Embed Size (px)
Citation preview

https://tvuni.academia.edu/mvinayagam Page 0
V. MAGENDIRA MANI
Assistant Professor of Biochemistry
Islamiah College (Autonomous)
Vaniyambadi
Vellore District
Mail: [email protected]
Download Science at: [email protected]
https://tvuni.academia.edu/mvinayagam

https://tvuni.academia.edu/mvinayagam Page 1
GENE EXPRESSION AND ITS REGULATION
Regulation of gene expression includes a wide range of mechanisms that are used by cells to
increase or decrease the production of specific gene products (protein or RNA), and is informally
termed gene regulation. Gene regulation is essential for viruses, prokaryotes and eukaryotes as it
increases the versatility and adaptability of an organism by allowing the cell to express protein
when needed.
Regulated stages of gene expression
Any step of gene expression may be modulated, from the DNA-RNA transcription step to post-
translational modification of a protein. The following is a list of stages where gene expression is
regulated; the most extensively utilized point is Transcription Initiation:
Chromatin domains
Transcription
Post-transcriptional modification
Translation
Modification of DNA - Chromatin domains
In eukaryotes, the accessibility of large regions of DNA can depend on its chromatin structure,
which can be altered as a result of histone modifications directed by DNA methylation, ncRNA,
or DNA-binding protein. Hence these modifications may up or down regulate the expression of a
gene. Some of these modifications that regulate gene expression are inheritable and are referred
to as epigenetic regulation.
Structural
Transcription of DNA is dictated by its structure. In general, the density of its packing is
indicative of the frequency of transcription. Octameric protein complexes called nucleosomes are
responsible for the amount of supercoiling of DNA, and these complexes can be temporarily
modified by processes such as phosphorylation or more permanently modified by processes such
as methylation. Such modifications are considered to be responsible for more or less permanent
changes in gene expression levels.

https://tvuni.academia.edu/mvinayagam Page 2
Chemical
Methylation of DNA is a common method of gene silencing. DNA is typically methylated by
methyltransferase enzymes on cytosine nucleotides in a CpG dinucleotide sequence.
Histone acetylation is also an important process in transcription. Histone acetyltransferase
enzymes (HATs) such as CREB-binding protein also dissociate the DNA from the histone
complex, allowing transcription to proceed. Often, DNA methylation and histone deacetylation
work together in gene silencing. The combination of the two seems to be a signal for DNA to be
packed more densely, lowering gene expression.
Regulation of transcription
Regulation of transcription thus controls when transcription occurs and how much RNA is
created. Transcription of a gene by RNA polymerase can be regulated by at least five
mechanisms:
Specificity factors alter the specificity of RNA polymerase for a given promoter or
set of promoters, making it more or less likely to bind to them (i.e., sigma factors
used in prokaryotic transcription).
Repressors bind to the Operator, coding sequences on the DNA strand that are close
to or overlapping the promoter region, impeding RNA polymerase's progress along
the strand, thus impeding the expression of the gene.
General transcription factors position RNA polymerase at the start of a protein-
coding sequence and then release the polymerase to transcribe the mRNA.
Activators enhance the interaction between RNA polymerase and a particular
promoter, encouraging the expression of the gene. Activators do this by increasing the
attraction of RNA polymerase for the promoter, through interactions with subunits of
the RNA polymerase or indirectly by changing the structure of the DNA.
Enhancers are sites on the DNA helix that are bound by activators in order to loop
the DNA bringing a specific promoter to the initiation complex. Enhancers are much
more common in eukaryotes than prokaryotes, where only a few examples exist.
Silencers are regions of DNA sequences that, when bound by particular transcription
factors, can silence expression of the gene

https://tvuni.academia.edu/mvinayagam Page 3
Post-transcriptional regulation
After the DNA is transcribed and mRNA is formed, there must be some sort of regulation on
how much the mRNA is translated into proteins. Cells do this by modulating the capping,
splicing, addition of a Poly (A) Tail, the sequence-specific nuclear export rates, and, in several
contexts, sequestration of the RNA transcript. These processes occur in eukaryotes but not in
prokaryotes. This modulation is a result of a protein or transcript that, in turn, is regulated and
may have an affinity for certain sequences.
Regulation of translation
The translation of mRNA can also be controlled by a number of mechanisms, mostly at the level
of initiation. Recruitment of the small ribosomal subunit can indeed be modulated by mRNA
secondary structure, antisense RNA binding, or protein binding. In both prokaryotes and
eukaryotes, a large number of RNA binding proteins exist, which often are directed to their
target sequence by the secondary structure of the transcript, which may change depending on
certain conditions, such as temperature or presence of a ligand. Some transcripts act as
ribozymes and self-regulate their expression.

