Embed Size (px)
Citation preview

Introduction
Industrial composting is a microbial, aerobic, self-heating and solid-phase biodegradation process of organic-waste materials (Miller, 1996). During the ther-mogenic phase of the composting process, a large
Geobacillus toebii subsp. decanicus subsp. nov., a hydrocarbon-degrading, heavy metal resistant
bacterium from hot compost
Annarita Poli,1 Ida Romano,1 Gaetano Caliendo,4 Giancarlo Nicolaus,3 Pierangelo Orlando,2 Antonio de Falco,4
Licia Lama,1 Agata Gambacorta,1 and Barbara Nicolaus1,*
1 Istituto di Chimica Biomolecolare (ICB), CNR, Via Campi Flegrei 34, 80078 Pozzuoli, Napoli, Italy2 Istituto di Biochimica delle Proteine (IBP), CNR, Napoli, Italy
3 Istituto di Ricerche di Biologia Molecolare “P. Angeletti” IRBM, Pomezia, Rome, Italy4 Pomigliano Ambiente s.p.a., Napoli, Italy
(Received December 14, 2005; Accepted August 4, 2006)
A thermophilic, spore-forming bacterial strain L1T was isolated from hot compost “PomiglianoEnvironment” s.p.a., Pomigliano, Naples, Italy. The strain was identified by using a polyphasictaxonomic approach. L1T resulted in an aerobic, gram-positive, rod-shaped, thermophilic with anoptimum growth temperature of 68°C chemorganotrophic bacterium which grew on hydrocar-bons as unique carbon and energy sources and was resistant to heavy metals. The G�C DNAcontent was 43.5 mol%. Phylogenetic analysis of 16S rRNA gene sequence and Random Ampli-fied Polymorphic DNA-PCR (RAPD-PCR) analysis of L1T and related strains showed that it formswithin Geobacillus toebii, a separate cluster in the Geobacillus genus. The composition of cellu-lar fatty acids analyses by Gas-Mass Spectroscopy differed from that typical for the genusGeobacillus in that it is lacking in iso-C15 fatty acid, while iso-C16 and iso-C17 were predomi-nant. Isolates grew on a rich complex medium at temperatures between 55–75°C and presented adoubling time (td) of 2 h and 6 h using complex media and hydrocarbon media, respectively.Among hydrocarbons tested, n-decane (2%) was the more effective to support the growth (1 g/Lof wet cells). The microorganism showed resistance to heavy metal tested during the growth.Furthermore, intracellular aa-galactosidase and aa-glucosidase enzymatic activities were de-tectable in the L1T strain. Based on phenotypic, phylogenetic, fatty acid analysis and resultsfrom DNA-DNA hybridization, we propose assigning a novel subspecies of Geobacillus toebii, tobe named Geobacillus toebii subsp. decanicus subsp. nov., with the type strain L1T (�DSM17041�ATCC BAA 1004).
Key Words——alkanes; DNA-DNA hybridization; fatty acid; Geobacillus; heavy metals; hot compost;lipid; PCR finger print; thermozymes
J. Gen. Appl. Microbiol., 52, 223–234 (2006)
* Address reprint requests to: Dr. Barbara Nicolaus, Istituto diChimica Biomolecolare (ICB), CNR, Via Campi Flegrei 34,80078 Pozzuoli, Napoli, Italy.
Tel: �39–081–8675245 Fax: �39–081–8041770E-mail: [email protected]
Full Paper

central zone remains at temperatures higher than70°C for many weeks (Fugio and Kume, 1991; Strom,1985a, b). In this “hot-zone” a high number of ther-mophilic bacteria, forming a different microbial commu-nity, belonging to Thermus, Bacillus and Hy-drogenobacter species were identified (Beffa et al.,1996; Blanc et al., 1999; Lyon et al., 2000; Strom,1985a).
Thermophiles isolated from compost in recent yearshave become extremely interesting from a technologi-cal point of view (Lyon et al., 2000; Strom 1985b).Many are the studies undertaken for defining the prin-cipal metabolic pathways and the peculiar propertiesof their molecules. Hot compost is considered to offera favorable habitat for thermophilic bacilli. Strom(1985b) isolated more than 750 heterotrophic spore-forming strains from compost. Most of these microor-ganisms grew under 60°C, and only Bacillus coagu-lans, Geobacillus stearothermophilus and Geobacillustoebii were isolated at 65°C (Strom, 1985a; Sung etal., 2002). Thermophilic bacteria related to the genusGeobacillus has been widely isolated from geothermaland man-made environments throughout the world(Maugeri et al., 2002; Nazina et al., 2001, 2005; Rheeet al., 2002; Sung et al., 2002).
Thermophilic Bacillus species of the group 5 rRNAare a phenotypically and phylogenetic coherent groupdisplaying very high similarity among their 16S rRNAsequences (98.5–99.2%) (Ash et al., 1991; Rainey etal., 1994; Sunna et al., 1997); this group was trans-ferred to a new genus Geobacillus that comprised at that time 12 validated species G. stearother-mophilus, G. thermocatenulatus, G. thermoleovorans,G. kaustophilus, G. thermoglucosidasius, G. thermo-denitrificans, G. subterraneus, G. uzenensis, G. cal-doxylosilyticus and G. toebii (Ahmad et al., 2000;Claus and Berkeley, 1986; Fortina et al., 2001a, b;Golovacheva et al., 1975; Logan and Berkeley, 1984;Manichini et al., 2000; Nazina et al., 2001; Priest et al.,1988; Sung et al., 2002; Suzuki et al., 1983; Tomita etal., 2003; White et al., 1993; Zarrilla and Perry, 1987).
Other thermophilic established species belonged toGeobacillus group such as Bacillus thermantarcticus(Nicolaus et al., 1996) validated in Int. J. Syst. Evol.Microbiol. 2002 and G. vulcani (Caccamo et al., 2000;Gugliandolo et al., 2003; Maugeri et al., 2001; Nazinaet al., 2004). The asporogenous Saccharococcus ther-mophilus representing a separate line of descent (Naz-ina et al., 2001; Nystrand, 1984) and more recently G.
caldoproteolyticus (Chen et al., 2004), G. gargensis(Nazina et al., 2004), and G. jurassicus (Nazina et al.,2005) were assigned to the Geobacillus group.
This paper describes the isolation of a new ther-mophilic Geobacillus strain, designated L1T, from ther-mogenic compost made from 12-week-old organicwaste samples, able to grow at temperatures up to75°C. The characteristics of this isolate were com-pared with its nearest neighbor. Data on L1T strainability to grow using hydrocarbons as unique carbonsources and its resistance to heavy metals, were alsoreported.
Materials and Methods
Compost characteristics. Compost samples werefrom “Pomigliano Environment” s.p.a., Pomiglianod’Arco (Campania Region, southern Italy). The firststep, in the bio-fermentation process, is essentiallybased on a static heap covered by a sandwich consti-tuted of two supports of polyester with interposed a foilof Gore-Tex® and on a forced airing cycle-treatment of30 days. The process is monitored by a computerizedsystem (BIOE® s.r.l, Milan, Italy) that checks the oxy-gen, the temperature and the humidity levels. The tem-perature of the inner core of the heap is above 60°C.At 30 days from pretreatment, the pile is moved to thethreshing floor of maturation where, for a further 60days, it is turned weekly to favor the oxygenation andconsequently, the degradation of the more slowlybiodegradable matrixes (lignin). The material is refinedthrough a phase of drum sifting by obtaining the “com-post.” The presence of heavy metals was determinedby atomic spectrometry (Shimadzu AA6200) (Poli etal., 2005). Bacterial tests were performed by the meth-ods of Koneman (Koneman, 1984).
