Embed Size (px)
Citation preview

3
>Functional expression of CXCR3
in cultured mouse and human
astrocytes and microglia<
K. Biber, I.M. Dijkstra, C.Trebst, C.J.A. De Groot, R.M. Ransohoff, H.W.G.M. Boddeke
Published in Neuroscience, 2002
77> G e n e r a l i n t r o d u c t i o n <

78> G e n e r a l i n t r o d u c t i o n <

AbstractIt has been established recently that inflammation of the central nervous system(CNS) is accompanied by an expression of chemokines within the CNS. Severallines of evidence suggest that chemokines within the CNS initiate and orchestratethe infiltration of the inflamed brain by blood leukocytes. In line with this assump-tion is the finding that glial cells expressing the chemokine ligands CXCL10 andCXCL9 accompany the presence of CXCR3 positive T-cells in the CNS in demyeli-nating diseases.
It is also known that endogenous cells of the CNS express functional chemokinereceptors, raising the possibility that chemokines may be involved in intercellularsignaling between brain cells during brain inflammation. Since it was shown recent-ly that CXCL10, but also CCL21 expression is induced rapidly in damaged neuronsit seemed of interest to investigate whether CXCR3 is expressed in brain cells.Astrocytes and microglia from neonatal mouse brain and adult human brain weregrown in culture and investigated for CXCR3 expression by the means of varioustechniques. RT-PCR experiments and in situ hybridization analysis revealed thatastrocytes and microglia from both mouse and human sources express CXCR3mRNA. Protein expression of CXCR3 in all cell types was detected by immunocy-tochemistry. Moreover, stimulation of cultured glial cells with chemokine ligands forCXCR3 induced intracellular calcium transients and chemotaxis, indicating thefunctional expression of CXCR3.
These results indicate that glial cells in culture functionally express the chemokinereceptor CXCR3. Since damaged neurons rapidly induce the expression ofchemokines that activate CXCR3 it is suggested that glial expression of CXCR3might contribute to an intercellular signaling system in the CNS related to patho-logical conditions.
79> C X C R 3 i n g l i a l c e l l s <

IntroductionChemokines are small chemotactic cytokines of approximately 10 kDa, whichorchestrate the inflammatory response by attracting leukocytes to sites of inflamma-tion and by controlling the homing of dendritic cells, T cells and B cells (for reviewsee: 34,35). Chemokines and their receptors, all of which are G-protein coupled, aresubdivided into four families: CXC-, CC-, C- and CX3C-chemokines 29. Due tomouse and human genome projects and EST databases a large number ofchemokines and chemokine receptors has been found in the last few years 35. Inhumans more than 40 chemokines and 18 chemokine receptors have been identi-fied 29. Beyond their functions in immune and inflammatory reactions, chemokinesmediate a wide variety of different processes throughout the body. It has been shownfor example that chemokines and their receptors are also involved in development,maturation of leukocytes, angiogenesis, metastasis, wound healing and allograftrejection 21,35,36.
Chemokines and their receptors are not only found in the periphery. It has beenshown that chemokines and their receptors are also expressed in the central nervoussystem (CNS) during development and pathology (for review see: 4,5,14,19).Several lines of evidence indicate that chemokine expression in the CNS is primari-ly involved in the infiltration of the CNS by blood leukocytes in response to CNSdiseases 4,28,46. In addition, new findings suggest that neurons and most glial cell typesalso express chemokine receptors. This has raised the suggestion that chemokinesmay contribute to an intercellular signalling system related to pathological condi-tions within the CNS 5,22. Two CXC chemokines, CXCL10 (formerly known as IP-10, interferon-γ inducible protein 10) and CXCL9 (formerly known as MIG,monokine induced by interferon-γ) have been found in brain areas affected byischemic brain damage 44,45, in experimental autoimmune encephalomyelitis (EAE)17,31 and in multiple sclerosis 6,39,40. CXCL9- and CXCL10-positive cells within theCNS are mainly glial cells, whereas the corresponding receptor CXCR3 27 in CNStissue was predominantly found in T-cells 6,39,40.
Recent data indicate that chemokines in the CNS are not only expressed by glialcells. Chemokine ligands for CXCR3 are rapidly expressed by neurons afterischemic brain damage 8,44,45, raising the question on the function of neuronalchemokines. Although in some reports expression of CXCR3 in CNS tissue hasbeen demonstrated, few data are available on the expression of CXCR3 in specificbrain cells. We therefore studied the expression of CXCR3 in cultured glial cells.
80> C X C R 3 i n g l i a l c e l l s <

Experimental procedures
ChemicalsDulbecco’s modified Eagle Medium and HAMF10 was purchased from Gibco BRLLife Technologies (Breda, Netherlands); Percoll from Pharmacia Biotech (Upsalla,Sweden); TA vector pCRII from Invitrogen (Leek, Netherlands); digoxigenin-con-jugated UTP and alkaline phosphatase conjugated sheep-anti-digoxigenin fromBoehringer Mannheim (Mannheim, Germany); biotin-labeled anti-mouse IgGfrom Sigma-Aldrich (Bornhem, Belgium); steptavidin horseradish peroxidase(HRP) from Amersham (Leek, The Netherlands); recombinant mouse and humanchemokines Pepro Tech EC Ltd (London, United Kingdom); antibodies for mouseGFAP from Chemicon (Temencula, USA); human GFAP from Dako (Denmark);human CXCR3 from R&D systems, murine affinity-purified rabbit anti-murineCXCR3 (14920, kindly provided by Dr. Julie DeMartino, Merck ResearchLaboratories, Rahway, NJ, USA). Fura-2 AM and all other chemicals were obtainedfrom Sigma-Aldrich.
Cell cultures
Mixed mouse astrocyte cell cultures and microgliaMixed astrocyte cell cultures were established as described previously 7. Cultureswere maintained up to 4 weeks in DMEM containing 10% fetal calf serum with0,01% penicillin and 0,01% streptomycin in a humidified atmosphere (5% CO2) at37°C. Culture medium was changed the second day after preparation and every 6days thereafter. These cultures contained 70-75% astrocytes (GFAP-positive cells)and 20-30% microglia (F4/80-positive cells); 1-5% of the cells did not stain foreither GFAP or F4/80 and were most likely endothelial cells and/or fibroblasts. Forin situ hybridization and immunocytochemistry mixed astrocyte cultures were incu-bated with 0.25% w/v pocine trypsin at 37°C for 10 min and seeded on poly-L-lysine-coated glass coverslips. Floating microglia were harvested from confluentmixed astrocyte cultures and plated on new culture dishes. Microglia cultures werepure (> 95%) as tested by cell specific markers (F4/80). For calcium measurementscells were seeded on glass coverslips and left in suspension for chemotaxis assay.
Human glial cellsHuman brain tissue was obtained by rapid autopsy according to standardized pro-cedures under the management of the Netherlands Brain Bank (Amsterdam, TheNetherlands; coordinator Dr. R. Ravid). All patients or their next of kin had givenwritten consent for autopsy during life, and for use of their brain tissue for research
81> C X C R 3 i n g l i a l c e l l s <

