Embed Size (px)
Citation preview

Functional G-CSF pathways in t(8;21) leukemic cells allow fordi�erentiation induction and degradation of AML1-ETO
Nicolas Da Silva1,4, Sandrine Meyer-Monard1,4, Marie-Laurence Menot1, Antonio Parrado1, Annie Lebel1,Nicole Balitrand1, Pierre Fenaux3, Jean-Michel Micle a1,2, Philippe Rousselot1,2, Laurent Degos2,Herve Dombret2 and Christine Chomienne*,1
1LBCH (Laboratoire de Biologie Cellulaire HeÂmatopoõÈeÂtique, INSERM E 00-03, et EA 316 Universite Paris 7, HoÃpitalSaint-Louis, Paris, France; 2Service Clinique des Maladies du Sang, HoÃpital Saint-Louis, Paris, France; 3CHU de Lille,France
Introduction: E�cacy of di�erentiating agents requires that their speci®c cellular targets arestill expressed and functional in the leukemic cells. One hypothesis to target sensitive cells isto select leukemic clones which harbor disrupted transcription factors. CBFa and CBFb arecore-binding proteins which have been identi®ed as transcription regulators of hematopoieticgenes and shown to be altered in numerous leukemias. In M2 AML, the t(8;21) translocation,CBFa (AML1) is altered and produced as the AML1-ETO fusion protein. The fusion proteinblocks transcription and di�erentiation mediated by G-CSF. Interestingly, AML1-ETOleukemic cell lines are sensitive to numerous cytokines in vitro and can be induced todi�erentiate in the presence of G-CSF and PMA.Materials and methods: As in the APL di�erentiation model, primary culture provides auseful tool for therapeutic screening of di�erentiation inducers, we analysed the in vitrosensitivity of 10 fresh M2 AML t(8;21) leukemic samples to G-CSF and the functionality ofG-CSF intracellular pathways. In vitro data were compared with in vivo data from fourpatients treated with rhG-CSF at the dosage of 5 mg/kg/day i.v. for two to three weeks beforethe initiation of AML induction chemotherapy and immunophenotypic analysis performedweekly to monitor in vivo di�erentiation.Results: In vitro, an increase in CD34+ cells expressing di�erentiation antigens (CD11b,CD13 or CD15) was noted along with a decrease of immature CD34+/di�erentiation antigennegative cells. After two weeks of a daily rhG-CSF administration in vivo, a signi®cant, albeittransient, decrease of blast count was achieved, concomitant with an increase in di�erentiatedleukemic cells suggesting that in vivo di�erentiation occurs. Fresh t(8;21) leukemic cellspossess functional G-CSF signaling pathways as normal activity and kinetics of STAT1 andSTAT3 binding was observed. Furthermore, di�erentiation induction leads to a subsequentdegradation of the AML1-ETO oncoprotein.Conclusion: The data presented here supports the claim that G-CSF can induce in vitro andin vivo di�erentiation of M2 AML t(8;21) cells.The Hematology Journal (2000) 1, 316 ± 328
Keywords: leukemia di�erentiation; AML1-ETO; G-CSF; therapy
Introduction
Combination of di�erentiation therapy with cytotoxicchemotherapy is strikingly e�ective in acute promyelo-cytic leukemia (APL) to eradicate the leukemic clone.1
However, few di�erentiating agents are known to bee�ective on other leukemic subtypes in primary cultures.2
The APL-di�erentiation model raises the hypothesisthat the e�cacy of a given agent in a leukemic cell maybe related to the stage of di�erentiation arrest and tothe accessibility of the di�erentiating agent's targetgenes. APL is identi®ed by the reciprocal translocationt(15;17) resulting in the PML-RARa fusion proteinwhich blocks retinoic acid (RA)-induced granulocyticdi�erentiation.3 Induction of di�erentiation by RA in
*Correspondence: C Chomienne, Hoà pital St Louis, 1 Avenue ClaudeVellefaux, 75754 Paris, Cedex 10, France;Tel: +33 (1) 42 49 42 34; Fax: +33 (1) 42 00 01 60E-mail: [email protected] ®rst two authors contributed equally to the workReceived October 1999; accepted 15 May 2000
The Hematology Journal (2000) 1, 316 ± 328ã 2000 The European Haematology Association All rights reserved 1466 ± 4680/00 $15.00
www.nature.com/thj

only this subtype of leukemia suggests that RAdi�erentiation target genes are accessible. E�cacy ofleukemic cell di�erentiation induction is closely relatedto the blockage of di�erentiation resulting from alteredkey proteins such as RARa. Thus acute leukemiascharacterized by speci®c alterations of hematopoieticregulatory proteins should be the ®rst targets ofdi�erentiation therapy. Among these, ®gure a subsetof M2 AML leukemias characterized by the t(8;21)translocation.4
The t(8;21) generates the AML1-ETO fusionprotein.5 ± 8 The AML1 gene on chromosome 21encodes a protein also known as core binding factoralpha (CBFa), an important key transcriptionalmyeloid protein9 which forms a heterodimer with thecore binding factor beta (CBFb). This CBF hetero-dimer is an important regulator of a number of targetgenes involved in hematopoiesis. It binds to theenhancer core motif (TGT/cGGT) which is present inthe promoter region of AML1 target genes, such asgenes coding for myeloperoxidase, NSE,10 IL-3,11 GM-CSF,12 M-CSF receptor13 or TCRb.14 Maximaltransactivating e�cacy of AML1 is enhanced by thepresence of c-myb15 and by the interaction of AML1with MEF (myeloid ELF-16 like factor), a recentlycloned member of the ETS family.16 The role of AML1in hematopoiesis is also re¯ected in the severealteration of fetal liver hematopoiesis of murineembryos which lack AML1,17 as well as in the numberof acute leukemias de®ned by chromosomal transloca-tions t(8;21) and t(3;21) in AML, and t(12;21) in ALL,resulting in alteration of the AML1 gene.18,19 AML1 isalso indirectly targeted through inv16 which disruptsthe core binding factor beta.6
The ETO gene of chromosome 8 implicated in thet(8;21) translocation equally encodes a nuclearphosphoprotein whose function is becoming moreexplicit. It is the homolog of the Drosophila Nervyprotein20 and includes a domain closely related to theDrosophila coactivator of TAF110 (the transcriptionactivating factor 110).21 It is expressed weakly inCD34+ hematopoietic cells and high levels are foundin the brain.22 It contains two zinc ®nger motifs andproline rich, or proline/serine/threonine rich, AP-1regions. It has recently been shown to bind to proteinwhich control chromatin modi®cations such as HDAC(histone deacetylase) or corepressors (N-CoR) andSin3A.23± 26
The t(8;21) translocation fuses 177 amino-acids ofthe amino terminal portion of AML1 to nearly the full-length ETO. The portion of AML1 included in thefusion protein retains the AML1 runt DNA-bindingdomain and acts as a dominant negative on theexpression of AML1 target genes. AML1-ETO retainsthe ability to recognize the AML1 consensus bindingsite and to dimerize with CBFb.9 The interaction ofAML1-ETO with the normal pathways of AML1 hasbeen clearly demonstrated both in vitro and in vivo.AML1-ETO inhibits AML1 functions and induces theaberrant expression and function of ETO in themyeloid stem cell.12 The transforming activity of
AML1-ETO may be through activation of AP-1.27
AML1-ETO, and not AML1a or AML1b, have beenfound to increase the transactivation of the BCL2promoter28 and is certainly a major component of theleukemogenetic process.
AML1-ETO can bind to the transcription factor C/EBPa, inhibit C/EBP/a-dependent transcription andblock granulocyte di�erentiation.29,30 This fusionprotein is also known to inhibit G-CSF-induceddi�erentiation of the myeloid progenitor cell line32Dcl331 and erythroid di�erentiation of K562 cellsby low dose Ara-C32 and knock in of AML-ETOinduces hematopoietic disorders.33,34 However, thepresence of AML1-ETO does not appear to irrever-sibly inhibit myeloid di�erentiation. Using RT±PCR,AML1-ETO transcripts have been detected in CFU-GM, BFU-E, CFU-MK and CFU-Mix progenitors,suggesting that AML1-ETO does not impede the invitro di�erentiation of the myeloid stem cell, in thepresence of cytokines.35 M2 AML t(8;21) leukemias areknown to exhibit prominent granulocytic differentia-tion, and extramedullary in®ltration.36 ± 39 The presenceof AML1-ETO in long-term remission, as in maturegranulocytes, re¯ects in vivo di�erentiation of theleukemic clone.35,40,41 Furthermore, establishment ofthe t(8;21) cell lines Kasumi-1 and SKNO-1,42,43 hasfurther allowed the documentation of the in vitrosensitivity of t(8;21) leukemic cells to growth factors,such as IL-3, GM-CSF, IL-5 and TPA and G-CSF.42,44
Kasumi-1 cells can di�erentiate into macrophage-likecells.45 Two patient samples have been reported withsome degree of induction of di�erentiation in thepresence of IL-5 and G-CSF.46,47
Though M2 AML t(8;21) patients have a goodsensitivity to chemotherapy, survival at ®ve years isstill poor, and an improvement of the therapeuticregimens is necessary. One patient with M2 AMLt(8;21) was recently shown to achieve completeremission after induction therapy with G-CSF alone.48
In order to test whether M2 AML patients could bene®tfrom di�erentiation therapy we studied the predictivityof in vitro di�erentiation of fresh M2 AML t(8;21)samples. We show: (a) the in vitro and in vivodi�erentiation of fresh M2 AML t(8;21) cells uponaddition of G-CSF; (b) the integrity of G-CSF signalingpathways; and (c) the degradation of AML1-ETOprotein after G-CSF-induced di�erentiation.
