Embed Size (px)
Citation preview

Plant Pathology
(2006)
55
, 639–649 Doi: 10.1111/j.1365-3059.2006.01437.x
© 2006 The Authors Journal compilation © 2006 BSPP
639
Blackwell Publishing Ltd
Genetic, phenotypic and pathogenic diversity among xanthomonads isolated from pistachio (
Pistacia vera
) in Australia
A. Marefat
a
*, E. S. Scott
a
, K. Ophel-Keller
b
and M. Sedgley
c
a
School of Agriculture and Wine, University of Adelaide, Waite Campus, PMB 1, Glen Osmond, South Australia 5064;
b
South Australian Research and Development Institute, GPO Box 397, Adelaide, South Australia 5001; and
c
Faculty of the Sciences, University of New England, Armidale, New South Wales 2351, Australia
Repetitive extragenic palindromic polymerase chain reaction (rep-PCR), sequencing of the 16S
−
23S rDNA internaltranscribed spacer (ITS), biochemical and physiological tests, the Biolog microplate system, polyacrylamide gel electro-phoresis (PAGE) of whole-cell proteins, and pathogenicity tests were used to characterize variability among xanthomonadsisolated from pistachio trees suffering from bacterial dieback in four regions of Australia. ITS sequencing and rep-PCRrevealed two distinct genotypes among the strains. The ITS sequencing suggested that the pistachio strains were closelyrelated to
Xanthomonas translucens
pathovars, in particular
X. translucens
pv
. poae
. Results of physiological andbiochemical tests, as well as Biolog microplate analysis and protein profiling, confirmed the existence of two groups.Furthermore, pathogenicity and host-range studies indicated that the two groups were biologically different. There wasan association between the two groups and the geographical origin of the strains.
Keywords
: 16S
−
23S rDNA sequencing, Biolog microplate, pathovars, repetitive PCR, SDS–PAGE,
Xanthomonas translucens
Introduction
Commercial planting of pistachio (
Pistacia vera
) in Aus-tralia began in the early 1980s, using the scion cv. Sirora,selected from Red Aleppo, a Syrian cultivar that wasimported as seed from California (Maggs, 1982). Whenthe trees bore fruit in commercial quantities, at about 6–7 years old, dieback became evident. This disease, causedby
Xanthomonas
genotypes, was first observed in 1992(Edwards & Taylor, 1998) and has since been reported inmost pistachio-growing regions of Australia. Symptomson mature trees include (i) shoot death and dieback;(ii) limb and trunk lesions; (iii) excessive resinous exudate;and (iv) discoloration of woody tissue in shoots >2 yearsold (Edwards & Taylor, 1998; Facelli
et al
., 2002). Inpreliminary studies,
Xanthomonas
strains similar to
Xan-thomonas translucens
were identified as the causal agentof dieback of pistachio in Australia, and the pathogenicityof selected strains to young pistachio trees was partiallydemonstrated in that bacteria injected into the trunkcolonized the wood and caused extensive discoloration
(Facelli
et al
., 2002, 2005). However, little was knownabout the phenotypic, genetic and pathogenic diversity ofthe pathogen.
Historically, DNA-independent methods such as phys-iological and biochemical tests and protein profiling havebeen used for characterization and study of phenotypicvariation among xanthomonads (Vauterin
et al
., 1991a,1991b; Verniere
et al
., 1993). Techniques that assessvariation in genomic DNA have since provided more reli-able, reproducible and robust tools to evaluate variability(Louws
et al
., 1999). DNA fingerprinting based on thepresence of repetitive elements dispersed in bacterialgenomes (rep-PCR) (Louws
et al
., 1994) has proved asensitive and reliable technique to assess the diversity ofxanthomonads at the species, subspecies and strain levels(Louws
et al
., 1995; Lopes
et al
., 2001; Scortichini
et al
.,2001; Scortichini & Rossi, 2003; Mkandawire
et al
., 2004).In addition, sequencing of the rDNA fragment has shownpotential for estimating genetic diversity within and among
Xanthomonas
species (Cubero & Graham, 2002;Goncalves & Rosato, 2002; Sahin
et al
., 2003).The evaluation of genetic diversity within a plant path-
ogen population improves understanding of its taxonomy,epidemiology and diagnosis (Milgroom & Fry, 1997). Thisstudy was initiated to (i) assess the genetic diversity of 65
*E-mail: [email protected]
Accepted 28 February 2006
Plant Pathology
(2006)
55
, 639–649
640
A. Marefat
et al.
strains of xanthomonads obtained from pistachio trees indifferent geographical regions of Australia using rep-PCR;and (ii) verify the relatedness of the strains by comparison oftheir characteristics in SDS–PAGE profiling, physiologicaland biological tests, pathogenicity tests and ITS sequences.
Materials and methods
Collection of plant material
Symptoms of pistachio dieback have been observed in fourregions of Australia: Renmark in eastern South Australia;Kyalite in southern New South Wales; and Red Cliffs andRobinvale in north-west Victoria. A collection was estab-lished comprising 30 xanthomonad strains obtained fromdiseased pistachio trees at the four locations in 2000 and35 strains obtained from Kyalite and Robinvale in 2003.The latter strains were collected to assess possible variationwithin the pathogen population after several years, andfor this purpose one orchard was selected at each of thetwo locations, in which 15–20 trees with dieback weresampled and five 2-year-old shoots with internal stainingwere collected from each tree.
Bacterial strains
Shoots were rinsed with tap water, dipped in 70% ethanolfor 50–60 s, then rinsed three times in sterile distilled water.Segments of 4
×
4 mm were cut from woody tissue of eachsample and soaked in a few drops of sterile 0·85% NaClfor 15 min at room temperature (
c
. 22
°
C). Two loopfulsof the resulting suspension were then streaked on nutrientagar (NA) and yeast dextrose carbonate (YDC) agar (Schaad
et al
., 2001). Plates were incubated at 28
°
C and inspectedfor pale yellowish colonies from 2 days onwards, and suchcolonies were purified on YDC. Pure cultures were stored at
−
70
°
C in nutrient broth containing 30% glycerol (Schaad
et al
., 2001).
Xanthomonas translucens
pv.
translucens
DAR 35705, isolated from wheat in Australia, the onlyrepresentative of this species available in Australia at thetime, was obtained from the Australian Collection of Plant
Pathogenic Bacteria (ACPPB), Agricultural Institute,Orange, NSW, Australia.
Strains that had been collected in 2000 and stored at
−
70
°
C or freeze-dried were revived on YDC at 28
°
C, thengrown on NA and YDC at 28
°
C for 3 days for use in sub-sequent tests. The strains used in this study are shown inTable 1. Strains 7 and 25, representative of groups A andB based on all tests described below, have been depositedin the International Collection of Micro-organisms fromPlants, Auckland, New Zealand, under accession numbersICMP 16316 and ICMP 16317, respectively.