purposes. Medical records were obtained for all patients prior to autopsy, andpathology evaluation was performed to confirm the clinical diagnosis. Primaryhuman astrocyte cultures were isolated as described previously 13.
Brain tissue samples were taken from the subcortical white matter or corpus cal-losum from normal control patients. The brain tissue samples were collected inDulbecco’s modification of Eagle’s medium (DMEM) with 4500 mg/l glucose andpyridoxine HCl (Gibco BRL Life Technologies, The Netherlands) diluted 1:1 withNutrient mixture F10 (HAMF10, Gibco) with L-glutamine (Gibco) and supple-mented with 50 µg/ml gentamycin (Gibco). Tissue devoid of meninges and bloodvessels was minced into small fragments (< 2 mm3) and digested with a 0.25%trypsin solution (porcine pancreas, Sigma, USA) containing 0.05% DNase (bovinepancreas, Boehringer Mannheim, Germany; 10 ml of this solution was added per 1g wet weight tissue). After digestion, the cell suspensions were gently triturated, andwashed with DMEM/HAMF10 culture medium containing 10% (v/v) fetal bovineserum (FBS, heat inactivated, Integro, The Netherlands), 2 mM L-glutamine, 100IU/ml penicillin and 50 µg/ml streptomycin-sulphate and transferred to 80 cm2 cul-ture flasks (Greiner, The Netherlands) previously coated with poly-L-lysine (PLL; 15µg/ml; Sigma, P6282). Flasks were incubated at 37°C in a humidified atmosphereof 5% CO2 and 95% air for 48 h after which the culture medium was changed toremove unattached cells and myelin debris. Subsequently, the culture medium waschanged once a week with fresh medium and oligoclonal astrocyte cultures weregrown to reach confluency. For passaging, the adherent cells they were rinsed withPBS, incubated for 5-10 min with 0.25% w/v porcine trypsin at 37°C and washedonce with culture medium containing 10% v/v FCS.
To obtain primary microglial cell cultures, human postmortem brain tissue sam-ples were treated as described previously 13. In brief, after digestion of the brain tis-sue samples (see above) cell suspensions were resuspended in culture medium andpassed through a 100 µm nylon mesh filter (Becton & Dickinson, USA). After fil-tering, the cell suspension was centrifuged at 275 X g for 7 min, whereafter the cellpellet was taken up in a final volume of 40 ml of Percoll solution with a density of1.03 g/ml. This Percoll solution is composed of 11.7 ml Percoll (Pharmacia Biotech,Sweden), 1.3 ml 1.5 M NaCl, and 40 ml myelin-gradient buffer. The myelin-gradi-ent buffer consists of sodium phosphatebuffer (composed of a solution of 0.78 g/lNaH2PO4.H2O which was adjusted to pH 7.4 by adding 3.56 g/l Na2HPO4.2H2O)containing 8.0 g/l NaCl, 0.4 g/l KCl, 2.0 g/l glucose (pH 7.4) and 0.2% (w/v; finalconcentration) bovine serum albumine (BSA, Boehringer, The Netherlands). Ontop of the cell suspension in the Percoll solution 6 ml of myelin-gradient buffer wasgently layered. The gradient was centrifuged at 950 X g for 30 min without brakes(acceleration to 950 X g in 120 seconds). Subsequently, the cell debris/myelin layer
82> C X C R 3 i n g l i a l c e l l s <

and interphase were gently aspirated with a 10 ml pipette and the cell pellet waswashed with culture medium at 275 X g for 7 min. For lysis of contaminating ery-throcytes, the pellet was resuspended with 5 ml of a NH4Cl (0.155 M) and KHCO3
(0.001 M) solution, containing 0.5 ml BSA and kept on ice for 15 min. After wash-ing the cells were transferred to 80 cm2 uncoated tissue culture flasks. The flasks wereincubated in a humidified incubator at 37°C pulsed with an atmosphere of 5% CO2
and 95% air. Microglial cells adhere to the bottom of the tissue culture flask within12 h.
To induce proliferation of microglial cells recombinant human granulocyte-macrophage colony stimulating factor (rhGM-CSF; stock solution 300 µg/ml;Leucomax, Sandoz, The Netherlands) was added (final concentration 25 ng/ml) tothe adherent cell cultures every three days. The culture medium was changed oncea week, using fresh culture medium. To passage the adherent cells they were har-vested using a rubber cell scraper (Costar, MA, USA) or by trypsinization with0.25% w/v porcine trypsin at 37°C.
Subcultured human astrocytes were immunoreactive (> 98%) for GFAP (Dako,Denmark) and glutamine synthetase (Affiniti, UK), and did not stain with mono-clonal antibodies LeuM5 (CD11c) and LeuM3 (CD14) (Becton & Dickinson,Belgium), indicating that contamination with macrophage/microglial cells was neg-ligible. Microglial cells had either a bipolar or an amoeboid morphology andimmunostained (> 98%) with LeuM5 (CD11c), LeuM3 (CD14), KP1 (CD68) andFcγRI (CD64) whereas they were negative for GFAP.
Human glial cultures were obtained from the Department of Pathology(University of Amsterdam, Amsterdam, The Netherlands). In contrast to culturedhuman astrocytes, it was not possible to transport cultured human microglia to theDepartment of Medical Physiology (University of Groningen, Groningen, TheNetherlands). Since in situ hybridization and calcium measurements were per-formed at the Department of Medical Physiology, it was not possible to use humanmicroglia in these experiments.
Reverse transcription polymerase chain reaction (RT-PCR)Cells were lysed in guanidinium isothiocyanate/mercaptoethanol buffer and totalRNA was extracted with one phenol-chloroform step, precipitated according toChomczynski and Sacchi (1987) 10. Subsequently RNA was treated with RNAse freeDNAse (Sigma-Aldrich, Bornhem, Belgium) for 2 h and precipitated. Reverse tran-scription (RT): 1 µg of total RNA was transcribed into cDNA as described 7. Thequality of the cDNA was controlled using glyceraldehyde-3-phosphate dehydroge-nase (GAPDH) primers 7 and potential contamination by genomic DNA waschecked for by running the reactions without reverse transcriptase and using
83> C X C R 3 i n g l i a l c e l l s <