Materials and methods
Proliferation, di�erentiation and apoptosisassessment
Bone marrow and blood samples were collected from10 patients diagnosed with t(8;21) AML after informedconsent. Mononuclear cells from samples wereobtained by Ficoll-Hypaque density gradient centrifu-gation and either used immediately for cell culture orstored in liquid nitrogen. Bone marrow and/or bloodsamples showed more than 40% blasts at diagnosis and
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
317
The Hematology Journal

cytospins of ®colled puri®ed leukemic samples pre-sented from 70 to 90% blast in®ltration.
Cells were grown in the presence or absence 50 ng/mlrhG-CSF (gift from Laboratoires Rhoà ne-PoulencRorer, Montrouge, France) or all-trans RA 1077 M
(gift from Ho�mann-Laroche, Basel, Switzerland) inRPMI 1640 medium supplemented with 10% fetal calfserum, 2 mmol/l L-glutamine, 100 U/ml penicillin and100 mg/ml streptomycine (all culture media fromBoehringer-Ingelheim Bioproducts, Gagny, France) atan initial concentration of 106 cells/ml. Cell proliferationand viability were studied by trypan blue exclusion.Maturation was monitored by May-GruÈ nwald Giemsa(MGG) staining of cytospin smears and expression ofcell surface antigens by ¯ow cytometry. Brie¯y, 56105
cells were incubated with monoclonal antibodies(Becton Dickinson, Le Pont de Claix, France) or withisotype-identical negative control antibody for 30 min at48C. After two washes in PBS, ¯ow cytometric analysiswas performed on a Becton Dickinson FACSCalibur.Apoptosis was measured on morphological criteria andpropidium iodide staining by ¯ow cytometry aspreviously described.49 In four patients, rhG-CSF wasadministered before conventional therapy, and monitor-ing of in vivo di�erentiation of the peripheral leukemiccells was performed. Management of G-CSF therapywas conducted according to the guidelines of ATRAdi�erentiation therapy in AML.1 Brie¯y, patients werescheduled to receive AML conventional therapy with apre-treatment by G-CSF (Lenograstim, Rhoà ne-PoulencRorer, France) at 5 mg/kg/day for 21 days, a dosepreviously administered in AML patients concomitantwith chemotherapy for the reduction of aplasia.50
Patients entering the assay had a previous diagnosis oft(8;21) AML1-ETO positive leukemia, were free ofgranulocytic sarcoma by tomodensitometry and gaveinformed consent. Blood count was monitored daily,bone marrow smears performed at day 0, 15 and 21. Inthe case of increased WBC during rhG-CSF therapy,administration of chemotherapy was anticipated andrhG-CSF withdrawn.
Preparation of nuclear extracts
Starved cells were stimulated at 378C with rhG-CSF(100 ng/ml). Cell pellets were solubilized in bu�er A:20 mM HEPES pH 8, 10 mM KCl, 0.1 mM EDTA,0.1% Nonidet P40, 0.1 mM Na2VO4, 1 mM DTT, 10%glycerol and proteases inhibitors (AEBSF 250 mg/ml,pepstatin, leupeptin and aprotinin 1 mg/ml). Aftercentrifugation (5 min at 2000 r.p.m.), nuclei wereextracted with bu�er B: 20 mM HEPES pH 8, 400 mM
NaCl, 10 mM KCl, 0.1 mM EDTA, 0.1 mM Na2VO4,1 mM DTT, 20% glycerol and proteases inhibitors.Cytoplasmic and nuclear extracts were immediatelyfrozen in liquid nitrogen and stored at 7708C.
Electrophoretic Mobility Shift Assays (EMSA)
Nuclear extracts (2 ml) were incubated for 20 min atroom temperature with 50 000 c.p.m. of 32P-labeled
double-stranded probe in binding bu�er: 200 mM
HEPES pH 7.5, 400 mM KCl, 10 mM MgCl2, 1 mM
EGTA, 5 mM DTT, 4% Ficoll and 2 mg/ml poly(dI-dC). The DNA-protein complexes were separated byelectrophoresis on a 4% non-denaturing polyacryla-mide gel in 0.256TBE and detected by autoradio-graphy. In some experiments, the radioactivity wasquanti®ed using a Molecular Imager (GS525, Biorad,Ivry sur Seine, France) and analyzed with MolecularAnalyst software (Biorad, Ivry sur Seine, France). Forcompetition analysis, a 100-fold excess of unlabeledprobe was added in the binding mix. For supershiftanalysis, nuclear extracts were pre-incubated for30 min with 1 mg of anti-STAT1 (raised against aprotein fragment corresponding to amino-acids 592 ±731 of STAT1, Transduction Laboratories, San Diego,USA) or anti-STAT3 (C-20, raised against a peptidecorresponding to amino-acids 750 ± 769 mapping at thecarboxy terminus of STAT3, Santa Cruz Biotechnol-ogy, CA, USA) antibodies. The oligonucleotide used inthis study was m67 (5'-GATCGACATTTCCCG-TAAATC-3'), containing a high a�nity mutant ofthe serum-inducible element (hSIE) of the human c-fospromoter.
Total protein extraction and Westernimmunoblotting
Five million cells were harvested by centrifugation andmixed with 0.5 ml lysis bu�er containing 320 mM
sucrose, 10 mM HEPES pH 7.5, 10 mM Tris-HCl pH6.8, 1% SDS, 2% glycerol and 5% b-mercaptoethanol.Protein concentration was determined according to themanufacturer's instructions. Equal loadings werechecked by Coomassie blue and Ponceau red stain-ings. After electroblotting, nitrocellulose membranes(Hybond-ECL, Amersham, Les Ulis, France) wereimmunoprobed overnight with an anti-ETO antibodyAP1170 (gift of HA Drabkin). After incubation withan anti-rabbit IgG antibody horse radish peroxidaseconjugate (Boehringer-Mannheim, Meylan, Germany),immunoreactive proteins were revealed using ECLchemiluminescence detection kit (Amersham, LesUlis, France).
Results
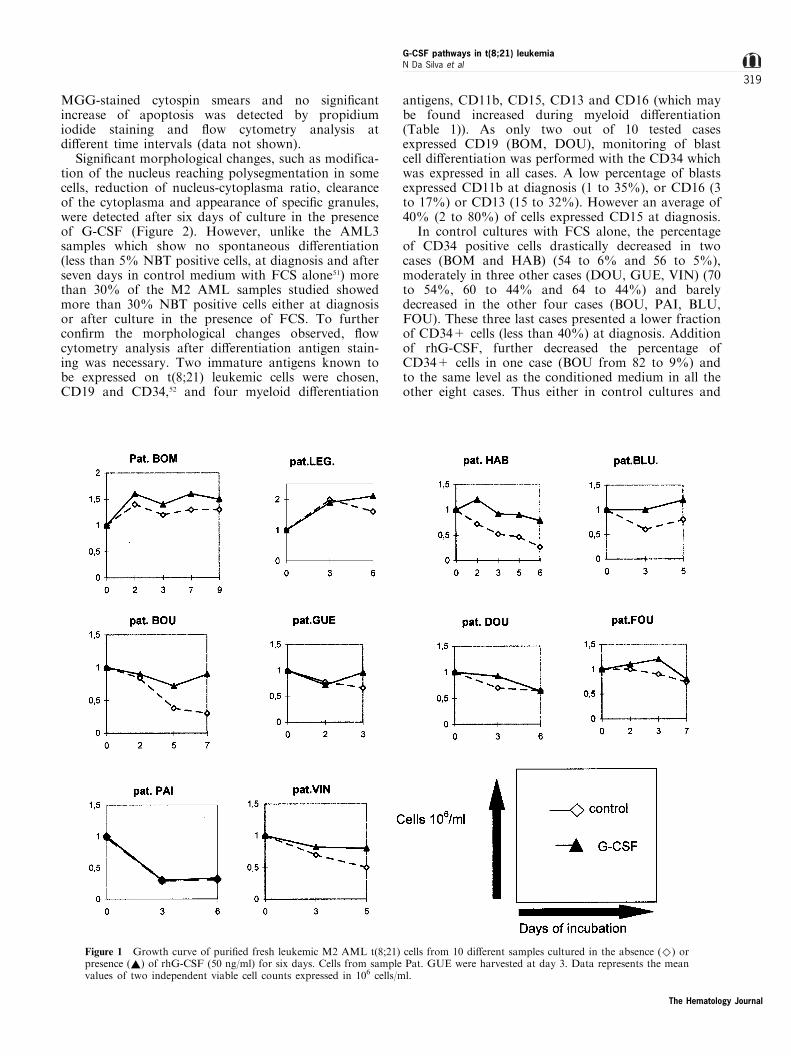
Di�erentiation of M2 AML t(8;21) blasts byrhG-CSF
In all but two patients (BOM, LEG) (eight out of 10)leukemic cell viability decreased during the seven-dayculture period by about 50%, in the presence of FCSalone. Addition of rhG-CSF: (a) allowed the main-tenance of a viable cell count in four patients (HAB,BLU, BOU, GUE); (b) had no or little e�ect in fourcases (FOU, DOU, PAI, VIN); and (c) was superior(though marginally) to the initial seeded cell concen-tration in only two cases (BOM and LEG) (Figure 1).Few apoptotic cells (less than 15%) were observed on
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
318
MGG-stained cytospin smears and no signi®cantincrease of apoptosis was detected by propidiumiodide staining and ¯ow cytometry analysis atdi�erent time intervals (data not shown).