Sampling and isolation. Thirty grams of compost(fresh wt) were placed in 200 ml of sterile water, ho-mogenized at room temperature on a shaker (150 rpm)for 20 min, and serially diluted (10�2 to 10�10) in the THmedium (8 g/L peptone (Oxoid), 4 g/L yeast extract(Oxoid), 2 g/L NaCl (Applichem) at pH 7.0). For opti-mum temperature determination the cultures were in-cubated without agitation from 50°C to 80°C for 1 to 6days. The pH dependence of growth was tested in thepH range 4.0 to 10.0. Pure strains were isolated at68°C at pH of 7.2 on the TH medium solidified withagar (2%, wt/v). The first pure culture obtained wascalled L1T and studied in detail.
224 POLI et al. Vol. 52

Reference strains and media. The referencestrains used were as follows: Geobacillus toebii DSM14590T (Sung et al., 2002), Geobacillus thermoleovo-rans ATCC 43513T (Zarrilla and Perry, 1987) andGeobacillus caldoxylosilyticus ATCC 42125T. Themedia utilized were: TH medium; TH agar medium,containing (g/L) 20.0 agar (Oxoid); YN medium, con-taining (g/L) 6.0 yeast extract, 6.0 NaCl at pH 6.5.
Other media were prepared as follows: M162 min-eral medium containing: (g/L) 4.0 NaCl (Applichem),0.53 NH4Cl (J.T. Baker), Solution A 60 ml/L, Solution B20 ml/L, Solution C 100 ml/L; Solution A: (g/L) 35.58Na2HPO4·2H2O (Applichem); Solution B: (g/L) 27.19KH2PO4 (Carlo Erba); Solution C: (g/L) 1.0 Nitrilotri-acetic acid (Applichem), 0.4 CaSO4·6H2O (CarloErba), 2.0 MgCl2· 6H2O (Applichem), 2.5 Ferric citrate(Carlo Erba), Nitsch’s trace elements 5.0 ml/L.
Minimal media were prepared using M162 mediumplus either 1% glycerol, glucose, lactose, Na-acetate,mannose, xylose, galactose, sucrose, cellobiose, ri-bose, maltose, fructose, ethanol, EDTA, sorbose, raffi-nose, malic acid, citric acid or trehalose as sole carbonsources, at pH 7.2 (Maugeri et al., 2002). To evaluatethe capacity of strain L1T to use different substrates assole carbon sources, the medium M162 was supple-mented with 1–2% of (w/v) n-decane (BDH—PooleEngland); pentadecane (Aldrich Chemie); n-esade-cane (Schuchardt); toluene (Carlo Erba); SDS (Ap-plichem); EDTA (Carlo Erba); tridecane (EGA-CHEMIE); squalane (FLUKA). All growth tests weredone at 68°C and pH 7.2. The growth was scored aspositive if the 540 nm absorbance was greater than 0.3optical density, after 3 days of incubation.
Morphological and physiological studies. Cellularmorphology was determined by phase-contrast mi-croscopy (Zeiss) and by scanning electron microscopy(SEM). For SEM analysis the samples were fixed for24 h in 2.5% glutaraldehyde. Samples were dehy-drated in ethyl alcohol, dried to critical point, goldcoated by sputtering (SEM BALTECMED 020) and ob-served by a Philips XL 20 ESEM.
Phenotypic characterization was performed after L1T
incubation at 68°C for 3 days in TH medium or THagar (Hudson et al., 1986; Maugeri et al., 2002; San-tos et al., 1989).
Sensitivity of the strain to antibiotics was tested byusing the enrichment-solid medium TH and sensi-discs(6 mm, Oxoid). The following antibiotics were tested(mg): neomycin (5, 30), erythromycin (30), penicillin G
(10 U), chloramphenicol (10, 50), kanamycin (5, 30),tylosin (10, 30), ampicillin (25), gentamicin (10, 30),novobiocin (30), nystatin (100), cycloheximide (30),bacitracin (10 U), lincomycin (15), fusidic acid (10),vancomycin (30), streptomycin (25) and tetracycline(30, 50) (Nicolaus et al., 2000).
Biochemical analysis. For the enzymatic assays,cells grown in YN medium were collected during sta-tionary growth phase by centrifugation at 9,000�g for30 min. Wet cells (about 2.0 g) were suspended in20 mM Tris-HCl at pH 8.0, lysed by ultrasonic treatment(Heat System Instrument) for 4 min, and centrifuged at15,000�g for 20 min. The supernatant (crude extract)was assayed for a-galactosidase and a-glucosidaseenzymatic activities, by incubating for 10 min at 68°C areaction mixture containing in 1 ml final volume: 0.1 mlof the crude extract, 40 mM Tris-HCl pH 7.0 and 1 mM
of the following substrates: p-nitrophenyl-a-D-(glucopy-ranoside, galactopyranoside, maltoside, arabinoside);p-nitrophenyl-b -(glucopyranoside, xylopyranoside,galactopyranoside, lactopyranoside, maltoside); p-ni-trophenyl N-acetyl-b-D-glucosamide and 2-nitrophenyl-2-acetamido-2-deoxy a-glucopyranoside. The reactionwas stopped by adding 1 ml of 1 M Na2CO3 followed by3 ml of H2O. The released p-nitrophenol was mea-sured as optical density at 420 nm (Nicolaus et al.,1998). Protein content was determined by the Bradfordmethod using the BioRad protein assay with bovineserum albumin as standard (Bradford, 1976).Aminopeptidase activity was assayed with “BactidentAminopeptidase Kit” from Merck (Germany) accordingto the manufacturer’s specifications. Hydrolysis of N�-benzoyl-arginine-p-nitroanilide (BAPA) stereoisomerswas tested according to Oren and Galinski (1994).
Lipid and fatty acid compositions. Cells, grownboth in TH and hydrocarbon media, were harvested inthe late exponential growth phase by centrifugation at9,000�g. Freshly harvested cells (5–10 g) werelyophilized and extracted by Soxhlet with CHCl3/MeOH(1 : 1 by vol.) for 5 h at 70°C (Nicolaus et al., 2001).Lipid spots were analyzed by thin layer chromatogra-phy (TLC) on silica gel (0.25 mm, F254, Merck) elutedwith CHCl3/MeOH/H2O (65 : 25 : 4 by vol.). Lipids weredetected by spraying the plates with 0.1% Ce(SO4)2
followed by heating at 100°C for 5 min. Staining testsfor complex lipids were performed using specificreagents for phospho-, amino- and glycolipids. Thetotal lipid extract was treated with two volumes of n-hexane at 30°C for 12 h. The quinone content was an-
2006 Geobacillus toebii subsp. decanicus subsp. nov. 225

alyzed by high-performance liquid chromatography(HPLC) using an RP-18 Lichrospher (250�4 mm) col-umn eluted with n-hexane/ethylacetate (99 : 1 v/v) witha flow rate of 1.0 ml/min. Compounds were identifiedby 1H Nuclear Magnetic Resonance (NMR) and MassSpectroscopy (MS) as previously described. Lipid hy-drolysis was performed by acid methanolysis. Gas-Chromatography and Mass Spectroscopy (GC-MS)analyses were performed with an HP5890 series IIplus-5989B equipped with an HP-V column with a fluxof 45 ml/min. Fatty acid methyl esters were detectedusing the temperature program of 120°C (1 min), from120 to 250°C at 2°C/min. The identification of the com-pounds was performed by parallel runs of pure stan-dards (Sigma), and by interpretation of mass spectra.