https://tvuni.academia.edu/mvinayagam Page 4
MOLECULAR MECHANISM OF REGULATION OF PROKARYOTIC OPERON
MODEL
Operons
Control of Gene Activity in Prokaryotic Cells
The activity of genes is controlled by the cell and the environment.
A. Inducible genes are inactive unless circumstances cause them to be activated
(“turned on”).
B. Repressible genes are active unless circumstances cause them to be inactivated
(“turned off”).
C. Constitutive gene functions are active continually, with no control exerted. This is
generally an abnormal situation.
In prokaryotic cells (and viruses) the control of gene activity is often in the form of operons.
D. Operons are a form of transcriptional control.
E. An operon consists of the structural gene (or genes) which actually code for specific
proteins and the controlling elements associated with the control of those genes. An
operon typically contains several genes, all under the same control mechanism.
F. Though rather similar controlling systems have been found for some eukaryotic
genes, control mechanisms in eukaryotes are generally more diverse and more
complex, and except for a few examples in simple eukaryotic organisms like yeasts,
multiple genes are not found to function under a single control mechanism. In other
words, eukaryotic cells do not have operons.
The first operon investigated was the lac operon in E. coli. This work came from Jacob and
Monod (1959 Journal of Molecular Biology V. 1).
G. The primary carbohydrate source for the cell is the sugar glucose, but there are a
large number of sugars which can be used if there isn’t enough glucose available to
support the energy needs of the cell. Sugars like lactose are “backup” carbohydrates.
This means that the cell only metabolizes lactose if glucose is low and lactose is
plentiful.

https://tvuni.academia.edu/mvinayagam Page 5
LAC OPERON
Lactose -----------> Glucose + Galactose
β-galactosidase
E.Coli cells grown in the absence of lactose contain an average number of 1 to 5 molecules of
β-galactosidase per cell, whereas bacteria grown in the presence of an excess of lactose or certain
lactose analogue contain 1000 to 10,000 β-galactosidase molecules per cell.
E.Coli can use lactose as a carbon sources, the enzymes required for the use of lactose as a
carbon source are only synthesized, when lactose is available as the sole carbon source. This can
be explained by Lac operon.
The cell uses a negative control system (the lac operon) to respond to the availability of lactose
in the environment. Negative control means that it’s a system in which the active substance acts
to turn off function. It is very wasteful to manufacture the enzymes needed to metabolize lactose
if there is no lactose to be used, so under normal conditions the genes for producing the enzymes
for metabolizing lactose need to be inactive. They should be activated only when lactose is
plentiful. So the purpose of the operon is to keep these genes turned off if there’s no lactose
around, and to turn them on if lactose is plentiful.
The lac operon contains three genes which code for enzymes necessary for the metabolism of the
sugar lactose. These three genes are side by side on the DNA molecule, and they are transcribed
as a single, polycistronic message. The operon consists of a promoter, an operator, the structural
genes, a termination sequence, and a repressor gene.

https://tvuni.academia.edu/mvinayagam Page 6
Structure of the lac operon
The lac operon consists of three structural genes, a promoter, a terminator, regulator and
an operator.
The three structural genes
are: lacZ, lacY, and lacA.
lacZ encodes β-galactosidase (LacZ), an
intracellular enzyme that cleaves
the disaccharide lactose into glucose and
galactose.
lacY encodes lactose permease (LacY), a transmembrane symporter that pumps β-
galactosides into the cell using a proton gradient in the same direction.
lacA encodes galactoside O-acetyl transferase (Lac A), an enzyme that transfers an acetyl
group from acetyl-CoA to β-galactosides. Only lacZ and lacY appear to be necessary for
lactose catabolism. All of the genes involved in controlling this pathway are located next to each
other on the E. coli chromosome. Together they form an operon.
There is a single promoter region which precedes the lac z gene (the first gene in the sequence).
The promoter region is where the RNA polymerase binds for transcription.
The promoter sequence of the lac operon is TATGTT at the -10 region; and TTTACA at the -35
region.
lac Operon Gene Gene Function
I Gene for repressor protein
P Promoter
O Operator
lac Z Gene for β-galactosidase
lac Y Gene for β-galactoside permease
lac A Gene for β-galactoside transacetylase

