Embed Size (px)
Citation preview

Running head: UA/SA of a food web bioaccumulation model 1
2
3
4
5
6
7
8
9
10
11
12
Corresponding author: 13
Stefano Ciavatta 14
Department of Physical-Chemistry, University of Venice, 15
Dorsoduro 2137, 16
30123 Venice, Italy 17
Phone: +39 041 234 8632 18
Fax: +39 041 234 8594 19
e-mail: [email protected] 20
Word count (text and references only): 9398 21

GLOBAL UNCERTAINTY AND SENSITIVITY ANALYSIS OF A FOOD WEB 22
BIOACCUMULATION MODEL 23
24
Stefano Ciavatta†,‡*, Tomas Lovato§, Marco Ratto|| and Roberto Pastres‡ 25
26
†EuroMediterranean Centre for Climate Change (CMCC) c/o Consorzio Venezia Ricerche, 27
Via della Libertà 12, 30175 Venezia, Italy 28
‡ Dept. of Physical-Chemistry, University of Venice, Dorsoduro 2137, 30123 Venezia, Italy 29
§ Dept. of Environmental Sciences, University of Venice, Dorsoduro 2137, 30123 Venezia, 30
Italy 31
|| European Commission, Joint Research Centre, TP361 Via Fermi 1, 21027 Ispra (VA), Italy 32

* Corresponding author: 33
Stefano Ciavatta 34
Dept. of Physical-Chemistry, University of Venice, 35
Dorsoduro 2137, 36
30123 Venezia, Italy 37
Phone: 0039 041 234 8632 38
Fax: 0039 041 234 8594 39
e-mail: [email protected] 40

Abstract 41
42
A global Uncertainty and Sensitivity Analysis (UA/SA) of a state of the art food web 43
bioaccumulation model was carried out. We used an efficient screening analysis technique to 44
identify the subset of the most relevant input factors among the whole set of 227 model 45
parameters. A quantitative UA/SA was then applied to this subset, in order to rank the 46
parameters and to partition the model output variance among them, by means of a non-linear 47
regression of the outcomes of one thousand Monte Carlo simulations. The concentrations of 48
four representative persistent organic pollutants (POPs) in two representative species of the 49
coastal marine food-web of the Lagoon of Venice, Italy, were taken as model outputs. 50
The screening analysis showed that the ranking was remarkably different in relation to the 51
species and chemical being considered. The subsequent Monte Carlo based quantitative 52
analysis showed that the relationships among some of the parameters and the model outputs is 53
non linear. The non linear regression showed that the fraction of output variance accounted 54
for by each parameter strongly depends upon the range of the octanol/water partition 55
coefficient (KOW) values which are being considered. In fact, for the low hydrophobic 56
chemicals, the main sources of model uncertainty were the parameters related to the 57
respiratory bioaccumulation, while, for the more hydrophobic chemicals, KOW and other 58
parameters related to the dietary uptake explained the largest fractions of the variance of the 59
organism concentrations of the chemicals. 60
The analysis pointed out that efforts are still needed for reducing uncertainty of model 61
parameters to get reliable results from the application of food web bioaccumulation models. 62
63
Key words: 64
non linear analysis, persistent organic pollutants, coastal food web, Lagoon of Venice 65
66
INTRODUCTION 67
Food-web bioaccumulation in aquatic systems represents a relevant issue in environmental 68
toxicology and risk assessment and, more recently, a focus of regulatory efforts, because of 69
the risks that persistent organic pollutants (POPs) may pose to humans and environment [1]. 70
Modelling tools of increasing complexity have been proposed since the 1970s for the 71
quantitative assessments of the concentration of chemicals in the environment and in the 72
organisms and at present, food web bioaccumulation models are regarded as reliable research 73
and management tools [2,3]. 74

These models allow one to take into account the contaminant migration and 75
biomagnification through the food web, due to the feeding habits of the most important 76
species. They usually include a large number of parameters, which are determined by means 77
of either complex laboratory, site-specific measurements or empirical relationships and 78
therefore are inherently uncertain, leading to uncertainties in estimates of chemical 79
concentrations in the target species[2,3]. 80
Thus, Uncertainty and Sensitivity analyses (UA/SA) of model outputs with respect to the 81
model parameters has become a quite common practice in evaluating the uncertainty in the 82
response of food web bioaccumulation models and both local and global analysis have been 83
applied [4]. Local methods are based on the assumption of a linear relationship among the 84
model parameters and model outputs [5-7]. In the context of bioaccumulation modelling In 85
most applications, they have been applied by changing the nominal values of the model 86
parameters one-at-a-time, by a small fixed amount, and computing the corresponding changes 87
of the model output (see for example [8-10]). Global UA/SA methods allow one to assess the 88
overall uncertainty in the model output, due to the simultaneous variation of the model 89
parameters (or "input factors") within their whole range of variability, accordingly to their 90
probability density functions (pdfs) [7]. MacLeod et al. [4] pointed out that the global “Monte 91
Carlo approach is virtually synonymous with uncertainty analysis in the field of chemical fate 92
assessment“ [4], since its application in Gobas [11] up to the recent work of Nfon and 93
Cousins [12]. Nevertheless, the Monte Carlo approach to UA/SA may pose difficulties, due to 94
both the very high number of model parameters and to the lack of knowledge about the pdfs 95
of the parameters [4]. In order to overcome these difficulties, MacLeod et al. [4] propose a 96
methodological approach were a local sensitivity screening method – based on a linear 97
approximation of the model equations by means of a Taylor series expansion - is applied in 98
order to identify the few most relevant parameters to be eventually included in a subsequent 99
UA. The variance of the output of the Monte Carlo trials is then usually partitioned among the 100
input factors by using linear methods, such as the rank correlation coefficients technique [4, 101
12]. Saloranta et al. [13] proposed to carry out UA/SA using an alternative global method, 102
namely the Fourier Amplitude Sensitivity Test, which was applied for evaluating the 103
sensitivity and the uncertainty of the bioaccumulation model proposed by Gobas in 1993 [11], 104
with respect to 15 arbitrarily pre-selected model parameters. 105
The aim of the present paper is to estimate the uncertainty of the state-of-the-art Arnot and 106
Gobas food web bioaccumulation model [14] associated to the uncertainty in the model 107
parameters and to partition the model output variance among them. To this aim, we propose 108

the application of a global UA/SA approach which accounts for the non linear relationships 109
among the parameters and the model outputs. Thus, we identified the subset of the most 110
relevant parameters by means of a preliminary screening analysis conducted by means of the 111
Morris method [15], as improved by Campolongo et al. [16]. The ranking and the contribution 112
of these parameters to the output variance were subsequently estimated by fitting a non linear 113
"State Dependent" regression model [17] to a Monte Carlo input-output sample, accordingly 114
to the UA/SA method proposed by Ratto et al. [18]. We selected as model outputs the 115
concentrations of four representative POPs, ranging from low up to very hydrophobic 116
chemicals, in two important species of a coastal marine food-web in the Lagoon of Venice, 117
Italy. In order to untie the UA/SA from site specific conditions, the model forcing functions 118
were kept constant to values representative of a temperate lagoon and the pdfs of the model 119
parameters were defined on the basis of a literature review. 120
121
METHODS 122
123
The model 124
125
The model analyzed in the present paper is the food web bioaccumulation model proposed 126
by Arnot and Gobas [14] and updated in Gobas and Arnot [2]. 127
The model mass-balance equation is based on the steady state assumption of a time 128
invariant concentration of the chemical in the species i of the food web [8]: 129
[ ] [ ]MGEiDDSWDPOWDiB kkkkCkCmCmkC +++⋅+⋅+⋅⋅= 2,,,01, /)( (1) 130
In Equation 1, CB,i (g/kg), CWD,O (g/L), CWD,S (g/L) and CD,i (g/kg) are the dissolved 131
concentrations of the chemical in the species i, in the water column, in the sediment 132
associated pore water and in the diet, respectively, while mO and mP are the fractions of the 133
respiratory ventilation that involve the overlying and pore water and 134
SWDPOWDWR CmCmC ,,0 ⋅+⋅= can be defined as the overall freely dissolved chemical 135
concentration in the water involved in the respiratory ventilation (g/L). The remaining 136
parameters characterize the uptake and loss routes, namely: k1 (L/kgּd) and kD (kg/kgּ d) are 137
the clearance rates through respiration and diet, while the terms k2, kE, kG and kM (d-1) 138
represent the loss rates for respiration, excretion, growth dilution and biotransformation, 139
respectively. In this application, as in [2, 14], the parameter kM was set to zero since we 140
considered non metabolized chemicals. 141

The model parameters are listed in Table 1, while the environmental model forcings are 142
listed in Table 2. Coherently with the original model formulation [14], the freely dissolved 143
concentration CWD,O in Equation 1 was estimated on the basis of the partitioning equilibrium 144
between the particulate and dissolved organic carbon (POC and DOC, respectively) and the 145
total concentration in water [2, 14]: 146
)1(
,,
DOCOWDOCPOCOWPOC
OWTOWD KK
CC
χαχα ++= (2) 147
where CWT,O is the total concentration of the chemical in the water (g/L), KOW is the octanol-148
water partitioning constant of the chemical, χPOC and χDOC are the POC and DOC 149
concentrations in water (kg/L) while αPOC and αDOC are dimensionless constants describing 150
the similarity in phase partitioning of POC and DOC, respectively, in relation to that of 151
octanol. 152
The concentrations in the pore water and in the water column were not assumed to be in 153
equilibrium. The freely dissolved concentration in pore water CWD,S in Equation 1 was 154
estimated from the concentration of the chemical in the sediment, by using the partitioning 155
equation proposed by Gobas and MacLean [19]: 156
,sOWOCs
sSWD KOC
CC
δα= (3) 157
where Cs is the chemical concentration in the sediment (g/kg), OCs and δs are, respectively, 158
the organic carbon fraction in the sediment and the sediment density, αOC is a constant which 159
represents the ratio between the sorption capacity of the organic carbon and that of octanol. 160
The dependence of KOW on water temperature, an environmental model forcing, was here 161
represented by means of the Van’t Hoff equation, as recently proposed in Gewurtz et al [10] 162
and Nfon and Cousins [12]: 163
−⋅
∆+=
w
OWOWwOW TR
KLogTKLog1
15.298
1
)10ln(
H )( 25, (4) 164
where Tw is the water temperature (K), KOW,25 is the value of the octanol-water partitioning 165
constant at TW =298.15K, R is the universal gas constant and ∆HOW is the internal enthalpy of 166
phase transfer of the chemical between octanol and water (J K-1 mol-1). 167
Accordingly to Equations 1 to 4, the fate of a contaminant is driven by its physical-168
chemical properties, characterized by the parameters X1 - X5 in Table 1, and by the 169
environmental forcing functions OCs, POC and DOC. Two more physical-chemical 170
parameters, X6=β and X7=βa in Table 1, characterize the partitioning of chemicals among 171

octanol/non-lipid organic matter and octanol/non-lipid organic carbon in heterotrophic and 172
autotrophic species, respectively. 173
In the Arnot and Gobas model (1), several other equations describe the physiological and 174
ecological processes driving the route of the contaminant throughout the organisms of the 175
site-specific food-web, in relation to the biological and dietary characteristics of the particular 176
species or trophic compartment [14]. These equations are characterized by the species-177
specific parameters - or compartment-specific parameters - from X8 to X23 in Table 1. In the 178
present paper, however, the dilution rate for heterotrophic species growth, kg (d-1) in Equation 179
1, was computed by means of: 180
-0.2i
20)-(T )w(1000(1.113)0.00586 w ⋅⋅⋅=gk (5) 181
as recently proposed in [2,10], where Tw is the water temperature and wi is the weight of the 182
organism (kg). For autotrophic species i, kg needs to be parameterized – parameter X21,i= kg,i 183
in Table 1 – and defined on the basis of site-specific measurements [14]. 184
Not all the species-specific parameters are independent. In particular, the values of the 185
water fraction in the heterotrophic species i (νw,i) are calculated as a function of the lipid (νl,i) 186
and nonlipid organic matter (νn,i) fractions in organisms [2]: 187
νw,i=1-(νl,i+νn,i) (6a) 188
while for autotrophic species the nonlipid organic carbon (νn’,i) fraction is considered: 189
νw,i=1-(νl,i+νn',i) (6b) 190
Moreover, the sum of the fractions of dietary preference of species i (pi,j, where species j is 191
a prey of species i) must equal 1. Thus, one of the dietary preference fractions, say pi,J, is fully 192
dependent and can be calculated from the remaining fractions: 193
∑−=in
jjiJi pp ,, 1 (7) 194
where ni is the number of preys of species i; we assumed, for each species i, the diet 195
preferences pi,j with the highest values as the dependent diet preference pi,J. 196
Finally, for the benthic species i, the fraction of the respiratory ventilation mO,i that 197
involves overlying water depends upon the pore water fractions mO,i [14]: 198
iPiO mm ,, 1−= (8) 199
while the value mO,i was set equal to one for the species that normally have no direct contact 200
with the pore water [14]. 201
202
The screening method 203

