Embed Size (px)
Citation preview

Plant Cell and Environment
(2003)
26
771ndash782
copy 2003 Blackwell Publishing Ltd
771
Blackwell Science LtdOxford UKPCEPlant Cell and Environment0016-8025Blackwell Science Ltd 2003 2003
26771782Original Article
Cotton responses to elevated CO
2
and UV-B radiationD Zhao
et al
Correspondence
K R Reddy Fax
+
1 662 325 9461 e-mailkrreddyramsstateedu
Growth and physiological responses of cotton (
Gossypium hirsutum
L) to elevated carbon dioxide and ultraviolet-B radiation under controlled environmental conditions
D ZHAO
1
K R REDDY
1
V G KAKANI
1
J J READ
2
amp J H SULLIVAN
3
1
Department of Plant and Soil Sciences Mississippi State University Box 9555 Mississippi State MS 39762 USA
2
USDA-ARS Crop Science Research Laboratory Mississippi State MS 39762 USA and
3
Department of Natural Resource Sciences and Landscape Architecture University of Maryland College Park MD 20742 USA
ABSTRACT
Better understanding of crop responses to projectedchanges in climate is an important requirement An exper-iment was conducted in sunlit controlled environmentchambers known as soilndashplantndashatmospherendashresearch unitsto determine the interactive effects of atmospheric carbondioxide concentration [CO
2
] and ultraviolet-B (UV-B)radiation on cotton (
Gossypium hirsutum
L) growthdevelopment and leaf photosynthetic characteristics Sixtreatments were used comprising two levels of [CO
2
] (360and 720
mmmm
mol mol----
1
) and three levels of 0 (control) 77 and151 kJ m----
2
d----
1
biologically effective UV-B radiationswithin each CO
2
level Treatments were imposed for 66 dfrom emergence until 3 weeks after the first flower stagePlants grown in elevated [CO
2
] had greater leaf area andhigher leaf photosynthesis non-structural carbohydratesand total biomass than plants in ambient [CO
2
] Neitherdry matter partitioning among plant organs nor pigmentconcentrations was affected by elevated [CO
2
] On theother hand high UV-B (151 kJ m----
2
d----
1
) radiation treat-ment altered growth resulting in shorter stem and branchlengths and smaller leaf area Shorter plants at high UV-Bradiation were related to internode lengths rather than thenumber of mainstem nodes Fruit dry matter accumulationwas most sensitive to UV-B radiation due to fruit abscis-sion Even under 77 kJ m----
2
d----
1
of UV-B radiation fruit dryweight was significantly lower than the control althoughtotal biomass and leaf photosynthesis did not differ fromthe control The UV-B radiation of 151 kJ m----
2
d----
1
reducedboth total (43) and fruit (88) dry weights due to smallerleaf area and lower leaf net photosynthesis Elevated [CO
2
]did not ameliorate the adverse effects of UV-B radiationon cotton growth and physiology particularly the bollretention under UV-B stress
Key-words
Gossypium hirsutum
L CO
2
yen
UV-B interac-tive effects dry matter production and partitioning non-structural carbohydrates pigments photosynthesis
INTRODUCTION
Continued depletion of the stratospheric ozone layermainly due to recent increases in atmospheric chlorofluro-carbons (CFCs) methane and nitrous oxide is of concernbecause the ozone column is the primary attenuator ofsolar UV-B radiation (280ndash320 nm) Reductions in theozone column have led to substantial increases in UV-Bradiation at the surface of the Earth with the amountdependent on atmospheric and geographic factors (WMO1995 Madronich
et al
1998) Intensified current and pro-jected UV-B radiation is known to alter and affect plantgrowth development and physiological processes (Dai
et al
1992 Gonzalez
et al
1996 Nogues
et al
1999) Dam-age due to UV-B radiation results in inhibition of photo-synthesis degradation of protein and DNA and increasedoxidative stress (Jordan Chow amp Anderson 1992 Stapleton1992 Sullivan 1997) In general enhanced UV-B radiationaffects plant growth andor yield of most crops (KrupaKickert amp Jaumlger 1998) The sensitivity of plants to UV-Bradiation however varies among species and is influencedby environmental factors such as water regime photosyn-thetically active radiation (PAR) and nutrient status(Murali amp Teramura 1985 Balakumar Hani amp Paliwal1993 Mark amp Tevini 1996) As in other crops enhancedUV-B radiation may alter cotton growth and developmentSong
et al