Biochemical and physiological tests
Nineteen strains selected randomly from the pathogencollection, strains 7 and 25, and
X. translucens
pv.
translu-cens
DAR 35705 were tested for Gram reaction using theKOH test, oxidase reaction, oxidative and fermentativemetabolism of glucose, gelatin liquefaction, starch hydro-lysis, H
2
S production from cysteine, nitrate reduction,urease production, growth in 5% NaCl, digestion of milkproteins, hypersensitive reaction on tobacco, action inlitmus milk, and mucoid growth on sucrose peptone agar(SPA), glucose yeast-extract carbonate agar (GYCA) andYDC, as described by Schaad
et al
. (2001). Growth at36
°
C and Tween 80 hydrolysis were assessed as describedby Fahy & Hayward (1983), whereas ice-nucleationactivity and aesculin hydrolysis were assessed accordingto Lindow (1990) and Sands (1990), respectively.
SDS–PAGE of whole-cell proteins
Sample preparation for sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS–PAGE) of solublewhole-cell protein extracts was performed for all strainslisted in Table 1, by the method of Laemmli (1970) asdescribed by Vauterin
et al
. (1991a). Discontinuous, one-dimensional SDS–PAGE was performed on a BRL verticalelectrophoresis system (model V16; Bethesda ResearchLaboratories). The gel was cast and electrophoresisperformed as per the manufacturer’s instructions. The
Table 1 List of Xanthomonas strains studied and their origins
Strain Location of origina Year isolated Source Groupb
2, 3, 4, 5, 6, 7, 8, 9, 10, 20 Kyalite, NSW 2000 NT99004c A21, 22, 23, 24, 25, 26, 27, 36 Robinvale, Vic 2000 NT99004 B11, 13, 14, 15, 16, Red Cliffs, Vic 2000 NT99004 A28, 29, 30, 31, 32, 33, 34 Renmark, SA 2000 NT99004 A72, 73, 74, 75, 76, 77, 78 Kyalite, NSW 2003 This study A44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71
Robinvale, Vic 2003 This study B
X. translucens pv. translucens DAR 35705 Tamworth, NSW 1981 ACPPBd
aNSW, New South Wales; Vic, Victoria; SA, South Australia.bTwo groups identified among pathogen strains used in this study: group A, strains from Kyalite, Red Cliffs and Renmark; group B, strains from Robinvale.cPistachio canker epidemiology, project NT99004, Horticulture Australia Limited.dACPPB, Australian Collection of Plant Pathogenic Bacteria, Agricultural Institute, Orange, NSW, Australia.

Plant Pathology
(2006)
55
, 639–649
Pistachio xanthomonads in Australia
641
concentrations of separating and stacking gels were 10and 5%, respectively, and a Tris–glycine electrophoresisbuffer [6 g Tris base, 28·8 g glycine, 10 mL SDS (100 g L
−
1
)and 1000 mL water, pH 8·3] was used for electrophoresis.Gels were run at 5 mA in a cold room (4
°
C) for
c
. 17 hthen stained overnight in Coomassie Blue solution (0·1 gCoomassie Brilliant Blue R in 100 mL methanol : aceticacid : water 45 : 10 : 45, filtered through Whatman no. 1filter paper). Gels were destained in the methanol : aceticacid solution without Coomassie Blue for 2 h and stored in7% acetic acid. Gels were analysed visually and scannedimages were analysed using the
gel
-
pro analyzer
program(Media Cybernetics). To check reproducibility, proteinsamples were prepared from all strains at three differenttimes and the extracts were separated on three gels.
Preparation of DNA and rep-PCR genomic fingerprinting
All strains listed in Table 1 were grown on tryptone soyagar (TSA): 15 g tryptone, 5 g soy peptone, 5 g NaCl,15 g agar in 1000 mL water, pH 7·3 at 28
°
C for 48 h, andgenomic DNA was extracted following the method ofRademaker & de Bruijn (1997). DNA concentration wasdetermined by spectrophotometer and diluted to 50 ng
µ
L
−
1
.All strains were subjected to BOX-PCR using the BOXA1R primer (5
′
-CTACGGCAAGGCGACGCTGACG-3
′
);REP-PCR using the REP 1R primer (5
′
-IIIICGICGICAT-CIGGC-3
′
) and REP 2I primer (5
′
-ICGICTTATCIG-GCCTAC-3
′
); and ERIC-PCR using the ERIC 1R primer(5
′
-ATGTAAGCTCCTGGGGATTCAC-3
′
) and ERIC 2primer (5
′
-AAGTAAGTGACTGGGGTGAGCG-3
′
), withcycling conditions as described by Rademaker & deBruijn (1997). The primer sets were synthesized byProligo Pty Ltd and PCR amplification was performed ona Peltier Thermal Cycler model PTC-200 (MJ Research).PCR products were separated on 1·5% agarose gels in a16
×
35-cm horizontal gel electrophoresis apparatus using1
×
Tris-acetate-EDTA buffer at 70 V and 4
°
C for 17 h.Gels were stained with ethidium bromide (0·6
µ
g mL
−
1
),visualized on a UV transilluminator and photographed.The banding patterns were determined using
gel
-
proanalyzer
and scored in binary form, 1 and 0 indicating thepresence and absence of a band, respectively. A similaritymatrix was obtained using the Jaccard coefficient and thesoftware package
ntsys
-
pc
(ver. 2·02K, Applied Biostatis-tics). To determine the relationship among strains, clusteranalysis was performed with UPGMA (unweighted pair-group method using arithmetic averages) in the
sahn
pro-gram of the
ntsys
-
pc
software. A dendrogram was generatedfrom the BOX, REP and ERIC combined binary matrix. Tocheck the reproducibility of the technique, DNA extraction,rep-PCR and gel analysis were performed three times.
Sequencing of the 16S−−−−
23S rDNA spacer region
The nucleotide sequence of the 16S
−
23S rDNA spacerregion was determined for five strains selected randomlyacross the two groups distinguished by rep-PCR, and for
strains 7 and 25. The fragment was amplified in PCR usinga forward 20-bp primer (C1), 5
′
-AGTCGTAACAAGG-TAAGCCG-3
′
, derived from
Escherichia coli
16S rDNApositions 1493–1513; and a reverse 20-bp primer (C2),5
′
-C(T/C)(A/G)(T/C)TGCCAAGGCATCCACC-3
′
, corre-sponding to the
E. coli
23S rDNA sequence positions23–43 (Lane, 1991). PCR was performed in a total volumeof 50
µ
L, using 0·5
µ
L
Taq
polymerase (5 U
µ
L
−
1
, Qiagen),5
µ
L 10
× PCR buffer, 0·4 µL dNTP mixture (25 mm each),2·5 µL of C1 and C2 primers (10 µm each), 1 µL sample(50 ng µL−1) and 38·1 µL distilled water. PCR conditionswere: 1 × 95°C for 15 min, 29 × (95°C for 45 s, 50°C for1 min, 72°C for 2 min) and 72°C for 10 min. PCR product(10 µL) was run on an agarose gel (1%) and stainedwith ethidium bromide. The Perfectprep Gel Cleanup Kit(Brinkmann Instruments) was used to purify the fragmentfrom the gel, as recommended by the manufacturer. Thepurified fragment was directly sequenced in both directionsusing the Taq DyeDeoxy Terminator Sequencing Kit(Applied Biosystems) as indicated by the manufacturer.An isopropanol protocol was used to precipitate the exten-sion product, and the sequence was determined using anApplied Biosystem Model 3700 automated sequencer at theSequencing Centre, Institute of Medical and VeterinaryScience, Adelaide, Australia. The chromas (ver. 2) software(Technelysium) was used for editing and generating thesequences. Alignment and comparison of the sequenceswere performed with the programs clustalx andgenedoc, available on the Bioinformatics. Net database(www.bioinformatics.vg). Using the Basic Local AlignmentSearch Tool (blast), sequences were compared with thosein the GenBank, EMBL, DDBJ and PDB databases.Samples were prepared for sequencing twice, each timefrom different DNA preparations.