Degradation of hydrocarbons. The evaluation ofgrowth rate and hydrocarbon hydrolysis for the L1T
strain was carried out at 68°C and pH 7.2 in the M162medium supplemented by 1–2% (w/v) n-decane or tri-decane by a 3 liter fermenter (Chemap), with low me-chanical agitation (100 rpm) and an aeration flux of20 ml min�1 for liter of broth. Cell density was esti-mated by measuring absorbance at 540 nm by directinsertion of culture tubes into DMS 90 VARIAN UV/VISspectrophotometer. Hydrocarbon depletion was evalu-ated using GC-MS and NMR analyses on cell free su-pernatants at interval times of growth. The super-natants were extracted with dichloromethane (v/v) anddried under vacuum. GC-MS was performed on aHewlett-Packard 5890-5970 instrument, equipped withan HP-V column. The analysis was performed at flowrate of 0.7 ml min�1; the temperature program usedwas: 60°C for 2 min, ramping from 60 to 280°C at 3°C min�1 and final step of 10 min at 280°C. NMRspectra were recorded on a Bruker AMX-500 instru-ment (500.13 MHz for 1H NMR) using CDCl3 as sol-vent for hydrocarbon samples.
Heavy metal resistance. The chemicals assessedfor toxicity were NiSO4·6H2O (Carlo Erba),ZnSO4·7H2O (Aldrich), Co(NO3)2· 6H2O (J.T. Baker),HgCl2 (J.T. Baker), MnCl2· 4H2O (J.T. Baker),Cr(NO3)3· 9H2O (J.T. Baker), K2Cr2O7 (J.T. Baker),CuSO4·5H2O (J.T. Baker), FeCl3 (Carlo Erba), CdSO4
(Aldrich). Solutions were prepared by dissolving in dis-tilled water and subsequently filtered. The chosenmetal concentrations were calculated as mg/L (ppmpart per million) of each metal (Poli et al., 2005).
Cells were grown by inoculating 90 ml media in a250-ml Erlenmeyer flask using the minimal medium
containing (1%) glucose as sole carbon source. Theculture was incubated for 12 h, after which the cellswere transferred to fresh media to get an optical den-sity of 0.1 (A540 nm). Each heavy metal was added tofresh media at appropriate concentrations at To (time).Bacterial growth controls and other samples werechecked by measuring spectrophotometrically at540 nm, using UV Spectrophotometer (Varian DMS90). Cells were harvested in late exponential growthphase by centrifugation at 9,000�g for 30 min.
Genetic studies. The almost complete 16S rRNAgene sequence was determined by direct sequencingof PCR-amplified 16S rDNA. Genomic DNA extraction,PCR mediated amplification of the 16S rDNA and pu-rification of PCR products were carried out as de-scribed previously (Rainey et al., 1996). Purified PCRproducts were sequenced using the ABI PRISMtm DyeTerminator Cycle Sequencing Ready Reaction Kit (Ap-plied Biosystems, Germany) as specified in the manu-facturer’s protocol. Sequence reactions were elec-trophoresed using the Applied Biosysthems 373A Se-quencer (Rainey et al., 1994). The resulting sequencedata were put into the alignment editor ae2 softwareand compared with representative 16S rDNA se-quences of organisms belonging to the Geobacillusgroup (Maidak et al., 1999). For comparison 16S rDNAsequences were obtained from the EMBL data base(Maidak et al., 1999). The 16S rDNA sequence ofstrain L1T has been deposited in EMBL databaseunder the accession number AJ966346. The results ofalignments are presented, in RESULTS AND DISCUSSION,as similarity matrix and phylogenetic tree (Saitou andNei, 1987). The 16S rDNA similarity values were cal-culated by pair-wise comparison of the sequenceswithin the alignment. For construction of the phyloge-netic dendrogram, the PHYLIP package was used(Felsenstein, 1993); pair-wise evolutionary distanceswere computed from percent similarities by the correc-tion of Jukes and Cantor (1969) and the phylogenetictree was constructed by the neighbour-joining method(Higgins et al., 1992).
The G�C content was determined by HPLC method(Mesbah et al., 1989; Tamaoka and Komagata, 1984).The calibration was performed with non-methylatedLambda-DNA (Sigma) (G�C content 49.85 mol%) andwith Halomonas pantelleriensis DNA (G�C content65.02 mol%).
Random Amplified Polymorphic DNA-PCR (RAPD-PCR) assay was used to produce fingerprint patterns
226 POLI et al. Vol. 52

of L1T and of reference strains Geobacillus toebii DSM14590T and Geobacillus caldoxylosilyticus ATCC42125T, according to Ronimus et al. (1997). DNA am-plification was performed in a 50 m l PCR reaction mix-ture containing: 50–200 ng of genomic DNA, 1� PCRbuffer (supplied as component of the DNA polymerasekit), 3 mM MgCl2, 250 mM dNTPs, 0.5 mM of OPR-2primer (5�-CACAGCTGCC-3�) or OPR-13 primer (5�-GGACGACAAG-3�) and 2.5 units of Platinum® TaqDNA polymerase (Invitrogen). The mixtures were am-plified in a thermocycler iCycler® (BIO RAD). The am-plification profile consisted of an initial denaturation of2 min at 92°C and 35 cycles of 15 s at 94°C, annealingfor 15 s at 36°C (previously optimized by temperaturegradient amplification) and elongation for 2 min at72°C. A final extension of 7 min was carried out at72°C. Ten to 20 m l of PCR products were elec-trophoresed on 2% agarose gel (Agarose-1000, Invit-rogen) in 1� TAE buffer at 5 V/cm for 4 h. Ethidiumbromide (0.1 mg/ml) was included both in the gel andelectrophoresis buffer and PCR products were de-tected by UV visualization and recorded on Polapan55 films (Polaroid).
DNA-DNA hybridization. DNA was isolated using aFrench pressure cell (Thermo Spectronic) and was pu-rified by chromatography on hydroxyapatite as de-scribed by Cashion et al. (1977). DNA-DNA hybridiza-tion was carried out as described by De Ley et al.(1970) under consideration of the modifications described by Huss et al. (1983) using a model Cary100 Bio UV/VIS-spectrophotometer equipped with aPeltier-thermostatted 6�6 multicell changer and atemperature controller with in situ temperature probe(Varian).
