 Predicted CE and SE on LinGDP-ManPPase model
CE and SE predicted by the Kulkarni-Kale and col. Algorithm
CE No. AD within 6A of Ref. AD Res. within 6 A of Ref. AD
Predicted CE
1. 1MNQDK5 1 103: ERYGdP :108 R56, R200
2. 13GGKgER18 1333GDiKNLVSELRKNKTLQKYTE353
3. 24RIS26 1 62NASlKKSiLIqEKN75
4. 62NASlKKSiLIqEKN75 1 83IePEGkN89 R42, D55, K79, N80
5. 83IePEGkN89 1 62NASlKKSiLIqEKN75 ? 226KKAGElAA233 ? 236KIiPSEPvDI245
6. 161KSTDA165 1 168AvKSfYEKpDVKTaLK183
7. 168AvKSfYEKpDVKTaLK183 1 161KSTDA165 ? 186KKKN189 ? 236KIiPSEPvDI245 P150, I152, G153, G155, E249
8. 186KKKN189 1 168AvKSfYEKpDVKTaLK183
9. 226KKAGElAA233 1 83IePEGkN89 ? 236KIiPSEPvDI245 K220, E221, F223
10. 236KIiPSEPvDI245 1 83IePEGkN89 ? 168AvKSfYEKpDVKTaLK183 ? 226KKAGElAA233 E249
11. 274RvMPGDELGNrHmGkTiLFHK294 1303RKEFtAvLGVND314 K33
12. 303RKEFtAvLGVND314 1 274RvMPGDELGNrHmGkTiLFHK294 ? 321EDVLfIsTkS330
13. 321EDVLfIsTkS330 ? 303RKEFtAvLGVND314 ? 333GDiKNLVSELRKNKTLQKYTE353
14. 333GDiKNLVSELRKNKTLQKYTE353 ? 13GGKgER18 ? 321EDVLfIsTkS330
Predicted SE
1. 209KTYsP213 E206
2. 36SNK38 K33
3. 137KEH139 R200, R253
4. 120HPVKE124 K127, S260, G262
In capital letters are highlighted the residues with an Accessible Surface Area (ASA) higher than or equal to 30%. In the third column can be
observed the residues which are less than 6 A from the Antigenic Determinant (AD) highlighted in bold. These residues should be taken into
account as a part of the CE. It can also be noticed that some AD could participate in more than one CE, e.g.: AD 161KSTDA165 belong to CE 6
and 7
Arch Microbiol
123

References
Altschul SF et al (1997) Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucleic Acids
Res 25:3389–3402
Arnold K, Bordoli L, Kopp J, Schwede T (2006) The SWISS-
MODEL workspace: a web-based environment for protein
structure homology modelling. Bioinformatics 22:195–201
Ballicora MA, Iglesias AA, Preiss J (2003) ADP-glucose pyrophos-
phorylase, a regulatory enzyme for bacterial glycogen synthesis.
Microbiol Mol Biol Rev 67:213–225 table of contents
Ballicora MA, Iglesias AA, Preiss J (2004) ADP-glucose pyrophos-
phorylase: a regulatory enzyme for plant starch synthesis.
Photosynth Res 79:1–24
Barnett JK et al (1999) Expression and distribution of leptospiral
outer membrane components during renal infection of hamsters.
Infect Immun 67:853–861
Berman HM et al (2000) The protein data bank. Nucleic Acids Res
28:235–242
Bharti AR et al (2003) Leptospirosis: a zoonotic disease of global
importance. Lancet Infect Dis 3:757–771
Blankenfeldt W, Asuncion M, Lam JS, Naismith JH (2000) The
structural basis of the catalytic mechanism and regulation of
glucose-1-phosphate thymidylyltransferase (RmlA). EMBO J
19:6652–6663
Bomfim MR, Ko A, Koury MC (2005) Evaluation of the recombinant
LipL32 in enzyme-linked immunosorbent assay for the serodi-
agnosis of bovine leptospirosis. Vet Microbiol 109:89–94
Boonyod D, Poovorawan Y, Bhattarakosol P, Chirathaworn C (2005)
LipL32, an outer membrane protein of Leptospira, as an antigen
in a dipstick assay for diagnosis of leptospirosis. Asian Pac J
Allergy Immunol 23:133–141
Bowie JU, Luthy R, Eisenberg D (1991) A method to identify protein
sequences that fold into a known three-dimensional structure.
Science 253:164–170
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254
Brede G, Fjaervik E, Valla S (1991) Nucleotide sequence and
expression analysis of the Acetobacter xylinum uridine diphos-
phoglucose pyrophosphorylase gene. J Bacteriol 173:7042–7045
Bulach DM, Kalambaheti T, de la Pena-Moctezuma A, Adler B
(2000a) Functional analysis of genes in the rfb locus of
Leptospira borgpetersenii serovar Hardjo subtype Hardjobovis.