204
The screening technique adopted in the present work for the preliminary ranking of the 205
parameters Xi listed in Table 1, which are also named "input factors" in the framework of 206
UA/SA [5], is the improved version of the Morris method [15], proposed in Campolongo et 207
al. [16]. The key idea of this method, exemplified in is to average "Elementary Effects", 208
resembling "derivatives", over the space of values of the input factors, which is approximated 209
by a grid of p levels. In the example shown in Figure 1, the parallelepiped represents the 210
space of k=3 input factors, whose marginal distribution are subdivided into p=4 levels. 211
The Elementary Effects (EEs) are defined as 212
( ) ( )ji
jjjjjji
jjjjj
ikkiii
xxxYxxxxxxYEE
∆−∆+
= +−,...,,...,,,...,
211121 (9) 213
where, for the factor iX of the p-level grid, an incremental ratio of the model output Y is 214
computed with respect to an increment ji∆ of the value j
ix . By incrementing each factor in 215
random sequence, one obtains a "trajectory" j in the factor space (the black arrows in Figure 216
1), whose (k+1) nodes represent sets of parameter values used to run the model and to 217
compute k values of EEs. A number of j=1,2, ..., r trajectories can be built by randomly 218
selecting r starting grid points (a second trajectory is represented in grey in Figure 1), leading 219
for each input factor to a sample of r EEs evaluated at different points within the whole range 220
of variability of the factor. 221
This sample of EEs allows one to estimate various sensitivity measures, such as the 222
average or the standard deviation [15]: 223
∑
∑
=
=
−=
=
rj i
jii
rj
jii
EE
EE
122
1
])[( µσ
µ, (10) 224
However, as suggested in [16], we used as sensitivity index the average of the absolute 225
values of the elementary effects: 226
∑ == rj
jii EE1
* ||µ (11) 227
where r is the sample size for each factor, usually of the order of 10 [16], for an overall 228
computational cost of )1( +kr model runs. Thus, despite the single jiEE in Equation 8 are 229
‘local’ incremental ratios, the sensitivity index *µ is estimated by averaging several jiEE 230
computed at different points sampled in the whole input space. Therefore, this method can be 231
regarded as a "global" screening technique and allows one to perform a reliable screening for 232
highly non linear models, while local derivative-based methods can not. It has a number of 233

advantages –in terms of lower computational costs, numbers of assumptions and reliability of 234
the results - with respect to other screening methods which are widely accepted in the 235
literature, as demonstrated in [5-7, 16]. In particular, Campolongo et al. [16] compared the 236
sensitivity *µ with a quantitative variance-based measures and demonstrated that µ* is a 237
good proxy of the total sensitivity index ST, which is a measure of the overall effect of a 238
factor on the output (inclusive of interactions) and corresponds to the expected variance of the 239
model output Y that is left when all factors are fixed except Xi. Moreover, Saltelli et al. [7] 240
showed that *µ is particularly suitable for screening purposes because it is rather resilient 241
against type II errors, meaning that, if a factor is found as non-influential by *µ , it is unlikely 242
to be found as influential by applying any other sensitivity measure. Finally, in contrast to µ 243
in Equation 10, the sensitivity index *µ has the advantage that it can be adjusted to provide 244
the overall sensitivity of subsets of input factors (see [16] for details). The last property 245
allowed us to collect the constrained input factors in Equation 1 into groups - such as the ones 246
on the right sides of Equations 7 and 8 - and to evaluate their overall index *µ through the 247
stochastic variations of the single factors but still respecting their mathematical relationships. 248
The cost of grouping factors is the loss of information on the relative importance of the 249
factors belonging to the same group [16]. 250
The routines to carry out the Morris screening analysis are coded in the freely available 251
computer program SimLab [20]. 252
253
The Monte Carlo analysis and the variance partition method 254
255
A Monte Carlo sampling-based analysis [7] was employed in order to explore the 256
relationship between the Equation 1 output and the subset of i=1,2,..., m input factors selected 257
on the basis of the results of the screening sensitivity analysis and collected in an m 258
dimensional input factor vector, X=(X1,..., Xm). 259
A crude Monte Carlo sampling scheme [7] was used to generate a number of t=1,2, ..., N 260
realizations of the input factor vector, and the input-output relationship was represented by 261
means of a multiple regression model, accordingly to the method proposed by Ratto et al. 262
[18]: 263
t
m
itiit eXffy ++= ∑
=1,0 )( ),0( 2σNet = (12a) 264
where: 265

tititii XbXf ,,, )( = (12b) 266
In Equation 12, yt, t=1,2., ... N, represents the tth Monte Carlo output corresponding to the 267
input factor sample Xt=(X1,t, ..., Xm,t); et represents a Gaussian stochastic variable with mean 0 268
and variance σ; and bi,t, i=1,2,..m, is the regression coefficient i, whose value is related to the 269
tth sample of the input factor vector i: )( ,tiii Xbb = . They are collected in the parameter vector 270
bt=(b1,t,..., bm,t). The functional terms )( ,tiii Xff = in Equation 12 are a non linear functions 271
with respect the input factors, due to the dependency of bi,t, upon Xi,t: ( ) titiii XXbf ,, ⋅= . The 272
regression model in Equation 12 was proposed in the framework of time series and system 273
analysis [17], and it is also named "State Dependent Regression" (SDR) model, because it is a 274
function of the "State Dependent Parameters" (SDPs) )( ,tiii Xbb = . 275
Accordingly to [18], the portion Vi of the variance of the model output y explained by each 276
single input Xi was calculated as the ratio of the variance of the associated term fi and the 277
variance of the model output: 278
)var(
)var(
y
fV i
i = (13) 279
where var stands for variance. The variance Vi in Equation 13, represent also an estimator of 280
the main effect sensitivity measure Si of the of the input Xi with respect to the model output, 281
and thus they were used to rank the relevance of the parameters of the model in Equation 1 282
[18]. 283
In order to calculate Vi in Equation 13, one needs to estimate the SDPs bi,t which, 284
accordingly to Equation 12, define the functional terms fi. Coherently with [18], we assumed 285
that bi,t are stochastic variables whose values are defined by an integrated random walk model 286
[17], allowing one to rewrite the regression model (12) in the following State-Space form: 287
tttt ey +⋅= bX ),0( 2σNet = (Observation Equation) (14a) 288
1,1,, −− += tititi dbb (State Equations) (14b) 289
tititi dd ,1,, η+= − ),0( 2, ,ti
Nti ηση = (14c) 290
where di,t ,i=1,2,...,m and t=1,2,...,N, are random variables which provide the stochastic 291
stimulus for the changes of the regression parameters bi,t, through the Gaussian stochastic 292
variables ηi,t, which has mean 0 and variance 2
,tiησ . The variables ηi,t are assumed 293
independent of each other and independent of et [17,18]. Referring for details to [18], the 294
sequence of bi,t values in Equation 14 is estimated by using a recursive estimation approach. 295
This approach requires that the input output data matrix is iteratively sorted along the 296

coordinate of each input factor Xi, and that the ty values are processed one-at-a-time and in 297
turn with respect to each sorted input factor, leading to a corresponding series of optimal bi,t 298
estimates. Coherently with [18], the recursive Kalman Filter and an associated Fixed Interval 299
Smoothing algorithm were used to obtain smoothed least square recursive estimates of bi,t, on 300
the basis of maximum likelihood estimates of the hyperparameters σ and ti ,ησ in Equation 14 301
(see [17] for details). The SDP analysis was carried out by using the freely available computer 302
program SS-ANOVA-R [21]. 303
The SDP approach presented here is equivalent to a smoothing spline analysis of variance 304
(ANOVA) model, with the advantage of the recursive algorithms, which avoid matrix 305
inversions that are computationally demanding [22,23]. Moreover, it is worth highlighting 306
that Equation (12) represents a classical Multiple Linear Regression model (MLR model), if 307
the values of the regression coefficients bi are set constant. A MLR model can been exploited 308
to estimate the portion )var(/)var(22, yXbV iiiil =≈ β of the variance of the model output y 309
explained by each single input Xi [7], where var indicates the variance, bi are the constant 310
regression coefficients estimated by using least square methods and βi are their standardized 311
values. Nevertheless, the above linear estimate Vl,i are reliable if the coefficient of 312
determination R2 of the MLR model is close to unity, that means, if the relationship between 313
the Monte Carlo output y and the factors Xi is quasi linear [7]. Differently, the fraction of 314
variance estimates given by Equation 13 allows one to partition the output variance despite 315
non linear relationships, because the latest are fitted by means of the non linear functional 316
terms fi obtained in the SDR estimation, as we show in the present paper, also through a 317
comparison of the output variance explained by SDR and MLR models. 318
Finally, it is worth highlighting that Monte Carlo based UA/SA - regardless the choice of 319
applying a SDR or a MLR model for the parameter ranking and variance partitioning - is 320
remarkably advantaged by the pre-selection of the input factor by means of the Morris 321
screening analysis. The latest, in fact, allows one to optimize the computational efficiency of 322
the analysis, reducing the number of input factors and thus of required Monte Carlo 323
simulation necessary for carrying out statistically significative regression analysis. But, even 324
more important, it completes the overall UA/SA approach because it allows one to identify 325
the input factors which have negligible effects on the model output, on the basis of a reliable 326
measure of total sensitivity, which currently can not be reliably provided by any 327
metamodelling technique, neither by SDR or MLR models [18]. 328
329

Set up of the analysis 330
331
In order to carry out the UA/SA of the model Equation 1 with respect to the input factors 332
in Table 1, we considered four representative POPs: lindane and three polychlorinated 333
biphenyls (PCB) congeners PCB 15, PCB 101 and PCB 194. As one can see in Table 1, the 334
literature values of the octanol-water coefficients of these chemicals span the range from ∼3.8 335
(lindane) up to ∼8.7 (PCB 194) and, thus, they represent POPs ranging from low hydrophobic 336
up to very hydrophobic, accordingly to a classification used in [1]. 337
Equation 1 was adapted to represent the bioaccumulation of the above chemicals through a 338
coastal food web in the temperate zone, that is the one for the Lagoon of Venice presented in 339
[24,25] and summarized in Tables S.1 and S.2 [SETAC Supplemental Data Archive, Item 340
XXX]. In particular, we selected as model output the concentrations of the four POPs in two 341
key species of the food web, which are the filter-feeder clam Tapes philippinarum and the top 342
predator fish Zosterissesor ophiocephalus. 343
The choice of this food web, lead to include in the model 227 input factors, and the UA/SA 344
required to specify the pdfs of each of them. Accordingly to the objectives of the present 345
work, uniform probability distributions among minimum-maximum ranges were assumed for 346
all the factors, as in [12,13]. The reference values and the minimum-maximum ranges were 347
defined on the basis of a literature review – see Table 1 and Tables S.1 and S.2 [SETAC 348
Supplemental Data Archive, Item XXX]. When the literature ranges were not available, a 349
minimum-maximum range of ±30% of the reference value was arbitrarily assumed. An 350
exception was done for the input factors εl,i and σi - which indicate fractions - when they have 351
a value, say l, in the range 0.77<l<1. In these cases, the ranges were set equal to l ± (1-l), that 352
means less than 30%, in order to avoid the random selection of fraction values greater than 1 353
from the pdfs in the UA/SA. 354
The environmental forcing functions were not included in the UA/SA and their values 355
were kept constant and equal to the values shown in the last column of Table 2, defined on the 356
basis of data collected in the Lagoon of Venice [26]. Moreover, the set of input factors does 357
not include neither the diet preference fractions pi,j equal to 1 - because that means that the 358
species j is the unique prey of species i -, nor the model parameters νw,i and pi,J, because their 359
values are determined by the constrained input factors at the right hand sides of Equations 6 360
and 7. These latest constrained factors were collected into two groups for each of the species, 361
leading to 41 groups collecting altogether 119 factors, among the total number of 227, and the 362
remaining (227-119)=108 factors represented single factor groups. Thus, a total number of 363