(1999) found that enhanced UV radiationreduced growth and yield in field-grown cotton
Along with increase in UV-B radiation climate projec-tions indicate changes in other major environmental factorssuch as the increase in atmospheric carbon dioxide concen-tration [CO
2
] temperature and variability in precipitation(Houghton
et al
2001) The [CO
2
] in the atmosphere hasincreased by more than 28 since the beginning of theIndustrial Revolution mainly because of the burning offossil fuels and deforestation The most recent future sce-narios of greenhouse gases in the atmosphere indicate that[CO
2
] could increase from current levels of approximately360
m
mol mol
-
1
to between 540 and 970
m
mol mol
-
1
by theend of the twenty-first century (Houghton
et al
2001) Ofthe environmental variables CO
2
is an important green-house gas and its increase in the atmosphere is known toincrease C
3
-type crop yields (Kimball 1983) Elevated
772
D Zhao
et al
copy 2003 Blackwell Publishing Ltd
Plant Cell and Environment
26
771ndash782
atmospheric [CO
2
] generally enhances leaf and canopy CO
2
assimilation rates because CO
2
is not only the substrate forphotosynthesis but also a competitive inhibitor of oxygen-ation of RuBP carboxylase and reduces photorespiration(Lawlor amp Mitchell 2000)
The response of crop plants to UV-B radiation in thepresence of other environmental stress factors is not clearand has not received much attention Studies conducted sofar indicate that effects of UV-B radiation with other envi-ronmental variables such as enhanced [CO
2
] (TeramuraSullivan amp Ziska 1990 Rozema
et al
1997 Visser
et al
1997) high temperature (Mark amp Tevini 1996) water defi-cit (Nogues
et al
1998 Alexieva
et al
2001) are both addi-tive and interactive Few studies have evaluated the effectsof UV-B radiation on the growth and development of cot-ton an important agronomic crop grown in a wide rangeof environmental conditions spanning latitudes 40
infin
N and40
infin
S although the influences of other environmental fac-tors such as atmospheric [CO
2
] temperature water andnutrients on plant growth and development have beenwell investigated in cotton (Christiansen 1986 GerikOosterhuis amp Torbet 1998 Reddy Hodges amp Kimball2000)
Reddy
et al
(2000) reported that canopy photosynthesisof cotton increased by 40 when [CO
2
] was increased from320 to 720
m
mol mol
-
1
for plants grown at 20 and 32
infin
C andincreased by about 80 when plants were grown underoptimum temperature (26ndash28
infin
C) conditions Kimball ampMauney (1993) found that cotton grown under550
m
mol CO
2
mol
-
1
had a 35 higher biomass 40 higherfruit weight and 60 higher lint yield than plants grownunder 350
m
mol CO
2
mol
-
1
under free-air CO
2
enrichmentstudies However there is a significant knowledge gap onthe interactive effects of UV-B radiation and [CO
2
] onplants We hypothesize that the CO
2
enhancement effectmay be reduced by concurrent exposure of plants to ele-vated UV-B radiation The objective of this study was thusto determine the interactive effects of elevated [CO
2
] andUV-B radiation on cotton growth development and phys-iology under controlled environment conditions Informa-tion related to whole plant and leaf-level processes will
contribute to filling gaps in our knowledge base andunderstanding
MATERIALS AND METHODS
Soilndashplantndashatmosphere research units
The experiment was conducted at the Agriculture and For-estry Experiment Station of Mississippi State (33
infin
28
cent
N88
infin
47
cent
W) Mississippi USA in 2001 using six soilndashplantndashatmosphere research (SPAR) units The SPAR facility hasthe capability to precisely control temperature and [CO
2
]at predetermined set points for plant growth studies undernear ambient levels of PAR Details of the SPAR units havebeen described by Reddy
et al
(2001) Each SPAR unitconsists of a steel soil bin (1 m deep by 2 m long by 05 mwide) and a Plexiglas chamber (25 m tall by 2 m long by15 m wide) to accommodate aerial plant parts a heatingand cooling system and an environment monitoring andcontrol system The Plexiglas chamber is completelyopaque to solar UV radiation but transmits more than 95of incoming PAR (wavelength 400ndash700 nm) (Table 1) Dur-ing the experiment the incoming solar radiation (285ndash2800 nm) outside the SPAR units was measured with apyranometer (Model 4ndash48 The Eppley Laboratory IncNewport RI USA) The average daily total solar radiationduring the experiment was 18