GAPDH primers in subsequent polymerase chain reaction (PCR) amplifications.PCR: 2 µl of the RT reaction was used in subsequent PCR amplification asdescribed 7. Primer sequences for mouse CXCR3 according to Soto et al. (1998)(accession number AF045146)41: forward primer: 5’-AACAGCACCTCTCCC-TACGA- 3’; backward primer: 5’-AAGGCCCCTGCATAGAAGTT- 3’. Primersequences for human CXCR3 according to Loetscher et al. (1996) (accession num-ber X95876)27: forward primer: 5’-TGCCAATACAACTTCCCACA- 3’; backwardprimer: 5’-CGGAACTTGACCCCTACA AA-3’. Cycle numbers were 35 andannealing temperature was 58°C for both primer pairs. Identification of all PCRproducts was verified by TA cloning into pCRII (Invitrogen) and subsequentsequencing. In brief PCR products were stored on ice after the amplification. 20 µlof the resulting PCR product was checked by gel electrophoresis and 2 µl was usedfor ligation into linearized pCRII (Invitrogen) according to the instructions of themanufacturer. The ligation product was used to transform competent bacteria(TOP-10F, Invitrogen) and resulting bacterial colonies were grown overnight in 5ml LB medium. Plasmid preparation was done by standard methods and positiveplasmids were sequenced by ALF (sequencing facility; State University Groningen).
In-situ hybridizationFor in situ hybridization pCRII vectors containing mouse or human CXCR3 PCRproducts were linearized with BamHI or XhoI. CXCR3 sense and antisense probeswere synthesized by run off transcription and the use of digoxigenin-conjugatedUTP according to the manufacturer’s protocol (Boehringer Mannheim). Culturedglial cells were seeded on poly-L-lysine-coated glass cover-slides and fixed in 4%paraformaldehyde for 30 min. In situ hybridization procedure was carried out asdescribed earlier and alkaline phosphatase-conjugated sheep anti-digoxigenin wasused for the immunological detection of the digoxigenin-labeled RNA-RNA com-plex 11. Positive cells were counted in randomly chosen areas with a scored eyepiece.Control experiments included hybridization with digoxigenin-labeled sense probesand hybridization after treatment of the sections with RNAse.
ImmunocytochemistryImmunocytochemistry in human cultured glia cells was carried out as described pre-viously 8. All antisera were diluted in phosphate-buffered saline (PBS) containing0.3% Triton X-100, 1% bovine serum albumin (BSA) and heparin (5 mg/ml). Cellswere seeded on poly-L-lysine-coated glass cover-slides, fixed in 4% paraformalde-hyde (30 min) and rinsed in PBS. Cells were preincubated in 5% BSA in Tris-buffered saline (TBS) for 30 min and incubated overnight with mouse anti-humanCXCR3 antibody (R&D Systems). Antibody-antigen reactions were detected using
84> C X C R 3 i n g l i a l c e l l s <

the biotin-streptavidin method (biotin-labeled anti-mouse IgG and streptavidinHRP) and the complex was visualized with 3,3’-diaminobenzidine (DAB)/H2O2.The slides were mounted in DEPEX and analyzed. Control experiments were per-formed in the absence of primary antibody.
For immunocytochemistry cultured mouse microglia cells were seeded on glasscoverslips and fixed in methanol:aceton (50:50) and dried. After rehydration in PBSfor 10 min, slides were blocked with 10% normal human antibody serum for 2 hand incubated with primary rabbit anti-murine CXCR3 antibody (kindly providedby Merck) overnight at 4°C. Antibody-antigen reactions were detected with biotiny-lated secondary antibody (goat anti-rabbit, Vector Laboratories, Burlingame, CA,USA) and Neutralite Avidin-FITC (Southern Biotechnologies, Biotechnologies,Birmingham, USA). After the final washing step slides were mounted in Vectashieldmounting media containing DAPI for nuclear counterstain. Control experimentswere performed in the absence of primary antibody.
Determination of intracellular calciumFor calcium measurements, microglia were cultured on glass coverslips. In order toload the cells with Fura-2 AM the cells were incubated for 15 min at 37°C in load-ing buffer containing: (in mM) NaCl 120, HEPES 5, KCL 6, CaCl2 2, MgCl2 1,glucose 5, NaHCO3 22, Fluo-3 AM 0.005; pH 7.4. The coverslips were fixed in aperfusion chamber (37°C) and mounted on an inverted microscope. Fluorometricmeasurements were performed using a sensicam CCD camera supported by AxolabR
2.1 imaging software. Digital images of the cells were obtained at an emission wave-length of 510 nm using paired exposures to 340 and 380 nm excitation wavelengthsampled at a frequency of 1 Hz. Fluorescence values representing spatial averagesfrom a defined pixel area were recorded on-line. Increases in intracellular calciumconcentrations were expressed as the 340/380 ratio of the emission wavelengths.Compounds were administered using a pipette positioned at a distance of 100-300µm from the cells.
Chemotaxis assayCell migration in response to chemokines was assessed using a 48-well chemotaxismicrochamber (NeuroProbe, Bethesda, MD, USA) as described earlier 8.Chemokine stock solutions (10 µM) were prepared in PBS and further diluted foruse in the assay. Culture medium without chemokines served as a control. Formicroglia and astrocytes 2·104 and 5·104 cells respectively per 50 µl were used in theassay. Determinations were done in quadruplicate. The chamber was incubated at37°C, 5% CO2 in a humidified atmosphere for 120 min in case of microglia and240 min for astrocytes. At the end of incubation the filter was washed, fixed in
85> C X C R 3 i n g l i a l c e l l s <

methanol and stained with Toluidine Blue. Migrated cells were counted with ascored eyepiece (three fields (1 mm2) per well) and migrated cells per chamber werecalculated. The data are presented as mean values ± S.D. and were analyzed byStudent’s t-test. P values ≤ 0.01 were considered significant.
86> C X C R 3 i n g l i a l c e l l s <