k=108+41=149 groups were the object of the screening sensitivity analysis. The number of 364
levels chosen for each factor was p = 4, meaning that the grid over which the elementary 365
effects are computed was constructed by dividing each marginal distribution into four levels. 366
The number of trajectories was r=10, leading to N= r (k+1) = 1500 required model 367
executions. 368
Finally, the input factors with the highest values of the sensitivity indexes µ* with respect 369
any of the model outputs obtained in the screening analysis, were selected to be included in 370
the Monte Carlo based UA/SA. This choice leads to define the same set of factors to be 371
investigated in the quantitative UA/SA, regardless the model output being considered, 372
allowing one the quantitative inter-comparison of the relevance of a model parameter with 373
respect different species and chemicals. In the absence of a theoretical threshold for the µ* 374
values of non-relevant factors [16], a subjective practical threshold was defined with respect 375
to each output, in correspondence of a plateau in the µ* values ordered in a descending 376
sequences, as in the practical case studies presented in [16]. 377
378
RESULTS 379
380
Screening Sensitivity analysis 381
382
The results of the application of the Morris method with respect each model output, are 383
synthesized in the first eight rows of Table 3, which lists the input factors with the sensitivity 384
indices *iµ above the selection thresholds, whose values are indicated in Tables S.3 and S.4 385
[SETAC Supplemental Data Archive, Item XXX]. As one can see in the Table 3, the input 386
factor ranking is a function of the species and chemical being considered. Moreover, in 387
general, sets including just few factors emerged as relevant with respect the chemical 388
concentrations in Zosterisessor ophiocephalus, due to the fact that the sequences of *iµ values 389
quickly dropped to low plateau values (see Tables S.3 and S.4 in [SETAC Supplemental Data 390
Archive, Item XXX]). Differently, when Tapes philippinarum is considered, the *iµ values 391
drop slower and thus larger sets of input factors ranked the highest positions with *iµ values 392
remarkably higher with respect the assumed plateau thresholds. 393
The lower part of Table 3 orders the parameters which appeared at least once in the upper 394
part. Thus, these parameters represent the set of factors which were found to be relevant for, 395
at least, one of the outputs and this set was thus investigated in the subsequent UA/SA. It 396

includes sixteen factors, six of which are physical-chemical parameters, while the remaining 397
ten are species-specific parameters. 398
399
Monte Carlo analysis and variance decomposition 400
401
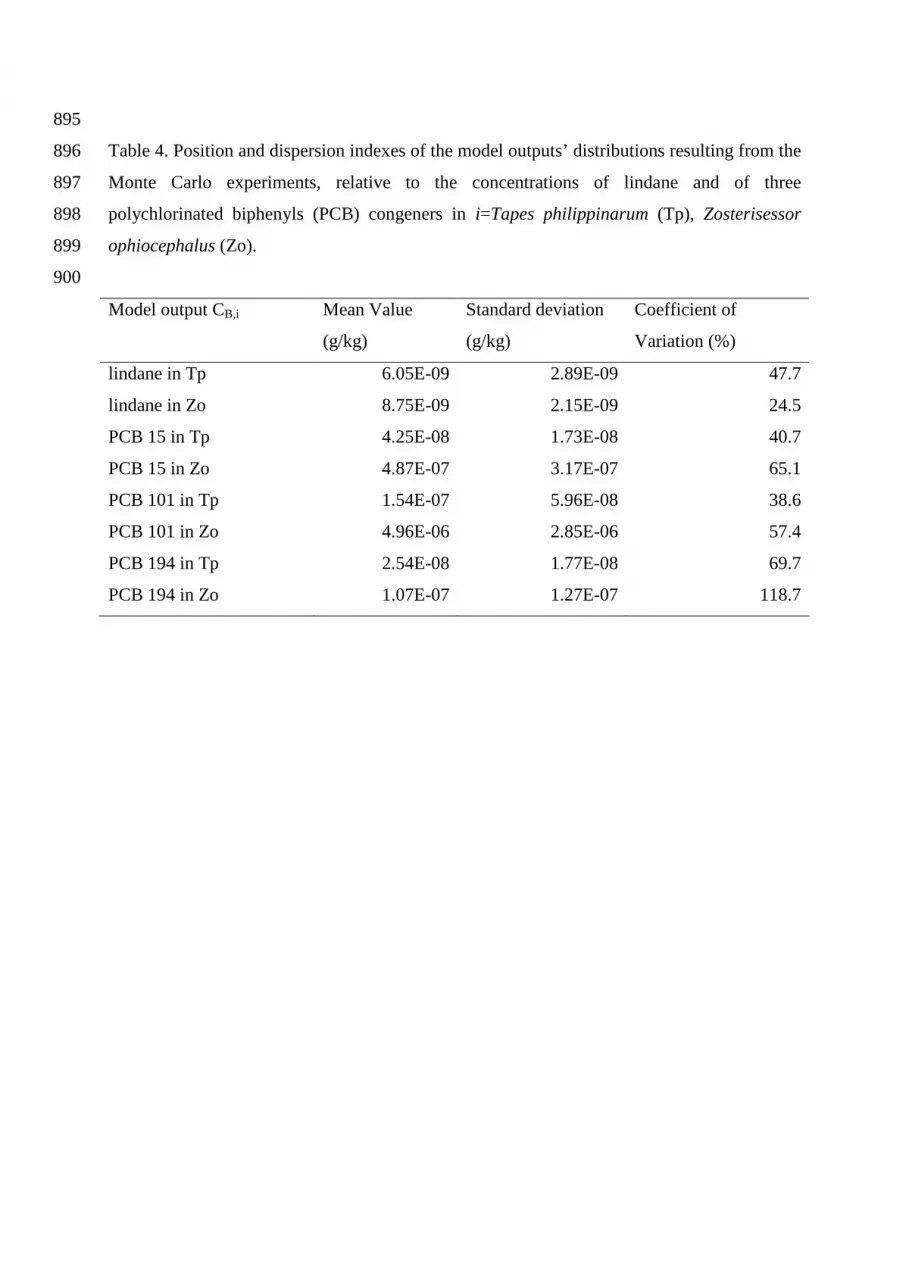
The results of the UA of the model output with respect to the input factors listed in the last 402
rows of Table 3, are summarized in Table 4, which presents the mean values, the standard 403
deviation and the coefficient of variation (CV) of each model output. Table 4 shows that the 404
coefficient of variations -and thus the model uncertainty - were markedly different with 405
respect to the different species and chemicals, ranging from 25% up to 119% for, respectively, 406
lindane and PCB 194 in Zosterisessor ophiocephalus. The mean concentrations and the 407
standard deviations are, in general, remarkably higher for the fish, with the only exception of 408
lindane, whose mean concentrations and standard deviations are of the same order of 409
magnitude with respect to the two species. The CVs are maxima when the very hydrophobic 410
PCB 194 is considered, and their values are comparable when the hydrophobic PCB 15 and 411
PCB 101 are considered. In fact, the CVs of both chemicals are equal to ∼40% and ∼60% for, 412
respectively, the clam and the fish. The CV of the low hydrophobic lindane is relatively low 413
with respect to the fish, but it assumes an intermediate value of ∼50% when the clam is 414
considered. 415
The results of the UA/SA based on the State Dependent Regression (SDR) model in 416
Equation 11 are presented in Table 5 and synthesized graphically in Figure 2. The Table 417
shows the fractions of variance Vi - see Equation 13 - explained by all the input factors while 418
the figure represents the fractions explained by just the most relevant ones. Table 5 reports 419
also the factor rankings based on the Vi values, and in the last row, we compare the sums 420
sum(Vi) - which furnish overall output variance explained by the SDR model - with the 421
fraction of output of variance explainable by fitting MLR models to the Monte Carlo input-422
output samples, sum(Vl,i). The latest sums were calculated on the basis of the Vl,i values 423
reported in Table S.5 [SETAC Supplemental Data Archive, Item XXX]. As one can see, the 424
values of sum(Vi) are higher than 90%, and resulted in all the cases higher than sum(Vl,i), in 425
particular for PCB 101 in both the species (sum(Vl,i) ∼ 60%), for lindane in the clam and PCB 426
194 in the fish, highlighting a significant degree of non-linearity in the model, which can not 427
be accounted for by the MLR model. 428
Table 5 and Figure 2 show differences in the factor ranking with respect to the species and 429
chemicals. Remarkably, the octanol water partition coefficient at 25°C (KOW,25) is the most 430

relevant parameter, ranked first, for most of the outputs, but the Vi values of this factor are 431
highly variable, ranging from 50% (PCB 101 concentration in the clam) up to 96% (PCB 194 432
in the fish). The octanol-carbon proportionality constant (αOC) and the lipid fraction (νl) 433
explain the major part of the uncertainty in the estimated bioaccumulation of lindane, 434
respectively, in the clam (Vi=71%) and in the fish (Vi=50%), while in these two cases Log 435
KOW,25 ranks just 6th (Vi=0.6%) and second (Vi=38%). The nonlipid organic matter-octanol 436
proportionality constant (β) tends to be quite relevant for the estimated chemical 437
concentrations in the clam and, in particular, this factor ranks second and explains ∼10% of 438
the variance of the less hydrophobic lindane and PCB 15. The fraction of the respiratory 439
ventilation which involves pore water (mp) is also important for the estimated 440
bioaccumulation of lindane in the clam, ranking third and explaining 7% of the variance. For 441
both the species, the lipid content of the organism are more important for the less hydrophobic 442
chemicals, while the parameters related to the bioavailability of the chemicals in the water 443
column (αPOC and αDOC in the partition Equation 2) tend to be more relevant with respect to 444
the two more hydrophobic ones- ranking second or third - and, in particular, αDOC explains 445
Vi=25% the variance of PCB 101 in the clam. Finally, it is worth noting that the internal 446
enthalpy ∆HOW and the diet fractions pi,j resulted among the less important factors in 447
determining the uncertainty in the outputs, since they ranked the last positions, with a 448
maximum contribute of 1.4% given by the diet fraction of the fish's prey Micro-Meio benthos 449
(pZo,MiMeb) to the uncertainty in the concentration of PCB 101 in Zosterisessor ophiocephalus. 450
451
DISCUSSION 452
453
The UA/SA approach we have adopted in the present paper allowed to us to partition the 454
variance of the outputs of the Arnot and Gobas model with respect to the most important 455
model parameters. The results showed that the model uncertainty and the parameter ranking 456
are strongly dependent upon the model output which is considered. Thus, for example, the 457
uncertainty in the estimated bioaccumulation was found to be intermediate for lindane in the 458
clam and very high for PCB 194 in the fish (Table 4), due mainly, respectively, to the 459
uncertainty in the organic carbon/octanol constant αOC and in the octanol/water partition 460
coefficient Log KOW,25 (Table 5 and Figure 2). Such differences are related to the physical-461
chemical properties of the chemicals and to the trophic level of the target species, which 462
imply that different uptake and loss processes - and thus model parameters - are involved as 463
main drivers of the estimated POP bioaccumulation. However, the results were, evidently, 464

influenced also by our methodological approach to the UA/SA. In particular, by our 465
assumptions upon the partial distribution functions (pdfs) of the factors, by our choice of 466
preferring global non linear methods to local linear ones, and by the criteria adopted to select 467
the set of input factors investigated in the UA/SA, on the basis of the screening results. In the 468
following, these methodological key points are firstly deepened before discussing the model 469
uncertainty and the variance partitioning among the factors in relation to the equations in the 470
Arnot and Gobas model (1). 471
472
The pdfs of the input factors 473
474
The definition of the probability density functions (pdfs) of the input factors is a critical 475
step in carrying out the UA/SA analysis of a model. In fact, factors characterized by the same 476
sensitivity indexes contribute to the output uncertainty in a measure somehow depending 477
upon their assumed variability. 478
In the application presented in this paper, we made some arbitrary assumptions with 479
respect the pdfs of the factors in Table 1, because their shapes and ranges are not univocally 480
defined in the literature (see for example the different pdfs assumed in [2,4,12,13]). Since the 481
theoretical applicability of both the Morris method and the SDR model does not rely on the 482
choice of any particular pdf, we assumed independent uniform distributions for of all the input 483
factors, like for example in [12,13], because two main reasons made this choice preferable to 484
others in the present application. First, using independent inputs makes the variance 485
decomposition unique, while assuming dependencies among input factors implies infinite 486
possible variance decomposition schemes, thus making the interpretation of the sensitivity 487
analysis results much more problematic (see for example Saltelli et al. [6] for a discussion of 488
the use of independent inputs in sensitivity also in the presence of dependency among input 489
factors). Second, using uniform distributions, as in [12,13], makes the UA/SA less prone to 490
possible mis-specified pdf’s, because it avoids to estimate the sensitivity index µ* and Vi on 491
the basis of random factor samples that are wrongly ‘too dense’ in some part of the 492
uncertainty space and ‘too sparse’ in some other regions of the space. 493
On the other hand, the ranges of the uniform distributions were set equal to the minimum-494
maximum ranges found in the literature and, in most instances, are consistent with those 495
applied in other UA/SA of bioaccumulation models (see for example [13]). As pointed out in 496
the Methods, when ranges were not available, we decided to set a range of ±30% the nominal 497
values for most of the species-specific parameters, which is lower than the 50% used in 498