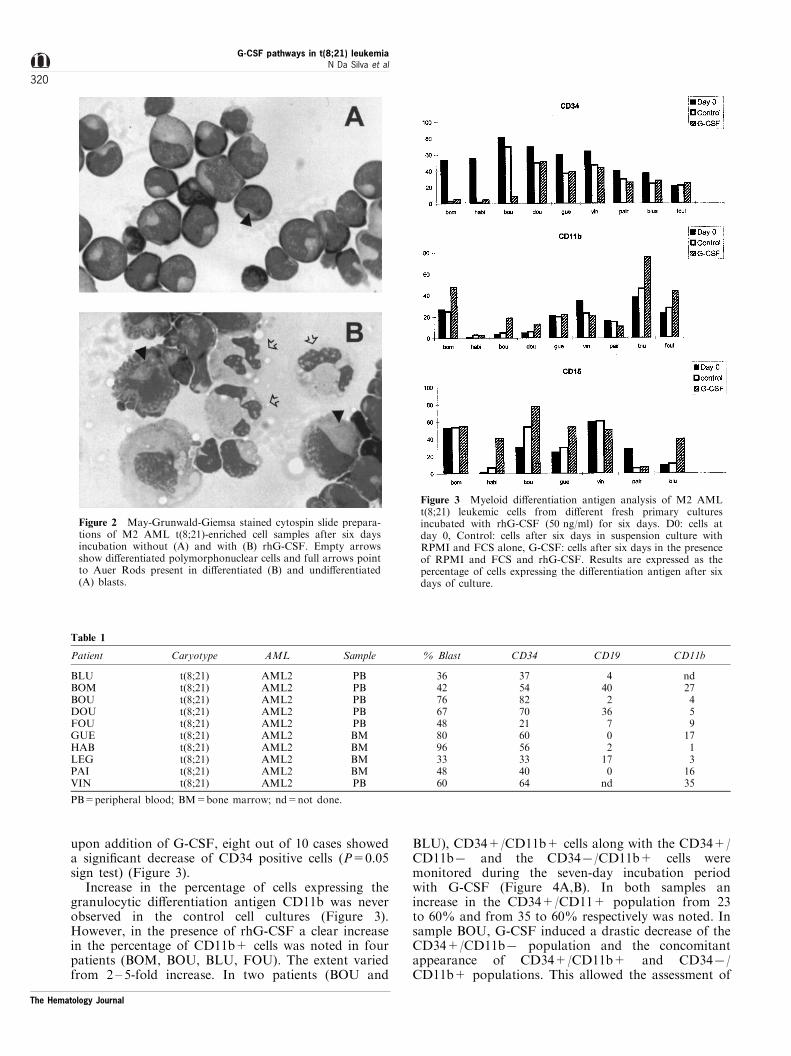
Signi®cant morphological changes, such as modifica-tion of the nucleus reaching polysegmentation in somecells, reduction of nucleus-cytoplasma ratio, clearanceof the cytoplasma and appearance of speci®c granules,were detected after six days of culture in the presenceof G-CSF (Figure 2). However, unlike the AML3samples which show no spontaneous di�erentiation(less than 5% NBT positive cells, at diagnosis and afterseven days in control medium with FCS alone51) morethan 30% of the M2 AML samples studied showedmore than 30% NBT positive cells either at diagnosisor after culture in the presence of FCS. To furthercon®rm the morphological changes observed, ¯owcytometry analysis after di�erentiation antigen stain-ing was necessary. Two immature antigens known tobe expressed on t(8;21) leukemic cells were chosen,CD19 and CD34,52 and four myeloid di�erentiation
antigens, CD11b, CD15, CD13 and CD16 (which maybe found increased during myeloid di�erentiation(Table 1)). As only two out of 10 tested casesexpressed CD19 (BOM, DOU), monitoring of blastcell di�erentiation was performed with the CD34 whichwas expressed in all cases. A low percentage of blastsexpressed CD11b at diagnosis (1 to 35%), or CD16 (3to 17%) or CD13 (15 to 32%). However an average of40% (2 to 80%) of cells expressed CD15 at diagnosis.
In control cultures with FCS alone, the percentageof CD34 positive cells drastically decreased in twocases (BOM and HAB) (54 to 6% and 56 to 5%),moderately in three other cases (DOU, GUE, VIN) (70to 54%, 60 to 44% and 64 to 44%) and barelydecreased in the other four cases (BOU, PAI, BLU,FOU). These three last cases presented a lower fractionof CD34+ cells (less than 40%) at diagnosis. Additionof rhG-CSF, further decreased the percentage ofCD34+ cells in one case (BOU from 82 to 9%) andto the same level as the conditioned medium in all theother eight cases. Thus either in control cultures and
Figure 1 Growth curve of puri®ed fresh leukemic M2 AML t(8;21) cells from 10 di�erent samples cultured in the absence (^) orpresence (~) of rhG-CSF (50 ng/ml) for six days. Cells from sample Pat. GUE were harvested at day 3. Data represents the meanvalues of two independent viable cell counts expressed in 106 cells/ml.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
319
upon addition of G-CSF, eight out of 10 cases showeda signi®cant decrease of CD34 positive cells (P=0.05sign test) (Figure 3).
Increase in the percentage of cells expressing thegranulocytic di�erentiation antigen CD11b was neverobserved in the control cell cultures (Figure 3).However, in the presence of rhG-CSF a clear increasein the percentage of CD11b+ cells was noted in fourpatients (BOM, BOU, BLU, FOU). The extent variedfrom 2 ± 5-fold increase. In two patients (BOU and
BLU), CD34+/CD11b+ cells along with the CD34+/CD11b7 and the CD347/CD11b+ cells weremonitored during the seven-day incubation periodwith G-CSF (Figure 4A,B). In both samples anincrease in the CD34+/CD11+ population from 23to 60% and from 35 to 60% respectively was noted. Insample BOU, G-CSF induced a drastic decrease of theCD34+/CD11b7 population and the concomitantappearance of CD34+/CD11b+ and CD347/CD11b+ populations. This allowed the assessment of
Figure 2 May-Grunwald-Giemsa stained cytospin slide prepara-tions of M2 AML t(8;21)-enriched cell samples after six daysincubation without (A) and with (B) rhG-CSF. Empty arrowsshow di�erentiated polymorphonuclear cells and full arrows pointto Auer Rods present in di�erentiated (B) and undi�erentiated(A) blasts.
Figure 3 Myeloid di�erentiation antigen analysis of M2 AMLt(8;21) leukemic cells from di�erent fresh primary culturesincubated with rhG-CSF (50 ng/ml) for six days. D0: cells atday 0, Control: cells after six days in suspension culture withRPMI and FCS alone, G-CSF: cells after six days in the presenceof RPMI and FCS and rhG-CSF. Results are expressed as thepercentage of cells expressing the di�erentiation antigen after sixdays of culture.