plusmn
49 MJ m
-
2
d
-
1
The air tem-perature and [CO
2
] in each SPAR unit were monitored andadjusted every 10 s throughout the experiment
The dew point temperatures were monitored with goldmirror hygrometers installed inside the return airline andwere also collected at 10 s intervals and summarized over900 s intervals The data were separated into day and nightperiods The daytime vapour pressure deficit (VPD) of eachtreatment was calculated based on air temperature and dewpoint temperature The PAR transmittance of all chamberswere determined on two typical clear days between 1000and 1400 h using seven 1 m line quantum sensors (Li-CorInc Lincoln NE USA) Daily mean temperatures aver-age daytime VPD and PAR transmittances of all treat-ments are presented in Table 1
Table 1
Measured [CO
2
] and Plexiglas transmittance of photosynthetically active radiation (PAR) from a typical day daily mean UV-B radiation dosage mean temperature (
T
) and daytime vapour pressure deficit (VPD) during the experimental period for each treatment
Treatment
CO
2
(
m
mol mol
-
1
)Mean UV-B(kJ m
-
2
d
-
1
)Mean T(
infin
C)VPD(kPa)
Transmittanceof PAR ()
CO
2
(
m
mol mol
-
1
)UV-B(kJ m
-
2
d
-
1
)
360 00 3597
plusmn
07 000
plusmn
000 2658
plusmn
004 168
plusmn
028 962
plusmn
0677 3613
plusmn
18 763
plusmn
005 2659
plusmn
003 196
plusmn
015 963
plusmn
05151 3601
plusmn
07 1503
plusmn
008 2645
plusmn
009 159
plusmn
008 973
plusmn
05720 00 7231
plusmn
10 000
plusmn
000 2659
plusmn
003 164
plusmn
020 986
plusmn
0277 7250
plusmn
14 774
plusmn
004 2659
plusmn
003 189
plusmn
021 957
plusmn
05151 7233
plusmn
10 1512
plusmn
009 2650
plusmn
005 179
plusmn
019 960
plusmn
05
Each value represents the mean plusmn SE of data of 40 values from a day for [CO2] and PAR transmittance or 65 d from 6 August to 10 October2001 for UV-B radiation VPD and temperature
Cotton responses to elevated CO2 and UV-B radiation 773
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Plant culture
Seeds of cotton cv NuCOTN 33B were sown on 1 August2001 in fine sand of the SPAR soil bins in 11 rows of fiveplants per row Emergence date was 5 d later The temper-atures in all units were maintained at 3022 infinC (daynight)during the experiment Plants were watered three times aday with half-strength Hoaglands nutrient solution deliv-ered at 0800 1200 and 1700 h to ensure favourable nutrientand water conditions for plant growth Irrigation was pro-vided through an automated and computer-controlled dripsystem Variable-density black shade cloths around theedges of plants were adjusted regularly to match plantheight in order to simulate natural shading in the presenceof other plants
Treatments
The six treatments included two CO2 levels of360 mmol mol-1 (ambient) and 720 mmol mol-1 (elevated)and three daily biologically effective UV-B radiation inten-sities of 0 (control) 77 and 151 kJ m-2 within each CO2
level The [CO2] and UV-B radiation treatments wereimposed from emergence The UV-B doses of 77 and151 kJ m-2 d-1 are designed to simulate the maximum levelof UV-B that would be received on a clear day on thesummer solstice with either ambient levels of ozone or witha 30 ozone depletion in Mississippis cotton productionregion as calculated by an empirical model (Green Crossamp Smith 1980)
Square-wave UV-B supplementation systems were usedto provide respective UV-B radiation in this study undernear ambient PAR The UV-B radiation was delivered toplants for 8 h each day from 0800 to 1600 h by eight fluo-rescent UV-313 lamps (Q-Panel Company Cleveland OHUSA) driven by 40 W dimming ballasts The lamps werewrapped with pre-solarized 008 mm cellulose diacetatefilm to filter UV-C (lt 280 nm) radiation The cellulose diac-etate film was changed at 3ndash4 d intervals The amount ofUV-B energy delivered at the top of the plant canopy waschecked daily at 1000 hours with a UVX digital radiometer(UVP Inc San Gabriel CA USA) and calibrated againstan Optronic Laboratory (Orlando FL USA) Model 754Spectroradiometer which was used to initially quantifylamp output The lamp power was adjusted as needed tomaintain the appropriate UV-B radiation level in eachtreatment and to maintain the distance from lamps to thetop of plants at about 05 m throughout the experiment Themean UV-B dosages measured at three different locationsin each SPAR unit corresponding to the permanent rowsfor all treatments during the experimental period are pre-sented in Table 1 After averaging the radiation dosages forthe plants within UV-B treatments the actual dosages forthe two levels of UV-B treatments were 77 and 151 kJ m-2