https://tvuni.academia.edu/mvinayagam Page 7
Between the Promoter and the lac z gene is a region called the Operator. This is the primary
control site for the operon.
Element purpose
Operator (Lac O) binding site for repressor
Promoter (Lac P) binding site for RNA polymerase
Repressor (Lac I) gene encoding lac repressor protein
Binds to DNA at operator and blocks binding of RNA polymerase at promoter
Pi promoter for Lac I
CAP binding site for cAMP/CAP complex
The repressor gene (lac I) is not contiguous with the other portions of the operon. The repressor
codes for the production of a diffusible repressor protein. So our entire operon could be
diagrammed like this:
Lac I is the name for the repressor gene; P is the promoter, O the operator, and T the termination
sequence. Note that the prokaryotic genome is a circular DNA molecule, and we are viewing just
one segment of that circle. Also note that the normal condition for a prokaryotic cell is to have a
single copy of its genome in the cell.

https://tvuni.academia.edu/mvinayagam Page 8
5. When glucose is plentiful and/or lactose is low, the desired situation is for these genes to be
repressed (inactive).
6. The repressor gene codes for the production of a diffusible repressor protein, which is present
in low numbers in the cell at all times. The repressor has an affinity for the operator of the lac
operon, and unless something happens to intervene, the repressor molecule will bind to the O site
and block the movement of an RNA polymerase from the promoter to the structural genes, thus
preventing transcription of those genes.
7. When lactose is plentiful (and glucose is not) the repression needs to be removed so the genes
can be activated. The trigger for this activation needs to be lactose. Besides being attracted to the
operator sequence of the operon, the repressor protein also has an affinity for a slightly unusual
form of lactose called allolactose, which will be present as a small percentage of the lactose
available whenever there’s lactose around. When the repressor binds to allolactose, the
configuration (three-dimensional shape) of the repressor is altered, and it is no longer able to
bind to the operator. It falls off, and there is nothing to prevent RNA polymerase from reading
through the operator to the structural genes, and the three enzymes will thus be made.