Table 1
Patient Caryotype AML Sample % Blast CD34 CD19 CD11b
BLUBOMBOUDOUFOUGUEHABLEGPAIVIN
t(8;21)t(8;21)t(8;21)t(8;21)t(8;21)t(8;21)t(8;21)t(8;21)t(8;21)t(8;21)
AML2AML2AML2AML2AML2AML2AML2AML2AML2AML2
PBPBPBPBPBBMBMBMBMPB
36427667488096334860
37548270216056334064
440236702170
nd
nd2745917131635
PB=peripheral blood; BM=bone marrow; nd=not done.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
320
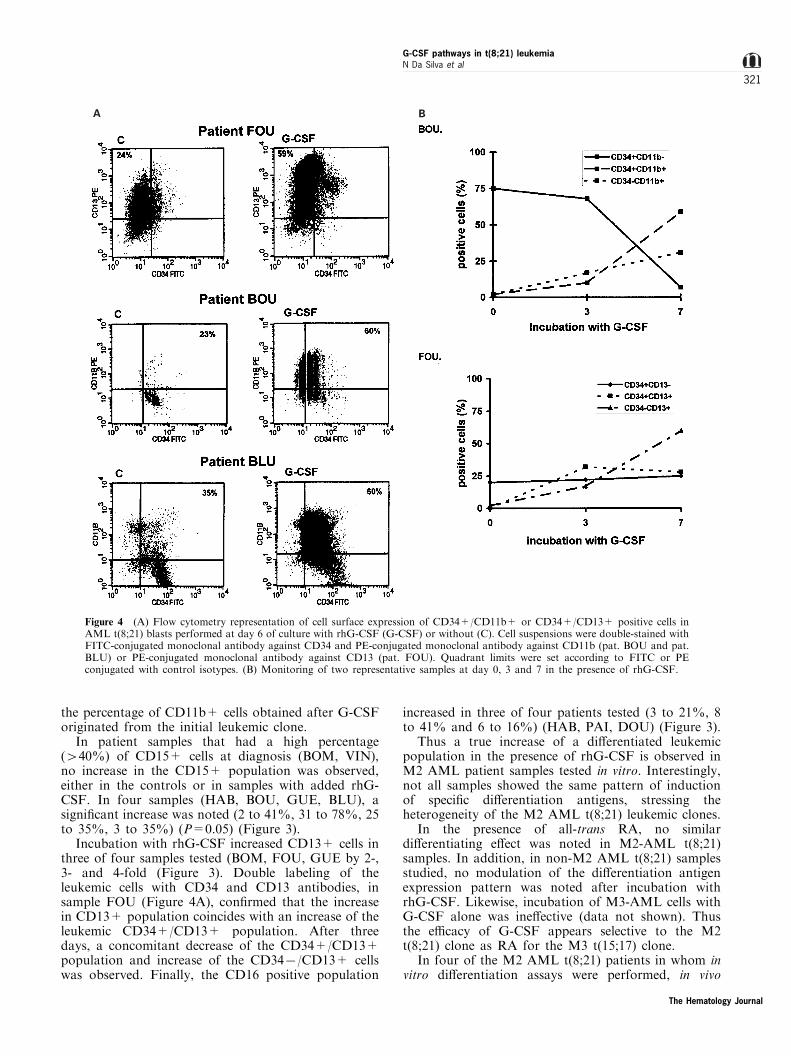
the percentage of CD11b+ cells obtained after G-CSForiginated from the initial leukemic clone.
In patient samples that had a high percentage(440%) of CD15+ cells at diagnosis (BOM, VIN),no increase in the CD15+ population was observed,either in the controls or in samples with added rhG-CSF. In four samples (HAB, BOU, GUE, BLU), asigni®cant increase was noted (2 to 41%, 31 to 78%, 25to 35%, 3 to 35%) (P=0.05) (Figure 3).
Incubation with rhG-CSF increased CD13+ cells inthree of four samples tested (BOM, FOU, GUE by 2-,3- and 4-fold (Figure 3). Double labeling of theleukemic cells with CD34 and CD13 antibodies, insample FOU (Figure 4A), con®rmed that the increasein CD13+ population coincides with an increase of theleukemic CD34+/CD13+ population. After threedays, a concomitant decrease of the CD34+/CD13+population and increase of the CD347/CD13+ cellswas observed. Finally, the CD16 positive population
increased in three of four patients tested (3 to 21%, 8to 41% and 6 to 16%) (HAB, PAI, DOU) (Figure 3).
Thus a true increase of a di�erentiated leukemicpopulation in the presence of rhG-CSF is observed inM2 AML patient samples tested in vitro. Interestingly,not all samples showed the same pattern of inductionof speci®c di�erentiation antigens, stressing theheterogeneity of the M2 AML t(8;21) leukemic clones.
In the presence of all-trans RA, no similardi�erentiating e�ect was noted in M2-AML t(8;21)samples. In addition, in non-M2 AML t(8;21) samplesstudied, no modulation of the di�erentiation antigenexpression pattern was noted after incubation withrhG-CSF. Likewise, incubation of M3-AML cells withG-CSF alone was ine�ective (data not shown). Thusthe e�cacy of G-CSF appears selective to the M2t(8;21) clone as RA for the M3 t(15;17) clone.
In four of the M2 AML t(8;21) patients in whom invitro di�erentiation assays were performed, in vivo
A B
Figure 4 (A) Flow cytometry representation of cell surface expression of CD34+/CD11b+ or CD34+/CD13+ positive cells inAML t(8;21) blasts performed at day 6 of culture with rhG-CSF (G-CSF) or without (C). Cell suspensions were double-stained withFITC-conjugated monoclonal antibody against CD34 and PE-conjugated monoclonal antibody against CD11b (pat. BOU and pat.BLU) or PE-conjugated monoclonal antibody against CD13 (pat. FOU). Quadrant limits were set according to FITC or PEconjugated with control isotypes. (B) Monitoring of two representative samples at day 0, 3 and 7 in the presence of rhG-CSF.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
321
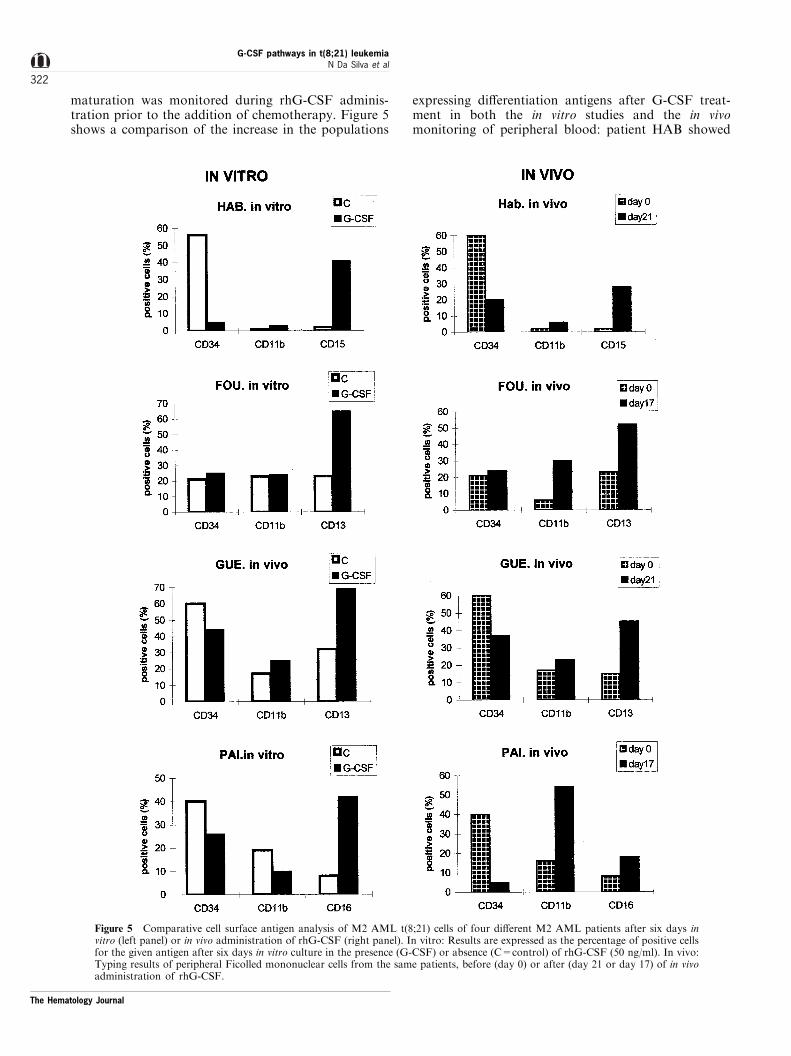
maturation was monitored during rhG-CSF adminis-tration prior to the addition of chemotherapy. Figure 5shows a comparison of the increase in the populations
expressing di�erentiation antigens after G-CSF treat-ment in both the in vitro studies and the in vivomonitoring of peripheral blood: patient HAB showed
Figure 5 Comparative cell surface antigen analysis of M2 AML t(8;21) cells of four di�erent M2 AML patients after six days invitro (left panel) or in vivo administration of rhG-CSF (right panel). In vitro: Results are expressed as the percentage of positive cellsfor the given antigen after six days in vitro culture in the presence (G-CSF) or absence (C=control) of rhG-CSF (50 ng/ml). In vivo:Typing results of peripheral Ficolled mononuclear cells from the same patients, before (day 0) or after (day 21 or day 17) of in vivoadministration of rhG-CSF.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
322
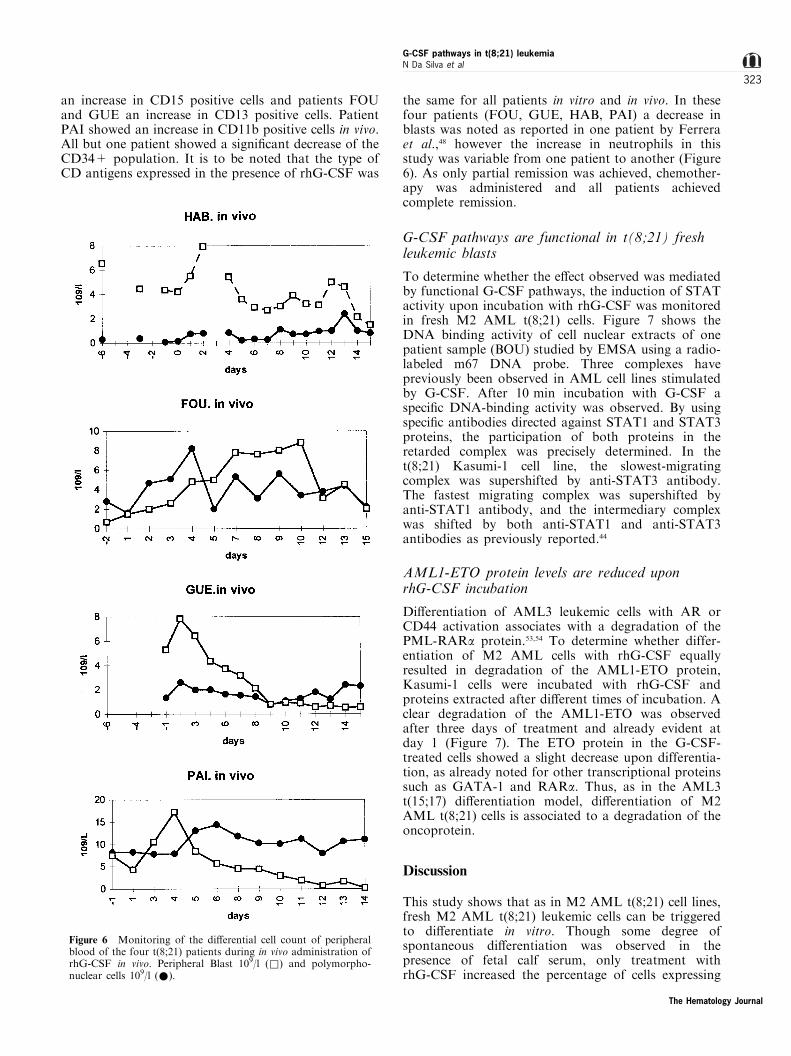
an increase in CD15 positive cells and patients FOUand GUE an increase in CD13 positive cells. PatientPAI showed an increase in CD11b positive cells in vivo.All but one patient showed a signi®cant decrease of theCD34+ population. It is to be noted that the type ofCD antigens expressed in the presence of rhG-CSF was
the same for all patients in vitro and in vivo. In thesefour patients (FOU, GUE, HAB, PAI) a decrease inblasts was noted as reported in one patient by Ferreraet al.,48 however the increase in neutrophils in thisstudy was variable from one patient to another (Figure6). As only partial remission was achieved, chemother-apy was administered and all patients achievedcomplete remission.
G-CSF pathways are functional in t(8;21) freshleukemic blasts
To determine whether the e�ect observed was mediatedby functional G-CSF pathways, the induction of STATactivity upon incubation with rhG-CSF was monitoredin fresh M2 AML t(8;21) cells. Figure 7 shows theDNA binding activity of cell nuclear extracts of onepatient sample (BOU) studied by EMSA using a radio-labeled m67 DNA probe. Three complexes havepreviously been observed in AML cell lines stimulatedby G-CSF. After 10 min incubation with G-CSF aspeci®c DNA-binding activity was observed. By usingspeci®c antibodies directed against STAT1 and STAT3proteins, the participation of both proteins in theretarded complex was precisely determined. In thet(8;21) Kasumi-1 cell line, the slowest-migratingcomplex was supershifted by anti-STAT3 antibody.The fastest migrating complex was supershifted byanti-STAT1 antibody, and the intermediary complexwas shifted by both anti-STAT1 and anti-STAT3antibodies as previously reported.44