Biolog GN2 microplate system
Six strains of the pathogen, including strains 7 and 25, wereselected from the clusters generated by rep-PCR to testtheir ability to utilize 95 substrates as sole carbon sourcesin Biolog GN2 microplates (Biolog Inc.). Xanthomonastranslucens pv. translucens DAR 35705 was used for com-parison. A single colony of each strain was cultured onNA and then on TSA for 24 h. The bacteria were swabbedfrom the surface of the plates. Otherwise, inoculation,incubation and analysis were performed as per the manu-facturer’s instructions. Colour development was recordedwith a MAXLine microplate reader (Molecular Devices)and the results checked by visual observation. Metabolicfingerprint patterns were compared and identified after24, 48 and 72 h using the microlog gn ver. 4·01A soft-ware. This test was performed twice.
Pathogenicity and host-range tests
The pathogenicity was tested of representatives of groups Aand B, as indicated by rep-PCR, PAGE, ITS sequencing andbiochemical tests, to members of the Anacardiaceae andPoaceae. The Poaceae were included because of the close

Plant Pathology (2006) 55, 639–649
642 A. Marefat et al.
relatedness of the pistachio pathogen to X. translucens, indi-cated by ITS sequencing and Biolog GN2 microplate studies.
Anacardiaceae
The pathogenicity of strains 7 and 25 was first determinedby inoculation of 2-year-old P. vera cv. Sirora trees graftedon Pistacia terebinthus in the glasshouse. Xanthomonastranslucens pv. translucens DAR 35705 and sterile distilledwater were used as negative controls. Inocula were preparedfrom 48-h-old cultures on YDC and adjusted photomet-rically to 1·2 × 108 colony forming units (CFU) mL−1 steriledistilled water. There were three replicate plants pertreatment. Using a sterile syringe with a 25-gauge needle,bacterial suspension (c. 0·1 mL) was injected at three loca-tions per plant: in the trunk 5–7 cm above the graft union;at the junction of the trunk and the oldest branch; and at thejunction of a petiole and the oldest branch. Inoculationwas performed in spring (November) 2004. Followinginoculation, plants were watered, incubated individuallyin transparent plastic bags at 24/17°C day/night temperaturewith a 12-h photoperiod for 72 h, and monitored forsymptoms at 7- to 9-day intervals for 90 days after inoc-ulation. At the end of the experiment, internal symptomswere assessed, and leaf and discoloured stem tissues werecollected for pathogen reisolation. Plant tissues weremacerated in sterile distilled water and the suspensionsplated on YDC after 15 min. Plates were monitored foryellow, mucoid colonies after 48 h, and such bacteriawere purified on YDC then compared with the inoculumstrains using BOX-PCR.
To compare the host ranges of groups A and B, detachedshoots of Anacardiaceae were inoculated with a mixtureof two strains from each group of the pathogen: strains7 and 72 from group A; and strains 25 and 50 from groupB. A suspension of killed bacteria (all four strains); viableX. translucens pv. translucens DAR 35705; and distilledwater were used as negative controls. The plants used werePistacia vera, P. terebinthus, P. atlantica, P. chinensis,P. lentiscus, P. palaestina, Rhus leptodictya, R. tripartita,Schinus latifolius, S. lentiscifolius and S. polygamus.Excised shoots of P. terebinthus were obtained fromCSIRO Plant Industry, Merbein, Victoria, Australia; otherplant materials were collected from the Waite Arboretum,University of Adelaide, South Australia. Shoots 15–20 cmlong were collected from the current season’s growth.Bacterial suspensions were prepared from 48-h-oldcultures on YDC and adjusted to 1·2 × 107 CFU mL−1 steriledistilled water by spectrophotometry. Three replicateswere used for each inoculum–plant combination, andthree leaves and one bud were inoculated per replicate.Before inoculation, shoots were surface-sterilized using70% alcohol and rinsed three times with sterile distilledwater. Using a syringe with a 26-gauge needle, c. 0·2 mLof the bacterial suspension was infiltrated into the leaflamina and major veins until a water-soaked area wasvisible. Also, 0·1 mL of the suspension was injected intothe bud using a 25-gauge needle. Inoculated shoots wereplaced in autoclaved Sigmaware (C5916) culture tubes
containing 55 mL Murashige and Skoog basal medium(M5519; Sigma-Aldrich). Each treatment was placedindividually in an autoclaved transparent Sun Bag (B7026)(Sigma-Aldrich) containing sterile wet paper towel.Treatments were distributed randomly in a glasshousewith 24/17°C day/night temperature and a 12-h photo-period, and checked for symptoms 3, 5, 7, 9 and 14 daysafter inoculation. After 2 weeks, infected leaves and budswere collected for pathogen reisolation. Leaves weremacerated in sterile distilled water and the suspensionswere spread on YDC after 15 min. These agar plates weremonitored for yellow, mucoid colonies after 48 h, andsuch bacteria were purified on YDC and compared withthe inoculum strains using BOX-PCR. The experimentwas performed three times.
Poaceae
Strains 7 and 25 were selected as representative of groupsA and B, respectively. A mixture of five strains from eachgroup (3, 7, 11, 28 and 73 from group A; 20, 24, 27, 50 and70 from group B) was used for comparison, X. translucenspv. translucens DAR 35705 was inoculated as a positivecontrol, and distilled water and a suspension of killedbacteria (a mixture of the two groups) were used asnegative controls. Pathogenicity was tested on bread wheat(Triticum aestivum cv. Frame), durum wheat (T. durumcv. Arrivato), barley (Hordeum vulgare cv. Sloop), rye(Secale cereale), oat (Avena sativa cv. Echidna), brome grass(Bromus inermis), triticale (× Triticosecale cv. Tickit),timothy (Phleum pratense), cocksfoot (Dactylis glomerata),barley grass (Hordeum leporinum) and perennial ryegrass(Lolium perenne cv. Victorian). Seeds were surface-sterilizedwith 1% sodium hypochlorite for 10 min, 70% ethanolfor 5 s and 2% sodium thiosulphate for 5 min, and rinsedthree times with sterile distilled water after each stage.Except for ryegrass seed, which was obtained from apistachio grower at Kyalite, New South Wales, seedswere obtained from the South Australian Research andDevelopment Institute, Waite Campus, South Australia.Seedlings were grown in pasteurized potting mix in aglasshouse with 22/17°C day/night temperature in naturallight. Pots containing three seedlings at the three-leaf stagewere transferred to a growth room at 25/17°C with a 12-hphotoperiod. Three replicates were used for each treatment–plant combination, and three leaves were inoculated ineach replicate. Bacterial suspensions were prepared from48-h-old cultures on YDC and adjusted to 1·2 × 107 CFUmL−1 sterile distilled water by spectrophotometry. Thesuspension (0·1 mL) was infiltrated into intercellular spacesusing a syringe with a 26-gauge needle and two soft rubberstoppers (Klement, 1990). Inoculated plants were sealed inplastic chambers with a humidifier to produce 100% RHfor 4 days, then transferred to the glasshouse as describedpreviously. Inoculated plants were watered from below andmonitored for symptom development 3, 5, 7 and 10 daysafter inoculation. Observation of greasy and water-soakedstreaks covered with exudate 3–5 days after inoculation wasconsidered a compatible reaction (Bragard et al., 1997),
Plant Pathology (2006) 55, 639–649
Pistachio xanthomonads in Australia 643
and further development of symptoms was recorded forthe plants individually. Diseased leaves were collected forreisolation of bacteria 10 days after inoculation. Leaveswere macerated in a few drops of sterile distilled water andthe suspensions were spread on YDC after 15 min. Theseagar plates were monitored for yellow, mucoid coloniesafter 48 h, and such bacteria were purified on YDC andcompared with the inoculum strains using BOX-PCR.