Results and Discussion
Compost parametersThe compost was obtained by using 50% of organic
urban waste and 50% green brush waste. Figure 1summarizes the flow-chart of the process. During thefermentation, the temperature ranged from 60 to 70°C,the oxygen concentration was 10–20% (v/v), the pHranged from 6.0 to 7.5 and the humidity decreasedfrom 60 to 20%. Chemical analysis of compost mate-rial was reported in Table 1. Zinc, Copper, and Leadwere the main heavy metal detected. Plastic materialwas found in a much lower amount. Among bacterialtests performed, only Enterobacteriaceae and Strepto-
2006 Geobacillus toebii subsp. decanicus subsp. nov. 227
Fig. 1. Flow-chart of composting process.
Table 1. Characteristics of hot compost from “Pomigliano Environment.”
Parameters UnityStandard Measured
values values
Cadmium mg/kg d.c. �1.5 1.34Chromium VI mg/kg d.c. �0.5 �0.5Mercury mg/kg d.c. �1.5 �1.0Nickel mg/kg d.c. �50 11.6Lead mg/kg d.c. �140 78.5Copper mg/kg d.c. �150 129.2Zinc mg/kg d.c. �500 256.9Plastics�3.33 mm % d.c. �0.45 �0.01Plastics�3.33 mm % d.c. �0.05 0.01Salmonella — Absent /25 g AbsentEnterobacteriaceae CFU/g �1�100 1,800Streptococcus faecalis NPM/g �1�1,000 200Nematode NPM/50 g Absent AbsentTrematode NPM/50 g Absent AbsentCestode NPM/50 g Absent AbsentInfestant seeds N Absent Absent
d.c., dry compost; CFU, colony forming units; NPM, numericponderable media; N, number.

coccus faecalis tests resulted positive (Table 1).
Characteristic of the isolatePhase-contrast microscopy observations of compost
samples show the presence of high numbers of ovalbacterial spores.
Bacterial growth occurred up to a 10�7 dilution of15% (w/v) compost in TH medium, and the strain L1T
was isolated from this dilution.Strain L1T exhibited morphological and chemical
characteristics that are consistent with those found inthe genus Geobacillus. Cells of strain L1T are aerobicand gram-positive motile rods, 2.0–3.0 mm long and0.5 mm wide (Fig. 2). Spores oval, located terminally,first appeared on medium TH with 2% MnCl2. Growthof strain L1T occurred at 55–75°C with an optimum of68°C (no growth was observed at 50°C and 80°C).Until now no strains of thermophilic bacilli isolated fromhot compost were able to grow at temperatures above70°C. At optimum temperature, growth occurred be-tween pH 5.0–9.0 with an optimum at pH 7.2.
The isolate was able to utilize a large variety of sug-ars and hydrocarbons. The bacterium utilized syntheticmedia and did not require any growth factors and vita-mins. Considerable growth was observed on TH agarand YN agar. The isolate was catalase, tyrosine de-composition, hippurate and gelatine hydrolysis, posi-tive. Isolate L1T was able to reduce NO3
� but not NO2�
and was sensible to lysozyme while it was oxidase, ca-sein and starch hydrolysis, indole production, phenyl-alanine deamination and urease negative.
Isolate L1T was negative for xylanase, b-glucosidase,a-mannosidase, a-amylase, esterase, b-galactosidaseactivities while it possessed a-galactosidase and a-glucosidase activities.
The isolate L1T utilized a wide range of carbonsources including ribose, glucose, glycerol, trehalose,maltose, cellobiose, ethanol and raffinose. The isolatewas sensitive to kanamycin (5 mg), bacitracin (10 U),novobiocin (30 mg), streptomycin (25 mg), tetracycline(30 mg) and penicillin G (10 U).
Lipid and fatty acid compositionsStrain L1T possessed complex lipids based on fatty
acid. The total lipid contents accounted for 10% and9% of dry weight for culture grown in TH medium andhydrocarbon media, respectively. Under these condi-tions three major phospholipids and one phosphogly-colipid were present. In particular, using n-decane
medium an additional phospholipid was detected. Thefatty acid compositions, determined from cells grown inTH medium or n-decane medium, were characterizedby the abundance of branched acyl chains; in particu-lar strain L1T contained major amounts of iso-C16:0(46%) and iso-C17 : 0 (28%) when cultured in THmedium, and iso-C16 : 0 (55%), iso-C17 : 0 (11%), n-C17 (16%) using the n-decane medium. The mem-brane lipids are characterized by an elevated numberof phospholipids composed of branched fatty acidsand by the presence of a quinone MK7 type. Iso-C16 : 0 and iso-C17 : 0 fatty acids were predominantwhile iso-C15 : 0 was absent, in contrast with the typi-cal fatty acid composition of bacteria from the genusGeobacillus. The pattern of the complex lipids has al-lowed to establish the taxonomic position of L1T by as-signing it to the Bacteria Domain. The phenotypiccharacteristics are coherent for strain L1T belonging tothe genus Geobacillus.
Growth on hydrocarbonsGrowth studies of strain L1T on hydrocarbon media
revealed its ability to utilize different hydrocarbon sub-strates such as pentadecane, tridecane, n-decane,squalane, and toluene, as unique carbon and energysources (Fig. 3). In particular n-decane supported effi-ciently the growth reaching 1 g/L of wet cells. Growthcurves of L1T strain in the TH and n-decane media arecompared (Fig. 4). The doubling time (td) of strain L1T
was 2 h in TH medium and 6 h using hydrocarbon sub-
228 POLI et al. Vol. 52
Fig. 2. Scanning electron microscopy of strain L1T grownon n-decane medium, in fermenter culture at 68°C.
The sample morphology is observed by using a SEM PhilipsXL 20 series microscope.
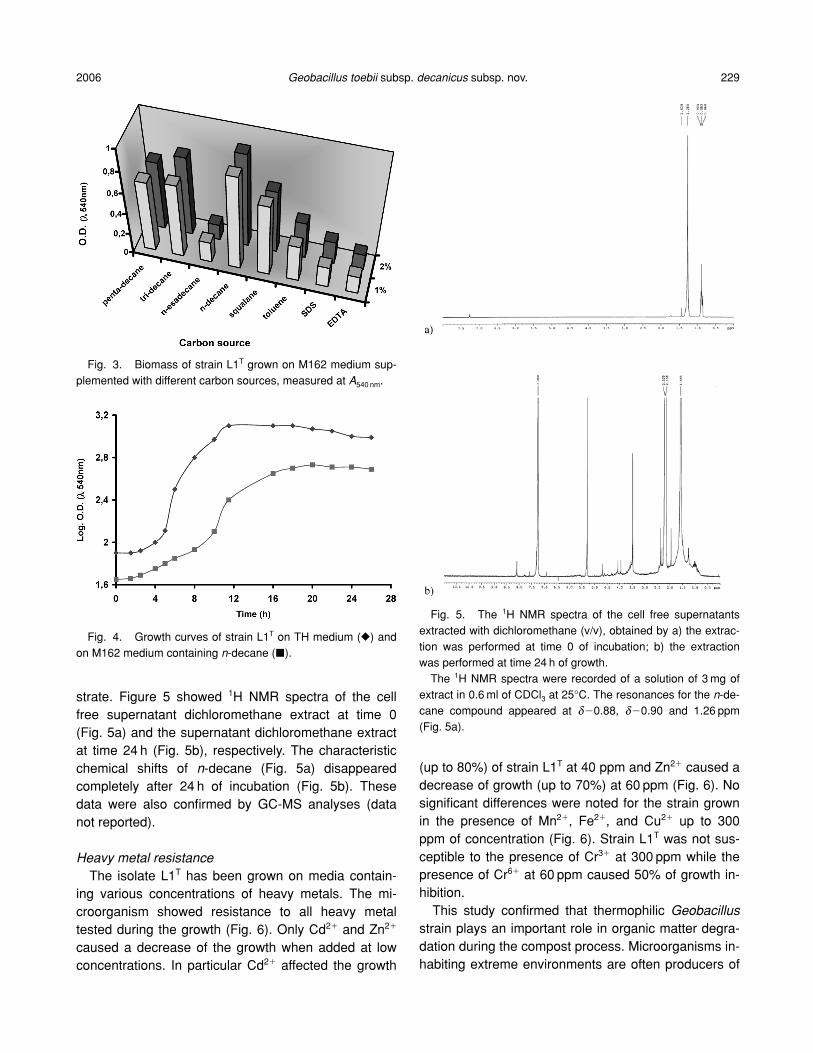
strate. Figure 5 showed 1H NMR spectra of the cellfree supernatant dichloromethane extract at time 0(Fig. 5a) and the supernatant dichloromethane extractat time 24 h (Fig. 5b), respectively. The characteristicchemical shifts of n-decane (Fig. 5a) disappearedcompletely after 24 h of incubation (Fig. 5b). Thesedata were also confirmed by GC-MS analyses (datanot reported).