Infect Immun 68:3793–3798
Bulach DM, Kalambaheti T, de la Pena-Moctezuma A, Adler B
(2000b) Lipopolysaccharide biosynthesis in Leptospira. J Mol
Microbiol Biotechnol 2:375–380
Cupp-Vickery JR, Igarashi RY, Meyer CR (2005) Preliminary
crystallographic analysis of ADP-glucose pyrophosphorylase
from Agrobacterium tumefaciens. Acta Crystallogr Sect F Struct
Biol Cryst Commun 61:266–268
Cupp-Vickery JR, Igarashi RY, Perez M, Poland M, Meyer CR
(2008) Structural analysis of ADP-glucose pyrophosphorylase
from the bacterium Agrobacterium tumefaciens. Biochemistry
47:4439–4451
Fusari C, Demonte AM, Figueroa CM, Aleanzi M, Iglesias AA (2006)
A colorimetric method for the assay of ADP-glucose pyrophos-
phorylase. Anal Biochem 352:145–147
Garami A, Ilg T (2001) Disruption of mannose activation in Leishmaniamexicana: GDP-mannose pyrophosphorylase is required for
virulence, but not for viability. EMBO J 20:3657–3666
Ghosh HP, Preiss J (1966) Adenosine diphosphate glucose pyrophos-
phorylase. A regulatory enzyme in the biosynthesis of starch in
spinach leaf chloroplasts. J Biol Chem 241:4491–4504
Ginsburg V (1964) Sugar nucleotides and the synthesis of carbohy-
drates. Adv Enzymol Relat Areas Mol Biol 26:35–88
Gornik O, Dumic J, Flogel M, Lauc G (2006) Glycoscience—a new
frontier in rational drug design. Acta Pharm 56:19–30
Gouet P, Courcelle E, Stuart DI, Metoz F (1999) ESPript: analysis of
multiple sequence alignments in PostScript. Bioinformatics
15:305–308
Griffith ME, Hospenthal DR, Murray CK (2006) Antimicrobial
therapy of leptospirosis. Curr Opin Infect Dis 19:533–537
Griffith ME et al (2007) Efficacy of fluoroquinolones against
Leptospira interrogans in a hamster model. Antimicrob Agents
Chemother 51:2615–2617
Guex N, Peitsch MC (1997) SWISS-MODEL and the Swiss-
PdbViewer: an environment for comparative protein modeling.
Electrophoresis 18:2714–2723
Haake DA et al (1999) Leptospiral outer membrane proteins OmpL1
and LipL41 exhibit synergistic immunoprotection. Infect Immun
67:6572–6582
Haake DA et al (2000) The leptospiral major outer membrane protein
LipL32 is a lipoprotein expressed during mammalian infection.
Infect Immun 68:2276–2285
Jaeckel P, Krauss G, Menge S, Schierhorn A, Rucknagel P, Krauss GJ
(2005) Cadmium induces a novel metallothionein and phyto-
chelatin 2 in an aquatic fungus. Biochem Biophys Res Commun
333:150–155
Jiang XM, Neal B, Santiago F, Lee SJ, Romana LK, Reeves PR
(1991) Structure and sequence of the rfb (O antigen) gene cluster
of Salmonella serovar typhimurium (strain LT2). Mol Microbiol
5:695–713
Jin X, Ballicora MA, Preiss J, Geiger JH (2005) Crystal structure of
potato tuber ADP-glucose pyrophosphorylase. EMBO J 24:694–
704
Kleczkowski LA, Villand P, Preiss J, Olsen OA (1993) Kinetic
mechanism and regulation of ADP-glucose pyrophosphorylase
from barley (Hordeum vulgare) leaves. J Biol Chem 268:6228–
6233
Kobayashi H et al (1997) Structure of a cell wall mannan from the
pathogenic yeast, Candida catenulata: assignment of 1H nuclear
magnetic resonance chemical shifts of the inner alpha-1, 6-linked
mannose residues substituted by a side chain. Arch Biochem
Biophys 341:70–74
Kulkarni-Kale U, Bhosle S, Kolaskar AS (2005) CEP: a conformational
epitope prediction server. Nucleic Acids Res 33:W168–W171
Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680–685
Larsson P, Wallner B, Lindahl E, Elofsson A (2008) Using multiple
templates to improve quality of homology models in automated
homology modeling. Protein Sci 17:990–1002
Luthy R, Bowie JU, Eisenberg D (1992) Assessment of protein
models with three-dimensional profiles. Nature 356:83–85
Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a
laboratory manual. Cold Spring Harbor Laboratory, Cold Spring
Harbor
Martins LO et al (1999) Biosynthesis of mannosylglycerate in the
thermophilic bacterium Rhodothermus marinus. Biochemical
and genetic characterization of a mannosylglycerate synthase.