Results
Biochemical tests
The results divided the strains from pistachio into twogroups, A and B. All strains shared the following featureswith X. translucens pv. translucens DAR 35705: mucoid
growth on SPA and YDC; Gram negative; oxidativemetabolism of glucose; oxidase negative; starch, gelatin,esculin and Tween 80 hydrolysis positive; H2S productionand growth at 36°C positive; urease and growth in thepresence of 5% NaCl negative. Strains belonging to groupA differed from the other bacteria in digestion of milkproteins, ice-nucleation activity, reaction in litmus milkand hypersensitivity on tobacco leaves (Table 2).
SDS–PAGE of whole-cell proteins
Strains fell into two groups based on analysis of the patternsof total proteins following SDS–PAGE. These groups wereidentical to those indicated by biochemical tests. Proteinprofiles comprised 25 and 27 reproducible bands, rangingfrom c. 4 to >90 kDa for groups A and B, respectively.
Table 2 Comparison of pistachio dieback pathogen strains with Xanthomonas translucens pv. translucens (DAR 35705) by biochemical and physiological tests
Test
Xanthomonads isolated from pistachioX. translucens pv. translucens DAR 35705Group A Group B
Gram reaction – (11/11)a – (10/10) –Oxidative/fermentative growth O+, F– (11/11) O+, F– (10/10) O+, F–Oxidase – (11/11) – (10/10) –Urease production – (11/11) – (10/10) –H2S production from cysteine + (11/11) + (10/10) +Nitrate reduction – (11/11) – (10/10) –Growth at 36°C + (11/11) + (10/10) +Mucoid on SPA and YDC + (11/11) + (10/10) +Growth in 5% NaCl – (11/11) – (10/10) –Starch hydrolysis + (11/11) + (10/10) +Gelatin hydrolysis + (11/11) + (10/10) +Esculin hydrolysis + (11/11) + (10/10) +Tween 80 hydrolysis + (11/11) + (10/10) +Digestion of milk proteins – (11/11) + (10/10) +Ice nucleation at:
−4°C – (11/11) + (10/10) +−10°C + (6/11)
In litmus milk:acid formation – (11/11) – (10/10) –peptonization + (11/11) + (10/10) +alkaline reaction w (11/11) + (10/10) +reduction – (11/11) + (10/10) +
Tobacco hypersensitive reaction + (11/11) – (10/10) –Utilization in Biolog microplate:
Tween 40 + (3/3) – (3/3) –Tween 80 + (3/3) – (3/3) –L-Fucose + (3/3) – (3/3) –Succinic acid mono-methyl-ester + (3/3) – (3/3) +L-Aspartic acid – (3/3) + (3/3) –α-Keto glutaric acid + (3/3) + (3/3) –D,L-Lactic acid + (3/3) + (3/3) –Bromosuccinic Acid + (3/3) + (3/3) –L-Alaninamide + (3/3) + (3/3) –L-Alanine + (3/3) + (3/3) –L-Alanylglycine + (3/3) + (3/3) –L-Glutamic acid + (3/3) + (3/3) –L-Serine + (3/3) + (3/3) –
+, Positive reaction; –, negative reaction; w, weak reaction.aNumber of strains positive or negative/number of strains tested.
Plant Pathology (2006) 55, 639–649
644 A. Marefat et al.
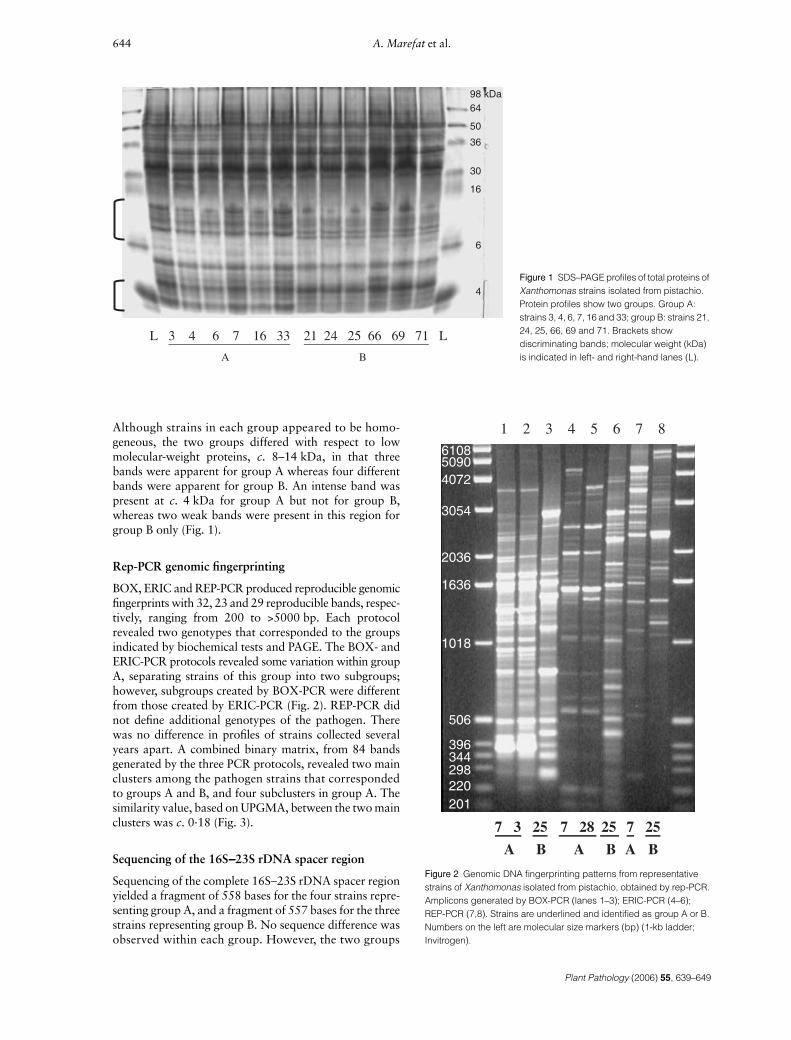
Although strains in each group appeared to be homo-geneous, the two groups differed with respect to lowmolecular-weight proteins, c. 8–14 kDa, in that threebands were apparent for group A whereas four differentbands were apparent for group B. An intense band waspresent at c. 4 kDa for group A but not for group B,whereas two weak bands were present in this region forgroup B only (Fig. 1).