Saloranta et al [13] and higher than the 10% applied in Nfon and Cousins [12]. This choice 499
aims at setting ranges of comparable size with respect to most of the input factors. In this way, 500
the ranks given by the screening sensitivity analysis are more likely to be determined by the 501
relevance of the factors rather than by differences in their ranges of variability. However, our 502
assumption does not remarkably affect the outcome of the screening step. In fact, in order to 503
check the robustness of the results, we repeated the screening analysis assuming a ±50% 504
range, as in Saloranta et al [13], for the species-specific parameters. The result, presented in 505
Table S.6 and S.7 [SETAC Supplemental Data Archive, Item XXX], showed that, despite the 506
variations in the µ* values, the parameters in Table 3 still ranked the highest positions with 507
respect the different model outputs and the parameters in the lower part of Table 3 would 508
have been again selected as input factors of the quantitative UA/SA on the basis of the same 509
threshold values. 510
511
Robustness of the screening analysis 512
513
The set of parameters in Table 3 were identified on the basis of the screening results and 514
defining arbitrary thresholds for the sensitivity measure µ*. In order to confirm that none 515
relevant model parameter was indeed classified as negligible and excluded from the 516
quantitative UA/SA, we performed a Monte Carlo experiment where all the model parameters 517
in Table 1, besides the sub set in Table 3, were uniformly sampled in their minimum-518
maximum ranges defined in Table 1. As a result of one thousand runs, the mean values and 519
standard deviations of the model outputs- presented in Table S.8 [SETAC Supplemental Data 520
Archive, Item XXX]- were found to be almost equal to those shown in Table 4. It was thus 521
proved that the few model parameters identified in Table 3 are by far more relevant than all 522
the remaining model parameters altogether in determining the model output variability. 523
524
Linear vs. non linear global methods 525
526
From the results in Table 5, one can infer that the relationships among some of the factors 527
and the model outputs were remarkably non-linear. In fact, the SDR models explained larger 528
fractions of the output variance with respect to the MLR models. An example of a non linear 529
relationship between an input factor and the model output is shown in Figure 3, where CB,i in 530
Equation 1 is plotted as a function of LogKOW values from 3 up to 9, which include the ranges 531
of variability of the octanol/water coefficient at 25°C (LogKOW,25) of the four chemicals 532

included in the UA/SA. One can see that CB in both the target species has a non linear bell-533
shape in the range of variability of PCB 101 or an exponential-like decrease in the range of 534
variability of PCB 194. As a consequence, the application of global and non linear methods in 535
the screening step and in the quantitative UA/SA avoids misleading parameter rankings and 536
variance partitions which could result from the application of local linear methods. 537
Thus, in the screening step, a local sensitivity index, such as the widely applied local 538
standardized sensitivities si, (see for example [9]) could have lead to classify the factor Log 539
KOW,25 of PCB 101 as a negligible one, if one had chosen a nominal values in the 540
neighborhood of the local maximum at Mx = Log KOW ∼ 6.6, evident in Figure 3. For 541
example, by changing by ∆x=0.6 (∼10%) a nominal value of x=6.35 and by evaluating the 542
corresponding fluctuations ∆yi in the estimated concentrations y of PCB 101 in Z. 543
ophiocephalus, one obtains 01.0)/()/( =⋅∆∆= iiiii yxxys and such an almost null value of 544
the sensitivity measure could lead one to classify the factor Log KOW,25 as negligible. This 545
misleading conclusion is avoided by using the Morris methods and the sensitivity index µ*, 546
which accounts for the absolute values of Elementary Effects calculated across the whole 547
range of variability of the parameter, leading to classify Log KOW,25 as the most important 548
parameter for the estimated bioaccumulation of PCB 101 in both the target species (Table 3). 549
Analogous considerations holds for any POPs of which the ranges of Log KOW,25 values 550
include the critical point Mx ∼6.6, such as, for example, PCB 105 and PCB 155 [27]. 551
On the other hand, as far as the Monte Carlo based UA/SA is concerned, Table 5 clearly 552
shows that the non linear SDR models explain larger fractions of the output variance with 553
respect to linear regression models. Thus, the relevant degree of non-linearity in the model 554
makes the MLR model inadequate for partitioning the variance of the model output. Thus, for 555
example, the better performances of the SDR models highlighted in Table 5 with respect to 556
the cases of the lindane concentration in the clam or the PCB 101 concentrations in the fish 557
were mainly due to the non linear effects of, respectively, X3=αOC and X1=KOW,25. In fact, in 558
Figure 4 one can see that, in both the cases, the non linear functions fi of the SDR models fit 559
much better the outputs of the Monte Carlo simulations if compared with the straight lines 560
bi⋅X i estimated by the MLR models. Consequently, the values of the variance fractions Vl,i 561
given for the above two factors by the linear model (Vl,3=58% and Vl,1=28% in Table S.5 562
[SETAC Supplemental Data Archive, Item XXX]) underestimate the fractions of explained 563
variance provided by the SDR model (Vl,3=71% and Vl,1=52% in Table 5). On the other hand, 564
when the coefficient of determination of the MLR models are close to one, the performances 565

of the linear regression compares well with those of the SDR model - as in the cases of the 566
results for the concentrations of PCB 15 in the clam or of lindane in the fish, in Table 5 and 567
Table S.5 [SETAC Supplemental Data Archive, Item XXX]. 568
569
The factor rankings in relation to the bioaccumulation processes 570
571
The results presented in Figure 2 and in Tables 4 and 5 show that the uncertainty in the 572
model output and the variance partitioning depend upon the trophic level of the species and 573
the hydrophobicity of the POP under investigation, which determine the relevance of the 574
different chemical uptake and loss routes. Thus, in the following, the factor rankings are 575
interpreted in relation to the equations and parameterizations of the bioaccumulation 576
processes in Equation (1), gathering insights into the performances of Arnot and Gobas 577
model. To this aim, we refer also to the graphs of Figures 5, 6 and 7, which show, 578
respectively, the uptake and loss rates, the functional expressions defining the uptake rates, 579
and the water concentrations of the chemical as functions of KOW. For the sake of clarity, in 580
the following we discuss separately low (Log KOW<5) and very hydrophobic (Log KOW>5) 581
chemicals. 582
Low hydrophobic chemicals. As one can see in Figure 5, the respiratory uptake is the main 583
inflow route of chemicals in the range Log KOW<5, which include lindane. This uptake is 584
counterbalanced by the high respiratory elimination rate, which keeps the lindane 585
concentration in the target species to the relatively low values shown in Table 4. Since the 586
respiratory route is highly relevant for low hydrophobic chemicals, the input factors related to 587
the POP bioavailability in the water (αOC), to its flux through the respiratory areas (mp), and 588
to its sorption by the organic matter (β and νl,i) rank high positions in Table 5 when the 589
lindane concentrations in the organisms are considered. 590
As far as differences among species are concerned, Figure 6 shows that the respiratory 591
uptake is higher for the clam due to both the higher values of the clearance rate constant k1 592
and to the higher chemical concentrations CWR in the water involved in the respiration, which 593
is in turn due to the high chemical concentrations in the pore water fraction (CWD,S in Figure 594
7). As a consequence, in Table 5, the factors αOC and mp, which respectively drive the 595
sediment/pore water partitioning of the POPs (see Equation 3) and quantify the pore water 596
fraction filtered by the clam (see Equation 1), rank the highest positions when the lindane 597
concentration in Tapes philippinarum is considered, explaining altogether ∼80% of the model 598
output variance. On the other hand, the relatively low rank of the factors characterizing the 599

bioavailability in the overlay water involved in respiration (αDOC and αPOC in Equation 2), 600
with respect to αOC, is mainly due to our choice of setting CS>>CWT,O in Table 2, which leads 601
the bioavailable concentrations in the overlay water (CWD,O in Figure 7) to be a negligible 602
source of chemical with respect to CWD,S when Log KOW< 5. 603
Accordingly to the Arnot and Gobas model, the chemical in the water entering the 604
respiratory area of the species is then sorbed by the organism in a measure which is 605
proportional to the coefficient iwOWinOWilBW KKkkK ,,,21 / νβνν +⋅⋅+⋅== [14]. Thus, the 606
lipid fraction νl,i and the nonlipid organic matter-octanol proportionality constant β become 607
critical parameters for the respiratory absorption of lindane and thus they rank, respectively, 608
first and second positions for, respectively, the relatively "fat" fish and the "thin" clam. This 609
outcome is in agreement with the indications given in Arnot and Gobas [14] and deBruyn and 610
Gobas [29], which highlights that further accurate estimates of β are particularly relevant for 611
reliable estimates of POPs bioaccumulation in species which lipid fraction is less than ∼5% 612
the dry-weight organic content (as Tapes philippinarum), since in these cases the sorptive 613
capacity can be dominated by the contribution from proteins rather than from lipids [29]. 614
Finally, the relatively high relevance of Log KOW,25 for lindane bioaccumulation in the fish is 615
partially due to its influence on KBW and on k1, the latest being proportional to the gill 616
chemical uptake 1)/15585.1( −+= OWW KE , whose slope reaches its maximum with respect to 617
Log KOW in the range Log KOW< 5. Nevertheless, the relevance of Log KOW,25 is also due to 618
its influence on the diet uptake rate, which, for the fish, is not negligible neither in the range 619
of low hydrophobic chemicals, as one can see in Figure 5. 620
Very hydrophobic chemicals. In the range of Log KOW>5, which include the three PCB 621
congeners we investigated, the relevance of the dietary uptake become comparable or much 622
higher with respect to the respiration uptake, in the cases of, respectively, the clam and the 623
fish, as one can see in Figure 5. Moreover, the diet uptake is quantitatively more relevant for 624
Zosterisessor ophiocephalus, due to the higher chemical concentrations in its diet, as shown 625
in Figure 6 by the relevant differences of the CD values for the two species, coherently with 626
the conceptual model of biomagnification of hydrophobic chemicals at the highest trophic 627
positions [1]. 628
Thus, on one hand, some of the input factors related to the respiratory route still explain 629
large fractions of the outputs for the clam, such β and αPOC and αDOC, which, in the range Log 630
KOW >5, have the main influence on CWR due to the relevance of CWD,O, as one can see in 631
Figure 7. On the other hand, the input factors more directly involved in the equations of the 632

dietary uptake (such as Log KOW,25, εl,i or σTp) increase their ranking positions when 633
considering the hydrophobic chemicals, with respect to both the species. 634
In particular, in Table 5, Log KOW,25 resulted by far the most relevant parameter with 635
respect to the three PCBs, explaining more than 50% of the variance of their concentrations in 636
the target species. Thus, it is worth noting that KOW directly influence the dietary route 637
accordingly to a lipid-water two-phase resistance model 17 )0.2100.3( −− +⋅⋅= OWD KE , which 638
enters in the formulations of the fecal elimination rate constant kE (see Figure 5) and of the 639
clearance rate constant kD (see Figure 6). This function has the highest slope with respect to 640
Log KOW when 5 > Log KOW > 8, leading to the high sensitivity of the dietary route and thus 641
of the PCB 15 and PCB 101 outputs with respect to Log KOW,25. Differently, the relevance of 642
this factor upon the model output was only secondarily determined by the influence of KOW 643
on the partition coefficient of the chemicals between the intestinal content and the organism 644
(KGB, [14]), since the function KGB has an almost constant plateau value with respect to Log 645
KOW, when considering hydrophobic chemicals. Finally, in the range Log KOW > 8, which 646
includes PCB 194, the estimated POP concentrations in the water rapidly drop to negligible 647
values, as one can see in Figure 7, coherently with the conceptual model of binding the more 648
hydrophobic chemicals to sediment and suspended organic matter [14,28]. However, 649
fluctuations of the Log KOW,25 value may lead, also in this case, to relevant changes in the 650
bioavailable water concentrations and in the estimated bioaccumulation, as highlighted by the 651
large fraction of variance explained by this factor in Table 5 and by the high uncertainty of the 652
estimated PCB 194 concentrations in the target species in Table 4. These results stress the 653
well known relevance of accurate KOW estimates for reliable applications of food web 654
bioaccumulation models (see for example the discussion in [39]), and, in particular, they 655
highlight that the model accuracy may become critical when very hydrophobic chemicals 656
characterized by uncertain KOW,25 values are considered. 657
658
CONCLUSIONS 659
660
A methodological approach for carrying out global UA/SA of food web bioaccumulation 661
models with respect the model parameters was proposed. The approach was then applied to 662
the analysis of a state of the art model and to a coastal marine food web, leading to the 663
quantitative estimation of the most relevant sources of model uncertainties, with respect to 664
eight representative outputs. 665