Results
Expression of CXCR3 mRNA in cultured glial cellsTo investigate CXCR3 mRNA expression in cultured glial cells, RT-PCR experi-ments were performed. Using specific primer pairs for mouse and human CXCR3specific PCR products of the expected size from astrocytes and microglia of bothmouse and humans were amplified (fig. 3-1). The PCR products were cloned,sequenced and found to be 100% identical with the published sequence for CXCR327,41 thus indicating expression of CXCR3 mRNA in cultured glial cells.
RT-PCR experiments do not allow conclusions concerning the cell type express-ing CXCR3 mRNA, especially not in mixed murine astrocyte cultures. Therefore,in situ hybridization experiments using specific CXCR3 probes were performed.Astrocytes in the cultures were identified by their size and their morphology (see fig.3-2A for the differences between astrocytes and microglia (microglia is indicated byan arrowhead)). Only background staining was found when cells were hybridizedwith the CXCR3 sense probe (fig. 3-2A). Specifc hybridization with DIG-labeledCXCR3 antisense probes was found in approximately 50% (58 out of 126 cells werepositive) of the cultured astrocytes (fig. 3-2B). The arrows show positive cells,dashed arrows indicate negative cells (fig. 3-2B). Due to the size of astrocytes andthe amount of cytoplasm in these cells the typical perinuclear staining for mRNAwas detected. More cells were found to be positive in pure microglial cultures wherealmost all of the microglia stained with the CXCR3 antisense probe (117 out of 125cells) (fig. 3-2D). Again, no or only background staining was found when the cul-tured cells were hybridized with the CXCR3 sense probe (fig. 3-2C).Microphotographs taken at higher magnification clearly show the difference instaining for the CXCR3 sense (fig. 3-2E) and the antisense probe (fig. 3-2F).
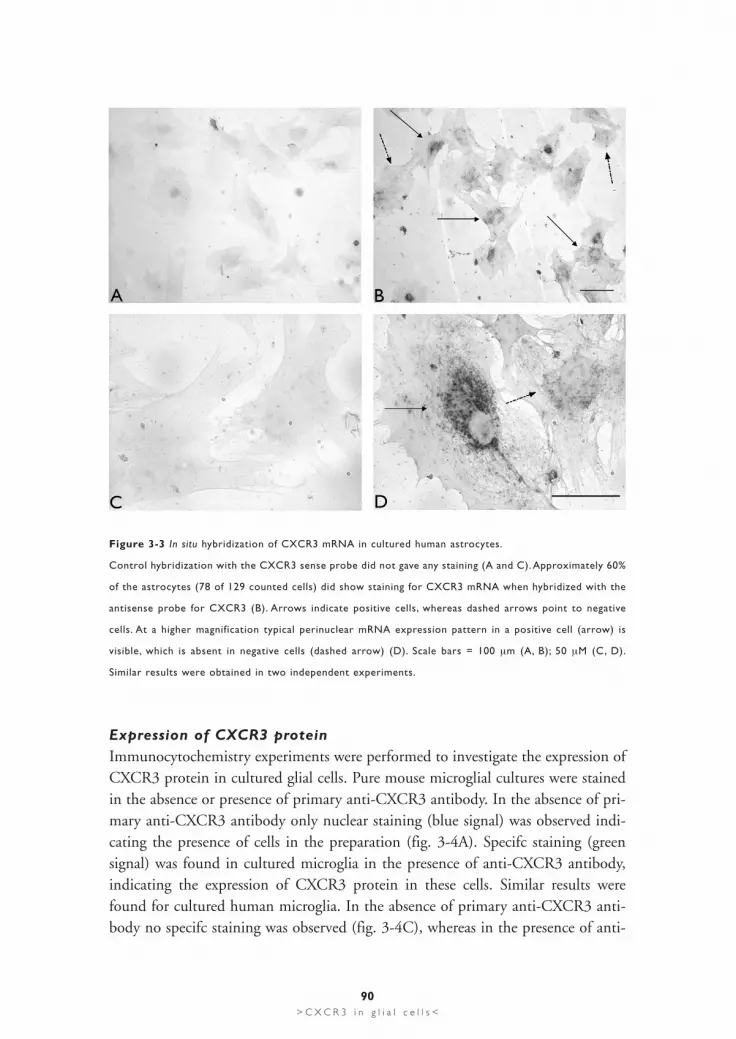
In situ hybridization analysis for human CXCR3 expression in cultured astrocytesgave similar results. No signal was found in experiments where the CXCR3 senseprobe was used (fig. 3-3A). Analysis with the CXCR3 antisense probe showedspecifc staining in approximately 60% of the astrocytes (78 out of 129 cells) (fig.3B). The arrows show positive cells, dashed arrows indicate negative cells (fig. 3B).Microphotographs taken at higher magnifcation revealed the clear differencebetween unspecifc (fig. 3-3C) and specifc staining (fig. 3-3D). Figure 3-3D more-over shows the difference between a positive cell (arrow) and a negative cell (dashedarrow) for CXCR3 mRNA expression.
87> C X C R 3 i n g l i a l c e l l s <

88> C X C R 3 i n g l i a l c e l l s <
Figure 3-1 RT-PCR analysis of CXCR3 mRNA expression in cultured human and mouse astrocytes and
microglia.
Using specific primer pairs CXCR3 mRNA expression was found in both cultured astrocytes (as) and
microglia (mic) derived from either human and mouse brain tissue as shown in the upper pannel. The lower
panel shows the results obtained with GAPDH house-keeping gene primers in order to verify the quality
of the cDNA used in the experiment. Similar results were obtained in 3 independent experiments. Number
of cycles used in PCR was 35 for CXCR3 and 28 for GAPDH; nc: PCR negative control; MM: molecular
weight marker, the highlighted band is 500 bp.

89> C X C R 3 i n g l i a l c e l l s <
Figure 3-2 In situ hybridization analysis of CXCR3 mRNA expression in cultured mouse glial cells.
A) Control hybridization with the CXCR3 sense probe in cultured astrocytes only resulted in background
staining. This panel moreover shows the difference in size and morphology between astrocytes and
microglia (arrowhead). B) Hybridization with the CXCR3 antisense probe showed CXCR3 mRNA expres-
sion in approximately 50% (58 of 126 counted cells were positive) cultured mouse astrocytes. Note the typ-
ical perinuclear mRNA expression pattern in positive astrocytes (arrows), whereas no staining was
detectable in negative cells (dashed arrows). Background staining was detected with the sense probe in cul-
tured microglia (C and E). Expression of CXCR3 mRNA in cultured microglia was found by hybridization
with the antisense probe for CXCR3 (D and F). Higher magnification clearly shows the difference between
sense (E) and antisense probes (F) in microglia. Similar results were obtained in three independent exper-
iments. Scale bars = 100 µm (A, B); 50 µm (C, D); 50 µm (E, F).
Expression of CXCR3 proteinImmunocytochemistry experiments were performed to investigate the expression ofCXCR3 protein in cultured glial cells. Pure mouse microglial cultures were stainedin the absence or presence of primary anti-CXCR3 antibody. In the absence of pri-mary anti-CXCR3 antibody only nuclear staining (blue signal) was observed indi-cating the presence of cells in the preparation (fig. 3-4A). Specifc staining (green signal) was found in cultured microglia in the presence of anti-CXCR3 antibody,indicating the expression of CXCR3 protein in these cells. Similar results werefound for cultured human microglia. In the absence of primary anti-CXCR3 anti-body no specifc staining was observed (fig. 3-4C), whereas in the presence of anti-
90> C X C R 3 i n g l i a l c e l l s <
Figure 3-3 In situ hybridization of CXCR3 mRNA in cultured human astrocytes.
Control hybridization with the CXCR3 sense probe did not gave any staining (A and C). Approximately 60%
of the astrocytes (78 of 129 counted cells) did show staining for CXCR3 mRNA when hybridized with the
antisense probe for CXCR3 (B). Arrows indicate positive cells, whereas dashed arrows point to negative
cells. At a higher magnification typical perinuclear mRNA expression pattern in a positive cell (arrow) is
visible, which is absent in negative cells (dashed arrow) (D). Scale bars = 100 µm (A, B); 50 µM (C, D).
Similar results were obtained in two independent experiments.