d-1 respectively Square-wave UV-B supplemental systemsin controlled environments usually provide higher UV-Bradiation and disproportionate spectral conditions com-pared to ambient condition (Musil et al 2002) and there is
no UV-B supplied in the control treatment This is likely toexaggerate the UV-B response in plants and thus cautionwill be necessary in extrapolating the findings from thisstudy to the field
Measurements
Plant height and the number of mainstem nodes weredetermined from nine plants (three centre plants in eachrow) at 3 d intervals from 10 to 66 d after emergence(DAE) These plants were also used to record dates ofappearance of the first floral bud of 3 mm in length (firstsquare FS) and first flower (FF) and durations in daysfrom emergence to FS and from FS to FF The plants wereharvested at the fourth true leaf stage (14 DAE) earlyfloral bud stage (29 DAE) and finally at 3 weeks after thefirst flower stage (66 DAE) Plants in the odd rows of eachSPAR unit were harvested at 14 DAE The five rows afterthe first sampling were spaced 33 cm apart The two evenrows were harvested 29 DAE and the remaining three rows(with a 67 cm row space) were finally harvested 66 DAEPlants were separated into leaves and stems at the first twoharvests and the leaf area and dry weights of leaves andstems were determined At the final harvest plant heightfruiting branch lengths numbers of mainstem nodes fruit-ing branches fruiting sites squares and bolls on each plantwere recorded The rooting medium was removed and rootswashed over a fine screen dried and weighed Mean inter-node length was calculated by dividing plant height by thenumber of mainstem nodes Fruit (squares + bolls) abscis-sion was calculated as the difference between the numberof fruiting sites and the number of fruits
Net photosynthetic rates (Pn) of the uppermost fullyexpanded mainstem leaf from five plants in each treatmentwere measured between 1000 and 1200 h using a LI-6400portable photosynthesis system (Li-Cor Inc) at 28 44 and64 DAE When measuring leaf Pn the PAR provided by a6400ndash02 LED light source was set to 1500 mmol m-2 s-1 Thetemperature in the leaf cuvette was set to 30 infinC and [CO2]matched CO2 treatments
Chlorophyll a chlorophyll b carotenoid and non-struc-tural carbohydrate (including glucose fructose sucroseand starch) concentrations of the leaves at the same posi-tion were also determined at 45 and 65 DAE For chloro-phyll measurement five leaf discs (0385 cm2 each) werepunched from a leaf and placed in a vial with 4 mL ofdimethyl sulphoxide Three samples were collected fromeach treatment and incubated at room temperature for24 h in the dark to allow for complete extraction of chlo-rophyll into the solution The absorbance of the extract wasmeasured using a Pharmacia UltraSpec Pro UVVIS spec-trophotometer (Pharmacia Cambridge UK) at 470 648and 664 nm and used to calculate chlorophyll a chlorophyllb and carotenoid concentrations (Chappelle Kim ampMcMurtrey 1992)
Five leaf discs from each leaf were collected at 0830 husing the same method described above and placed in a testtube with 2 mL of 80 (vv) cold ethanol Three leaf sam-
774 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