The use of this approach is suggested because it allows one to deal with large numbers of 666
model parameters, characterized by huge ranges of uncertainty, which may lead the model to 667
non-linear responses. In fact, it consists in a preliminary screening analysis aimed to select the 668
most relevant model parameters on the basis of a sensitivity index calculated by exploring 669
their whole range of variability. The selected parameters are then included as input factors in 670
a subsequent Monte Carlo based quantitative UA/SA. The screening and quantitative analysis 671
are carried out by means of two global methods - the refined Morris method and the State 672
Dependent Regression of the Monte Carlo input-output sample - not yet applied in the 673
framework of food web bioaccumulation modelling. These methods were proved to be 674
preferable to classical local and linear methods in the applications described in the present 675
paper. 676
Such applications provided valuable insights into the performances of the Arnot and Gobas 677
model, which is widely applied in the framework of risk assessments and regulatory efforts. 678
In particular, the analysis lead to the identification of the negligible model parameters, whose 679
values could be fixed to any value within their ranges of variability without influencing 680
significantly the model outputs. The modeller could even consider either to eliminate or to 681
simplify the parts of the model where these parameters are involved. This is the case, for 682
example, of the Van't Hoff Equation 4, due to the low relevance of the enthalpy of phase 683
transfer ∆HOW. On the other hand, the UA/SA indicated those factors which need to be further 684
investigated and correctly specified for reliable model applications, such as for example the 685
nonlipid organic matter-octanol proportionality constant β, or the lipid content of organisms 686
νl, or the fraction of pore water involved in respiratory uptake mp, which for low hydrophobic 687
chemicals can be more relevant than Log KOW,25 in determining the model output uncertainty. 688
Coherently with the objectives of the paper, the UA/SA focused on the model parameters 689
rather then on the model environmental forcing functions, such as the POP concentration in 690
the sediment or the water temperature. These forcings, in practical model applications, can 691
contribute considerably to the variability of the model predictions (see for example the 692
analysis in Nfon and Cousins [12] for the Baltic sea), and thus our estimates of model 693
uncertainty can not be compared the actual variability of the estimates of POPs 694
concentrations in the species in the Lagoon of Venice, for example those presented in 695
Micheletti et al [25]. But the extension of the global UA/SA approach to account also for the 696
environmental forcings is straightforward, once their pdfs are specified on the basis of site 697
specific data. Thus, in the framework of ongoing research, the approach is being applied to 698
compare the relevance of water temperature fluctuations in determining the output variability, 699

with respect to the relevance of the actual uncertainty in the model parameters. In fact, we 700
retain such preliminary analysis mandatory for reliable modelling assessments of the 701
consequences of future climatic scenarios upon the POP bioaccumulation in coastal food 702
webs. 703
704
Acknowledgement – The authors thank CMCC for partially founding the present work and 705
Ministero delle Infrastrutture-Magistrato alle Acque di Venezia for making data and technical 706
reports available. 707
708
REFERENCES 709
710
1. Kelly BC, Ikonomou MG, Blair JD, Morin AE, Gobas FAPC. 2007. Food web–711
specific biomagnification of persistent organic pollutants. Science 317:236 – 239. 712
2. Arnot JA, Gobas FAPC. 2006. A review of bioconcentration factor (BCF) and 713
bioaccumulation factor (BAF) assessments for organic chemicals in aquatic 714
organisms. Environ Rev 14:257-297. 715
3. Mackay D, Fraser A. 2000. Bioaccumulation of persistent organic chemicals: 716
Mechanisms and models. Environ Pollut 110:375–391 717
4. MacLeod M, Fraser AJ, Mackay D. 2002. Evaluating and expressing the propagation 718
of uncertainty in chemical fate and bioaccumulation models. Environ Toxicol Chem 719
21:700-709. 720
5. Saltelli A, Chan K, Scott M, eds. 2000. Handbook of Sensitivity Analysis. John Wiley 721
& Sons, New York, USA. 722
6. Saltelli A, Tarantola S, Campolongo F, Ratto M. 2004. Sensitivity analysis in practice. 723
A guide to assessing scientific models. John Wiley & Sons, New York, USA. 724
7. Saltelli A, Ratto M, Andres T, Campolongo F, Cariboni J, Gatelli D, Saisana M, 725
Tarantola, S. 2008. Global Sensitivity Analysis, The Primer, Wiley & Sons, 726
Chichester, UK. 727
8.- Morrison HA, Gobas FAPC, Lazar R, Whittle DM, Haffner GD. 1997. Development 728
and verification of a benthic/pelagic food web bioaccumulation model for PCB 729
congeners in Western Lake Erie. Environ Sci Technol 31:3267-3273. 730
9. Burkhard LP. Comparison of two models for predicting bioaccumulation of 731
hydrophobic organic chemicals in a Great Lakes food web. 1998. Environ Toxicol 732
Chem, 17:383-393. 733

10. Gewurtz SB, Lposa R, Gandhi N, Guttorm C, Evenset A, Gregor D, Diamond ML. 734
2006. A comparison of contaminant dynamics in arctic and temperate fish: A 735
modelling approach. Chemosphere 63:1328-1341. 736
11. Gobas FAPC. 1993. A model for predicting the bioaccumulation of hydrophobic 737
organic chemicals in aquatic food webs: Application to Lake Ontario. Ecol Model 738
69:1–17. 739
12. Nfon E, Cousins IT. 2007. Modelling bioaccumulation in a Baltic food web. Environ 740
Pollut 148: 73-82. 741
13. Saloranta TM, Andersen T, Naes K. 2006. Flow of dioxins and furans in coastal food 742
webs: inverse modeling, sensitivity analysis, and applications of linear system theory. 743
Environ Toxicol Chem 25:253-264. 744
14. Arnot JA, Gobas FAPC. 2004. A food web bioaccumulation model for organic 745
chemicals in aquatic ecosystems. Environ Toxicol Chem 23:2343-2355. 746
15. Morris MD. 1991. Factorial sampling plans for preliminary computational 747
experiments. Technometrics 33:161-174. 748
16. Campolongo F, Cariboni J, Saltelli A. 2007. An effective screening design for 749
sensitivity analysis of large models. Environmental Modelling & Software 22:1509-750
1518. 751
17. Young, PC. 2000. Stochastic, dynamical modelling and signal processing: Time 752
variable and state dependent parameter estimation. In: Fitzgerald WJ, Smith RL, 753
Walden AT, Young PC, eds, Nonlinear and Nonstationary Signal Processing, 754
Cambridge University Press, Cambridge, USA, pp 74-114. 755
18. Ratto M, Pagano A, Young P. 2007. State dependent parameter meta-modelling and 756
sensitivity analysis. Computer Physics Communications 177:863-876. 757
19. Gobas FAPC, Maclean LG. 2003. Sediment–water distribution of organic 758
contaminants in aquatic ecosystems: The role of organic carbon mineralization. 759
Environ Sci Technol 37:735–741. 760
20. SimLab. Joint Research Centre (European Commission), Ispra, Italy. 761
http://simlab.jrc.ec.europa.eu 762
21. SS-ANOVA-R. 2007. Joint Research Centre (European Commission), Ispra, Italy. 763
http://eemc.jrc.ec.europa.eu/softwareSS-ANOVA-R-Dowload.htm. 764
22. Wahba G. 1990. Spline Models for Observational Data. In Series in Applied 765
Mathematics, Vol 59. Society for Industrial and Applied Mathematics. Philadelphia, 766
USA. 767

23. Gu C. 2002. Smoothing Spline ANOVA Models. Springer, New York, USA. 768
24. Libralato S, Pastres R, Pranovi F, Raicevich S, Granzotto A, Giovanardi , Torricelli P. 769
2002. Comparison between the energy flow networks of two habitats in the Venice 770
Lagoon. Mar Ecol 23:228–236 771
25. Micheletti C, Lovato T, Critto A, Pastres R, Marcomini A. Spatially distributed 772
Ecological risk for fish of a coastal food web exposed to dioxins. Environ Toxicol 773
Chem 27:1217-1225. 774
26. Penna G, Barbanti A, Bernstein AG, Boato S, Casarin R, Ferrari G, Montobbio L, 775
Parati P, Piva MG, Vazzoler M. 2007. Institutional monitoring of the Venice Lagoon, 776
its watershed and coastal waters: a continuous updating of their ecological quality. In 777
Fletcher CA, Spencer DC, eds, Flooding and Environmental Challenges for Venice 778
and its Lagoon. State of Knowledge. Cambridge University, Cambridge, United 779
Kingdom, pp 517-528. 780
27. Li N, Wania F, Lei YD, Daly GL. 2003. A comprehensive and critical compilation, 781
evaluation, and selection of physical-chemical property data for selected 782
Polychlorinated Biphenyls. J Phys Chem Ref Data 32:1545-1590. 783
28. Erickson RJ, MacKim JM. 1990. A model for exchange of organic chemicals at fish 784
gills: flow and diffusion limitations. Aquat Toxicol 18: 175-198. 785
29. deBruyn AMH, Gobas FAPC. 2007. The sorptive capacity of animal protein. Environ 786
Toxicol Chem 26:1803-1808. 787
30 Paschke A, Schüürmann G. 1998. Ocatanol/water-partitioning of four HCH isomers at 788
5, 25 and 45 °C. Fresenius Envir Bull 7:258-263. 789
31 Paschke A, Popp P, Schüürmann G. 1998. Water solubility and octanol/water-790
partitioning of hydrophobic chlorinated organic substances determined by using 791
SPME/GC. Fresenius J Anal Chem 360: 52-57. 792
32 Paschke A, Schüürmann G. 2000. Concentration dependence of the octanol/water 793
partition coefficients of the Hexachlorocyclohexane isomers at 25°C. Chem Eng 794
Technol 23: 666-670. 795
33 Harner T, Bidleman TF, Jantunen LMM, MacKay D. 2001. Soil-air exchange model 796
of persistent pesticides in the United States cotton belt. Environ Toxicol Chem 797
20:1612-1621. 798
34 MacKay D, Shiu WY, Ma KC. 1997. Illustrated handbook of physical-chemical 799
properties and environmental fate for organic chemicals. Volume V Pesticide 800
Chemicals. Lewis Publishers, New York, USA. 801

35 Ruelle P, Kesselring UW. 1998. The hydrophobic effect. 3. A key ingredient in 802
predicting n-octanol-water partition coefficient. Journal of Pharmaceutical Sciences 803
87:1015-1024 804
36 Paassivirta J, Sinkkonen S., Mikkelson P, Rantio T, Wania F. 1999. Estimation of 805
vapour pressures, solubilities and Henry's law constants of selected persistent organic 806
pollutants as function of temperature. Chemosphere 39:811-832 807
37 Seth R, Mackay D, Muncke J. 1999. Estimating the organic carbon partition 808
coefficient and its variability for hydrophobic chemicals. Environ Sci Technol 809
33:2390–2394. 810
38. Burkhard LP. 2000. Estimating dissolved organic carbon partition coefficients for 811
nonionic organic chemicals. Environ Sci Technol 34:4663–4668. 812
39. Rebecca R. 2002. The KOW Controversy. Environ Sci Technol, 36:411A-413A.813

Figure 1 Graphical representation of the Morris sampling strategy in a three dimensional input 814
factor space which define the ranges of variability of k=3 input factors Xi (i=1,...,k) 815
subdivided into a grid of four levels (p=4). The trajectory j (j=1, 2, ...,r), represented by the 816
black arrows, connect a sequence of grid points which differ in one of coordinates by the 817
quantity ji∆ . The grey arrows represent a second trajectory drawn by changing the starting 818
grid point and the sequence of the factor variations. 819
820
Figure 2 Fractions of the variance of the concentrations of lindane and of three 821
polychlorinated biphenyls (PCB) congeners in i=Tapes philippinarum, Zosterisessor 822
ophiocephalus explained by the input factors included in the quantitative UA/SA. 823
824
Figure 3 Estimated organism concentration (CB) of persistent organic pollutants in i=Tapes 825
philippinarum, Zosterisessor ophiocephalus as a function of Log KOW values in the range 826
from 3 up to 9, which includes the ranges of variability of lindane and of three 827
polychlorinated biphenyls (PCB) congeners accounted for in the UA/SA. 828
829
Figure 4 Scatter-plots of the lindane concentrations in Tapes philippinarum (CTp) and of the 830
polychlorinated biphenyls (PCB) congener PCB 101 in Zosterisessor ophiocephalus (CZp) 831
versus the values of, respectively, the organic carbon – octanol proportionality constant (αOC) 832
and the octanol-water partition coefficient at 25°C (Log KOW,25), obtained by the Monte Carlo 833
runs. The values of the corresponding state dependent regression functions fi are represented 834
by means of continuous black lines and the linear regression functions bi⋅X i by means of 835
dashed lines 836
837
Figure 5 Estimated values of the respiratory uptake (k1⋅CWR), dietary uptake (kD⋅CD), 838
respiratory elimination rate (k2), excretion elimination rate (kE) and loss rate due to growth 839
dilution (kG) for Tapes philippinarum (graphs A and C) and Zosterisessor ophiocephalus 840
(graphs B and D) versus Log KOW. 841
842
Figure 6 Estimated values of the respiratory clearance rate (k1), chemical concentration ⋅in the 843
respired water (CWR), dietary clearance rate (kD) and chemical concentration ⋅in the prey items 844
(⋅CD) of Tapes philippinarum and Zosterisessor ophiocephalus versus Log KOW values in the 845
range from 3 up to 9. 846
847