https://tvuni.academia.edu/mvinayagam Page 9
8. As the cell metabolizes the available lactose, eventually the concentration of lactose will fall
far enough that there won’t be enough around to maintain the complexes with the available
repressor proteins, the repressor will return to its former configuration, and will once again bind
to the operator and repress the structural genes.
C. There are actually two conditions which must be met if the cell needs these enzymes to be
produced. Not only must lactose be plentiful, but glucose must be low. The cell responds to
glucose availability through a positive control mechanism involving cyclic AMP. Positive
control means that it’s a system in which the active substance acts to turn on function.
1. When abundant glucose is available, it isn’t efficient for the cell to metabolize lactose, even if
it is plentiful, because it takes more energy to metabolize lactose than to metabolize glucose. So
the lac operon has a second control system which keeps the operon turned off when glucose is
abundant.
2. This repression is called catabolite repression, and involves a protein called catabolite-
activating protein (CAP).
3. CAP has an affinity for the promoter region of the lac operon, and unless CAP is bound to that
region, RNA polymerase will not bind to the promoter, and transcription will not occur.
(Contrast this to the situation with the lac repressor. The repressor prevents transcription when
bound to the Operator site; CAP allows transcription when bound to the operator site. This is
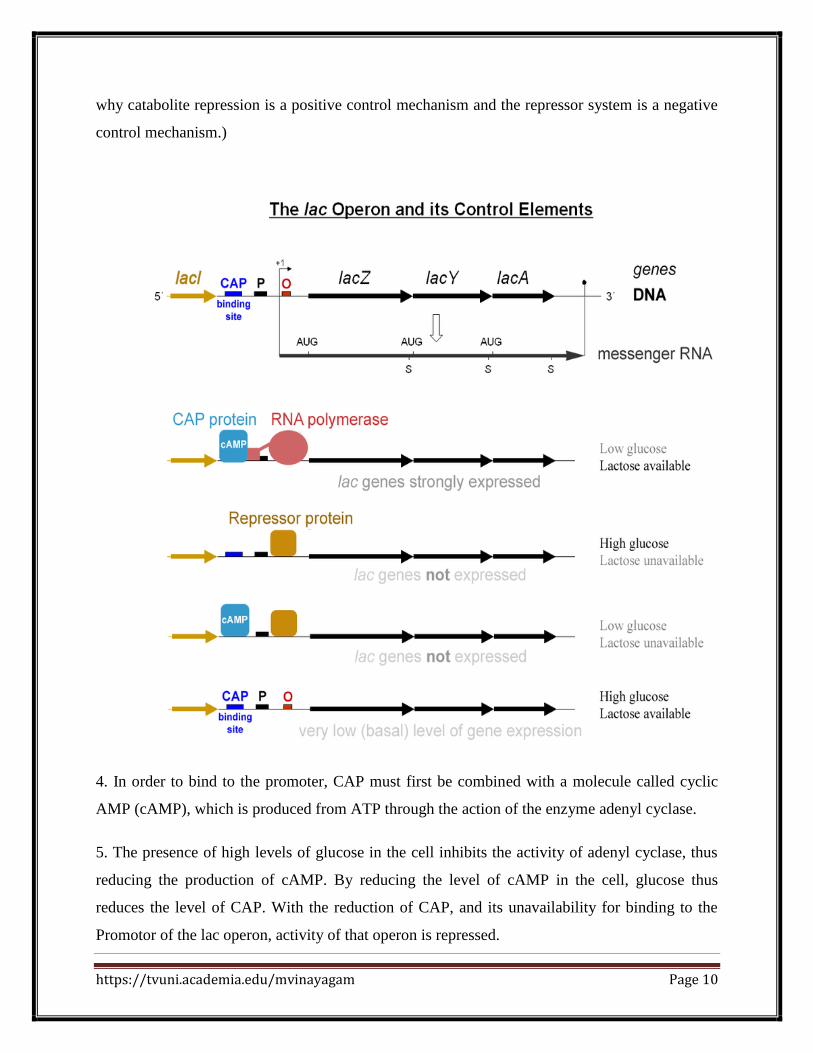
https://tvuni.academia.edu/mvinayagam Page 10
why catabolite repression is a positive control mechanism and the repressor system is a negative
control mechanism.)
4. In order to bind to the promoter, CAP must first be combined with a molecule called cyclic
AMP (cAMP), which is produced from ATP through the action of the enzyme adenyl cyclase.
5. The presence of high levels of glucose in the cell inhibits the activity of adenyl cyclase, thus
reducing the production of cAMP. By reducing the level of cAMP in the cell, glucose thus
reduces the level of CAP. With the reduction of CAP, and its unavailability for binding to the
Promotor of the lac operon, activity of that operon is repressed.

https://tvuni.academia.edu/mvinayagam Page 11
D. So these three lactose-metabolysis genes are under dual control.
1. The CAP system prevents lac operon activity when glucose is plentiful because high glucose
levels lead to reduced CAP availability, and CAP is necessary for this operon to function.
2. The repressor system prevents lac operon activity when lactose is not available because the
repressor protein binds to the Operator site and prevents transcription. The repression can only
be removed when lactose is high because allolactose is necessary to inactivate the repressor.
3. The combination of these two control mechanisms ensures that these enzymes will be
produced only under conditions in which glucose is low and lactose is high.
E. In low glucose conditions (when the only control mechanism functioning is the repressor
system), mutations in the various components of the operon have been studied.