91> C X C R 3 i n g l i a l c e l l s <
Figure 3-4 CXCR3 protein expression in cultured glial cells.
Control immunocytochemistry experiments in mouse microglia without primary antibody yielded no stain-
ing: DAPI nuclear staining (blue signal) reveals the presence of cells in the preparation (A).
Immunocytochemistry with specific antibody for mouse CXCR3 revealed specific staining (green signal) in
mouse microglia (B). Similar results were also found for human microglia (C and D) and astrocytes (E and
F). Control stainings without primary antibody gave only background staining (C and E). Experiments with
primary antibody for CXCR3 indicated the expression of CXCR3 in microglia (D) and in astrocytes (F). A
partial staining for CXCR3 expression was found in 64 out of 132 counted cells (dashed arrow), whereas
61 out of 132 cells showed CXCR3 expression throughout the cell (arrow). Scale bars = 50 µm (A-D and
E-F). Similar results were obtained in two independent experiments.

CXCR3 antibody specifc staining was found in these cells (fig. 3-4D). A significantamount of the staining for CXCR3 in cultured human microglia was found intra-cellular (fig. 3-4D). This staining pattern was not influenced by other fixation tech-niques or by further dilution of the primary antibody (data not shown).
Similar experiments were also performed with cultured human astrocytes. In theabsence of primary anti-CXCR3 antibody no specific staining was found (fig. 3-4E),whereas in the presence of anti-CXCR3 antibody the expression of CXCR3 proteinwas indicated by specific staining (fig. 3-4F). CXCR3 protein was detected in morethan 95% of the cells, but approximately 50% of the cells (64 out of 132) onlyshowed partial, intracellular staining round the nuclei of the cells (fig. 3-4F) (indi-cated by a dashed arrow). Complete staining for CXCR3 was found in 61 out of 132cells as indicated in fig. 3-4F (indicated by an arrow). Different fixation techniquesor further dilution of the primary antibody did not influence the staining pattern(data not shown).
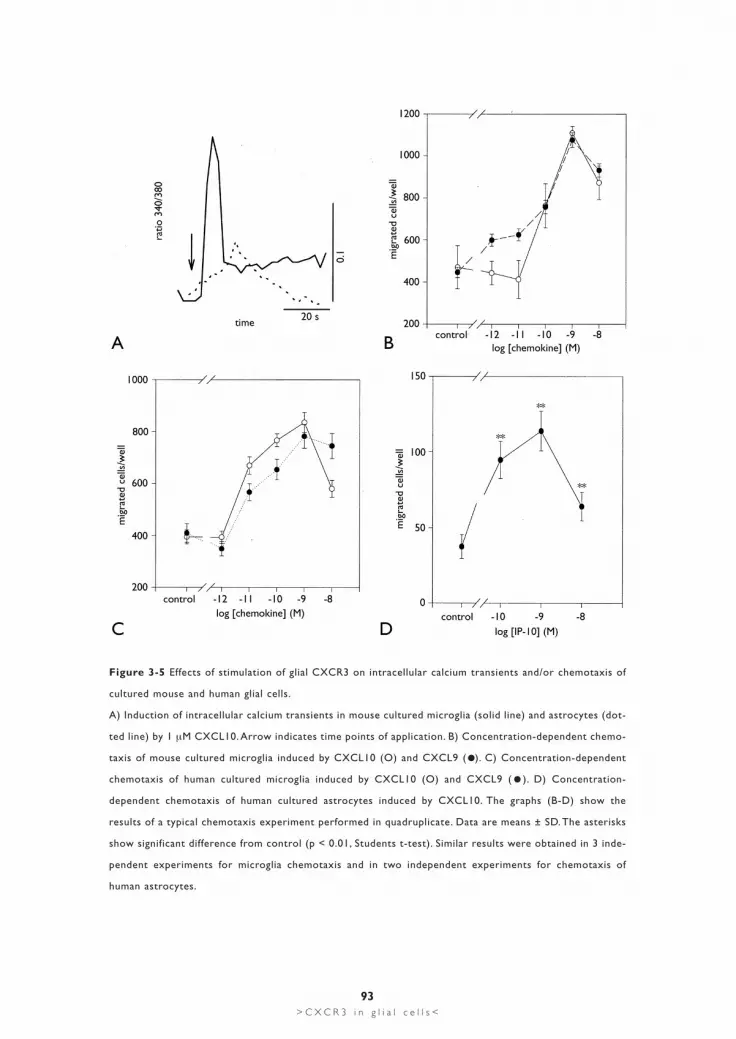
Effects of stimulation of CXCR3 on calcium signaling and chemotaxis of cultured glial cellsStimulation of cultured mouse microglia with 1 µM CXCL10 induced intracellularcalcium transients (fig. 3-5A) in approximately 50% of the cells investigated (51positive cells out of 96). Smaller increases in intracellular calcium were found using100 nM of CXCL10, whereas higher concentrations (5 µM) induced oscillatory cal-cium responses in cultured mouse microglia data not shown). Similar experimentsperformed with cultured mouse astrocytes revealed in approximately 10% of thecells (17 positive cells out of 146) small increases in intracellular calcium when stim-ulated with 1 µM CXCL10 (fig. 3-5A). No increases in intracellular calcium havebeen observed in cultured astrocytes with lower CXCL10 concentrations (100 nM).For technical reasons it was not possible to perform calcium experiments with cul-tured human microglia (see Experimental procedures).
Stimulation of CXCR3 induced chemotaxis of cultured mouse microglia. Figure3-5B shows the CXCL10- induced concentration-dependent chemotactic effect incultured microglia, with a peak effect at 1 nM CXCL10. Similar results wereobtained in experiments with another CXCR3 ligand: CXCL9 (fig. 3-5B).Furthermore, CXCL10- and CXCL9-induced chemotaxis in mouse microglia wasquantitatively comparable to chemotaxis induced by other chemokines like CCL2and CCL3 (data not shown). We detected ≥ 20% microglia in mixed murine astro-cytic cultures. It was therefore not possible to discriminate between astrocytes andmicroglia in the chemotaxis assay, which made chemotaxis experiments with cul-tured murine astrocytes too difficult to interpret.Human microglia migrated in the chemotaxis chamber in response to both CXCR3
92> C X C R 3 i n g l i a l c e l l s <
93> C X C R 3 i n g l i a l c e l l s <
Figure 3-5 Effects of stimulation of glial CXCR3 on intracellular calcium transients and/or chemotaxis of
cultured mouse and human glial cells.
A) Induction of intracellular calcium transients in mouse cultured microglia (solid line) and astrocytes (dot-
ted line) by 1 µM CXCL10. Arrow indicates time points of application. B) Concentration-dependent chemo-
taxis of mouse cultured microglia induced by CXCL10 (O) and CXCL9 ( ). C) Concentration-dependent
chemotaxis of human cultured microglia induced by CXCL10 (O) and CXCL9 ( ). D) Concentration-
dependent chemotaxis of human cultured astrocytes induced by CXCL10. The graphs (B-D) show the
results of a typical chemotaxis experiment performed in quadruplicate. Data are means ± SD. The asterisks
show significant difference from control (p < 0.01, Students t-test). Similar results were obtained in 3 inde-
pendent experiments for microglia chemotaxis and in two independent experiments for chemotaxis of
human astrocytes.