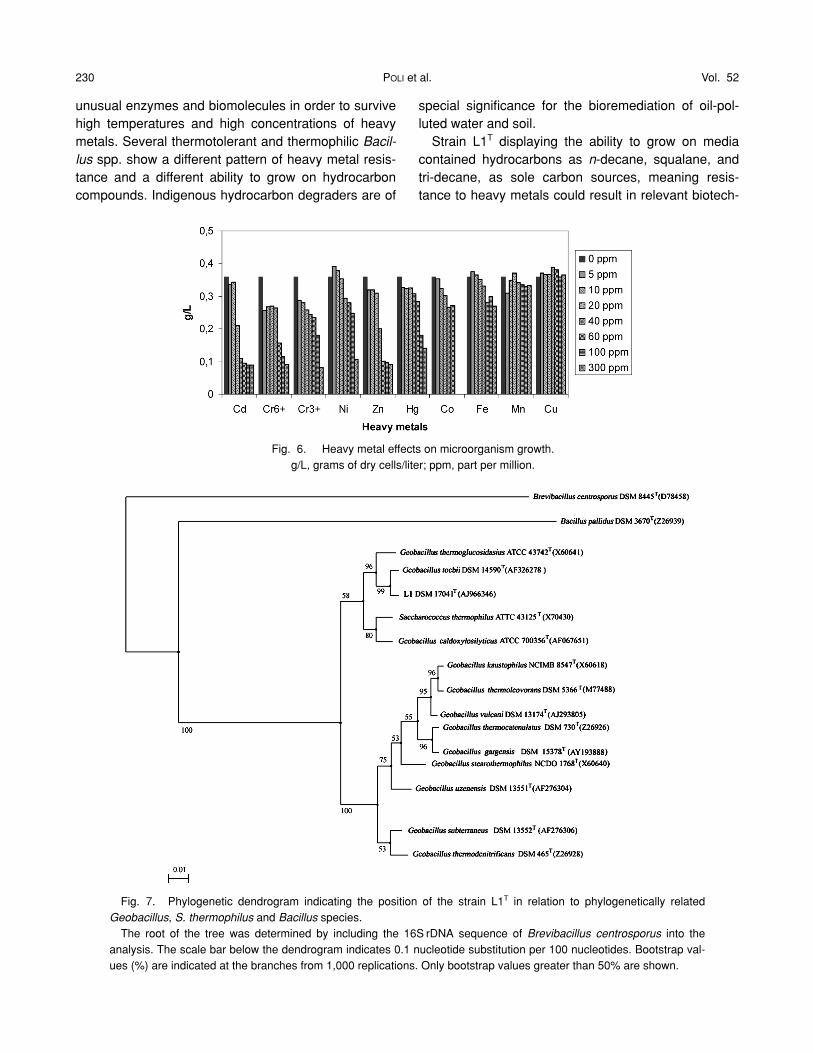
Heavy metal resistanceThe isolate L1T has been grown on media contain-
ing various concentrations of heavy metals. The mi-croorganism showed resistance to all heavy metaltested during the growth (Fig. 6). Only Cd2� and Zn2�
caused a decrease of the growth when added at lowconcentrations. In particular Cd2� affected the growth
(up to 80%) of strain L1T at 40 ppm and Zn2� caused adecrease of growth (up to 70%) at 60 ppm (Fig. 6). Nosignificant differences were noted for the strain grownin the presence of Mn2�, Fe2�, and Cu2� up to 300ppm of concentration (Fig. 6). Strain L1T was not sus-ceptible to the presence of Cr3� at 300 ppm while thepresence of Cr6� at 60 ppm caused 50% of growth in-hibition.
This study confirmed that thermophilic Geobacillusstrain plays an important role in organic matter degra-dation during the compost process. Microorganisms in-habiting extreme environments are often producers of
2006 Geobacillus toebii subsp. decanicus subsp. nov. 229
Fig. 3. Biomass of strain L1T grown on M162 medium sup-plemented with different carbon sources, measured at A540 nm.
Fig. 4. Growth curves of strain L1T on TH medium (�) andon M162 medium containing n-decane (�).
Fig. 5. The 1H NMR spectra of the cell free supernatantsextracted with dichloromethane (v/v), obtained by a) the extrac-tion was performed at time 0 of incubation; b) the extractionwas performed at time 24 h of growth.
The 1H NMR spectra were recorded of a solution of 3 mg ofextract in 0.6 ml of CDCl3 at 25°C. The resonances for the n-de-cane compound appeared at d�0.88, d�0.90 and 1.26 ppm(Fig. 5a).
unusual enzymes and biomolecules in order to survivehigh temperatures and high concentrations of heavymetals. Several thermotolerant and thermophilic Bacil-lus spp. show a different pattern of heavy metal resis-tance and a different ability to grow on hydrocarboncompounds. Indigenous hydrocarbon degraders are of
special significance for the bioremediation of oil-pol-luted water and soil.
Strain L1T displaying the ability to grow on mediacontained hydrocarbons as n-decane, squalane, andtri-decane, as sole carbon sources, meaning resis-tance to heavy metals could result in relevant biotech-
230 POLI et al. Vol. 52
Fig. 7. Phylogenetic dendrogram indicating the position of the strain L1T in relation to phylogenetically relatedGeobacillus, S. thermophilus and Bacillus species.
The root of the tree was determined by including the 16S rDNA sequence of Brevibacillus centrosporus into theanalysis. The scale bar below the dendrogram indicates 0.1 nucleotide substitution per 100 nucleotides. Bootstrap val-ues (%) are indicated at the branches from 1,000 replications. Only bootstrap values greater than 50% are shown.
Fig. 6. Heavy metal effects on microorganism growth.g/L, grams of dry cells/liter; ppm, part per million.

nological importance in industrial uses. One exampleis the possibility to use such microorganisms to cleanup oil spills in high temperature environments.