ples were obtained from each treatment and immediatelystored at -80 infinC to determine leaf non-structural carbohy-drates The modified method of Hendrix (1993) was usedto extract and quantify the concentrations of leaf non-structural carbohydrates Briefly soluble sugars wereextracted by incubating leaf tissue with 2 mL of the ethanolin an 80 infinC water bath for 15 min The extraction wasrepeated three times and the supernatants were combinedand brought to a 6 mL volume with 80 ethanol Sixtymilligrams of finely ground activated charcoal was addedto each sample tube covered and shaken by hand After5 min the tubes were centrifuged at 3000 yen g for 15 min toobtain a clear extract Four 40 mL aliquots from each sam-ple were pipetted into separate wells of a microtitrationplate and dried at 50 infinC for 30 min to remove the ethanolEight wells in the first column of each microtitration platewere filled with 20 mL glucose standard solutions of 0 5 1325 50 125 250 and 500 mg L-1 respectively To each samplewell 20 mL deionized water was added and then 100 mLglucose-6P dehydrogenaseiodonitrotetrazolium violet(INT) mixture (glucose kit 115 A Sigma Chemical Com-pany St Louis MO USA) was added under reduced roomillumination Plates were incubated at 37 infinC for 15 min andthe absorbance measured at 490 nm with an Emax preci-sion microplate reader (Emax Sunnyvale CA USA) Aglucose standard curve was obtained for each plate basedon the standard concentrations and absorbance Sampleglucose concentration was calculated according to the stan-dard curve absorbance extract volume and leaf disc areaSubsequently 10 mL phosphoglucose isomerase (PGIenzyme 025 units) and 10 mL invertase (83 units) wereadded to each well separately re-incubated at 37 infinC for15 min each time The absorbance was determined at490 nm to obtain concentrations of fructose and sucrose
After extraction of the soluble sugars 1 mL of 01 MKOH was added to the test tube containing the tissue res-idue and the tubes placed for 1 h in a boiling waterbathAn a-amylase and amyloglucosidase preparation was usedto hydrolyse starch to glucose as described by Hendrix(1993) After centrifuging the hydrolysate the supernatantwas collected and the glucose concentration determined asabove Starch concentration in the sample was calculatedaccording to the glucose concentration multiplied by 09 toaccount for water loss when glucose units are linked Thesum of glucose fructose sucrose and starch was defined astotal non-structural carbohydrate
Data analysis
A uniformity test of the SPAR units conducted in a previ-ous study by determining sorghum growth parametersunder the same controlled environments indicated no sta-tistical differences among all SPAR units (Reddy personalcommunication 2000) Therefore six treatments were ran-domly arranged in six identical SPAR units Except for thetwo treatment factors of [CO2] and UV-B radiation theother growth conditions were the same in all treatments(Table 1) All growth measurements were made on nine
plants and physiological measurements were made oneither three or five plants for each treatment Linear andsigmoid regressions were performed for the number ofmainstem nodes and plant height respectively against daysafter emergence Data were analysed by the analysis ofvariance (ANOVA) procedures in SAS (SAS Institute Inc1997) to determine the main and interactive effects of[CO2] and UV-B radiation on growth dry matter accumu-lation leaf Pn and concentrations of leaf pigments and non-structural carbohydrates If the hypothesis of equal meanswas rejected by the ANOVA test Fisher LSD procedures atP = 005 probability level (SAS Institute Inc 1997) wereemployed to distinguish differences among treatmentmeans for the growth and physiological processesmeasured
RESULTS
Plant growth and development
Plants grown under 720 mmol CO2 mol-1 were typically 25ndash50 cm taller than those grown in 360 mmol CO2 mol-1depending on UV-B exposure (Fig 1) Changes in plantheight from emergence up to 3 weeks after first floweringfollowed a sigmoid growth pattern The maximum main-stem elongation rate was achieved between 35 and 45 DAEfor all treatments but the elongation rates differed substan-tially among the treatments Plants irradiated with151 kJ m-2 d-1 UV-B radiation had significantly lower stemelongation rates in both CO2 levels compared to plants ineither the 0 or 77 kJ m-2 d-1 UV-B treatments At the finalharvest (66 DAE) plants grown in 720 mmol CO2 mol-1
were 22 taller than plants grown under360 mmol CO2 mol-1 averaged across the UV-B treatmentsPlants irradiated with UV-B radiation were significantlyshorter than the controls (except for 77 kJ UV-B treatmentat 360 mmol CO2 mol-1)