Rep-PCR genomic fingerprinting
BOX, ERIC and REP-PCR produced reproducible genomicfingerprints with 32, 23 and 29 reproducible bands, respec-tively, ranging from 200 to >5000 bp. Each protocolrevealed two genotypes that corresponded to the groupsindicated by biochemical tests and PAGE. The BOX- andERIC-PCR protocols revealed some variation within groupA, separating strains of this group into two subgroups;however, subgroups created by BOX-PCR were differentfrom those created by ERIC-PCR (Fig. 2). REP-PCR didnot define additional genotypes of the pathogen. Therewas no difference in profiles of strains collected severalyears apart. A combined binary matrix, from 84 bandsgenerated by the three PCR protocols, revealed two mainclusters among the pathogen strains that correspondedto groups A and B, and four subclusters in group A. Thesimilarity value, based on UPGMA, between the two mainclusters was c. 0·18 (Fig. 3).
Sequencing of the 16S−−−−23S rDNA spacer region
Sequencing of the complete 16S−23S rDNA spacer regionyielded a fragment of 558 bases for the four strains repre-senting group A, and a fragment of 557 bases for the threestrains representing group B. No sequence difference wasobserved within each group. However, the two groups
Figure 1 SDS–PAGE profiles of total proteins of Xanthomonas strains isolated from pistachio. Protein profiles show two groups. Group A: strains 3, 4, 6, 7, 16 and 33; group B: strains 21, 24, 25, 66, 69 and 71. Brackets show discriminating bands; molecular weight (kDa) is indicated in left- and right-hand lanes (L).
Figure 2 Genomic DNA fingerprinting patterns from representative strains of Xanthomonas isolated from pistachio, obtained by rep-PCR. Amplicons generated by BOX-PCR (lanes 1–3); ERIC-PCR (4–6); REP-PCR (7,8). Strains are underlined and identified as group A or B. Numbers on the left are molecular size markers (bp) (1-kb ladder; Invitrogen).

Plant Pathology (2006) 55, 639–649
Pistachio xanthomonads in Australia 645
differed in 1, 6 and 5 bases in ITS1–3, respectively. The16S−23S rDNA spacer region, in all strains studied,contained tRNAala and tRNAile genes. The sequences forstrains 7 and 25, representing groups A and B, respectively,were deposited in the GenBank database under accessionnumbers AY579378 and AY579379, respectively. Com-parison with sequences in the GenBank, EMBL, DDBJ andPDB databases revealed that the 16S−23S rDNA sequence ofstrains in groups A and B most closely matched the regionin X. translucens pv. poae, with 98 and 97% similarity,respectively.
Biolog GN2 microplate system
All six strains tested were identified as X. campestris(syn. translucens) pv. translucens by their metabolicfingerprint. At the first reading, after 24 h, most wells werenegative, whereas more positive wells were read at 48and 72 h; there was no difference between the second andthird readings. Strains in groups A and B differed from oneanother in utilization of Tween 40, Tween 80, l-fucose,succinic acid mono-methyl-ester and l-aspartic acid(Table 2), whereas metabolic activity was recorded onthe other carbon substrates 48 h after inoculation for allstrains tested. Xanthomonas translucens pv. translucensDAR 35705 was also confirmed by the test as X. translucenspv. translucens. This strain did not metabolize α-ketoglutaric acid, d,l-lactic acid, bromosuccinic acid, l-alaninamide, l-alanine, l-alanylglycine, l-glutamic acid orl-serine, all of which were utilized by strains from pista-chio representing both groups (Table 2).
Pathogenicity on Anacardiaceae
Strains 7 and 25 induced symptoms on young pistachioplants. Necrosis developed at the branch–petiole junction7–10 days after inoculation, and at the trunk–branchjunction 20–25 days after inoculation. The necrotic areastended to enlarge in plants inoculated with strains 7 and25. No symptoms were visible at the inoculation site near
the graft union on the trunk. In plants inoculated with thepistachio strains, necrosis and dieback began on treatedbranches and foliage arising from them about 45 days afterinoculation, while untreated branches and their shootsremained healthy. At the end of the experiment, stainedwood was observed in two treated branches, as well as oneuntreated branch adjacent to a branch inoculated withstrain 7, while only one branch inoculated with strain 25showed staining. No symptoms, internal or external, wereobserved on controls, except for tiny necrotic areas atinjection points. Xanthomonads isolated from plants withsymptoms were assigned to group A or B by BOX-PCR.
Inoculation of detached shoots with the pistachio strainsresulted in water-soaked spots on leaves of all the Anac-ardiaceae tested, except R. leptodictya (Table 3), 3–4 daysafter inoculation with strains from groups A and B. Anecrotic area expanded from the middle of this spot 5–7days after inoculation. Such symptoms were identicalfor all three replicates for each treatment. Spots weresurrounded by a yellowish halo on some inoculated leaves,and discoloration spread along the veins in some hosts.Complete necrosis and collapse of some infected leaves wereobserved 14 days after inoculation. Strains from group Aappeared to be more aggressive than those from group B.White–grey bacterial ooze was observed from some budsinoculated with strains from group A. Strains from groupA were pathogenic to P. lentiscus, whereas strains fromgroup B failed to induce any symptoms on this species.Leaves treated with sterile distilled water, killed bacteriaor X. translucens pv. translucens DAR 35705 did not showwater-soaked lesions. Xanthomonads isolated from inocu-lated shoots were assigned to group A or B by BOX-PCR.
Figure 3 Dendrogram generated from a combined binary matrix derived from amplicons obtained by BOX-, ERIC- and REP-PCR, demonstrating fingerprint groups of 65 strains of Xanthomonas isolated from pistachio in Australia. Numbers on the right denote the number of strains falling within groups A and B.
Table 3 Differences among xanthomonads isolated from pistachio and Xanthomonas translucens pv. translucens DAR 35705 from wheat, based on pathogenicity to detached leaves of species of Anacardiaceae
Host
Pistachio strainsa
X. translucens pv. translucens DAR 35705Group A Group B
Pistacia vera + + –Pistacia atlantica + + –Pistacia terebinthus + + –Pistacia chinensis + + –Pistacia lentiscus + – –Pistacia palaestina + + –Rhus leptodictya – – –Rhus tripartita + + –Schinus latifolius + + –Schinus lentiscifolius + + –Schinus polygamus + + –
aInoculum comprised a mixed suspension of two strains representing group A or two strains representing group B.+, Positive reaction (water-soaked area on leaf lamina within 4 days of inoculation followed by necrosis); –, negative reaction or chlorotic symptoms restricted to inoculated areas only. Symptoms were identical in three replicates for each treatment. There were no symptoms on controls treated with sterile distilled water or killed bacteria (data not shown).

Plant Pathology (2006) 55, 639–649
646 A. Marefat et al.
Pathogenicity on Poaceae
Strains belonging to groups A and B induced water-soaked,greasy areas with bacterial exudate within 3–5 days ofinoculation, on bread wheat, durum wheat, barley, rye, oat,brome grass, triticale, barley grass and cocksfoot (Table 4).Streaks with a greasy appearance developed after 7 days.A translucent lesion with a yellow to light brown halooccurred on some leaves. Strains from group A failed toinduce lesions on timothy grass, whereas strains fromgroup B produced water-soaked streaks 3–5 days afterinoculation, followed by necrosis and collapse 7–10 daysafter inoculation. Likewise, strains from group B inducedno symptoms on ryegrass, whereas strains from group Aproduced symptoms as described above. Xanthomonastranslucens pv. translucens DAR 35705 was pathogenicon all plants tested except cocksfoot and ryegrass. Leavestreated with water or killed bacteria remained healthy.Xanthomonads isolated from plants with symptoms wereassigned to group A or B, or DAR 35705, by BOX-PCR.