Genetic studiesComparison of 16S rDNA strain sequence in data
bank (EMBL AJ966346), revealed that strain L1T fallswithin the radiation of the cluster comprising Geobacil-lus species and forms a coherent cluster withGeobacillus toebii DSM 14590T and Geobacillus cal-doxylosilyticus ATCC 42125T (Fig. 7). The level of 16SrDNA sequence similarity between strain L1T and G.toebii SK-1T was 97.0%. The sequence similarity toother species within the genus Geobacillus (withvalidly published names) was less than 95.0%. TheG�C content of the DNA of strain L1T was 43.5 mol%.
RAPD-PCR was used to produce fingerprint pat-terns of L1T isolate and related strains, as described inMATERIALS AND METHODS. Both OPR-2 (Fig. 8, lines 1,2, 3) and OPR-13 (Fig. 8, lines 4, 5, 6) primers pro-duced fingerprints patterns characterized by relevantbands within 2,500–400 bp. L1T and G. toebii differedat level of bands 500 and 700 bp utilizing OPR-2primer and at level of band 2,500 bp using OPR-13primer. L1T and Geobacillus caldoxylosilyticus showeda different fingerprints using both primers.
The phylogenetic analysis (16S rDNA % of similarityand PCR-fingerprint) confirms that strain L1T belongsto the genus Geobacillus (Stackebrandt and Goebel,1994).
DNA-DNA hybridization experiments between strainL1T and G. toebii SK-1T, showing low 16S rDNA simi-larity level (97%), revealed a border line re-associationvalue between the new isolate with G. toebii SK-1T
(73.7%) and a lower value with Geobacillus caldoxy-losilyticus (55.9%) (Wayne et al., 1987).
Table 2 indicates the difference in the phenotypicand biochemical characteristics between strain L1T
and G. toebii. Strain L1T differed from G. toebii in opti-mal growth temperature, pH range, reduction of NO2
�,utilization of substrates, casein and gelatin hydrolysisand fatty acid compositions (Table 2).
On the basis of these results, we propose that ther-mophilic Geobacillus strain L1T is a novel subspeciesof Geobacillus toebii, for which the name Geobacillustoebii subsp. decanicus is proposed.
2006 Geobacillus toebii subsp. decanicus subsp. nov. 231
Fig. 8. RAPD-PCR fingerprint patterns of L1T and refer-ence strains.
PCR analysis was performed as described in MATERIALS AND
METHODS by OPR-2 primer (5�-CACAGCTGCC-3�) (lanes 1–3)or OPR-13 primer (5�-GGACGACAAG-3�)(lanes 4–6). MW, 100bp DNA ladder (Invitrogen); lanes 1 and 4, Geobacillus toebiiDSM 14590T; lanes 2 and 5, strain L1T; lanes 3 and 6,Geobacillus caldoxylosilyticus ATCC 42125T.
Table 2. Comparison of the phenotypic characteristics ofstrain L1T with related species.
Characteristic Strain L1Geobacillus toebii
strain SK-1a
Temperature range (°C) 60–70 45–70Optimum temperature (°C) 68 60pH range 5.0–9.0 6.0–9.0Gelatin hydrolysis � �
Casein hydrolysis � �
Fermentation of glucose � �
Denitrification � �
G+C content (mol%) 43.5 44.0FAME composition (%):nC15 : 0 6.2 �
iC15 : 0 � 34.03iC16 : 0 46.1 17.46nC16 : 0 8.6 �
iC17 : 0 28.1 34.86nC17 : 0 6.1 �
nC18 : 0 4.8 �
Geobacillus toebii strain SK-1T.a Data were obtained from Sung et al. (2002).

Description of Geobacillus toebii subsp. decanicussubsp. nov.
G. toebii subsp. decanicus (de. ca. ni. cus. N.L. n.decane common name for hydrocarbon; able to growon n-decane; N.L. masc. adj. decanicus pertaining ton-decane).
Gram positive, aerobic, motile, spore forming bac-terium, 2.0–3.0 mm long and 0.5 mm wide, growth oc-curs at 55–75°C with optimum growth occurring at68°C. It grows at pH 5.0–9.0 (optimal pH 7.2). Growson M162 mineral medium up to and in 1–2% n-dec-ane, tridecane, pentadecane, or squalane as sole car-bon sources. It is able to grow on glucose, ribose, tre-halose, glycerol, maltose, cellobiose, ethanol, raffi-nose. It is resistant to different heavy metals addedduring the growth: Cu2�, Mn2�, Fe2�, Co2�, Hg2� andNi2�. It is catalase, tyrosine decomposition, hippurateand gelatine hydrolysis positive, reduces NO3
� to NO2�;
it is sensitive to lysozyme, negative for oxidase, starchand casein hydrolysis, urease, indole production,phenylalanine deamination. Positive results are ob-tained for a-galactosidase and a-glucosidase activi-ties. MK7 is the predominant quinone. iC16 : 0 andiC17 : 0 are the major fatty acids (74% of total fattyacids). The following antibiotics inhibit the growth:kanamycin (5 mg), bacitracin (10 U), novobiocin (30 mg),streptomycin (25 mg), tetracycline (30 mg) and penicillinG (10 U). The mol% GC content of DNA is 43.5 mol%.The EMBL accession number for the 16S rDNA se-quence of strain L1T is AJ 966346. Type strain is L1T
(DSM 17041T�ATCC BAA 1004T). Isolated from hotcompost “Pomigliano Environment” s.p.a., Pomigliano,Naples, Italy.
Acknowledgments
This work was partially supported by Regione Campania. Wethank Valeria Calandrelli and Eduardo Pagnotta for technicalassistance, Ottavio De Luca for LC-MS analyses, VincenzoMirra and Salvatore Zambardino for NMR service.
References
Ahmad, S., Scopes, R. K., Rees, G. N., and Patel, B. K. C.(2000) Saccharococcus caldoxylosilyticus sp. nov., an ob-ligately thermophilic, xylose-utilizing, endospore-formingbacterium. Int. J. Syst. Evol. Microbiol., 50, 517–523.
Ash, C., Farrow, J. A. E., Wallbanks, S., and Collins, M. D.(1991) Phylogenetic heterogeneity of the genus Bacillusrevealed by comparative analysis of small-subunit-riboso-mal RNA. Lett. Appl. Microbiol., 13, 202–206.
Beffa, T., Blanc, M., Lyon, P. F., Vogt, G., Marchiani, M., Fisher,J. L., and Aragno, M. (1996) Isolation of Thermus strainfrom hot composts (60–80°C). Appl. Environ. Microbiol.,62, 1723–1727.
Blanc, M., Marilley, L., Beffa, T., and Aragno, M. (1999) Ther-mophilic bacterial communities in hot composts as re-vealed by most probable number counts and molecular(16S rDNA) methods. FEMS Microbiol. Ecol., 28, 141–149.
Bradford, M. M. (1976) A rapid and sensitive method for quanti-tation of microgram quantities of protein using the princi-ples of protein-dye binding. Anal. Biochem., 72, 248–254.
Caccamo, D., Gugliandolo, C., Stackebrandt, E., and Maugeri,T. L. (2000) Bacillus vulcani sp. nov., a novel thermophilicspecies isolated from a shallow marine hydrothermal vent.Int. J. Syst. Evol. Microbiol., 50, 2009–2012.