Unlike plant height mainstem nodes increased linearly(R2 = 099) as plants aged in all treatments (data notshown) The node appearance rates (slopes of the lines) didnot differ among the UV-B treatments but elevated [CO2]caused a slight increase in node addition rate (Y = 033X - 128 for 360 mmol CO2 mol-1 and Y = 036X - 161 for720 mmol CO2 mol-1 where Y is the number of mainstemnodes and X is days after emergence) Averaged across theUV-B treatments it took about 30 d (slope = 033) to pro-duce a leaf (or node) on the main stem under ambient[CO2] and about 28 d (slope = 036) under elevated [CO2]At the final harvest mean internode lengths of the 0 77and 151 kJ UV-B-treated plants were 85 79 and 45 cmrespectively under ambient [CO2] and 94 83 and 54 cmrespectively under elevated [CO2]
No CO2 yen UV-B radiation interaction effect was detectedfor leaf area development (Table 2) Plants grown in ele-vated [CO2] had significantly greater leaf area than plantsin ambient [CO2] at 14 DAE Leaf area was reduced by UV-B treatments of 77 and 151 kJ m-2 d-1 at 24 DAE At 66DAE only plants exposed to 151 kJ UV-B had significantly
Cotton responses to elevated CO2 and UV-B radiation 775
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
smaller leaf area (47ndash50) than plants in other treatmentsof CO2 and UV-B radiation
Neither elevated [CO2] nor UV-B radiation affected thedates of appearance of the first floral buds and flowers (datanot shown) A significant CO2 yen UV-B interactive effectwas detected for the number of fruiting branches branchlengths and fruit abscission (Table 3) Elevated [CO2] sig-nificantly increased the numbers of fruiting branches fruit-ing sites and squares per plant and mean branch length butdid not affect the number of bolls retained per plant Incontrast to [CO2] UV-B radiation significantly decreasedmost reproductive growth parameters except for the num-
ber of fruiting branches Fruit abscission in UV-B-treatedplants was significantly higher than in control plants in bothambient and elevated CO2 levels Floral bud abscission wasmore than young boll abscission when plants were grownunder UV-B radiation (data not shown)
Dry matter accumulation and partitioning
Elevated [CO2] significantly increased dry matter accumu-lation in leaves and stems at all sampling dates (Tables 4 amp5) Dry weights of leaves and stems did not differ amongthe UV-B radiation treatments at 14 DAE but at 24 DAE
Figure 1 Changes in plant height and stem elongation rate of cotton plants grown under 360 and 720 mmol CO2 mol-1 as affected by UV-B radiation Each data point represents the mean plusmn SE of nine plants The lines are regressions fitted to the data using a sigmoid equation for plant height and a first deriva-tive of the sigmoid equation for stem elon-gation rate
Table 2 Effects of elevated CO2 and UV-B radiation on leaf area (cm2 plant-1) of cotton plant 14 24 and 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Day after emergence
14 24 66
360 00 98 plusmn 6 ab 589 plusmn 48 b 6111 plusmn 627 c77 87 plusmn 3 bc 471 plusmn 50 c 6776 plusmn 331 bc
151 80 plusmn 6 c 455 plusmn 27 c 3064 plusmn 251 e720 00 102 plusmn 6 a 721 plusmn 32 a 8456 plusmn 432 a
77 104 plusmn 4 a 546 plusmn 42 bc 7738 plusmn 440 ab151 110 plusmn 7 a 588 plusmn 31 b 4502 plusmn 354 d
P (CO2) 00002 00011 lt 00001P (UV-B) 05991 00009 lt 00001P (CO2 yen UV-B) 00702 07166 02611
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
776 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
total dry weights of 77 and 151 kJ UV-B-treated plantswere 21 and 30 lower than the control plants respec-tively At the final harvest plants under 720 mmol CO2
mol-1 had a 36 higher total biomass than plants under360 mmol CO2 mol-1 averaged across the three UV-B treat-ments Total biomass produced was lowered by 12 for the77 kJ UV-B treatment and by 69 for the 151 kJ UV-Btreatment at 66 DAE compared to the control (Table 5)Elevated [CO2] increased fruit dry matter accumulationonly in no UV-B control plants because fruit dry weight wassimilar in the ambient or elevated CO2 treatments whenplants were grown under 77 or 151 kJ m-2 d-1 UV-B radi-ation The 77 kJ UV-B radiation did not affect leaf andstem dry weights but significantly decreased fruit dryweight compared to the control The 151 kJ UV-B radia-tion decreased dry matter accumulations of all plant partsand caused the greatest decrease in fruit weight amongplant organs