Figure 7 Estimated values of the freely dissolved concentrations in pore water (CWD,S) and in 848
the water column (CWD,O) versus Log KOW values in the range from 3 up to 9 estimated by 849
means of the partitioning Equations 2 and 3. 850

851
(x1j,x2
j+∆2j, x3
j)x2
x3
(x1j +∆1
j,x2j+∆2
j, x3j)
(x1j +∆1
j,x2j+∆2
j, x3j+∆3
j)
x1
(x1j,x2
j, x3j) (x1
j,x2j+∆2
j, x3j)
x2
x3
(x1j +∆1
j,x2j+∆2
j, x3j)
(x1j +∆1
j,x2j+∆2
j, x3j+∆3
j)
x1
(x1j,x2
j, x3j)
852
853
854
Figure 1 855

856
857
Figure 2 858

859
860
Figure 3 861

862
Figure 4 863

864
865
866
867
Figure 5 868

869
870
871
872
Figure 6 873

874
875
876
Figure 7 877
878

879
Table 1 Summary of the input factors accounted for in the UA/SA analysis, units, range and 880
reference values and their sources. 881
882
883
Definition Input factor Units Range Reference
value (r.v.)
Octanol-water partition
coefficient at 25°C
X1=
Log KOW,25 unitless
lindane: 3.7-3.9 a
PCB 15: 4.8-5.5 b
PCB 101: 5.9-7.6 b
PCB 194: 7.6-8.7 b
3.8 1
5.17 1
6.8 1
8.15 1
Internal enthalpy of phase
transfer octanol-water
X2=∆HOW J K-1 mol-
1
lindane: 9.88-10.92 2
PCB 15: 19.95-20.5 2
PCB 101: 22.8-25.2 2
PCB 194: 25.65-
28.35 2
10.4 c
21 b
24 b
27 b
Organic carbon – octanol
proportionality constant
X3=αOC unitless 0.14-0.89 3 0.35d
Dissolved organic carbon –
octanol proportionality
constant
X4=αDOC unitless 0.004-1.6 4 0.08e
Particulate organic carbon –
octanol proportionality
constant
X5=αPOC unitless 0.14-0.89 5 0.35f
Nonlipid organic matter-
octanol proportionality
constant
X6=β unitless 0.0105-0.0595 6 0.035f
Nonlipid organic carbon-
octanol proportionality
constant
X7=βa unitless 0.14-0.89 7 0.35f
Weight of species i X8,i=wi kg Table A in Appendix
Lipid fraction of species i X9,i=νl,i kg /kg Table A in Appendix
Nonlipid organic matter
fraction of heterotrophic
X10,i=νn,i kg /kg 0.14-0.26 8 0.20d

species i
Nonlipid organic carbon
fraction of autotrophic
species i
X11,i=νn’,i kg /kg 0.14-0.25 8 0.195d
Water fraction of species i X12,i=νw,i kg /kg Eq. (8)
Dietary absorption
efficiency of lipid for
heterotrophic species i
X13,i=εl,i %
plankton 0.50-0.94 8
invertebrates 0.53-
0.988
fishes 0.84-1.00 9
0.72 d
0.92 d
0.72 d
Dietary absorption
efficiency of nonlipid
organic matter for
heterotrophic species i
X14,i=εn,i %
plankton 0.50-0.94 8
invertebrates 0.53-
0.988
fishes 0.42-0.78 8
0.72 d
0.75 d
0.60 d
Dietary absorption
efficiency of water for
heterotrophic species i
X15,i=εw,i % 0.35-0.65 8 0.50 h
Fraction of dietary
preference of species i with
respect species j
X16,i=pi,j % Table B in Appendix
Weight of sediment/detritus X17,i=ws kg 1.8-3.3 8 2.57 h
Lipid fraction in
sediment/detritus
X18,i=νl,s % 0.0035-0.0065 8 0.005 h
Nonlipid organic fraction in
sediment/detritus
X19,i=νn,s % 0.028-0.052 8 0.04 h
Water fraction in
sediment/detritus
X20,i=νw,s % 0.24-0.45 8 0.345 h
Growth rate of autotrophic
species i
X21,i=kg,i d-1 0.056-0.0.104 8 0.08 d
Fraction of the respiratory
ventilation of the benthic
species i which involve
pore water.
X22,i=mp,i % 0.035-0.065 8 0.05 d
Scavenging efficiency of
particles for filter-feeding
X23,i=σi % 0.80-1.00 9 0.90 10

species i
a Range of lindane KOW values at 25 °C in: Paschke and Schüürmann [30], Paschke et al. [31],
Paschke and Schüürmann [32], Harner et al [33], MacKay et al. [34], Ruelle and Kesselring
[35], Paassivirta et al. [36] b Li et al [27]; c Paschke and Schüürmann [30]; d Seth et al. [37]; e Burkhard [38]; fArnot and
Gobas [14]; g Gewurtz et al. [10]; hMicheletti et al [25] 1 Reference value defined as the central value of the literature range; 2 Range defined as ±5% the r.v. 3 Calculated by considering a variation by a factor 2.5 in either direction for the regression
Koc=0.35 Kow [37]; 4 Calculated by considering a 95% confidence limit of a factor 20 in either direction for the
regression KDOC=0.08 Kow [38]; 5 Calculated in analogy with respect αDOC as suggested by Arnot and Gobas [14] 6 Range defined as ±70% the r.v. value, same order of variability of αOC and αPOC; 7 Range defined equal to the range of αOC due to the analogy among the two parameters [14]; 8 Range defined arbitrarily in the present work as ±30% the r.v.; 9 Range defined as 2 x (1-r.v.); 10 Reference value changed in the present work from 1 in [25] to 0.9 in order to include the
parameter among the input factors of the UA/SA

Table 2. Environmental forcing functions of the food web bioaccumulation model. The 884
reference values compares to the values found in the Lagoon of Venice [26]. 885
886
Environmental forcing Label Unit Reference Value
Total chemical concentration in overlay water CWT,O g/L 1E-11 1
Chemical concentration in sediment Cs g/kg 1E-7 1
Organic carbon fraction in sediment OCs % 0.032
Sediment density δs kg/L 1.67
Concentration of suspended solids in water Css g/L 3.1E-5
Particulate organic carbon concentration in water χPOC kg/L 1.0E-6
Dissolved organic carbon concentration in water χDOC kg/L 1.0E-6
Water temperature T °C 17.5
Dissolved oxygen concentration 2 Cox mgO2/L 8.1
1 The concentrations of the four chemicals were set equal to a single arbitrary value for
comparison purposes.

Table 3. The most important input factors identified by the screening sensitivity analysis with 887
respect the target species i=Tapes philippinarum (Tp), Zosterisessor ophiocephalus (Zo). The 888
diet fractions pi,j refer to the target species' preys j=Phytoplankton (Fp), Bacterioplankton 889
(Bp), Zooplankton (Zp), Micro-Meiobenthos (MiMeB), Macrobenthos omnivorous filter-890
feeder (MaBoff), Macrobenthos omnivorous mixed-feeder (MaBomf), Carcinus 891
mediterraneus (Cm), Atherina boyeri (Ab). 892
893
Model output CB,i Input Factors
lindane in Tp αOC, νl,Tp/νn,Tp, β mP,Tp, LogKOW,25, σTp, pTp,j, ∆HOW, αDOC
PCB 15 in Tp LogKOW,25, νl,Tp/νn,Tp, β, αDOC, αOC, αPOC, σTp, mp,Tp
PCB 101 in Tp LogKOW,25, αDOC, αPOC, νl,Tp/νn,Tp, β, σTp, pTp,j, εl,Tp
PCB 194 in Tp LogKOW,25, αDOC, αPOC, σTp, wTp
lindane in Zo νl,Zo/νn,Zo, Log KOW,25, αOC, β
PCB 15 in Zo LogKOW,25, νl,Zo/νn,Zo, εn,Zo, αDOC, β
PCB 101 in Zo LogKOW,25, αDOC, εn,Zo, αPOC
PCB 194 in Zo LogKOW,25, αDOC, αPOC
Subset included in the Monte Carlo based UA/SA
Physical – chemical input factors
Log KOW,25 ∆HOW αOC αDOC αPOC β
Species-specific input factors
wi νl,i νn,i εl,i εn,i (for Tp and Zo)
pTp,Fp pTp,Bp pTp,Zp σTp mp,Tp (for Tp)
pZo,MiMeb pZo,MaBoff pZo,MaBomf pZo,Cm pZo,Ab (for Zo)
894
895
Table 4. Position and dispersion indexes of the model outputs’ distributions resulting from the 896
Monte Carlo experiments, relative to the concentrations of lindane and of three 897
polychlorinated biphenyls (PCB) congeners in i=Tapes philippinarum (Tp), Zosterisessor 898
ophiocephalus (Zo). 899
900
Model output CB,i Mean Value
(g/kg)
Standard deviation
(g/kg)
Coefficient of
Variation (%)
lindane in Tp 6.05E-09 2.89E-09 47.7
lindane in Zo 8.75E-09 2.15E-09 24.5
PCB 15 in Tp 4.25E-08 1.73E-08 40.7
PCB 15 in Zo 4.87E-07 3.17E-07 65.1
PCB 101 in Tp 1.54E-07 5.96E-08 38.6
PCB 101 in Zo 4.96E-06 2.85E-06 57.4
PCB 194 in Tp 2.54E-08 1.77E-08 69.7
PCB 194 in Zo 1.07E-07 1.27E-07 118.7

Table 5 Values of the fractions of variance explained by the input factors (Vi) and correspondent factors' rank obtained by means of the state 901
dependent regression (SDR) model, with respect the estimated concentrations of lindane and of three polychlorinated biphenyls (PCB) congeners in 902
i=Tapes philippinarum (Tp), Zosterisessor ophiocephalus (Zo). The diet fractions pi,j refer to the target species' preys j=Phytoplankton (Fp), 903
Bacterioplankton (Bp), Zooplankton (Zp), Micro-Meiobenthos (MiMeB), Macrobenthos omnivorous filter-feeder (MaBoff), ), Macrobenthos 904
omnivorous mixed-feeder (MaBomf), Carcinus mediterraneus (Cm), Atherina boyeri (Ab). In the last row, sum (Vi) and sum (Vl,i) are the total 905
fraction of variance explained by, respectively, the SDR model and the multiple linear regression (MLR) model. 906
907
Tapes philippinarum Zosterisessor ophiocephalus
Input Factor lindane PCB 15 PCB 101 PCB 194 Input Factor lindane PCB 15 PCB 101 PCB 194
rank Vi (%) rank Vi (%) rank Vi (%) rank Vi (%) rank Vi (%) rank Vi (%) rank Vi (%) rank Vi (%)
LogKOW,25 6 0.6 1 69.9 1 52.2 1 84.9 LogKOW,25 2 38.4 1 86.0 1 79.9 1 96.3
∆HOW 14 1.E-3 12 8.E-3 12 2.3.E-2 8 8.E-3 ∆HOW 12 8.E-3 10 0.11 14 6.E-3 7 1.3.E-2
αOC 1 71.4 6 2.00 13 2.0.E-2 14 1.1.E-3 αOC 3 5.8 9 0.12 16 1.E-4 16 6.E-6
αDOC 10 6.E-3 5 2.05 2 25.1 2 6.6 αDOC 8 2.1.E-2 3 1.4 2 6.9 2 1.3
αPOC 11 3.1.E-3 7 0.3 3 6.4 3 1.7 αPOC 14 3.E-4 7 0.20 4 2.0 3 0.4
β 2 9.1 2 12.3 4 6.2 7 3.7.E-2 β 4 2.8 5 0.7 9 3.E-2 10 5.E-3
νl,Tp 4 5.1 3 7.5 5 2.9 15 1.E-4 νl,Zo 1 44.9 2 4.4 5 1.5 4 5.E-2
νn,Tp 5 1.4 4 2.2 6 0.8 10 5.1.E-3 νn,Zo 5 0.2 13 1.E-2 11 1.E-2 14 1.E-3
εl,Tp 7 5.E-2 16 9.E-5 7 0.7 6 4.4.E-2 εl,Zo 11 1.E-2 4 0.9 3 3.8 5 3.E-2
εn,Tp 12 2.9.E-3 14 6.E-4 10 3.3.E-2 16 3.E-5 εn,Zo 7 2.4.E-2 8 0.19 7 0.4 9 6.E-3
wTp 16 7.E-5 13 6.E-3 15 3.E-4 5 0.2 wZo 6 3.E-2 11 0.1 15 1.E-4 15 4.E-4

pTp,Fp 15 7.E-4 15 1.E-4 14 1.E-3 12 2.7.E-3 pZo,MiMeb 16 3.E-7 6 0.4 6 1.4 6 1.3.E-2
pTp,Bp 8 2.E-2 10 2.E-2 16 2.E-4 9 5.5.E-3 pZo,MaBoff 9 1.7.E-2 12 2.E-2 8 0.2 8 7.E-3
pTp,Zp 13 2.E-3 11 1.E-2 8 0.52 11 3.0.E-3 pZo,MaBomf 10 1.3.E-2 14 2.1.E-3 13 7.E-3 13 3.8.E-3
σTp 9 1.E-2 8 0.09 9 0.46 4 0.4 pZo,Cm 13 4.E-3 15 1.8.E-3 10 2.E-2 12 3.9.E-3
mp,Tp 3 7.4 9 0.06 11 2.8.E-2 13 1.5.E-3 pZo,Ab 15 9.E-5 16 1.6.E-3 12 8.E-3 11 4.3.E-3
sum (Vi)
(sum (Vl,i))
95.1
(82)
96.6
(94)
95.4
(59)
94.0
(88)
sum (Vi)
(sum (Vl,i))
92.3
(91)
94.4
(87)
96.2
(61)
98.1
(75)
908