Cashion, P., Hodler-Franklin, M. A., McCully, J., and Franklin,M. (1977) A rapid method for base ratio determination ofbacterial DNA. Anal. Biochem., 81, 461–466.
Chen, X. G., Stabnikova, O., Tay, J. H., Wang, J. Y., and Tay, S.T. L. (2004) Thermoactive extracellular proteases ofGeobacillus caldoproteolyticus, sp. nov., from sewagesludge. Extremophiles, 8, 489–498.
Claus, D. and Berkeley, R. C. W. (1986) Genus Bacillus Cohn1872, 174AL. In Bergey’s Manual of Systematic Bacteriol-ogy, ed. by Sneath, P. H. A., Mair, N. S., Sharpe, M. E.,Holt, J. G., Williams & Wilkins, Baltimore, Vol. 2, pp.1105–1139.
De Ley, J., Cattoir, H., and Reynaerts, A. (1970) The quantita-tive measurement of DNA hybridization from renaturationrates. Eur. J. Biochem., 12, 133–142.
Felsenstein, J. (1993) PHYLIP Phylogeny Inference Package,version 3.5.1. Department of Genetics, University of Wash-ington, Seattle, USA.
Fortina, M. G., Mora, D., Schumann, P., Parini, C., Manichini, P.L., and Stackebrandt, E. (2001a) Reclassification of Sac-charococcus caldoxylosilyticus as Geobacillus caldoxylosi-lyticus (Ahmad et al. 2000) comb. nov. Int. J. Syst. Evol. Mi-crobiol., 51, 2063–2071.
Fortina, M. G., Pukall, R., Schumann, P., Mora, D., Parini, C.,Manichini, P. L., and Stackebrandt, E. (2001b) Ureibacillusgen. nov., a new genus to accommodate Bacillus thermo-sphaericus (Andersson et al. 1995), emendation of Urei-bacillus thermosphaericus and description of Ureibacillusterrenus sp. nov. Int. J. Syst. Evol. Microbiol., 51, 447–455.
Fugio, Y. and Kume, S. J. (1991) Isolation and identification ofthermophilic bacteria from sewage sludge compost. J. Fer-ment. Bioeng., 72, 334–337.
Golovacheva, R. S., Loginova, L. G., Salikhov, T. A.,Kolesnikov, A. A., and Zaitseva, G. N. (1975) A new ther-mophilic species Bacillus thermocatenulatus sp. nov. Micro-biology (English translation of Mikrobiologiya), 44, 230–233.
Gugliandolo, C., Maugeri, T., Caccamo, D., and Stackebrandt,E. (2003) Bacillus aeolius sp. nov. a novel thermophilichalophilic marine Bacillus species from Eolian Islands
232 POLI et al. Vol. 52

(Italy). Syst. Appl. Microbiol., 26, 172–176.Higgins, D. G., Bleasby, A. T., and Fuchs, R. (1992) Clustal W:
improved software for multiple sequence alignment.CABIOS, 8, 189–191.
Hudson, J. A., Morgan, H. W., and Daniel, R. M. (1986) A nu-merical classification of some Thermus isolates. J. Gen.Microbiol., 13, 532–540.
Huss, V. A. R., Festl, H., and Schleifer, K. H. (1983) Studies onthe spectrophotometric determination of DNA hybridizationfrom renaturation rates. Syst. Appl. Microbiol., 4, 184–192.
Jukes, T. H. and Cantor, C. R. (1969) Evolution of protein mole-cules. In Mammalian Protein Metabolism, ed. by Munro, H.N., Academic Press, New York, pp. 21–132.
Koneman, E. W., Allen, S. D., Dowell, V. R., and Sommers, H.M. (1984) Colour Atlas and Textbook of Diagnostic Microbi-ology, ed. by Lippincott JB Company, Philadelphia, Toronto.
Logan, N. A. and Berkeley, R. C. W. (1984) Identification ofBacillus strains using the API system. J. Gen. Microbiol.,130, 1871–1882.
Lyon, P. F., Beffa, T., Blanc, M., Auling, G., and Aragno, M.(2000) Isolation and characterization of highly thermophilicxylanolytic Thermus thermophilus strains from hot com-posts. Can. J. Microbiol., 46, 1029–1035.
Maidak, B. L., Cole, J. R., Parker, C. T., Garrity, G. M., Larsen,N., Li, B., Liburn, T. G., McCaughey, M. J., Olsen, G. J.,Overbeek, R., Pramanik, S., Schmidt, T. M., Tiedje, J. M.,and Woese, C. R. (1999) A new version of the RDP (Ribo-somal Database Project). Nucleic Acids Res., 27, 171–173.
Manichini, P. L., Mora, D., Nicastro, G., Parini, C., Stackebrandt,E., Pukall, R., and Fortina, M. G. (2000) Bacillus thermo-denitrificans sp. nov., nom. rev. Int. J. Syst. Evol. Microbiol.,50, 1331–1337.
Maugeri, T. L., Gugliandolo, C., Caccamo, D., and Stacke-brandt, E. (2001) A polyphasic taxonomic study of ther-mophilic bacilli from shallow marine vents. Syst. Appl. Mi-crobiol., 24, 572–587.
Maugeri, T. L., Gugliandolo, C., Caccamo, D., Panico, A., Lama,L., Gambacorta, A., and Nicolaus, B. (2002) A halophilicthermotolerant Bacillus isolated from a marine hot springable to produce a new exopolysaccharide. Biotechnol.Lett., 24, 515–519.
Mesbah, M., Premachandran, U., and Whitman, W. (1989) Pre-cise measurement of the G�C content of deoxyribonucleicacid by high performance liquid chromatography. Int. J.Syst. Bacteriol., 39, 159–167.
Miller, F. C. (1996) Composting of municipal solid waste and itscomponents. In Microbiology of Solid Waste, ed. byPalmisano, A. C. and Barlaz, N. A., CRC Press, BocaRaton, Fla., pp. 115–154.
Nazina, T. N., Lebedeva, E. V., Poltaraus. A. B., Tourova, T. P.,Grigoryan, A. A., Sokolova, D. S., Lysenko, A. M., and Os-ipov, G. A. (2004) Geobacillus gargensis sp. nov., a novelthermophile from a hot spring, and the reclassification ofBacillus vulcani as Geobacillus vulcani (Caccamo et al.,2000) comb. nov. Int. J. Syst. Evol. Microbiol., 54, 2019–
2024.Nazina, T. N., Sokolova, D. S., Grigoryan, A. A., Shestakova, N.
M., Mikhailova, E. M., Poltaraus, A. B., Tourova, T. P., Ly-senko, A. M., Osipov, G. A., and Belyaev, S. S. (2005)Geobacillus jurassicus sp. nov., a new thermophilic bac-terium isolated from a high-temperature petroleum reser-voir and validation of the Geobacillus species. Syst. Appl.Microbiol., 28, 43–53.