Leaf photosynthesis
Net photosynthetic rate (Pn) of the uppermost fullyexpanded mainstem leaves of control plants was stimu-
lated 44 by elevated [CO2] (Fig 2) Leaf Pn of plantsgrown under 77 kJ m-2 d-1 of UV-B radiation did not dif-fer from that of the control at either CO2 level at anytime of measurements Averaged across CO2 treatmentsleaf Pn of plants exposed to 151 kJ m-2 d-1 of UV-B radia-tion was decreased 33 at 28 DAE 47 at 44 DAE and42 at 64 DAE in comparison with the control plants Asignificant CO2 yen UV-B interactive effect was detected inleaf Pn at 44 and 64 DAE For instance in comparisonwith the control plants the 151 kJ UV-B-treated plantshad a 38 (44 DAE) or 33 (64 DAE) lower leaf Pn inambient [CO2] but 53 or 49 lower leaf Pn under ele-vated [CO2]
In general plants grown in elevated [CO2] had 11ndash19lower stomatal conductance than plants grown in ambient[CO2] (data not shown) The effect of UV-B radiation onstomatal conductance was similar to the trends observed inleaf Pn Intercellular [CO2] on the other hand did not differamong the UV-B treatments in ambient CO2 level with amean of about 260 mmol mol-1 (data not shown) but thevalues of intercellular [CO2] of 0 77 and 151 kJ UV-B-treated plants were 499 560 and 586 mmol mol-1 respec-tively under elevated CO2 conditions
Table 3 Effect of elevated CO2 and UV-B radiation on fruiting branch length (cm branch-1) the number of fruiting branches fruiting sites bolls floral buds and fruit abscission (no plant-1) 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Fruitingbranches
Branchlength
Fruitingsites Bolls
Floralbuds
Fruitabscission
360 00 141 plusmn 04 c 241 plusmn 15 b 473 plusmn 57 c 187 plusmn 24 ab 243 plusmn 36 c 43 plusmn 06 d77 158 plusmn 03 b 302 plusmn 08 a 558 plusmn 24 bc 147 plusmn 15 bc 341 plusmn 16 b 70 plusmn 08 c
151 142 plusmn 06 c 119 plusmn 13 d 339 plusmn 34 d 52 plusmn 11 d 214 plusmn 31 c 72 plusmn 19 c720 00 162 plusmn 05 b 312 plusmn 13 a 631 plusmn 69 ab 219 plusmn 29 a 344 plusmn 36 b 68 plusmn 09 cd
77 163 plusmn 03 b 310 plusmn 15 a 674 plusmn 30 a 144 plusmn 16 c 412 plusmn 19 a 118 plusmn 09 b151 176 plusmn 05 a 195 plusmn 14 c 514 plusmn 34 c 38 plusmn 12 d 328 plusmn 19 b 149 plusmn 10 a
P (CO2) lt 00001 lt 00001 lt 00001 06681 lt 00001 lt 00001P (UV-B) 00944 lt 00001 lt 00001 lt 00001 lt 00001 lt 00001P (CO2 yen UV-B) 00078 00240 06715 02686 06122 00156
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Table 4 Effect of elevated CO2 and UV-B radiation on above-ground dry matter accumulation (g plant-1) in cotton 14 and 24 d after emergence (DAE)
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
14 DAE 24 DAE
Leaves Stems Total Leaves Stems Total
360 00 050 plusmn 003 b 015 plusmn 001 c 065 plusmn 004 b 227 plusmn 024 b 140 plusmn 010 b 367 plusmn 035 b77 049 plusmn 002 bc 015 plusmn 001 c 064 plusmn 003 bc 190 plusmn 014 bc 111 plusmn 008 c 301 plusmn 022 c
151 040 plusmn 003 c 012 plusmn 001 c 052 plusmn 004 c 152 plusmn 010 c 082 plusmn 006 d 234 plusmn 015 d720 00 070 plusmn 003 a 023 plusmn 001 ab 093 plusmn 004 a 291 plusmn 026 a 192 plusmn 014 a 483 plusmn 039 a
77 065 plusmn 003 a 020 plusmn 001 b 085 plusmn 004 a 232 plusmn 025 b 142 plusmn 015 b 372 plusmn 040 b151 066 plusmn 004 a 024 plusmn 002 a 090 plusmn 006 a 230 plusmn 016 b 133 plusmn 007 bc 363 plusmn 023 b
P (CO2) lt 00001 lt 00001 lt 00001 00003 lt 00001 00002P (UV-B) 00789 02948 01427 00025 lt 00001 00023P (CO2 yen UV-B) 03110 00443 02874 06443 05071 05561
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

772
D Zhao
et al
copy 2003 Blackwell Publishing Ltd
Plant Cell and Environment
26
771ndash782
atmospheric [CO
2
] generally enhances leaf and canopy CO
2
assimilation rates because CO
2
is not only the substrate forphotosynthesis but also a competitive inhibitor of oxygen-ation of RuBP carboxylase and reduces photorespiration(Lawlor amp Mitchell 2000)
The response of crop plants to UV-B radiation in thepresence of other environmental stress factors is not clearand has not received much attention Studies conducted sofar indicate that effects of UV-B radiation with other envi-ronmental variables such as enhanced [CO
2
] (TeramuraSullivan amp Ziska 1990 Rozema
et al
1997 Visser