SETAC - SUPPLEMENTAL DATA ARCHIVE 909
Table S.1. Trophic compartments included in the food web of the Lagoon of Venice [24,25] 910
exploited to carry out the UA/SA of the Arnot and Gobas model [14] and corresponding 911
nominal values of the organism wet weights (wi), of the lipid fractions (νn,i) and nonlipid 912
organic matter (νn,i).The range of variability of these factors were set equal to ±30% the 913
nominal values. 914
915
Trophic
level
Compartment Label wi
(kg)
νl,i(kg/kg) νn,i(kg/kg)
Sediment/Detritus SD 2.57 5.0E-3 0.04
Phytoplankton Fp 3.2E-
3
5.0E-3 0.195*
Phytobentos Fb 6.5E-
12
5.0E-3 0.2*
Bacterioplankton Bp 3.0E-
5
5.0E-2 0.2
Plankton
Zooplankton Zp 3.4E-
5
5.0E-2 0.2
Micro-Meio benthos MiMeB 1.0E-
4
1.4E-2 0.2
Macrobenthos detritivorous MaBd 3.2E-
4
1.4E-2 0.2
Macrobenthos erbivorous/detritivorous MaBhd 2.5E-
5
1.4E-2 0.2
Macrobenthos omnivorous filter-feeder MaBoff 6.7E-
3
1.2E-2 0.2
Tapes philippinarum juveniles TpJ 1.0E-
3
1.3E-2 0.2
Tapes philippinarum Tp 7.0E-
3
1.3E-2 0.2
Macrobenthos omnivorous mixed-feeder MaBomf 1.4E-
3
2.6E-2 0.2
Benthos
Carcinus mediterraneus Cm 1.0E-
2
5.0E-2 0.2

Macrobenthos omnivorous predator MaBop 1.6E-
3
5.0E-2 0.2
Chelon labrosus juveniles ClJ 1.7E-
2
6.8E-2 0.2
Chelon labrosus Cl 2.7E-
2
6.8E-2 0.2
Atherina boyeri Ab 3.2E-
3
9.6E-2 0.2
Zosterisessor ophiocephalus Zo 1.6E-
1
1.0E-1 0.2
Nekton carnivorous benthic feeder Ncarbf 8.2E-
2
8.0E-2 0.2
Sparus aurata juveniles SaJ 4.3E-
2
9.7E-2 0.2
Sparus aurata Sa 1.3E-
1
9.7E-2 0.2
Dicentrarcus labrax juveniles DlJ 4.0E-
2
1.3E-1 0.2
Fish
Dicentrarcus labrax Dl 2.5E-
1
1.3E-1 0.2
* Nonlipid organic carbon fraction of autotrophic species (νn',i; kg/kg)
916

Table S.2. Dietary preference matrix for the trophic web of the Lagoon of Venice [24,25], relative to Phytoplankton (Fp), Phytobentos (Fb),
Bacterioplankton (Bp),Zooplankton (Zp), Micro-Meiobenthos (MiMeB), Macrobenthos detritivorous (MaBd), Macrobenthos
erbivorous/detritivorous (MaBhd), Macrobenthos omnivorous filter-feeder (MaBoff), Tapes philippinarum juveniles (TpJ), Tapes philippinarum
(Tp), Macrobenthos omnivorous mixed-feeder (MaBomf), Carcinus mediterraneus (Cm), Macrobenthos omnivorous predator (MaBop), Chelon
labrosus juveniles (ClJ), Chelon labrosus (Cl), Atherina boyeri (Ab), Zosterisessor ophiocephalus (Zo), Nekton carnivorous benthic feeder
(Ncarbf), Sparus aurata juveniles (SaJ), Sparus aurata (Sa), Dicentrarcus labrax juveniles (DlJ), Dicentrarcus labrax (Dl).
Pre
dato
r/
Pre
y
SD
Fp
Fb
Bp
Zp
MiM
eB
MaB
d
MaB
hd
MaB
off
TpJ
Tp
MaB
omf
Cm
MaB
op
ClJ
Cl
Ab
Zo
Nca
rbf
SaJ
Sa
DlJ
Dl
Fp
Fb
Bp 1**
Zp .5* .3 .2
MiMeB .99* .01
MaBd .66* .34
MaBhd .1 .9*
MaBoff .2 .5 .06 .24*
TpJ .52* .2 .22 .06
Tp .83* .075 .085 .01
MaBom
f .34* .25 .05 .23 .09 .04
Cm .25* .15 .2 .1 .04 .06 .1 .1
MaBop .25* .2 .04 .2 .16 .15

ClJ .69* .31
Cl .45* .11 .32 .12
Ab .38* .12 .28 .15 .01 .06
Zo .08 .44* .12 .23 .01 .12
Ncarbf .04 .41* .18 .15 .15 .01 .06
SaJ .52* .24 .12 .12
Sa .2 .22 .26* .21 .09 .005 .005 .005 .005
DlJ .12 .48* .18 .16 .01 .05
Dl .03 .05 .1 .52* .05 .05 .05 .08 .07
* Dependent input factor in the UA/SA analysis: the value is calculated as the difference between 1 and the sum of the other diet fractions relative
prey i
** Parameter not included among the input factors in the UA/SA

Table S.3 The first twenty highest values of the sensitivity index µ* estimated by means of the
Morris screening analysis with respect to the concentrations of lindane and three polychlorinated
biphenyls (PCBs) in Tapes philippinarum (Tp). Some of the input factors are relative to
Phytoplankton (Fp), Phytobentos (Fb), Bacterioplankton (Bp),Zooplankton (Zp). The symbol †
highlight the first µ* value of each sequence which is below to the corresponding threshold value in
the last row.
Rank lindane PCB 15 PCB 101 PCB 194
Factor µ* Factor µ* Factor µ* Factor µ*
1 αOC 6.5Ε−9 LogKOW,25 3.6Ε−8 LogKOW,25 1.12E-07 LogKOW,25 3.7Ε−8
2 νl,Tp/νn,Tp 2.5Ε−9 νl,Tp/νn,Tp 1.7Ε−8 αDOC 7.67E-08 αDOC 1.0Ε−8
3 β 2.5Ε−9 β 1.4Ε−8 αPOC 5.11E-08 αPOC 6.6Ε−9
4 mTp 2.2Ε−9 αDOC 6.1Ε−9 νl,Tp/νn,Tp 3.91E-08 σTp 1.9Ε−9
5 LogKOW,25 5.7Ε-10 αOC 4.4Ε−9 β 3.71E-08 wTp 1.4Ε−9
6 σTp 5.2Ε-11 αPOC 4.3Ε−9 σTp 1.55E-08 νl,Tp/νn,Tp 6.2Ε-10†
7 pTp 1.7Ε-11 σTp 1.6Ε−9 pTp 1.48E-08 ∆HOW 5.1Ε-10
8 ∆HOW 1.2Ε-11 mTp 1.0Ε−9 εl,Tp 1.23E-08 pTp 4.6Ε-10
9 αDOC 1.0Ε-11 νn,Bp 4.8Ε-10† νn,Bp 8.7E-09† β 4.5Ε-10
10 αPOC 5.4Ε-12† ∆HOW 4.6Ε-10 wTp 5.49E-09 εl,Tp 4.2Ε-10
11 εl,Tp 2.1Ε-12 εl,Tp 3.8Ε-10 νl,s 5.04E-09 αOC 2.7Ε-10
12 βa 1.1Ε-12 pTp 2.8Ε-10 εn,Tp 3.46E-09 mTp 1.5Ε-10
13 νn,Bp 8.2Ε-13 βa 2.0Ε-10 εl,Bp 3.12E-09 νl,s 1.3Ε-10
14 νl,s 6.9Ε-13 kgFp 1.4Ε-10 αOC 2.74E-09 εn,Tp 1.0Ε-10
15 εn,Tp 5.0Ε-13 νl,s 1.1Ε-10 νn,s 1.24E-09 εl,Bp 3.9Ε-11
16 νn',Fp 4.0Ε-13 εn,Tp 1.0Ε-10 νn,Zp 1.09E-09 νn,s 3.7Ε-11
17 wTp 3.2Ε-13 νn',Fp 7.9Ε-11 εn,Bp 9.76E-10 νn,Bp 2.6Ε-11
18 kgFp 2.7Ε-13 wTp 7.3Ε-11 mTp 9.12E-10 pZp 1.8Ε-11
19 νn,s 1.9Ε-13 νn,s 4.0Ε-11 pZp 8.8E-10 wBp 8.2Ε-12
20 νn,Zp 7.4Ε-14 wBp 3.7Ε-11 ∆HOW 7.88E-10 εn,Bp 5.6Ε-12
Thresholds 1.0E-11 1.0E-9 1.0E-8 1.0E-9

Table S.4 The first twenty highest values of the sensitivity index µ* estimated by means of the
Morris screening analysis with respect to the concentrations of lindane and three polychlorinated
biphenyls (PCBs) in Zosterisessor ophiocephalus (Tp). Some of the input factors are relative to
Zooplankton (Zp), Micro-Meiobenthos(MiMeB), Macrobenthos detritivorous (MaBd),
Macrobenthos omnivorous filter-feeder (MaBoff), Macrobenthos omnivorous mixed-feeder
(MaBomf) and Atherina boyeri (Ab). The symbol † highlight the first µ* value of each sequence
which is below to the corresponding threshold value in the last row.
Rank lindane PCB 15 PCB 101 PCB 194
Factor µ* Factor µ* Factor µ* Factor µ*
1 νl,Zo /νn,Zo 3.7Ε−9 LogKOW,25 7.1Ε−7 LogKOW,25 6.2Ε−6 LogKOW,25 2.5Ε−7
2 Log KOW,25 3.1Ε−9 νl,Zo /νn,Zo 1.4Ε−7 αDOC 1.7Ε−6 αDOC 3.1Ε−8
3 αOC 9.7Ε-10 εn,Zo 7.2Ε−8 εn,Zo 1.2Ε−6 αPOC 1.7Ε−8
4 β 9.1Ε-10 αDOC 6.8Ε−8 αPOC 1.2Ε−6 εn,Zo 9.4Ε−9†
5 νn,MaBa 1.7Ε-10† β 6.0Ε−8 νl,Zo /νn,Zo 8.1Ε−7† νn,MaBd 5.7Ε−9
6 mMaBd 1.6Ε-10 νn,Αb 4.1Ε−8† pZo 7.0Ε−7 νl,Zo /νn,Zo 5.6Ε−9
7 νn,MaBomf 1.6Ε-10 εw,Zo 3.5Ε−8 εw,Zo 6.7Ε−7 ∆HOW 4.1Ε−9
8 mMaBomf 1.5Ε-10 pZo 3.3Ε−8 εn,Αb 4.7Ε−7 νl,s 4.0Ε−9
9 wZo 9.8Ε-11 αPOC 3.3Ε−8 β 4.4Ε−7 pZo 3.7Ε−9
10 ∆HOW 6.7Ε-11 αOC 2.8Ε−8 νn,MaBd 4.1Ε−7 εl,MaBd 3.5Ε−9
11 pZo 5.9Ε-11 νn,MaBomf 2.1Ε−8 νn,MiMeB 3.2Ε−7 εw,Zo 3.5Ε−9
12 αDOC 5.7Ε-11 νn,MaBd 2.0Ε−8 εl,MaBe 3.2Ε−7 pMaBomf 3.3Ε−9
13 εn,Zo 5.4Ε-11 wZo 1.6Ε−8 εl,MiMeB 3.2Ε−7 pMaBd 2.4Ε−9
14 νn,Αb 4.6Ε-11 εn,Αb 1.4Ε−8 νn,Zp 3.1Ε−7 wZo 1.6Ε−9
15 mMaBoff 4.1Ε-11 νn,MaBoff 1.1Ε−8 νl,s 3.0Ε−7 νn,MiMeB 1.5Ε−9
16 mMiMeB 3.2Ε-11 ∆HOW 9.6Ε−9 pZp 2.6Ε−7 εn,MaBd 1.4Ε−9
17 νn,MiMeB 2.9Ε-11 νn,MiMeB 8.3Ε−9 pMaBd,i 2.6Ε−7 εl,MiMeB 1.2Ε−9
18 νn,MaBoff 2.7Ε-11 νn,Zp 8.0Ε−9 νn,MaBomf 2.0Ε−7 νn,MaBomf 1.2Ε−9
19 αPOC 2.6Ε-11 wMaBomf 6.2Ε−9 εl,MaBomf 1.8Ε−7 εn,Αb 1.0Ε−9
20 εw,Zo 2.3Ε-11 ba 6.1Ε−9 εw,Αb 1.7Ε−7 εl,MaBomf 9.3Ε-10
Thresholds 5.0E-10 5.0E-8 1.0E-6 1.0E-8