Nazina, T. N., Tourova, T. P., Poltaraus, A. B., Novikova, E. V.,Grigoryan, A. A., Ivanova, A. E., Lysenko, A. M., Petrun-yaka, V. V., Osipov, G. A., Belyaev, S. S., and Ivanov, M. V.(2001) Taxonomic study of aerobic thermophilic bacilli: De-scription of Geobacillus subterraneus gen. nov., sp. nov.and Geobaacillus uzenensis sp. nov. from petroleum reser-voirs and transfer of Bacillus stearothermophilus, Bacillusthermocatenulatus, Bacillus thermoleovorans, Bacilluskaustophilus, Bacillus thermoglucosidasius and Bacillusthermodenitrificans to Geobacillus as the new combinationsG. stearothermophilus, G. thermocatenulatus, G. ther-moleovorans, G. kaustophilus, G. thermoglucosidasius andG. thermodenitrificans. Int. J. Syst. Evol. Microbiol., 5,433–446.
Nicolaus, B., Improta, R., Manca, M. C., Lama, L., Esposito, E.,and Gambacorta, A. (1998) Alicyclobacilli from an unex-plored geothermal soil in Antarctica: Mount Rittmann. PolarBiol., 19, 133–141.
Nicolaus, B., Lama, L., Esposito, E., Manca, M. C., Di Prisco,G., and Gambacorta, A. (1996) “Bacillus thermoantarcticus”sp. nov., from Mount Melbourne, Antarctica: A novel ther-mophilic species. Polar Biol., 16, 101–104.
Nicolaus, B., Manca, M. C., Lama, L., Esposito, E., and Gamba-corta, A. (2001) Lipid modulation by environmentalstresses in two models of extremophiles isolated fromAntarctica. Polar Biol., 24, 1–8.
Nicolaus, B., Panico, A., Manca, M. C., Lama, L., Gambacorta,A., Maugeri, T., Gugliandolo, C., and Caccamo, D. (2000) Athermophilic Bacillus isolated from an eolian shallow hy-drothermal vent, able to produce exopolysaccharides. Syst.Appl. Microbiol., 23, 426–432.
Nystrand, R. (1984) Saccharococcus thermophilus gen. nov.,sp. nov. isolated from beet sugar extraction. Syst. Appl. Mi-crobiol., 5, 204–219.
Oren, A. and Galinski, E. A. (1994) Hydrolysis of N�-benzoyl-arginine-p-nitroanilide stereoisomers as a phenotypic test:A study of Gram-positive halotolerant bacteria. Syst. Appl.Microbiol., 17, 710–716.
Poli, A., Esposito, E., Lama, L., Pelliccione, S., Gambacorta, A.,and Nicolaus, B. (2006) Heavy metal effects on Ex-tremophiles and on enzyme biosynthesis in a new Bacillusstrain from Mount Rittmann, Antarctica. In Modern Multidis-ciplinary Applied Microbiology, ed. by Mendez-Vilas, A.,Wiley-VCH Verlag GmbH & Co. KgaA, Weinheim, pp.83–88.
Priest, F. G., Goodfellow, M., and Todd, C. (1988) A numericalclassification of the genus Bacillus. J. Gen. Microbiol., 134,
2006 Geobacillus toebii subsp. decanicus subsp. nov. 233

1847–1882.Rainey, F. A., Fritze, D., and Stackebrandt, E. (1994) The phylo-
genetic diversity of thermophilic members of the genusBacillus as revealed by 16S rDNA analysis. FEMS Micro-biol. Lett., 115, 205–212.
Rainey, F. A., Ward-Raney, N., Kroppenstedt, R. M., andStackebrandt, E. (1996) The genus Nocardiopsis repre-sents a phylogenetically coherent taxon and a distinctactinomycete lineage; Proposal of Nocardiopsaceae fam.nov. Int. J. Syst. Bacteriol., 46, 1088–1092.
Rhee, S. K., Jeon, C. O., Bae, J. W., and 9 other authors.(2002) Characterization of Symbiobacterium toebii, an ob-ligate commensal thermophile isolated from compost. Ex-tremophiles, 6, 57–64.
Ronimus, R. S., Parker, L. E., and Morgan, H. W. (1997) Theutilization of RAPD-PCR for identifying thermophilic andmesophilic Bacillus species. FEMS Microbiol. Lett., 147,75–79.
Saitou, N. and Nei, M. (1987) The neighbour-joining method: Anew method for reconstructing phylogenetic trees. Mol.Biol. Evol., 4, 406–425.
Santos, M. A., Williams, R. A. D., and da Costa, M. S. (1989)Numerical taxonomy of Thermus isolates from hot spring inPortugal. Syst. Appl. Microbiol., 12, 310–315.
Stackebrandt, E. and Goebel, B. M. (1994) Taxonomic note: Aplace for DNA-DNA reassociation and 16S rRNA sequenceanalysis in the present species definition in bacteriology.Int. J. Syst. Bacteriol., 44, 846–849.
Strom, P. F. (1985a) Effect of temperature on bacterial speciesdiversity in thermophilic solid-waste composting. Appl. En-viron. Microbiol., 50, 899–905.
Strom, P. F. (1985b) Identification of thermophilic bacteria insolid-waste composting. Appl. Environ. Microbiol., 50,906–913.
Sung, M. H., Kim, H., Bae, J. W., Rhee, S. K., Jeon, C. O., Kim,K., Kim, J. J., Hong, S. P., Lee, S. G., Yoon, J. H., Park, Y.H., and Baek, D. H. (2002) Geobacillus toebii sp. nov., anovel thermophilic bacterium isolated from hay compost.Int. J. Syst. Evol. Microbiol., 52, 2251–2255.
Sunna, A., Tokajian, S., Burghardt, J., Raney, F., Antranikian,G., and Hashwa, F. (1997) Identification of Bacilluskaustophilus, Bacillus thermocatenulatus and BacillusStrain HSR as members of Bacillus thermoleovorans. Syst.Appl. Microbiol., 20, 232–237.
Suzuki, Y., Kishigami, T., Inoue, K., Mizoguchi, Y., Eto, N., Taka-gi, M., and Abe, S. (1983) Bacillus thermoglucosidasius sp.nov., a new species of obligately thermophilic bacilli. Syst.Appl. Microbiol., 4, 487–495.
Tamaoka, J. and Komagata, K. (1984) Determination of DNAbase composition by reversed-phase-high-performance liq-uid chromatography. FEMS Microbiol. Lett., 25,125–128.
Tomita, K., Ikeda, N., and Ueno, A. (2003) Isolation and charac-terization of a thermophilic bacterium, Geobacillus thermo-catenulatus, degrading nylon 12 and nylon 66. Biotechnol.Lett., 25, 1743–1746.
Wayne, L. G., Brenner, D. J., Colwell, R. R., Grimont, P. A. D.,Kandler, O., Krichevsky, M. I., Moore, L. H., Moore, W. E.C., Murray, R. G. E., Stackebrandt, E., Starr, M. P., andTrupper, H. G. (1987) Report of the Ad Hoc Committee onreconciliation of approaches to bacterial systematics. Int. J.Syst. Bacteriol., 37, 463–464.
White, D., Sharp, R. J., and Priest, F. G. (1993) A polyphasictaxonomic study of thermophilic bacilli from a wide geo-graphical area. Antonie van Leeuwenhoek, 64, 357–386.
Zarilla, K. A. and Perry, J. J. (1987) Bacillus thermoleovorans,sp. nov., a species of obligately thermophilic hydrocarbonutilizing endospore-forming bacteria. Syst. Appl. Microbiol.,9, 258–264.
234 POLI et al. Vol. 52





