AML1-ETO protein levels are reduced uponrhG-CSF incubation
Di�erentiation of AML3 leukemic cells with AR orCD44 activation associates with a degradation of thePML-RARa protein.53,54 To determine whether differ-entiation of M2 AML cells with rhG-CSF equallyresulted in degradation of the AML1-ETO protein,Kasumi-1 cells were incubated with rhG-CSF andproteins extracted after di�erent times of incubation. Aclear degradation of the AML1-ETO was observedafter three days of treatment and already evident atday 1 (Figure 7). The ETO protein in the G-CSF-treated cells showed a slight decrease upon differentia-tion, as already noted for other transcriptional proteinssuch as GATA-1 and RARa. Thus, as in the AML3t(15;17) di�erentiation model, di�erentiation of M2AML t(8;21) cells is associated to a degradation of theoncoprotein.
Discussion
This study shows that as in M2 AML t(8;21) cell lines,fresh M2 AML t(8;21) leukemic cells can be triggeredto di�erentiate in vitro. Though some degree ofspontaneous di�erentiation was observed in thepresence of fetal calf serum, only treatment withrhG-CSF increased the percentage of cells expressing
Figure 6 Monitoring of the di�erential cell count of peripheralblood of the four t(8;21) patients during in vivo administration ofrhG-CSF in vivo. Peripheral Blast 109/l (&) and polymorpho-nuclear cells 109/l (*).
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
323

myeloid di�erentiation antigens, both in vitro and invivo. This increase in myeloid marker expression wasassociated with a decrease of CD34+ leukemic cells. Invivo, this di�erentiation is concomitant with an increasein neutrophils and the disappearance of blast cells; invitro, it is accompanied by a degradation of the AML1-ETO oncoprotein. The fact that normal induction andbinding of STAT3 and STAT1 homodimers andheterodimers were observed upon G-CSF treatmentin both the Kasumi-1 cell line and the fresh leukemiccells is in favor of G-CSF functional pathways in theseleukemic cells.
This novel di�erentiating model bears some analogywith the RA-APL model: (a) e�cacy of a physiologicalgranulocyte di�erentiating factor;55 ± 57 (b) e�ectiveactivation of the agent's signaling pathways;58 and (c)degradation of the oncoprotein.53,54 In both models,di�erentiation can be monitored by the progressiveappearance of a population expressing immature andmature di�erentiation markers (CD33 and CD15 forAPL cells, CD34 and CD11b or CD13 in M2 AMLt(8;21) cells.59,60 The induction of di�erentiation is alsolinked, both in vitro and in vivo, with a transientincrease of cell proliferation51,59 with no signi®cantincrease in apoptosis in vitro.49,61 In both models, theleukemic clone is linked to a speci®c translocationinvolving a myeloid transcription factor (RARa orAML1) and a nuclear protein (PML or ETO) found tobind to transcriptional nuclear complexes. The fusionprotein in both cases represses the transactivation ofthe target myeloid genes and inhibits the granulocytic
di�erentiation induced by the di�erentiation agent innormal sensitive cell lines31,62 ± 65 (Table 2).
None the less, the data presented in this study showsthat, unlike the RA-APL model, complete terminaldi�erentiation of the entire t(8;21) leukemic cellpopulation was not achieved in vitro in primarysamples. This may be related to the fact that someAML1-ETO leukemic cells are more immature thanothers, or it may be that the carcinogenesis of thisleukemia, contrary to APL, is associated with othermolecular events, preventing e�ective di�erentiation.Nevertheless, a speci®c di�erentiating response of G-CSF was evident in t(8;21) leukemia. In the patients ofthis study and with the criteria used, no signi®cantdi�erentiating e�ect was observed with all-trans RA orwith IL-3, GM-CSF or FCS alone. Likewise, G-CSFdid not allow di�erentiation of other subsets ofleukemias, contrary to the recent ®ndings with CD44stimulation.53 It is well known that speci®c leukemicsubtypes may confer speci®c sensitivity to variousagents (chemotherapy, or inducers of di�erentiation orapoptosis). Recently, it has been shown that dexa-methasone selectively inhibits Kasumi-1 and SKNO-1cells, and fresh M2 AML t(8;21) samples, atconcentrations as low as 1 nM by inducing apopto-sis.66,67
What molecular mechanisms inherent to the t(8;21)leukemic cells favor G-CSF e�cacy? While direct dataare not available, various results put forward severalworking hypotheses. Expression of AML1-ETO blocksG-CSF-induced di�erentiation of myeloid stem cells29 ±31 and this inhibition is abrogated by pharmacologicaldoses of G-CSF. This implies that di�erentiation genes,and especially G-CSF target genes, have remainedaccessible in the leukemic cells and that G-CSFpathways are functional. We have shown thatactivation and kinetics of activation of STAT1 andSTAT3 homo- and heterodimers in these cells upon G-CSF incubation are normal. On the other hand, theaccessible genes in each M2 AML t(8;21) leukemicclone may be speci®c to a given patient, as thedi�erentiation antigens induced were variable fromone clone to another. This further stresses that theblockage of di�erentiation in this leukemia may be atdi�erent stages of myeloid di�erentiation.