Discussion
Using DNA-independent and DNA-based analyses, thepistachio dieback pathogen, previously identified as similarto X. translucens, was found to be phenotypically, biolog-ically and genetically heterogeneous. Two distinct groupswere identified, supporting preliminary findings based onfive strains (Facelli et al., 2005). Group A included strains
from three areas (Kyalite, Renmark and Red Cliffs) andrep-PCR revealed genetic variation within this group.Group B comprised identical strains obtained from onearea only (Robinvale). Strains of the two groups have notyet been found in the same tree. The pathogen strainswere closely related to the X. translucens group, with ITSsequences 97–98% similar to X. translucens pv. poae.
BOX-, ERIC- and REP-PCR, which amplify specificconserved repetitive sequences distributed throughout thegenome of diverse bacteria, have proved effective in discrim-inating Xanthomonas strains, especially at the pathovarlevel (Louws et al., 1994). In the present study, ampliconsproduced following these PCR protocols revealed twogroups, A and B, among Xanthomonas strains from pista-chio. Furthermore, BOX- and ERIC-PCR protocols showedtwo subgroups within group A. The similarity value ofc. 0·18 between the two main clusters, based on a combinedbinary matrix generated by the three protocols, confirmedthat the two groups were genetically distinct. Proteinprofiles and ITS sequencing supported this grouping.Furthermore, differential host ranges in pathogenicity testson representative Anacardiaceae and Poaceae revealed abiological difference between the two groups.
Physiological and biochemical tests supported thisgrouping, and phenotypic properties of the pathogen wereagain similar to those of X. translucens. However, it shouldbe noted that, whereas positive reactions have been reportedfor X. translucens in digestion of milk proteins and ice-nucleation tests (Schaad et al., 2001), group A strains werenegative for milk digestion and several did not exhibit ice-nucleation activity. In the Biolog test, the data obtained48 h after inoculation of the plates were used because colourreactions at 48 h were considered more accurate thanthose at 24 h because of the slow growth of the bacteria,as reported by Vauterin et al. (1995). The discriminatorysubstrates indicated by this analysis are valuable for diag-nostic tests and may also be useful in designing a selectivemedium for each group.
Of the various techniques used in this study, only rep-PCRdiscriminated subgroups in group A, demonstrating thatrep-PCR had more resolution than the other techniques todiscriminate among closely related strains associated withpistachio dieback.
There was a relationship between the groups and thegeographical origin of the strains, in that group A strainsoriginated from Kyalite, Red Cliffs and Renmark, whereasgroup B comprised strains from Robinvale only. TheRobinvale orchard is situated between the orchards atKyalite and Red Cliffs. As growers at all four locations haveshared budwood in the past, the origin of this distributionis not clear, although it is possible that the two groupsrepresent different, recent introductions of the pathogen.Linkage between groups (indicated by rep-PCR) andgeographical origin has been reported for other diseasescaused by Xanthomonas. Scortichini et al. (2001) reportedthe existence of genetic diversity among strains of Xan-thomonas axonopodis pv. juglandis collected from differentgeographical areas, and showed that the pathogenpopulation was unique to each walnut-cultivation area.
Table 4 Differences among xanthomonads isolated from pistachio and Xanthomonas translucens pv. translucens DAR 35705 isolated from wheat based on pathogenicity to species of Poaceae
Plant
Pistachio strainsa X. translucens pv.
translucens DAR 35705A B
Wheat (bread) + + +Wheat (durum) + + +Barley + + +Rye + + +Oat + + +Brome grass + + +Triticale + + +Barley grass + + +Cocksfoot + + –Timothy – + +Ryegrass + – –
aInoculum comprised one strain or a mixture of five strains representing group A, or one strain or a mixture of five strains representing group B.+, Compatible reaction (water-soaked and greasy area covered by exudates on leaf lamina within 3–5 days of inoculation followed by necrosis); –, incompatible reaction (without any symptoms or chlorotic symptoms restricted to inoculated areas only). There was no difference between treatments inoculated with one strain or a mixture of five strains representing each group. Symptoms were identical in three replicates for each treatment and there were no symptoms on controls treated with sterile distilled water or killed bacteria (data not shown).

Plant Pathology (2006) 55, 639–649
Pistachio xanthomonads in Australia 647
Also, Massomo et al. (2003) observed variation amongX. campestris pv. campestris strains obtained from brassicasin Tanzania and linked each fingerprint pattern to ageographical area. Likewise, Mkandawire et al. (2004)revealed that X. campestris pv. phaseoli strains collectedfrom Malawi, Tanzania, East Africa and Wisconsincomprised three genotypes, two unique to East Africa andthe third associated with other locations. Selection for aspecialized niche can affect genome organization and dis-tribution of repetitive sequences in the bacterium genome,resulting in fingerprints unique to a specific pathovar orstrain (Louws et al., 1994).
As shown by ITS sequencing, as well as phenotypic andpathogenicity properties, the pistachio pathogen belongsto the translucens group, a group of closely related patho-vars causing bacterial leaf streak in small grains and grasses(Vauterin et al., 1992). This group, along with severalpathovars also isolated from the Poaceae that are not patho-genic on small grains, constitutes the species X. translucens(Vauterin et al., 1995). Because of the high degree ofhomogeneity within the group, the pathovars cannot bedistinguished reliably based on fatty acid analysis, SDS–PAGE of proteins (Stead, 1989; Vauterin et al., 1992),polyclonal antibodies (Azad & Schaad, 1988) or mono-clonal antibodies (Bragard & Verhoyen, 1993). Despiteseveral attempts to differentiate the pathovars by DNA-based techniques (Bragard et al., 1995, 1997; Alizadehet al., 1997), pathogenicity tests are required, especially forthose pathogenic on small grains. The close relatedness ofthe pistachio pathogen to X. translucens suggests that thesestrains may have been introduced to pistachio from grasses,possibly evolved from different ancestors (X. translucensindigenous and exotic to Australia). Xanthomonas trans-lucens strains have already been reported in Australia(Noble, 1935; Moffett & McCarthy, 1973), and an X.translucens-like bacterium, although different from thepistachio pathogen strains in rep-PCR fingerprint and ITSsequence, has been found regularly on grasses collectedfrom pistachio orchards (data not shown). It would be ofinterest to determine if these strains may have been intro-duced from grasses to pistachio, or if pistachio strains cancolonize grasses in the orchards.