https://tvuni.academia.edu/mvinayagam Page 12
1. Some mutations in the I gene (which codes for the repressor) produced an altered protein
which is unable to bind to the Operator. These mutations would result in constitutive gene
activity.
2. There are also I gene mutations which produce a repressor which is unable to bind to
allolactose. These mutations produce cells which are unable to activate the lac operon genes, and
which can therefore not metabolize lactose, even though the structural genes themselves may be
completely fine.
3. There are also mutations which alter the sequence of the Operator region such that the
repressor molecule is no longer able to bind. These would be constitutive for all of the genes in
the operon.
TRYPTOPHAN (TRP) OPERON
The tryptophan (trp) operon in E. coli is a negative control repressible system. It’s negative
control because the system produces a repressor which functions to turn off the operon. It’s
repressible because, unlike the lac operon, repression occurs when a critical substance is
abundant in the cell. (For the trp operon, that critical substance is the amino acid tryptophan; for
the lac operon the critical substance is lactose, and its presence removes repression.)
Tryptophan is an important amino acid which most E. coli can acquire in two ways. They can
extract it from the materials the cell consumes, or they can manufacture it themselves. The genes
controlled by the trp operon produce enzymes which are necessary for the cell to produce its own
tryptophan. Again, contrast to the lac operon. The lac operon enzymes function in the digestion
(catabolism) of a “food” molecule (lactose); the trp operon enzymes function in the
manufacture (anabolism) of a necessary amino acid for the construction of the cell’s own
proteins. The lac operon needs to be active when there’s lots of lactose around; the trp operon
needs to be active when tryptophan levels in the cell are low.

https://tvuni.academia.edu/mvinayagam Page 13
The components of the trp operon are basically the same as those for the lac operon.
This operon controls five structural genes which code for enzymes needed for the production of
tryptophan, including the key enzyme tryptophan synthetase. These genes are called trp E, trp D,
trp C, trp B and trp A.
In the trp operon, the repressor gene is named trp R.
The repressor gene (trp R) codes for a repressor protein. Unlike the lac repressor, this one is
inactive as it is produced by the cell. It requires a co-repressor in order to bind to the Operator.
trp Operon Gene Gene Function
P/O Promoter; operator sequence is found in the promoter
trp L Leader sequence; attenuator (A) sequence is found in the leader
trp E Gene for anthranilate synthetase subunit1
trp D Gene for anthranilate synthetase subunit2
trp C Gene for glycerolphosphate synthetase
trp B Gene for tryptophan synthetase subunit1
trp A Gene for tryptophan synthetase subunit2
The co-repressor is the amino acid tryptophan. Thus, when tryptophan is abundant, the repressor
is active and will bind to the Operator, preventing transcription of these genes. But if tryptophan
levels fall, the repressor will lose its trp co-repressor and will fall off the Operator, and the genes
will be transcribed and the enzymes constructed. This leads to exactly the control needed—if
there’s a lot of tryptophan around, the cell doesn’t want to make more, and it would be a waste of
energy and materials to have this set of genes active. It’s when tryptophan levels fall that the cell
needs to manufacture more, and needs this operon to be active.

https://tvuni.academia.edu/mvinayagam Page 14
When tryptophan is not present in the cell, the repressor by itself does not bind to the operator;
therefore, the operon is active and tryptophan is synthesized. Because the repressor protein
actively binds to the operator to keep the genes turned off, the trp operon is negatively regulated
and the proteins that bind to the operator to silence trp expression are negative regulators.
While the lac operon can be activated by a chemical (allolactose), the tryptophan (Trp) operon is
inhibited by a chemical (tryptophan). This operon contains five structural genes: trp E, trp D, trp
C, trp B, and trp A, which encode tryptophan synthetase. It also contains a repressive regulator
gene called trp R. Trp R has a promoter where RNA polymerase binds and synthesizes mRNA
for a regulatory protein. The protein that is synthesized by trp R then binds to the operator which
then causes the transcription to be blocked. In the lac operon, allolactose binds to the repressor
protein, allowing gene transcription, while in the trp operon; tryptophan binds to the repressor
protein effectively blocking gene transcription. In both situations, repression is that of RNA
polymerase transcribing the genes in the operon. Also unlike the lac operon, the trp operon
contains a leader peptide and an attenuator sequence which allows for graded regulation.