Could AML1-ETO play a part in the induction ofdi�erentiation? The runt domain of AML1-ETO, isrequired for cell di�erentiation of Kasumi-1 cells withPMA.45 Thus, as in the case of PML-RARa in APLcells, AML1-ETO may be involved in the initiation ofdi�erentiation induction. Whether this is due to aspeci®c and direct e�ect of AML1-ETO on differ-entiation target genes or a consequence of thepresence of AML1-ETO in these cells is not clear.Like PML-RARa in di�erentiated APL cells, AML1-ETO is degraded in G-CSF-treated-t(8;21) cells whichmay further increase the di�erentiation process.Indeed antisense experiments against the AML1-ETO junction increase the sensitivity to PMA, andantisense to AML1-ETO alone favors granulocyticdi�erentiation.
++
+++
+ ++ +
α-STAT1α-STAT3 N.S.
SUPERSHIFTS
STATs COMPLEXES
S.C.
–G-CSF +G-CSF
Kd D0 D1 D3 D4 D5 D1 D3 D4 D5105–
75–AML1/ETOETO
EXTRACT FROM PATIENT BOU EXTRACT FROM KASUMI-1
A
B
Figure 7 (A) Analysis of STAT-protein activity in rhG-CSF-induced DNA binding complexes. Nuclear extracts of M2 AMLt(8;21) blasts of patient BOU and the cell line Kasumi-1 wereincubated during 10 min with 100 ng/ml of rhG-CSF. EMSAanalysis was performed with the m67 speci®c oligonucleotideprobe and antibodies against STAT1 pr STAT3 proteins. (B)Immunoblot of t(8;21) leukemic cell extracts with a speci®cantibody against ETO. The 105 Kd band corresponds to theAML1-ETO oncoprotein which is degraded after rhG-CSFincubation.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
324

AML1-ETO induces an upregulation of cytokinereceptors and t(8;21) leukemic cells are known to besensitive to cytokines.35 A synergistic up-regulation ofthe M-CSFR by AML1 and AML1-ETO has beenobserved,13 and a higher expression of M-CSFR isnoted in Kasumi-1 cell lines and in cells from M2AML t(8;21) patients.13 One study reported high G-CSFR in one patient sample with M2 AML t(8;21).68
Whether this relates to G-CSF e�cacy in these cellsremains to be explored. As receptor structure is crucialfor the biological e�ect of a cytokine, the dose-a�nityrelationship between G-CSFR of t(8;21) cells and theconcentration of G-CSF should be taken into accountto distinguish between proliferative and di�erentiatione�ects and is crucial for clinical trials with rhG-CSF.Indeed hyperleukocytosis with ATRA di�erentiatingtherapy is a major complication of this novel therapywhen not controlled by chemotherapy.69 G-CSF hasbeen shown to increase the proliferation of AML cells70
and in this study, rhG-CSF increased viable cell countin nearly all samples tested in vitro while a discreteincrease in WBC count was noted in all patients. Thisis reminiscent of the increase in WBC upon ATRAtherapy in APL and testi®es to the in vivo e�ect of bothdi�erentiating agents.
Dose-response studies performed on cell lines and infresh patient samples with t(8;21) (Da Silva andChomienne, unpublished results) showed that, unlikewith other cytokines such as GM-CSF and IL-3, thereexists a dose-dependent sensitivity of these blasts to G-CSF and suggests that receptor a�nity may still beadjusted. In the RA-APL model, in vivo e�cacy isrelated to a structure-dose e�ect.53 Thus the e�ect of G-CSF in these cells could be adapted to the G-CSF
signaling pathways present in the leukemic cell (fromreceptor a�nity to transduction signals and targetgenes). In particular, STAT5 activation or the balanceof STAT3a and STAT3b response to G-CSF71 shouldbe interesting to target. As the preliminary in vivo dataof this study correlate well with in vitro results,improvement of di�erentiation achievement may bewell attempted on fresh primary cell cultures.
Prior conventional trials in AML have alreadyshown that addition of G-CSF to chemotherapy forAML patients prolongs remission,50 and this could berelated to the association of di�erentiation andcytotoxic chemotherapy, which is now known toprolong survival in APL patients. Indeed data fromFerrara et al.48 and our combined in vitro and in vivostudies, show that, as in the APL model, in vitrostudies on primary M2 AML t(8;21) cultures shouldallow the design of more e�ective therapeutic regimenssimultaneously associating di�erentiation, transcriptioninduction and cytotoxicity.
AcknowledgementsWe thank HA Drabkin (Denver, Colorado, USA) for theinterest in the study and the gift of the ETO antibody. Weare indebted to Rose Ann Padua for instructive discussionsand reviewal of the manuscript. This study was supportedby grants from the Association de la Recherche Contre leCancer, Fondation Saint-Louis. S Meyer-Monard is agrant holder from Ciba-Geigy JubilaÈ umstiftung, Krebsligabeider, Basel, Switzerland and Swiss National Foundation.N Da Silva was supported by a grant from AgenceNationale de la Recherche Technique (ANRT) andLaboratoires Rhoà ne-Poulenc Rorer, Montrouge, France.
References
1 Fenaux P, Chastang C, Chevret S, Sanz M, Dombret H,Archimbaud E, Fey M, Rayon C, Huguet F, Sotto JJ,Gardin C, Makhoul PC, Travade P, Solary E, FegueuxN, Bordessoule D, Miguel JS, Link H, Desablens B,Stamatoullas A, Deconinck E, Maloisel F, Castaigne S,Preudhomme C, Degos L. A randomized comparison ofall transretinoic acid (ATRA) followed by chemotherapyand ATRA plus chemotherapy and the role of main-tenance therapy in newly diagnosed acute promyelocyticleukemia. Blood 94: 1192, 1999.
2 Olsson I, Bergh G, Ehinger M, Gullberg U. Celldi�erentiation in acute myeloid leukemia. EuropeanJournal of Haematology 57: 1, 1996.
3 Grignani F, Fagioli M, Alcalay M, Longo L, Pandol® PP,Donti E, Biondi A, Lo Coco F, Grignani F, Pelicci PG.Acute promyelocytic leukemia: from genetics to treat-ment. Blood 83: 10, 1994.
4 Bennett JM, Catovsky D, Daniel MT, Flandrin G,Galton DA, Gralnick HR, Sultan C. Proposals for theclassi®cation of the acute leukaemias. French-American-British (FAB) co-operative group. British Journal ofHaematology 33: 451, 1976.
5 Erickson P, Gao J, Chang KS, Look T, Whisenant E,Raimondi S, Lasher R, Trujillo J, Rowley J, Drabkin H.Identi®cation of breakpoints in t(8;21) acute myelogen-ous leukemia and isolation of a fusion transcript, AML1/ETO, with similarity to Drosophila segmentation gene,runt. Blood 80: 1825, 1992.
6 Miyoshi H, Shimizu K, Kozu T, Maseki N, Kaneko Y,Ohki M. t(8;21) breakpoints on chromosome 21 in acutemyeloid leukemia are clustered within a limited region ofa single gene, Aml1. Proceedings of the National Academyof Science USA 88: 10431, 1991.
7 Nisson PE, Watkins PC, Sacchi N. Transcriptionallyactive chimeric gene derived from the fusion of the AML1gene and a novel gene on chromosome 8 in t(8;21)leukemic cells [published erratum appears in CancerGenetic and Cytogenetics 1993 Mar; 66(1): 81]. CancerGenetic and Cytogenetics 63: 81, 1992.
8 Miyoshi H, Kozu T, Shimizu K, Enomoto K, Maseki N,Kaneko Y, Kamada N, Ohki M. The t(8;21) transloca-tion in acute myeloid leukemia results in production of anAML1-MTG8 fusion transcript. EMBO Journal 12:2715, 1993.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
325

9 Meyers S, Downing JR, Hiebert SW. Identi®cation ofAML-1 and the (8;21) translocation protein (AML-1/ETO) as sequence-speci®c DNA-binding proteins: therunt homology domain is required for DNA binding andprotein-protein interactions. Molecular Cell Biology 13:6336, 1993.
10 Nuchprayoon I, Meyers S, Scott LM, Suzow J, Hiebert S,Friedman AD. PEBP2/CBF, the murine homolog of thehuman myeloid AML1 and PEBP2 beta/CBF beta proto-oncoproteins, regulates the murine myeloperoxidase andneutrophil elastase genes in immature myeloid cells.Molecular Cell Biology 14: 5558, 1994.
11 Uchida H, Zhang J, Nimer SD. AML1A and AML1B cantransactivate the human IL-3 promoter. Journal ofImmunology 158: 2251, 1997.
12 Frank R, Zhang J, Uchida H, Meyers S, Hiebert SW,Nimer SD. The AML1/ETO fusion protein blockstransactivation of the GM-CSF promoter by AML1B.Oncogene 11: 2267, 1995.
13 Rhoades KL, Hetherington CJ, Rowley JD, Hiebert SW,Nucifora G, Tenen DG, Zhang DE. Synergistic up-regulation of the myeloid-speci®c promoter for themacrophage colony-stimulating factor receptor byAML1 and the t(8;21) fusion protein may contribute toleukemogenesis. Proceedings of the National Academy ofScience USA 93: 11895, 1996.