J Biol Chem 274:35407–35414
May TB, Shinabarger D, Boyd A, Chakrabarty AM (1994) Identi-
fication of amino acid residues involved in the activity of
phosphomannose isomerase-guanosine 50-diphospho-D-mannose
pyrophosphorylase. A bifunctional enzyme in the alginate
biosynthetic pathway of Pseudomonas aeruginosa. J Biol Chem
269:4872–4877
Melo F, Feytmans E (1998) Assessing protein structures with a non-
local atomic interaction energy. J Mol Biol 277:1141–1152
Arch Microbiol
123

Melo A, Glaser L (1965) The nucleotide specificity and feedback
control of thymidine diphosphate D-glucose pyrophosphorylase.
J Biol Chem 240:398–405
Ning B, Elbein AD (1999) Purification and properties of mycobac-
terial GDP-mannose pyrophosphorylase. Arch Biochem Biophys
362:339–345
Ning B, Elbein AD (2000) Cloning, expression and characterization
of the pig liver GDP-mannose pyrophosphorylase. Evidence that
GDP-mannose and GDP-Glc pyrophosphorylases are different
proteins. Eur J Biochem 267:6866–6874
Opperdoes FR, Michels PA (2001) Enzymes of carbohydrate
metabolism as potential drug targets. Int J Parasitol 31:482–490
Preston A, Mandrell RE, Gibson BW, Apicella MA (1996) The
lipooligosaccharides of pathogenic gram-negative bacteria. Crit
Rev Microbiol 22:139–180
Ramachandran GN, Ramakrishnan C, Sasisekharan V (1963) Stereo-
chemistry of polypeptide chain configurations. J Mol Biol 7:95–
99
Russell RB, Barton GJ (1992) Multiple protein sequence alignment
from tertiary structure comparison: assignment of global and
residue confidence levels. Proteins 14:309–323
Saavedra-Lira E, Perez-Montfort R (1996) Energy production in
Entamoeba histolytica: new perspectives in rational drug design.
Arch Med Res 27:257–264
Sali A, Blundell TL (1993) Comparative protein modelling by
satisfaction of spatial restraints. J Mol Biol 234:779–815
Shinabarger D, Berry A, May TB, Rothmel R, Fialho A, Chakrabarty
AM (1991) Purification and characterization of phosphomannose
isomerase-guanosine diphospho-D-mannose pyrophosphorylase.
A bifunctional enzyme in the alginate biosynthetic pathway of
Pseudomonas aeruginosa. J Biol Chem 266:2080–2088
Sugahara MKN (2005) Crystal structure of mannose-1-phosphate
guanyltransferase from Thermus thermophilus Hb8. Riken
Structural GenomicsPROTEOMICS, Initiative (Rsgi)
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22:4673–
4680
Vega MC et al (2005) Regulation of the hetero-octameric ATP
phosphoribosyl transferase complex from Thermotoga maritimaby a tRNA synthetase-like subunit. Mol Microbiol 55:675–686
Veloso IF, Lopes MT, Salas CE, Moreira EC (2000) A comparison of
three DNA extractive procedures with Leptospira for polymerase
chain reaction analysis. Mem Inst Oswaldo Cruz 95:339–343
Verlinde CL et al (2001) Glycolysis as a target for the design of new
anti-trypanosome drugs. Drug Resist Updat 4:50–65
Yang Y-H, Kang Y-B, Lee K-W, Lee T-H, Park S-S, Hwang B-Y,
Kim B-G (2005) Characterization of GDP-mannose pyrophos-
phorylase from Escherichia coli O157:H7 EDL933. J Mol Catal
B: Enzym 37:1–8
Zhang L, Radziejewska-Lebrecht J, Krajewska-Pietrasik D, Toivanen
P, Skurnik M (1997) Molecular and chemical characterization of
the lipopolysaccharide O-antigen and its role in the virulence of
Yersinia enterocolitica serotype O:8. Mol Microbiol 23:63–76
Zuccotti S, Zanardi D, Rosano C, Sturla L, Tonetti M, Bolognesi M
(2001) Kinetic and crystallographic analyses support a sequen-
tial-ordered bi bi catalytic mechanism for Escherichia coliglucose-1-phosphate thymidylyltransferase. J Mol Biol 313:831–
843
Zunino ME, Pizarro PR (2007) Leptospirosis: a literature review. Rev
Chilena Infectol 24:220–226
Arch Microbiol
123









![[Relation between characteristics of the small mammal population and occurrence of Leptospira antibodies]](https://img.pdfslide.net/doc/110x75/635d6736a0f1eac29f0c2c74/relation-between-characteristics-of-the-small-mammal-population-and-occurrence.jpg)



