https://tvuni.academia.edu/mvinayagam Page 15
The attenuator mechanism of Trp operon
The attenuator mechanism uses the 5’ end of the leader sequence (trpL) of the transcript. The
leader is about 140 ntds long; it has four segments of complementary sequences, call 1, 2, 3 and
4. These sequences facilitate partial base pairing between 1 and 2, 2 and 3 and 3 and 4 to
generate two stem loop structures.
The 4th segment when pairs with the 3rd, it forms a stem loop similar the rho independent
transcriptional terminator structure with terminal Us.
In the presence of Trp, formation of this structure prevents the transcription initiated by the
RNAP to proceed further and the transcription is terminated. So this is a fail-proof operation.

https://tvuni.academia.edu/mvinayagam Page 16
When Trp is absent, the leader sequence can still generate this chain termination stem loop
structure and terminate transcription, but transcription of the full-length transcript is required for
Tryptophan synthesis. In order to produce full-length transcript the terminator stem-loop
formation should be prevented by change in base pairing alignments among the four blocks.
When tryptophan levels are high, the
ribosome quickly translates sequence 1 (open
reading frame encoding leader peptide) and
blocks sequence 2 before sequence 3 is
transcribed. Continued transcription leads to
attenuation at the terminator-like structure
formed by sequences 3 and 4.
When tryptophan levels are low, the ribosome
pauses at the Trp codons in sequence l.
Formation of the paired structure between
sequences 2 and 3 prevents attenuation
because sequence 3 is no longer available to form the attenuator structure with sequence 4.

https://tvuni.academia.edu/mvinayagam Page 17
The trp operon attenuation mechanism uses signals encoded in four sequences within a 162
nucleotide leader region at the 5' end of the mRNA that precedes the initiation codon of the first
gene. At the end of the leader is a sequence called the attenuator, made up of sequences 3 and 4.
Sequences 3 and 4 base-pair to form a G=C rich stem and loop structure followed by a series of
uridylate residues, a structure that resembles a transcription terminator; transcription will halt
here when this structure forms.
C - VALUE PARADOX
C value - the amount of DNA per haploid cell (usually expressed as picograms) or the number of
kilobases per haploid cell.
Human C-values
The Human genome varies in size; however, the current estimate of the nuclear haploid size of
the reference human genome is 3,031,042,417 bp for the X gamete and 2,932,228,937 bp for the

https://tvuni.academia.edu/mvinayagam Page 18
Y gamete. (The X gamete size is larger because the X chromosome is larger than the Y. Added
together, the XX female diploid genome size is larger than the XY male.)
The human genome's GC content is about 41% Accounting for the autosomal, X, and Y
chromosomes, human haploid GC contents are 40.97460% for X gametes, and 41.01724% for Y
gametes.
REPETITIVE DNA
Eukaryote and also human DNA contains large portion of noncoding sequences. As for the
coding DNA, the noncoding DNA may be unique or in more identical or similar copies. DNA
sequences with high copy numbers are then called repetitive sequences. If the copies of a
sequence motif lie adjacent to each other in a block, or an array, we are speaking about tandem
repeats, the repetitive sequences dispersed throughout the genome as single units flanked by
unique sequence are interspersed repeats.
Single copy DNA sequences (60% of total)
Present in single or low copy numbers.
Includes coding sequence for structural genes (up to 1400 bp in size), which account for 3% of
the genome.
Moderately repetitive DNA sequences (30% of total)
Present at between 10 - 105
copies per genome. Found throughout the euchromatin.
Average 300bp in size
May be classed as :-
a) microsatellites / minisatellites (VNTR, DNA 'fingerprints)
b) dispersed-repetitive DNA, mainly transposable elements (LINES/ SINES)
Many moderately-repetitive sequences may be involved in regulation of gene expression. This is
supported by their interspersion with single-copy sequences and location adjacent to structural
genes.