14 Redondo JM, Hata S, Brocklehurst C, Krangel MS. A Tcell-speci®c transcriptional enhancer within the human Tcell receptor delta locus. Science 247: 1225, 1990.
15 Britos-Bray M, Friedman AD. Core binding factorcannot synergistically activate the myeloperoxidaseproximal enhancer in immature myeloid cells without c-Myb. Molecular Cell Biology 17: 127, 1997.
16 Mao S, Frank RC, Zhang J, Miyazaki Y, Nimer SD.Functional and physical interactions between AML1proteins and an ETS protein, MEF: implications for thepathogenesis of t(8;21)-positive leukemias. MolecularCell Biology 19: 3635, 1999.
17 Okuda T, van Deursen J, Hiebert SW, Grosveld G,Downing JR. AML1, the target of multiple chromosomaltranslocations in human leukemia, is essential for normalfetal liver hematopoiesis. Cell 84: 321, 1996.
18 Nucifora G, Begy CR, Erickson P, Drabkin HA, RowleyJd. The 3;21 translocation in myelodysplasia results in afusion transcript between the AML1 gene and the genefor EAP, a highly conserved protein associated with theEpstein-Barr virus small RNA EBER 1. Proceedings ofthe National Academy of Science USA 90: 7784, 1993.
19 Lo Coco F, Pisegna S, Diverio D. The AML1 gene: atranscription factor involved in the pathogenesis ofmyeloid and lymphoid leukemias. Haematologica 82:364, 1997.
20 Feinstein PG, Kornfeld K, Hogness DS, Mann RS.Identi®cation of homeotic target genes in Drosophilamelanogaster including nervy, a proto-oncogene homo-logue. Genetics 140: 573, 1995.
21 Hoey T, Weinzierl RO, Gill G, Chen JL, Dynlacht BD,Tjian R. Molecular cloning and functional analysis ofDrosophila TAF110 reveal properties expected ofcoactivators. Cell 72: 247, 1993.
22 Erickson PF, Robinson M, Owens G, Drabkin HA. TheETO portion of acute myeloid leukemia t(8;21) fusiontranscript encodes a highly evolutionarily conserved,putative transcription factor. Cancer Research 54: 1782,1994.
23 Wang J, Hoshino T, Redner RL, Kajigaya S, Liu JM.ETO, fusion partner in t(8;21) acute myeloid leukemia,represses transcription by interaction with the human N-CoR/mSin3/HDAC1 complex. Proceedings of the Na-tional Academy of Science USA 95: 10860, 1998.
24 Kitabayashi I, Ida K, Morohoshi F, Yokoyama A,Mitsuhashi N, Shimizu K, Nomura N, Hayashi Y, OhkiM. The AML1-MTG8 leukemic fusion protein forms acomplex with a novel member of the MTG8(ETO/CDR)family, MTGR1. Molecular Cell Biology 18: 846, 1998.
25 Lutterbach B, Westendorf JJ, Linggi B, Patten A,Moniwa M, Davie JR, Huynh KD, Bardwell VJ,Lavinsky RM, Rosenfeld MG, Glass C, Seto E, HiebertSW. ETO, a target of t(8;21) in acute leukemia, interactswith the N-CoR and mSin3 corepressors. Molecular CellBiology 18: 7176, 1998.
26 Gelmetti V, Zhang J, Fanelli M, Minucci S, Pelicci PG,Lazar MA. Aberrant recruitment of the nuclear receptorcorepressor-histone deacetylase complex by the acutemyeloid leukemia fusion partner ETO. Molecular CellBiology 18: 7185, 1998.
27 Frank RC, Sun X, Berguido FJ, Jakubowiak A, NimerSD. The t(8;21) fusion protein, AML1/ETO, transformsNIH3T3 cells and activates AP-1. Oncogene 18: 1701,1999.
28 Klampfer L, Zhang J, Zelenetz AO, Uchida H, NimerSD. The AML1/ETO fusion protein activates transcrip-tion of BCL-2. Proceedings of the National Academy ofScience USA 93: 14059, 1996.
29 Westendorf JJ, Yamamoto CM, Lenny N, Downing JR,Selsted ME, Hiebert SW. The t(8;21) fusion product,AML-1-ETO, associates with C/EBP-alpha, inhibits C/EBP-alpha-dependent transcription, and blocks granulo-cytic di�erentiation.Molecular Cell Biology 18: 322, 1998.
30 Behre G, Zhang P, Zhang DE, Tenen DG. Analysis of themodulation of transcriptional activity in myelopoiesisand leukemogenesis. Methods 17: 231, 1999.
31 Ahn MY, Huang G, Bae SC, Wee HJ, Kim WY, Ito Y.Negative regulation of granulocytic di�erentiation in themyeloid precursor cell line 32Dcl3 by ear-2, a mammalianhomolog of Drosophila seven-up, and a chimericleukemogenic gene, AML1/ETO. Proceedings of theNational Academy of Science USA 95: 1812, 1998.
32 Le XF, Claxton D, Kornblau S, Fan YH, Mu ZM, ChangKS. Characterization of the ETO and AML1-ETOproteins involved in 8;21 translocation in acute myelo-genous leukemia. European Journal of Haematology 60:217, 1998.
33 Yergeau D, Hetherington CJ, Wang Q, Zhang P, SharpeAH, Binder M, Marin-Padilla M, Tenen DG, Speck NA,Zhang DE. Embryonic lethality and impairment ofhaematopoiesis in mice heterozygous for an AML1-ETO fusion gene. Nature Genetics 15: 303, 1997.
34 Okuda T, Cai Z, Yang S, Lenny N, Lyu CJ, van DeursenJM, Harada H, Downing JR. Expression of a knocked-inAML1-ETO leukemia gene inhibits the establishment ofnormal de®nitive hematopoiesis and directly generatesdysplastic hematopoietic progenitors. Blood 91: 3134,1998.
35 Miyamoto T, Nagafuji K, Akashi K, Harada M, Kyo T,Akashi T, Takenaka K, Mizuno S, Gondo H, OkamuraT, Dohy H, Niho Y. Persistence of multipotentprogenitors expressing AML1/ETO transcripts in long-term remission patients with t(8;21) acute myelogenousleukemia. Blood 87: 4789, 1996.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
326

36 Yamasaki H, Era T, Asou N, Sanada I, Matutes E,Yamaguchi K, Takatsuki K. High degree of myeloiddi�erentiation and granulocytosis is associated witht(8;21) smoldering leukemia. Leukemia 9: 1147, 1995.
37 Nucifora G, Dickstein JI, Torbenson V, Roulston D,Rowley JD, Vardiman JW. Correlation between cellmorphology and expression of the AML1/ETO chimerictranscript in patients with acute myeloid leukemiawithout the t(8;21). Leukemia 8: 1533, 1994.
38 Hurwitz CA, Raimondi SC, Head D, Krance R, Mirro JrJ, Kalwinsky DK, Ayers GD, Behm FG. Distinctiveimmunophenotypic features of t(8;21)(q22;q22) acutemyeloblastic leukemia in children. Blood 80: 3182, 1992.
39 Tallman MS, Hakimian D, Shaw JM, Lissner GS, RussellEJ, Variakojis D. Granulocytic sarcoma is associatedwith the 8;21 translocation in acute myeloid leukemia.Journal of Clinical Oncology 11: 690, 1993.
40 Nucifora G, Larson RA, Rowley JD. Persistence of the8;21 translocation in patients with acutemyeloid leukemiatype M2 in long-term remission. Blood 82: 712, 1993.
41 Preudhomme C, Philippe N, Macintyre E, Henic N, LaiJL, Jouet JP, Cosson A, Fenaux P. Persistence of AML1/ETO fusion mRNA in t(8;21) acute myeloid leukemia(AML) in prolonged remission: is there a consensus?[letter]. Leukemia 10: 186, 1996.
42 Asou H, Tashiro S, Hamamoto K, Otsuji A, Kita K,Kamada N. Establishment of a human acute myeloidleukemia cell line (Kasumi-1) with 8;21 chromosometranslocation. Blood 77: 2031, 1991.
43 Matozaki S, Nakagawa T, Kawaguchi R, Aozaki R,Tsutsumi M, Murayama T, Koizumi T, Nishimura R,Isobe T, Chihara K. Establishment of a myeloidleukaemic cell line (SKNO-1) from a patient with t(8;21)who acquired monosomy 17 during disease progression.British Journal of Haematology 89: 805, 1995.
44 Da Silva N, Meyer-Monard S, Menot ML, Degos L,Baruchel A, Dombret H, Chomienne C. G-CSF activatesSTAT pathways in Kasumi-1 myeloid leukemic cells withthe t(8;21) translocation: basis for potential therapeutice�cacy. Cytokines Cellular and Molecular Therapy 3: 75,1997.