The virulence determination of a new pathogen,especially one that causes shoot death and dieback, usingthe hypersensitive reaction (HR) test, is critical. Strains ingroup A induced a typical HR on tobacco leaves, indicat-ing that they were pathogenic, whereas strains in group Bdid not. Groups A and B could also be distinguished bypathogenicity to the Anacardiaceae and Poaceae, whereasa preliminary study with a limited range of strains and hostsfailed to reveal differences between groups (Facelli et al.,2005). Group A appeared to be more aggressive than groupB in terms of the HR response, development of symptomsin inoculated plants and recovery of the pathogen frominoculated buds. Differences in aggressiveness have beennoted for X. translucens pv. undulosa strains from variousgeographical areas (Bragard & Maraite, 1994). Indeed, inthe translucens group, single strains can have different hostranges (Duveiller et al., 1997), and this can be affected by
transposon and chemical mutagenesis (Mellano & Cooksey,1988). Further studies are needed to determine the structureand function of the genes involved in pathogenicity andhost-range determination in pistachio strains.
Previous attempts to confirm the pathogenicity of oneXanthomonas strain isolated from pistachio by injectionof bacteria into the trunks of 2-year-old grafted trees inpots in autumn resulted in discoloration of wood andreisolation of the bacteria from various parts of the plants.Monitoring of inoculated trees for trunk and limb lesionscontinues on the basis that such symptoms are observed intrees 6–7 years old (Facelli et al., 2002, 2005). In the presentstudy, however, multiple inoculation points (especiallyclose to the apex), inoculation in spring, and providing amoist environment for several days after inoculationresulted in necrosis and dieback. Xanthomonas translucenspv. translucens DAR 35705 was not pathogenic topistachio, demonstrating that despite the close relatednessof pistachio and cereal strains, they differ in pathogenicity.Recently, canker, leaf spot and dieback have been reportedon 1-year-old pistachio seedlings in Iran and, based onlimited physiological and biochemical tests, a Xanthomonassp. was identified as the causal agent (Tarighi & Rahimian,2001). Some characteristics of this Xanthomonas, such asice-nucleation activity, tobacco hypersensitive reactionand use of some carbon sources, were similar to those ofAustralian group A. Furthermore, dieback and discolora-tion in woody tissues of pistachio, albeit as an unknowndisorder, have recently been reported in Turkey (Sarpkaya& Can, 2005).
Inoculation of excised shoots, as a manageable and lesstime-consuming method, was used to compare groups Aand B with respect to host range. Strains representative ofboth groups, including strain 25, apparently identicalto that tested previously by Facelli et al. (2002, 2005),induced lesions on leaves of shoots excised from trees ofvarious Anacardiaceae species, whereas such symptomshave not been observed in pistachio orchards. This mayreflect optimal conditions for infection in the bioassay, orthe fact that leaves incubated in moist conditions are moresusceptible than those on the tree. Inoculation of smallparts under optimal controlled conditions often shows arapid response (Randhawa & Civerolo, 1985). Detachedplant material has been used to demonstrate the pathogenic-ity of various xanthomonads (Sharon et al., 1982; Fahy &Hayward, 1983; Randhawa & Civerolo, 1985; Goncalves& Rosato, 2000). It is possible that Xanthomonas belongingto groups A and B infect foliage of plants other thancultivated pistachio in nature, albeit rarely, and that thepathogen may have been introduced to pistachio fromother species of Anacardiaceae. Further pathogenicitytests within the Anacardiaceae are in progress to assessthis possibility.
In summary, rep-PCR and ITS sequencing differentiatedXanthomonas strains associated with pistachio dieback inAustralia into two distinct groups, which differed in bio-chemical and physiological tests, protein profile analysis andhost range. A full taxonomic description of the bacteria,based on a polyphasic study, is in preparation. The 16S−23S

Plant Pathology (2006) 55, 639–649
648 A. Marefat et al.
rDNA sequences will provide the basis for the developmentof a PCR-based assay for specific detection of each group,for use in pathogen recognition and disease diagnosis.
Acknowledgements
We thank E. Facelli, C. Taylor and R. Emmett for theiruseful comments and assistance during this research. We alsoacknowledge K. Dowling for assistance with experimentaldesign and analysis. This research was supported by a grantfrom Horticulture Australia Limited (HAL, NT02007)and the Pistachio Growers Association of Australia.
References
Alizadeh A, Arlat M, Sarrafi A, Boucher CA, Barrault G, 1997. Restriction fragment length polymorphism analyses of Iranian strains of Xanthomonas campestris from cereals and grasses. Plant Disease 81, 31–5.
Azad H, Schaad NW, 1988. Serological relationships among membrane proteins of strains of Xanthomonas campestris pv. translucens. Phytopathology 78, 272–7.
Bragard C, Maraite H, 1994. Pathogenic variation in Xanthomonas campestris pv. undulosa. In: Lemattre M, Freigoun S, Rudolph K, Swings J, eds. Proceedings of the 8th International Conference on Plant Pathogenic Bacteria, Versailles, France. Paris: INRA/ORSTOM, 807–12.
Bragard C, Verhoyen M, 1993. Monoclonal antibodies specific for Xanthomonas campestris bacteria pathogenic on wheat and other small grains, in comparison with polyclonal antisera. Journal of Phytopathology 139, 217–28.
Bragard C, Verdier V, Maraite H, 1995. Genetic diversity among Xanthomonas campestris strains pathogenic for small grains. Applied and Environmental Microbiology 61, 1020–6.
Bragard C, Singer E, Alizadeh A, Vauterin L, Maraite H, Swings J, 1997. Xanthomonas translucens from small grains: diversity and phytopathological relevance. Phytopathology 87, 1111–7.
Cubero J, Graham JH, 2002. Genetic relationship among worldwide strains of Xanthomonas causing canker in citrus species and design of new primers for their identification by PCR. Applied and Environmental Microbiology 68, 1257–64.
Duveiller E, Bragard C, Maraite H, 1997. Bacterial leaf streak and black chaff caused by Xanthomonas translucens. In: Duveiller E, Fucikovski L, Rudolph K, eds. The Bacterial Diseases of Wheat: Concept and Methods of Disease Management. Mexico, D.F: CIMMYT.
Edwards M, Taylor C, 1998. Pistachio canker: the story so far. In: Wilkinson J, ed. Proceedings of the Eighth Australian Nut Industry Council Conference. Victoria, Australia: Australian Nut Industry Council, 31–2.
Facelli E, Taylor C, Scott E, Emmett R, Fegan M, Sedgley M, 2002. Bacterial dieback of pistachio in Australia. Australasian Plant Pathology 31, 95–6.
Facelli E, Taylor C, Scott E, Fegan M, Huys G, Emmett R, Noble D, Sedgley M, 2005. Identification of the causal agent of pistachio dieback in Australia. European Journal of Plant Pathology 112, 155–65.
Fahy PC, Hayward AC, 1983. Media and methods for isolation and diagnostic tests. In: Fahy PC, Persley GJ, eds. Plant Bacterial Diseases, A Diagnostic Guide. Sydney, Australia: Academic Press, 337–78.
Goncalves ER, Rosato YB, 2000. Genotypic characterization of xanthomonad strains isolated from passion fruit plants (Passiflora spp.) and their relatedness to different Xanthomonas species. International Journal of Systematic and Evolutionary Microbiology 50, 811–21.
Goncalves ER, Rosato YB, 2002. Phylogenetic analysis of Xanthomonas species based upon 16S−23S rDNA intergenic spacer sequences. International Journal of Systematic and Evolutionary Microbiology 52, 355–61.