https://tvuni.academia.edu/mvinayagam Page 19
Highly repetitive DNA sequences (satellite DNA) (10% of total)
Present at >106
copies per genome
Occurs as variable length motifs (5-100 bp), in long tracts of up to 100 Mb
Most is located in heterochromatic regions around the centromere / telomere.
Postulated functions include structural or organizational roles, role in chromosome pairing,
involvement in cross-over or recombination, junk.
eg. alpha-satellite DNA
This is a highly repetitive sequence, each centromere contains a tandem array of alpha-satellite
repeats that extend for millions of base pairs and are arranged in a hierarchy of higher order
repeats. These vary between 100-5000 on different chromosomes (0.2-10Mb). Some contain
17bp binding sites for the centromere-specific DNA binding protein CENP-B. They have been
recently cloned and used to construct artificial human chromosomes.
We can classify human repetitive
DNA as shown below. The highly
repetitive fraction is satellite DNA
that makes up most of the
heterochromatin of the human
genome. The middle repetitive
fraction consists of both tandem
repeats and interspered repeats. The
tandem repeats include multiple
copy genes found in tandem arrays
(rRNA and histone genes),
minisatellites like the VNTRs or STRs used in forensic DNA typing, and microsatellites that are
runs of dinucleotide repeats. The interspersed repeats consist mostly of retrotransposons (SINEs
and LINEs) and a small fraction of DNA transposons.

https://tvuni.academia.edu/mvinayagam Page 20
GENE DOSAGE
Gene dosage is the number of copies of a gene present in a cell or nucleus. An increase in gene
dosage can cause higher levels of gene product if the gene is not subject to regulation from
elsewhere in the body. Gene dosage is also known to be significantly associated with other
quantitative or qualitative phenotype traits.
Gene dosage refers to the number of copies of a gene present in an organism's genome, or
complete "library" of genetic information. Many organisms, including humans, store genetic
information on paired chromosomes. Each member of a pair of chromosomes contributes a
"single dose" of the genes contained on that chromosome. Sex chromosomes, however, tend to
differ between males and females; human males have a single X chromosome and a single Y
chromosome while human females have a pair of X chromosomes. Various regulatory processes
known as "dosage compensation" are in place to ensure that gene dosage remains at proper levels
in both males and females despite the genetic imbalance caused by different genes.
Generally speaking, more copies of a gene — or
higher gene dosage — will result in increased
expression of the proteins for which the genes code.
To a significant extent, however, the genes on male
and female sex chromosomes are expressed at
comparable levels despite the difference in gene
dosage. If this were not the case, females with their
two X chromosomes could overexpress certain
genes, or males with their single X and single Y
chromosomes could underexpress certain genes.
Either of these alternatives could cause severe
mutations or death, so it is important that the genes
are expressed at comparable levels in spite of the
difference in gene dosage.
Each member of a pair of chromosomes contributes
a "single dose" of the genes contained on that
chromosome.

https://tvuni.academia.edu/mvinayagam Page 21
A set of regulatory mechanisms and processes known as dosage compensation are responsible
for maintaining the expression of genes at appropriate levels. Different organisms have different
means of regulating the expression of their genes, and some even make use of multiple methods
of dosage compensation. Gene expression in human females is regulated through X-inactivation,
through which one of the female's two X chromosomes becomes an inactive "Barr body." The
result of X-inactivation is that males and females each only have a single X-chromosome that is
actually expressing its genetic information and contributing to gene dosage.
In some organisms, such as the fruit fly or Drosophila melanogaster, the expression of genes on
the male X chromosome is doubled to match the gene dosage of the female's two X
chromosomes. The roundworm, or C. elegans, presents an interesting case, as it exists most
commonly as a hermaphrodite with two X chromosomes, though some have only a single sex
chromosome, X, and are classified as male. Dosage compensation in C. elegans results in the
partial repression of the expression of genes on both of the X chromosomes in hermaphrodites.
GENE AMPLIFICATION
A selective increase in the number of copies of a gene coding for a specific protein without a
proportional increase in other genes is called gene amplification. It occurs naturally via the
excision of a copy of the repeating sequence from the chromosome and its extrachromosomal
replication in a plasmid, or via the production of an RNA transcript of the entire repeating
sequence of ribosomal RNA followed by the reverse transcription of the molecule to produce an
additional copy of the original DNA sequence. Laboratory techniques have been introduced for
inducing disproportional replication by unequal crossing over, uptake of DNA from lysed cells,
or generation of extrachromosomal sequences from rolling circle replication.
Gene amplification is an increase in the number of copies of a gene sequence. Cancer cells
sometimes produce multiple copies of genes in response to signals from other cells or their
environment. The term also can refer to polymerase chain reaction (PCR), a laboratory technique
that is used by scientists to amplify gene sequences in a test tube.





