ligands CXCL10 and CXCL9 (fig. 3-5C). Similar concentration dependencies forCXCL10 and CXCL9 were detected when compared to murine microglia. In par-ticular, both chemokines induced maximum migration at 1 nM (fig. 3-5C). Due tothe purity of cultured human astrocytes it was possible to perform chemotaxis exper-iments with these cells. Although the number of migrated astrocytes was rather lowwhen compared to cultured microglia, we detected significant migratory responseswhen cells were stimulated with 0.1-10 nM of CXCL10 (fig. 3-5D). Maximummigration was detected at a concentration of 1 nM CXCL10.
94> C X C R 3 i n g l i a l c e l l s <

Discussion
Expression of CXCR3 in glial cells Demyelinating autoimmune diseases of the CNS like MS or experimental allergicencephalitis (EAE) are characterized by a migration of CXCR3 positive Th1 cellsinto the CNS. Accordingly, glial cells (astrocytes and microglia) positive for thechemokines CXCL10 and CXCL9, have been found in post-mortem CNS tissue ofMS patients and after induction of EAE 6,17,31,39,40.
It is, however, well known that endogenous CNS cells (in vitro and in vivo) alsoexpress a variety of chemokine receptors 9,15,25. It was indicated in a recent report thatneurons express chemokine receptor CXCR3 47. Moreover, there are a few publishedexperiments suggesting the presence of CXCR3 in cultured microglia and glial celllines 8,12,44. We have investigated the expression of CXCR3 in primary cultured glialcells from both mouse and humans. The results presented here show that microgliaand astrocytes from murine and human sources express CXCR3 mRNA and pro-tein. Stimulation of CXCR3 induced chemotaxis and induced intracellular calciumtransients in these cells, which are typical responses after chemokine receptor stim-ulation. It is therefore suggested that CXCR3 is, at least in vitro, a functionalchemokine receptor expressed by two different types of glial cells, astrocytes andmicroglia. It is not possible to obtain a sufficient number of microglia from post-mortem tissue without the presence of GM-CSF in the culture medium 13. Whetherthe presence of GM-CSF in the culture medium might influence the expression ofCXCR3 in cultured human microglia is not known. However, the similar concen-tration response curves and the comparable number of cells migrated in the chemo-taxis assays in cultured microglia from mouse and humans do not indicate a signif-icant influence of GM-CSF on microglial CXCR3 expression. The functionalresponses (chemotaxis and intracellular calcium signals) observed in microglia afterstimulation of CXCR3 were quantitatively different from the rather weak and rareresponses observed in astrocytes. The difference in chemotaxis might be explainedby the different nature of astrocytes and microglia. Microglia are most likely bonemarrow-derived, monocytic cells that have a high migratory capacity. Culturedastrocytes on the other hand are known to have a rather low migratory capacity 20,30.The difference in chemotactic activity between astrocytes and microglia could alsobe explained by a low, insufficient number of CXCR3 at the surface of the astrocytes(see below). It has been shown for other types of chemokine receptors that intracel-lular calcium signals are only detectable when pronounced receptor numbers areexpressed 1, different receptor numbers could therefore explain the differences in cal-cium signals between astrocytes and microglia. Thus, the level of CXCR3 expressionin astrocytes may be insufficient to enable a robust coupling of CXCR3 to the intra-
95> C X C R 3 i n g l i a l c e l l s <

cellular calcium signaling system. This assumption is corroborated by the findingsthat pronounced CXCR3 expression (both mRNA and protein) was observed lessfrequently in astrocytes than in microglia. Moreover, most staining for CXCR3 pro-tein was found at intracellular structures, particularly astrocytes showed this stain-ing pattern. Intracellular expression of chemokine receptors in astrocytes and othercell types in the developing brain in vitro and in vivo, has recently been shown forseveral chemokine receptors 33,43. Nothing is yet known about the regulation ofchemokine receptor expression on the surface of glial cells. It is therefore only pos-sible to speculate that intracellular expression of chemokine receptors in these cellsis part of the normal transport of the receptors to the surface (see here also 33,43).Few data are published concerning the expression of CXCR3 in glial cells in vivo.Basal CXCR3 mRNA expression level in control rat brain was detected by Wang etal. (2000) using RT-PCR analysis 45. Moreover, an induction of CXCR3 mRNAexpression upon ischemic brain damage has been reported 45. Comparable expres-sion of CXCR3 has been observed in ischemic mouse brain in our laboratory 8.These results thus indicate that CXCR3 mRNA is expressed in brain. Whether glialcells are the cellular source of CXCR3 mRNA expression in vivo still needs to be elu-cidated. Antibody staining showed CXCR3-positive cells that had the morphologyof reactive astrocytes in multiple sclerosis lesions 39. Furthermore, glial expression ofCXCR3 has been observed during human brain development 43. Van der Meer et al.(2001), moreover, have reported expression of CXCR3 in microglia in vivo, but incontrast to the results presented here for cultured microglia less than 1% ofmicroglia in vivo showed CXCR3 expression 43. However, recent data obtained inour group showed that microglia in freshly prepared mouse brain slices react onCXCR3 activation with physiological responses, thus indicating microglial CXCR3expression in in vivo like preparations 32.
Possible function of CXCR3 and its chemokine ligands in CNSSeveral lines of evidence suggest that the expressions of CNS chemokines mediatethe infiltration of the brain by leukocytes 4,28. The first indications came from a studyshowing that treatment with anti-CCL3 antibodies prolonged the onset of EAE 26.Recent publications have shown that mice deficient for CCR2 or CCL2 haveimpaired monocyte recruitment in response to peripheral nervous system injury 38
and are resistant to EAE 16,23,24. The existence of CXCR3-positive T-cells in demyeli-nating CNS diseases therefore led subsequently to the assumption that CXCL10and CXCL9 are CNS attractors for Th1-cells 6,39,40. Recently found functions ofchemokines in development, maturation of leukocytes, angiogenesis, metastasis,wound healing and allograft rejection indicate a broad spectrum of chemokinergicactions far beyond a simple attraction of blood leukocytes 21,35.
96> C X C R 3 i n g l i a l c e l l s <