45 Sakakura C, Yamaguchi-Iwai Y, Satake M, Bae SC,Takahashi A, Ogawa E, Hagiwara A, Takahashi T,Murakami A, Makino K, Nakagawa T, Kamada N, ItoY. Growth inhibition and induction of di�erentiation oft(8;21) acute myeloid leukemia cells by the DNA-bindingdomain of PEBP2 and the AML1/MTG8(ETO)-speci®cantisense oligonucleotide. Proceedings of the NationalAcademy of Science USA 91: 11723, 1994.
46 Ema H, Kitano K, Suda T, Sato Y, Muroi K, Ohta M,Yoshida M, Sakamoto S, Eguchi M, Miura Y. In vitrodi�erentiation of leukemic cells to eosinophils in thepresence of interleukin-5 in two cases of acute myeloidleukemia with the translocation t(8;21)(q22;q22). Blood75: 350, 1990.
47 Touw I, Donath J, Pouwels K, van Buitenen C, SchipperP, Santini V, Hagemeijer A, Lowenberg B, Delwel R.Acute myeloid leukemias with chromosomal abnormal-ities involving the 21q22 region identi®ed by their in vitroresponsiveness to interleukin-5. Leukemia 5: 687, 1991.
48 Ferrara F, Di Noto R, Viola A, Russo C, Boccuni P,Costantini S, Dello Russo A, Lo Pardo C, Del Vecchio L.Complete remission in acute myeloid leukaemia witht(8;21) following treatment with G-CSF: ¯ow cytometricanalysis of in vivo and in vitro e�ects on cell maturation.British Journal of Haematology 106: 520, 1999.
49 Calabresse C, Barbey S, Venturini L, Balitrand N, DegosL, Fenaux P, Chomienne C. In vitro treatment withretinoids or the topoisomerase inhibitor, VP-16, evi-dences di�erent functional apoptotic pathways in acutepromyelocytic leukemic cells. Leukemia 9: 2049, 1995.
50 Dombret H, Chastang C, Fenaux P, Rei�ers J,Bordessoule D, Bouabdallah R, Mandelli F, Ferrant A,Auzanneau G, Tilly H, Yver A, Degos L. A controlledstudy of recombinant human granulocyte colony-stimu-lating factor in elderly patients after treatment for acutemyelogenous leukemia. AML Cooperative Study Group.New England Journal of Medicine 332: 1678, 1995.
51 Chomienne C, Ballerini P, Balitrand N, Daniel MT,Fenaux P, Castaigne S, Degos L. All-trans retinoic acidin acute promyelocytic leukemias. II In vitro studies:structure-function relationship. Blood 76: 1710, 1990.
52 Kita K, Nakase K, Miwa H, MasuyaM, Nishii K, MoritaN, Takakura N, Otsuji A, Shirakawa S, Ueda T, Nasu K,Kyo T, Dohy H, Kamada N. Phenotypical characteristicsof acute myelocytic leukemia associated with thet(8;21)(q22;q22) chromosomal abnormality: frequentexpression of immature B-cell antigen CD19 togetherwith stem cell antigen CD34. Blood 80: 470, 1992.
53 Charrad RS, Li Y, Delpech B, Balitrand N, Clay D,Jasmin C, Chomienne C, Smadja-Jo�e F. Ligation of theCD44 adhesion molecule reverses blockage of differentia-tion in human acute myeloid leukemia. Nature Medicine5: 669, 1999.
54 Yoshida H, Kitamura K, Tanaka K, Omura S, MiyazakiT, Hachiya T, Ohno R, Naoe T. Accelerated degradationof PML-retinoic acid receptor alpha (PML-RARA)oncoprotein by all-trans-retinoic acid in acute promye-locytic leukemia: possible role of the proteasome path-way. Cancer Research 56: 2945, 1996.
55 Douer D, Koe�er HP. Retinoic acid enhances growth ofhuman early erythroid progenitor cells in vitro. Journal ofClinical Investigation 69: 1039, 1982.
56 Gratas C, Menot ML, Dresch G, Chomienne C. Retinoidacid supports granulocytic but not erythroid differentia-tion of myeloid progenitors in normal bone marrow cells.Leukemia 7: 1156, 1993.
57 Micle a J, Chomienne C. E�ect of all-trans retinoic acidon CD34+ human myeloid cells. Leukemia 8: 214, 1994.
58 Chomienne C, Balitrand N, Ballerini P, Castaigne S, deThe H, Degos L. All-trans retinoic acid modulates theretinoic acid receptor-alpha in promyelocytic cells.Journal of Clinical Investigation 88: 2150, 1991.
59 Warrell Jr RP, Frankel SR, Miller Jr WH, ScheinbergDA, Itri LM, HittelmanWN, Vyas R, Andree�M, TafuriA, Jakubowski A, Grabilove J, Gordon NS, DmitrovskyE. Di�erentiation therapy of acute promyelocyticleukemia with tretinoin (all-trans-retinoic acid). NewEngland Journal of Medicine 324: 1385, 1991.
60 Castaigne S, Chomienne C, Daniel MT, Ballerini P,Berger R, Fenaux P, Degos L. All-trans retinoic acid as adi�erentiation therapy for acute promyelocytic leukemia.I. Clinical results. Blood 76: 1704, 1990.
61 Testa U, Grignani F, Samoggia P, Zanetti C, Riccioni R,Coco FL, Diverio D, Felli N, Passerini CG, Grell M,Pelicci PG, Peschle C. The PML/RARalpha fusionprotein inhibits tumor necrosis factor-alpha-inducedapoptosis in U937 cells and acute promyelocyticleukemia blasts. Journal of Clinical Investigation 101:2278, 1998.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
327

62 de The H, Lavau C, Marchio A, Chomienne C, Degos L,Dejean A. The PML-RAR alpha fusion mRNAgenerated by the t(15;17) translocation in acutepromyelocytic leukemia encodes a functionally alteredRAR. Cell 66: 675, 1991.
63 Kakizuka A, Miller Jr WH, Umesono K, Warrell Jr RP,Frankel SR, Murty VV, Dmitrovsky E, Evans RM.Chromosomal translocation t(15;17) in human acutepromyelocytic leukemia fuses RAR alpha with a novelputative transcription factor, PML. Cell 66: 663, 1991.
64 Grignani F, Ferrucci PF, Testa U, Talamo G, Fagioli M,Alcalay M, Mencarelli A, Grignani F, Peschle C,Nicoletti I, Pelicci P-G. The acute promyelocyticleukemia-speci®c PML-RAR alpha fusion proteininhibits di�erentiation and promotes survival of myeloidprecursor cells. Cell 74: 423, 1993.
65 Rousselot P, Hardas B, Patel A, Guidez F, Gaken J,Castaigne S, Dejean A, de The H, Degos L, Farzaneh F,Chomienne C. The PML-RAR alpha gene product of thet(15;17) translocation inhibits retinoic acid-inducedgranulocytic di�erentiation and mediated transactiva-tion in human myeloid cells. Oncogene 9: 545, 1994.
66 Miyoshi H, Ohki M, Nakagawa T, Honma Y.Glucocorticoids induce apoptosis in acute myeloidleukemia cell lines with A t(8;21) chromosome transloca-tion. Leukemia Research 21: 45, 1997.
67 Recher C, Larghero J, Rousselot P, Da Silva N, Meyer S,Mouas H, Micle a J, Chomienne C, Dombret H, Degos L.Dexamethasone induces in vitro and in vivo apoptosis ofleukemic cells in a patient with the t(8;21) translocation.Blood 90 (10) (Suppl 1): 245b, 1997.
68 Tsuchiya H, ElSonbaty SS, Nagano K, Watanabe M,Migita M, Mitsubuchi H, Kaneko Y, Matsuda I. Acutemyeloblastic leukemia (ANLL-M2) with t(8;21)(q22;q22)variant expressing lymphoid but not myeloid surfaceantigens with a high number of G-CSF receptors.Leukemia Research 17: 375, 1993.
69 De Botton S, Dombret H, Sanz M, Miguel JS, Caillot D,Zittoun R, Gardembas M, Stamatoulas A, Conde E,Guerci A, Gardin C, Geiser K, Makhoul DC, Reman O,de la Serna J, Lefrere F, Chomienne C, Chastang C,Degos L, Fenaux P. Incidence, clinical features, andoutcome of all trans-retinoic acid syndrome in 413 casesof newly diagnosed acute promyelocytic leukemia. TheEuropean APL Group. Blood 92: 2712, 1998.
70 Estey E. Hematopoietic growth factors in the treatmentof acute leukemia. Current Opinion in Oncology 10: 23,1998.
71 Chakraborty A, White SM, Schaefer TS, Ball ED, DyerKF, Tweardy DJ. Granulocyte colony-stimulating factoractivation of Stat3 alpha and Stat3 beta in immaturenormal and leukemic human myeloid cells. Blood 88:2442, 1996.
The Hematology Journal
G-CSF pathways in t(8;21) leukemiaN Da Silva et al
328





