Table S.5 Values of the fractions of variance explained by the input factors (Vl,i) and correspondent factors' rank obtained by means of the multiple
linear regression (MLR) model, with respect the estimated concentrations of lindane and of three polychlorinated biphenyls (PCB) congeners in
i=Tapes philippinarum (Tp), Zosterisessor ophiocephalus (Zo). The diet fractions pi,j refer to the target species' preys j=Phytoplankton (Fp),
Bacterioplankton (Bp), Zooplankton (Zp), Micro-Meiobenthos (MiMeB), Macrobenthos omnivorous filter-feeder (MaBoff), Macrobenthos
omnivorous mixed-feeder (MaBomf), Carcinus mediterraneus (Cm), Atherina boyeri (Ab). In the last row, sum (Vl,i) is the total fraction of variance
explained by the MLR models.
Tapes philippinarum Zosterisessor ophiocephalus
Input Factor lindane PCB 15 PCB 101 PCB 194 Input Factor lindane PCB 15 PCB 101 PCB 194
rank Vl,i (%) rank Vl,i (%) rank Vl,i (%) rank Vl,i (%) rank Vl,i (%) rank Vl,i (%) rank Vl,i (%) rank Vl,i (%)
LogKOW,25 6 0.39 1 68.55 1 22.78 1 80.01 LogKOW,25 2 38.14 1 78.57 1 46.19 1 73.92
∆HOW 13 0.01 12 0.01 11 0.04 6 0.03 ∆HOW 10 0.00 12 0.05 11 0.05 9 0.02
αOC 1 58.38 6 1.59 14 0.01 10 0.01 αOC 3 4.63 8 0.23 15 0.00 7 0.03
αDOC 14 0.01 4 2.11 2 18.76 2 5.38 αDOC 7 0.03 3 1.29 2 4.91 2 0.48
αPOC 9 0.04 7 0.27 3 7.03 3 1.54 αPOC 15 0.00 7 0.28 4 2.61 3 0.31
β 2 8.50 2 12.17 4 5.25 7 0.02 β 4 2.54 5 0.52 9 0.16 16 0.00
νl,Tp 4 5.31 3 7.00 5 3.32 14 0.00 νl,Zo 1 45.48 2 4.40 5 1.45 8 0.02
νn,Tp 5 1.53 5 2.10 6 1.27 8 0.02 νn,Zo 5 0.27 13 0.03 8 0.18 12 0.01
εl,Tp 7 0.15 16 0.00 8 0.34 16 0.00 εl,Zo 11 0.00 4 0.87 3 3.91 10 0.01
εn,Tp 8 0.06 14 0.00 12 0.01 13 0.00 εn,Zo 9 0.01 9 0.14 7 0.57 4 0.08
wTp 16 0.00 15 0.00 13 0.01 5 0.23 wZo 6 0.03 10 0.12 13 0.01 5 0.05

pTp,Fp 15 0.00 11 0.01 15 0.00 11 0.01 pZo,MiMeb 13 0.00 15 0.00 16 0.00 6 0.04
pTp,Bp 10 0.03 10 0.01 7 0.35 9 0.02 pZo,MaBoff 12 0.00 16 0.00 10 0.08 11 0.01
pTp,Zp 11 0.01 13 0.00 16 0.00 12 0.00 pZo,MaBomf 8 0.03 11 0.06 12 0.04 13 0.00
σTp 12 0.01 8 0.11 9 0.15 4 0.44 pZo,Cm 14 0.00 14 0.00 14 0.01 15 0.00
mp,Tp 3 7.24 9 0.06 10 0.05 15 0.00 pZo,Ab 16 0.00 6 0.38 6 0.59 14 0.00
sum (Vl,i) 81.7 94.0 59.4 87.7 sum (Vl,i) 91.2 86.9 60.8 75.0

54
Table S.6 The first twenty highest values of the sensitivity index µ* estimated by means of 1
the Morris screening analysis when the ranges of variability of the species specific input 2
factor were increased to 50% the reference values, with exception of the factors εl,i and εn,i 3
higher than 0.67 which were kept to ±30%. The model outputs are the concentrations of 4
lindane and three polychlorinated biphenyls (PCBs) in Tapes philippinarum (Tp). Some of the 5
input factors are relative to Phytoplankton (Fp), Phytobenthos (Fb), Bacterioplankton 6
(Bp),Zooplankton (Zp). The symbol † highlight the first µ* value of each sequence which is 7
below to the corresponding threshold value in the last row. 8
Rank lindane PCB 15 PCB 101 PCB 194
Factor µ* Factor µ* Factor µ* Factor µ*
1 αOC 5.7E-09 LogKOW,25 3.9E-08 LogKOW,25 9.0E-08 LogKOW,25 3.5E-08
2 mTp 3.4E-09 νn,Tp 2.0E-08 αDOC 8.5E-08 αDOC 5.9E-09
3 νl,Tp/νn,Tp 3.4E-09 β 1.7E-08 αPOC 4.6E-08 αPOC 4.1E-09
4 β 2.0E-09 αOC 6.6E-09 β 3.8E-08 wTp 3.0E-09
5 LogKOW,25 5.6E-10 αDOC 5.6E-09 νl,Tp/νn,Tp 3.5E-08 σTp 2.4E-09
6 σTp 4.3E-11 mTp 3.5E-09 σTp 1.5E-08 νl,Tp/νn,Tp 1.5E-09
7 pTp 2.6E-11 αPOC 3.2E-09 pTp 1.2E-08 pTp 1.1E-09
8 ∆HOW 1.3E-11 σTp 1.4E-09 νn,Bp 8.7E-09† εl,Tp 7.4E-10†
9 αDOC 1.2E-11 νn,Bp 8.3E-10† νl,s 8.0E-09 ∆HOW 5.0E-10
10 αPOC 5.0E-12† pTp 7.3E-10 εl,Tp 6.9E-09 β 4.9E-10
11 εl,Tp 2.3E-12 ∆HOW 5.1E-10 wTp 6.4E-09 νl,s 3.3E-10
12 νl,s 1.8E-12 εl,Tp 4.9E-10 εn,Tp 3.3E-09 mTp 2.4E-10
13 νn,BB 1.2E-12 νl,s 2.2E-10 εl,Bp 2.7E-09 αOC 1.7E-10
14 βa 1.0E-12 βa 1.9E-10 αOC 2.4E-09 εn,Tp 1.2E-10
15 εn,Tp 9.3E-13 νn',Fp 1.5E-10 νn,s 2.4E-09 νn,Bp 7.1E-11
16 νn',Fp 5.8E-13 kgFp 1.4E-10 νn,Zp 2.1E-09 εl,Bp 6.7E-11
17 wTp 4.7E-13 εn,Tp 1.4E-10 kgFp 1.8E-09 νn,s 6.0E-11
18 νn,s 2.8E-13 vl,Zp 1.0E-10 pZp 1.5E-09 pZp 2.1E-11
19 kgFp 2.6E-13 νn,s 8.6E-11 ∆HOW 1.1E-09 νn,Zp 1.9E-11
20 νn,Zp 1.3E-13 wTp 6.95E-11 wBB 1.1E-09 wBp 1.3E-11
Thresholds 1.0E-11 1.0E-9 1.0E-8 1.0E-9

55
Table S.7 The first twenty highest values of the sensitivity index µ* estimated by means of 9
the Morris screening analysis when the ranges of variability of the species specific input 10
factor were increased to 50% the reference values, with exception of the factors εl,i and εn,i 11
higher than 0.67 which were kept to ±30%. The model outputs are the concentrations of 12
lindane and three polychlorinated biphenyls (PCBs) in Zosterisessor ophiocephalus (Tp). 13
Some of the input factors are relative to Zooplankton (Zp), Micro-Meiobenthos(MiMeB), 14
Macrobenthos detritivorous (MaBd), Macrobenthos omnivorous filter-feeder (MaBoff), 15
Macrobenthos omnivorous mixed-feeder (MaBomf) and Atherina boyeri (Ab). The symbol † 16
highlight the first µ* value of each sequence which is below to the corresponding threshold 17
value in the last row. 18
Rank lindane PCB 15 PCB 101 PCB 194
Factor µ* Factor µ* Factor µ* Factor µ*
1 νl,Zo /νn,Zo 6.1E-09 LogKOW,25 5.6E-07 LogKOW,25 5.1E-06 LogKOW,25 2.4E-07
2 Log KOW,25 3.4E-09 νl,Zo /νn,Zo 2.8E-07 αDOC 1.6E-06 αDOC 3.4E-08
3 αOC 1.1E-09 pZo 6.1E-08 νl,Zo /νn,Zo 1.5E-06 νl,Zo /νn,Zo 1.1E-08
4 β 8.5E-10 νn,MaBomf 4.9E-08† εn,Zo 1.0E-06 αPOC 1.1E-08
5 νn,MaBa 3.1E-10† νn,Αb 4.8E-08 εw,Zo 8.6E-07† νn,MaBd 5.3E-09†
6 mMaBd 3.0E-10 νn,MaBd 4.6E-08 αPOC 7.9E-07 pZo 5.0E-09
7 mMaBomf 2.8E-10 β 4.4E-08 pZo 7.1E-07 ∆HOW 4.6E-09
8 νn,MaBomf 2.4E-10 αDOC 4.1E-08 νn,Zp 5.6E-07 νl,s 3.7E-09
9 wZo 2.0E-10 εn,Zo 3.8E-08 νn,MaBd 5.0E-07 εw,Zo 3.2E-09
10 pZo 1.3E-10 εw,Zo 2.8E-08 νn,Αb 4.9E-07 β 2.9E-09
11 mMaBoff 9.1E-11 νn,Zp 2.2E-08 εn,Ab 4.6E-07 νn,MiMeB 2.7E-09
12 νn,Αb 8.9E-11 αOC 1.9E-08 νl,s 4.0E-07 pMaBomf 2.6E-09
13 νn,MiMeB 7.4E-11 wZo 1.8E-08 εw,Αb 3.8E-07 εl,MiMeB 2.2E-09
14 αDOC 7.2E-11 αPOC 1.6E-08 pMaBd,i 3.3E-07 νn,s 1.6E-09
15 ∆HOW 6.8E-11 νn,MiMeB 1.2E-08 pMaBomf 2.9E-07 wZo 1.6E-09
16 εn,Zo 6.7E-11 εw,Ab 1.1E-08 pZp 2.7E-07 εl,MaBd 1.6E-09
17 νn,MaBoff 6.3E-11 νn,MaBoff 9.9E-09 εl,MaBd 2.7E-07 wMaBd 1.6E-09
18 mMiMeB 5.4E-11 wAb 8.2E-09 pAb 2.6E-07 pZp 1.5E-09
19 εw,Zo 3.9E-11 ∆HOW 7.2E-09 vn,MaBomf 2.4E-07 pMaBd 1.5E-09
20 αPOC 2.9E-11 νn,MaBoff 6.7E-09 β 2.4E-07 εn,Zo 1.5E-09

56
Thresholds 5.0E-10 5.0E-8 1.0E-6 1.0E-8

57
19
Table S.8. Position and dispersion indexes of the model outputs’ distributions resulting from 20
the Monte Carlo experiments where all the 227 model parameters were randomly sampled. 21
The indexes are relative to the concentrations of lindane and of three polychlorinated 22
biphenyls (PCB) congeners in Tapes philippinarum (Tp) and Zosterisessor ophiocephalus 23
(Zo). 24
25
Model output Cb Mean Value
(g/kg)
Standard deviation
(g/kg)
Coefficient of
Variation (%)
lindane in Tp 6.05E-09 2.89E-09 47.7
lindane in Zo 8.75E-09 2.15E-09 24.5
PCB 15 in Tp 4.25E-08 1.72E-08 40.6
PCB 15 in Zo 4.87E-07 3.23E-07 66.3
PCB 101 in Tp 1.55E-07 6.00E-08 38.8
PCB 101 in Zo 4.97E-06 2.91E-06 58.5
PCB 194 in Tp 2.54E-08 1.77E-08 69.7
PCB 194 in Zo 1.07E-07 1.26E-07 118.5
26





