These findings and the presence of chemokine receptors in endogenous brain cells(neurons and glial cells) imply a functional role of chemokines in an intercellularcommunication system within the brain 5,22. The activation of glial cells is one of thefirst events occurring after neuronal injury. Therefore, the existence of a neuronal-glial communication has been proposed by several groups 2,3,42. Chemokines may sig-nificantly contribute to this communication since induction of several chemokines(CCL2, CCL3 and CXCL10) has been described in injured neurons 18,37,45,48.Recently, it has been reported that another chemokine, CCL21 (formerly known asSLC, exodus-2, TCA4 and 6Ckine), is expressed in neurons during brain ischemia8. Induction of CCL21 expression occurred within 6 h after the ischemic insult andtherefore in a similar time-window as it has been reported for expression of neuronalCXCL10 in ischemic rats 45. Moreover, cross-desensitization studies indicated thatCXCL10 and CCL21 would activate the same receptor (CXCR3) on mousemicroglia 8. It is therefore tempting to speculate that CXCR3 expressed on glial cellsmight be the responsible receptor to detect CXCL10 and CCL21 under pathologi-cal conditions.
In summary, we have provided evidence that cultured mouse and human glial cells(astrocytes and microglia) express functional CXCR3 receptors. Chemokine ligandsfor CXCR3 (CXCL10, CXCL9 and CCL21) are highly expressed in ischemicand/or demyelinating CNS diseases. Since the CXCR3-activating chemokine lig-ands CXCL10 and CCL21 are also expressed in neurons under neurodegenerativeconditions our data raise the possibility that glial CXCR3 expression might con-tribute to the communication system between damaged neurons and glial cells.
AcknowledgementsThe authors would like to thank the Netherlands Brain Bank for supplying humanCNS tissue. We wish to thank Lizette Montagne for excellent technical assistance.This research was supported in part by the United States National Institutes ofHealth (2RO1NS32151 and 1PO1NS38667 to RMR).
97> C X C R 3 i n g l i a l c e l l s <

References
1. Albright AV, Shieh JT, Itoh T, Lee B, Pleasure D, O'Connor MJ, Doms RW, Gonzalez-Scarano F.
Microglia express CCR5, CXCR4, and CCR3, but of these, CCR5 is the principal coreceptor for
human immunodeficiency virus type 1 dementia isolates. J Virol 1999; 73:205-213.
2. Aldskogius H, Kozlova EN. Central neuron-glial and glial-glial interactions following axon injury. Prog
Neurobiol 1998; 55:1-26.
3. Aschner M, Allen JW, Kimelberg HK, LoPachin RM, Streit WJ. Glial cells in neurotoxicity development.
Annu Rev Pharmacol Toxicol 1999; 39:151-73.
4. Asensio VC, Campbell IL. Chemokines in the CNS: plurifunctional mediators in diverse states. Trends
Neurosci 1999; 22:504-512.
5. Bacon KB, Harrison JK. Chemokines and their receptors in neurobiology: perspectives in physiology
and homeostasis. J Neuroimmunol 2000; 104:92-7.
6. Balashov KE, Rottman JB, Weiner HL, Hancock WW. CCR5(+) and CXCR3(+) T cells are increased in
multiple sclerosis and their ligands MIP-1alpha and IP-10 are expressed in demyelinating brain
lesions. Proc Natl Acad Sci U S A 1999; 96:6873-8.
7. Biber K, Klotz KN, Berger M, Gebicke-Harter PJ, van Calker D. Adenosine A1 receptor-mediated
activation of phospholipase C in cultured astrocytes depends on the level of receptor expression. J
Neurosci 1997; 17:4956-64.
8. Biber K, Sauter A, Brouwer N, Copray SC, Boddeke HW. Ischemia-induced neuronal expression of the
microglia attracting chemokine Secondary Lymphoid-tissue Chemokine (SLC). Glia 2001; 34:121-133.
9. Boddeke EW, Meigel I, Frentzel S, Biber K, Renn LQ, Gebicke-Harter P. Functional expression of the
fractalkine (CX3C) receptor and its regulation by lipopolysaccharide in rat microglia. Eur J Pharmacol
1999; 374:309-13.
10. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-
phenol-chloroform extraction. Anal Biochem 1987; 162:156-159.
11. Copray JC, Brouwer N. Selective expression of neurotrophin-3 messenger RNA in muscle spindles of
the rat. Neuroscience 1994; 63:1125-1135.
12. Cross AK, Woodroofe MN. Chemokines induce migration and changes in actin polymerization in
adult rat brain microglia and a human fetal microglial cell line in vitro. J Neurosci Res 1999; 55:17-23.
13. De Groot CJ, Montagne L, Janssen I, Ravid R, Van d, V, Veerhuis R. Isolation and characterization of
adult microglial cells and oligodendrocytes derived from postmortem human brain tissue. Brain Res
Brain Res Protoc 2000; 5:85-94.
14. De Groot CJ, Woodroofe MN. The role of chemokines and chemokine receptors in CNS
inflammation. Prog Brain Res 2001; 132:533-544.
15. Dorf ME, Berman MA, Tanabe S, Heesen M, Luo Y. Astrocytes express functional chemokine receptors.
J Neuroimmunol 2000; 111:109-21.
16. Fife BT, Huffnagle GB, Kuziel WA, Karpus WJ. CC chemokine receptor 2 is critical for induction of
experimental autoimmune encephalomyelitis. J Exp Med 2000; 192:899-905.
98> C X C R 3 i n g l i a l c e l l s <