Klement Z, 1990. Inoculation of Plant Tissues. In: Klement Z, Rudolph K, Sands DC, eds. Methods in Phytobacteriology. Budapest, Hungary: Akademiai Kiado and Nyomda Vallalat, 103–4.
Laemmli UK, 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680–5.
Lane DJ, 1991. Small subunit ribosomal RNA sequences and primers. Large subunit ribosomal RNA sequences and primers. In: Goodfellow E, ed. Nucleic Acid Techniques in Bacterial Systematics. Chichester, UK: John Wiley & Sons, 148–75.
Lindow SE, 1990. Bacterial Ice-Nucleation Activity. In: Klement Z, Rudolph K, Sands DC, eds. Methods in Phytobacteriology. Budapest, Hungary: Akademiai Kiado and Nyomda Vallalat, 428–34.
Lopes SA, Damann KE, Grelen LB, 2001. Xanthomonas albilineans diversity and identification based on rep-PCR fingerprints. Current Microbiology 42, 155–9.
Louws FJ, Fulbright DW, Stephens CT, de Bruijn FJ, 1994. Specific genome fingerprints of phytopathogenic Xanthomonas and Pseudomonas pathovars and strains generated with repetitive sequences and PCR. Applied and Environmental Microbiology 60, 2286–95.
Louws FJ, Fulbright DW, Stephens CT, de Bruijn FJ, 1995. Differentiation of genomic structure by rep-PCR fingerprinting to rapidly classify Xanthomonas campestris pv. vesicatoria. Phytopathology 85, 528–36.
Louws FJ, Rademaker JLW, de Bruijn FJ, 1999. The three Ds of PCR-based genomic analysis of phytobacteria: diversity, detection, and disease diagnosis. Annual Review of Phytopathology 37, 81–125.
Maggs DH, 1982. An Introduction to Pistachio Growing in Australia. Adelaide, Australia: CSIRO.
Massomo SMS, Nielsen H, Mabagala RB, Mansfeld-Giese K, Hockenhull J, Mortensen CN, 2003. Identification and characterisation of Xanthomonas campestris pv. campestris strains from Tanzania by pathogenicity tests, Biolog, rep-PCR and fatty acid methyl ester analysis. European Journal of Plant Pathology 109, 775–89.
Mellano VJ, Cooksey DA, 1988. Development of host range mutants of Xanthomonas campestris pv. translucens. Applied and Environmental Microbiology 54, 884–9.
Milgroom MG, Fry WE, 1997. Contributions of population genetics to plant disease epidemiology and management. Advances in Botanical Research 24, 1–30.
Mkandawire ABC, Mabagala RB, Guzman P, Gepts P, Gilbertson RL, 2004. Genetic diversity and pathogenic variation of common blight bacteria (Xanthomonas

Plant Pathology (2006) 55, 639–649
Pistachio xanthomonads in Australia 649
campestris pv. phaseoli and X. campestris pv. phaseoli var. fuscans) suggests pathogen coevolution with the common bean. Phytopathology 94, 593–603.
Moffett ML, McCarthy JP, 1973. Xanthomonas translucens on Japanese millet (Echinochloa crus-galli var. frumentacea). Australian Journal of Experimental Agriculture and Animal Husbandry 13, 452–4.
Noble RJ, 1935. Notes on plant diseases recorded in New South Wales for the year ending 30th June, 1935. International Bulletin of Plant Protection 12, 270–3.
Rademaker JLW, de Bruijn FJ, 1997. Characterization and classification of microbes by rep-PCR genomic fingerprinting and computer-assisted pattern analysis. In: Caetano-Anolles G, Gresshoff PM, eds. DNA Markers: Protocols, Application and Overviews. New York, USA: John Wiley & Sons, 151–71.
Randhawa PS, Civerolo EL, 1985. A detached-leaf bioassay for Xanthomonas campestris pv. pruni. Phytopathology 75, 1060–3.
Sahin F, Kotan R, Abbasi PA, Miller SA, 2003. Phenotypic and genotypic characterization of Xanthomonas campestris pv. zinniae strains. European Journal of Plant Pathology 109, 165–72.
Sands DC, 1990. Physiological criteria – determinative tests. In: Klement Z, Rudolph K, Sands DC, eds. Methods in Phytobacteriology. Budapest, Hungary: Akademiai Kiado and Nyomda Vallalat, 133–43.
Sarpkaya K, Can C, 2005. Dieback: an unknown disorder in pistachio. In: Proceedings of the IV International Symposium on Pistachios and Almonds. Tehran, Iran: Iranian Pistachio Research Institute, 192.
Schaad NW, Jones JB, Chun W, 2001. Laboratory Guide for Identification of Plant Pathogenic Bacteria, 3rd edn. St Paul, MN, USA: APS Press.
Scortichini M, Rossi MP, 2003. Genetic diversity of Xanthomonas arboricola pv. fragariae strains and comparison with some other X. arboricola pathovars using repetitive PCR genomic fingerprinting. Journal of Phytopathology 151, 113–9.
Scortichini M, Marchesi U, Di Prospero P, 2001. Genetic diversity of Xanthomonas arboricola pv. juglandis (synonyms: X. campestris pv. juglandis; X. juglandis pv. juglandis) strains from different geographical areas shown by repetitive polymerase chain reaction genomic fingerprinting. Journal of Phytopathology 149, 325–32.
Sharon E, Okon Y, Bashan Y, Henis Y, 1982. Detached leaf enrichment: a method for detecting small numbers of Pseudomonas syringae pv. tomato and Xanthomonas campestris pv. vesicatoria in seed and symptomless leaves of tomato and pepper. Journal of Applied Bacteriology 53, 371–7.
Stead DE, 1989. Grouping of Xanthomonas campestris pathovars of cereals and grasses by fatty acid profiling. EPPO Bulletin 19, 57–68.
Tarighi S, Rahimian H, 2001. Canker and leafspot of pistachio caused by Xanthomonas sp. Iranian Journal of Plant Pathology 37, 161–2.
Vauterin L, Swings JG, Kersters K, 1991a. Grouping of Xanthomonas campestris pathovars by SDS–PAGE of proteins. Journal of General Microbiology 137, 1677–87.
Vauterin L, Yang P, Hoste B, Vancanneyt M, 1991b. Differentiation of Xanthomonas campestris pv. citri strains by sodium dodecyl sulphate polyacrylamide gel electrophoresis of proteins, fatty acid analysis, and DNA–DNA hybridization. International Journal of Systematic Bacteriology 41, 535–42.
Vauterin L, Yang P, Hoste B, Pot B, Swings J, Kersters K, 1992. Taxonomy of xanthomonads from cereals and grasses based on SDS–PAGE, fatty acid analysis and DNA hybridization. Journal of General Microbiology 138, 1467–77.
Vauterin L, Hoste B, Kersters K, Swings J, 1995. Reclassification of Xanthomonas. International Journal of Systematic Bacteriology 45, 472–89.
Verniere C, Pruvost O, Civerolo EL, Gambin O, Jacquemoud-Collet JP, Luisetti J, 1993. Evaluation of the Biolog substrate utilization to identify and assess metabolic variation among strains of Xanthomonas campestris pv. citri. Applied and Environmental Microbiology 59, 243–9.





