17. Fischer FR, Santambrogio L, Luo Y, Berman MA, Hancock WW, Dorf ME. Modulation of experimental
autoimmune encephalomyelitis: effect of altered peptide ligand on chemokine and chemokine
receptor expression. J Neuroimmunol 2000; 110:195-208.
18. Flugel A, Hager G, Horvat A, Spitzer C, Singer GM, Graeber MB, Kreutzberg GW, Schwaiger FW.
Neuronal MCP-1 expression in response to remote nerve injury. J Cereb Blood Flow Metab 2001;
21:69-76.
19. Glabinski AR, Ransohoff RM. Chemokines and chemokine receptors in CNS pathology. J Neurovirol
1999; 5:3-12.
20. Han Y, Wang J, Zhou Z, Ransohoff RM. TGFbeta1 selectively up-regulates CCR1 expression in primary
murine astrocytes. Glia 2000; 30:1-10.
21. Hancock WW, Lu B, Gao W, Csizmadia V, Faia K, King JA, Smiley ST, Ling M, Gerard NP, Gerard C.
Requirement of the chemokine receptor CXCR3 for acute allograft rejection. J Exp Med 2000;
192:1515-1520.
22. Hesselgesser J, Horuk R. Chemokine and chemokine receptor expression in the central nervous
system. J Neurovirol 1999; 5:13-26.
23. Huang DR, Wang J, Kivisakk P, Rollins BJ, Ransohoff RM. Absence of monocyte chemoattractant
protein 1 in mice leads to decreased local macrophage recruitment and antigen-specific T helper cell
type 1 immune response in experimental autoimmune encephalomyelitis. J Exp Med 2001; 193:713-26.
24. Izikson L, Klein RS, Charo IF, Weiner HL, Luster AD. Resistance to experimental autoimmune
encephalomyelitis in mice lacking the CC chemokine receptor (CCR)2. J Exp Med 2000; 192:1075-80.
25. Jiang Y, Salafranca MN, Adhikari S, Xia Y, Feng L, Sonntag MK, deFiebre CM, Pennell NA, Streit WJ,
Harrison JK. Chemokine receptor expression in cultured glia and rat experimental allergic
encephalomyelitis. J Neuroimmunol 1998; 86:1-12.
26. Karpus WJ, Lukacs NW, McRae BL, Strieter RM, Kunkel SL, Miller SD. An important role for the
chemokine macrophage inflammatory protein-1 alpha in the pathogenesis of the T cell-mediated
autoimmune disease, experimental autoimmune encephalomyelitis. J Immunol 1995; 155:5003-5010.
27. Loetscher M, Gerber B, Loetscher P, Jones SA, Piali L, Clark-Lewis I, Baggiolini M, Moser B.
Chemokine receptor specific for IP10 and mig: structure, function, and expression in activated
T-lymphocytes. J Exp Med 1996; 184:963-969.
28. Mennicken F, Maki R, de Souza EB, Quirion R. Chemokines and chemokine receptors in the CNS: a
possible role in neuroinflammation and patterning. Trends Pharmacol Sci 1999; 20:73-8.
29. Murphy PM, Baggiolini M, Charo IF, Hebert CA, Horuk R, Matsushima K, Miller LH, Oppenheim JJ,
Power CA. International union of pharmacology. XXII. Nomenclature for chemokine receptors.
Pharmacol Rev 2000; 52:145-76.
30. Peterson PK, Hu S, Salak-Johnson J, Molitor TW, Chao CC. Differential production of and migratory
response to beta chemokines by human microglia and astrocytes. J Infect Dis 1997; 175:478-481.
31. Ransohoff RM, Hamilton TA, Tani M, Stoler MH, Shick HE, Major JA, Estes ML, Thomas DM, Tuohy VK.
Astrocyte expression of mRNA encoding cytokines IP-10 and JE/MCP-1 in experimental autoimmune
encephalomyelitis. FASEB J 1993; 7:592-600.
99> C X C R 3 i n g l i a l c e l l s <

32. Rappert A, Biber K, Nolte C, Lipp M, Schubel A, Lu B, Gerard NP, Gerard C, Boddeke HW,
Kettenmann H. Secondary lymphoid tissue chemokine (CCL21) activates CXCR3 to trigger a Cl-
current and chemotaxis in murine microglia. J Immunol 2002; 168:3221-6.
33. Rezaie P, Trillo-Pazos G, Everall IP, Male DK. Expression of beta-chemokines and chemokine receptors
in human fetal astrocyte and microglial co-cultures: potential role of chemokines in the developing
CNS. Glia 2002; 37:64-75.
34. Rollins BJ. Chemokines. Blood 1997; 90:909-28.
35. Rossi D, Zlotnik A. The biology of chemokines and their receptors. Annu Rev Immunol 2000;
18:217-242.
36. Sallusto F, Lenig D, Mackay CR, Lanzavecchia A. Flexible programs of chemokine receptor expression
on human polarized T helper 1 and 2 lymphocytes. J Exp Med 1998; 187:875-883.
37. Schreiber RC, Krivacic K, Kirby B, Vaccariello SA, Wei T, Ransohoff RM, Zigmond RE. Monocyte
chemoattractant protein (MCP)-1 is rapidly expressed by sympathetic ganglion neurons following
axonal injury. Neuroreport 2001; 12:601-606.
38. Siebert H, Sachse A, Kuziel WA, Maeda N, Bruck W. The chemokine receptor CCR2 is involved in
macrophage recruitment to the injured peripheral nervous system. J Neuroimmunol 2000;
110:177-85.
39. Simpson JE, Newcombe J, Cuzner ML, Woodroofe MN. Expression of the interferon-gamma-inducible
chemokines IP-10 and Mig and their receptor, CXCR3, in multiple sclerosis lesions. Neuropathol Appl
Neurobiol 2000; 26:133-142.
40. Sorensen TL, Tani M, Jensen J, Pierce V, Lucchinetti C, Folcik VA, Qin S, Rottman J, Sellebjerg F, Strieter
RM, Frederiksen JL, Ransohoff RM. Expression of specific chemokines and chemokine receptors in the
central nervous system of multiple sclerosis patients. J Clin Invest 1999; 103:807-15.
41. Soto H, Wang W, Strieter RM, Copeland NG, Gilbert DJ, Jenkins NA, Hedrick J, Zlotnik A. The CC
chemokine 6Ckine binds the CXC chemokine receptor CXCR3. Proc Natl Acad Sci U S A 1998;
95:8205-10.
42. Streit WJ, Walter SA, Pennell NA. Reactive microgliosis. Prog Neurobiol 1999; 57:563-81.
43. van der Meer P, Goldberg SH, Fung KM, Sharer LR, Gonzalez-Scarano F, Lavi E. Expression pattern of
CXCR3, CXCR4, and CCR3 chemokine receptors in the developing human brain. J Neuropathol Exp
Neurol 2001; 60:25-32.
44. Wang X, Ellison JA, Siren AL, Lysko PG, Yue TL, Barone FC, Shatzman A, Feuerstein GZ. Prolonged
expression of interferon-inducible protein-10 in ischemic cortex after permanent occlusion of the
middle cerebral artery in rat. J Neurochem 1998; 71:1194-1204.
45. Wang X, Li X, Schmidt DB, Foley JJ, Barone FC, Ames RS, Sarau HM. Identification and molecular
characterization of rat CXCR3: receptor expression and interferon-inducible protein-10 binding are
increased in focal stroke. Mol Pharmacol 2000; 57:1190-8.
46. Wu DT, Woodman SE, Weiss JM, McManus CM, D'Aversa TG, Hesselgesser J, Major EO, Nath A,
Berman JW. Mechanisms of leukocyte trafficking into the CNS. J Neurovirol 2000; 6 Suppl 1:S82-S85.
47. Xia MQ, Bacskai BJ, Knowles RB, Qin SX, Hyman BT. Expression of the chemokine receptor CXCR3
100> C X C R 3 i n g l i a l c e l l s <

on neurons and the elevated expression of its ligand IP-10 in reactive astrocytes: in vitro ERK1/2
activation and role in Alzheimer's disease. J Neuroimmunol 2000; 108:227-35.
48. Xia MQ, Qin SX, Wu LJ, Mackay CR, Hyman BT. Immunohistochemical study of the beta-chemokine
receptors CCR3 and CCR5 and their ligands in normal and Alzheimer's disease brains. Am J Pathol
1998; 153:31-37.
101> C X C R 3 i n g l i a l c e l l s <

102> G e n e r a l i n t r o d u c t i o n <





