et al
1997) high temperature (Mark amp Tevini 1996) water defi-cit (Nogues
et al
1998 Alexieva
et al
2001) are both addi-tive and interactive Few studies have evaluated the effectsof UV-B radiation on the growth and development of cot-ton an important agronomic crop grown in a wide rangeof environmental conditions spanning latitudes 40
infin
N and40
infin
S although the influences of other environmental fac-tors such as atmospheric [CO
2
] temperature water andnutrients on plant growth and development have beenwell investigated in cotton (Christiansen 1986 GerikOosterhuis amp Torbet 1998 Reddy Hodges amp Kimball2000)
Reddy
et al
(2000) reported that canopy photosynthesisof cotton increased by 40 when [CO
2
] was increased from320 to 720
m
mol mol
-
1
for plants grown at 20 and 32
infin
C andincreased by about 80 when plants were grown underoptimum temperature (26ndash28
infin
C) conditions Kimball ampMauney (1993) found that cotton grown under550
m
mol CO
2
mol
-
1
had a 35 higher biomass 40 higherfruit weight and 60 higher lint yield than plants grownunder 350
m
mol CO
2
mol
-
1
under free-air CO
2
enrichmentstudies However there is a significant knowledge gap onthe interactive effects of UV-B radiation and [CO
2
] onplants We hypothesize that the CO
2
enhancement effectmay be reduced by concurrent exposure of plants to ele-vated UV-B radiation The objective of this study was thusto determine the interactive effects of elevated [CO
2
] andUV-B radiation on cotton growth development and phys-iology under controlled environment conditions Informa-tion related to whole plant and leaf-level processes will
contribute to filling gaps in our knowledge base andunderstanding
MATERIALS AND METHODS
Soilndashplantndashatmosphere research units
The experiment was conducted at the Agriculture and For-estry Experiment Station of Mississippi State (33
infin
28
cent
N88
infin
47
cent
W) Mississippi USA in 2001 using six soilndashplantndashatmosphere research (SPAR) units The SPAR facility hasthe capability to precisely control temperature and [CO
2
]at predetermined set points for plant growth studies undernear ambient levels of PAR Details of the SPAR units havebeen described by Reddy
et al
(2001) Each SPAR unitconsists of a steel soil bin (1 m deep by 2 m long by 05 mwide) and a Plexiglas chamber (25 m tall by 2 m long by15 m wide) to accommodate aerial plant parts a heatingand cooling system and an environment monitoring andcontrol system The Plexiglas chamber is completelyopaque to solar UV radiation but transmits more than 95of incoming PAR (wavelength 400ndash700 nm) (Table 1) Dur-ing the experiment the incoming solar radiation (285ndash2800 nm) outside the SPAR units was measured with apyranometer (Model 4ndash48 The Eppley Laboratory IncNewport RI USA) The average daily total solar radiationduring the experiment was 18
plusmn
49 MJ m
-
2
d
-
1
The air tem-perature and [CO
2
] in each SPAR unit were monitored andadjusted every 10 s throughout the experiment
The dew point temperatures were monitored with goldmirror hygrometers installed inside the return airline andwere also collected at 10 s intervals and summarized over900 s intervals The data were separated into day and nightperiods The daytime vapour pressure deficit (VPD) of eachtreatment was calculated based on air temperature and dewpoint temperature The PAR transmittance of all chamberswere determined on two typical clear days between 1000and 1400 h using seven 1 m line quantum sensors (Li-CorInc Lincoln NE USA) Daily mean temperatures aver-age daytime VPD and PAR transmittances of all treat-ments are presented in Table 1
Table 1
Measured [CO
2
] and Plexiglas transmittance of photosynthetically active radiation (PAR) from a typical day daily mean UV-B radiation dosage mean temperature (
T
) and daytime vapour pressure deficit (VPD) during the experimental period for each treatment
Treatment
CO
2
(
m
mol mol
-
1
)Mean UV-B(kJ m
-
2
d
-
1
)Mean T(
infin
C)VPD(kPa)
Transmittanceof PAR ()
CO
2
(
m
mol mol
-
1
)UV-B(kJ m
-
2
d
-
1
)
360 00 3597
plusmn
07 000
plusmn
000 2658
plusmn
004 168
plusmn
028 962
plusmn
0677 3613
plusmn
18 763
plusmn
005 2659
plusmn
003 196
plusmn
015 963
plusmn
05151 3601
plusmn
07 1503
plusmn
008 2645
plusmn
009 159
plusmn
008 973
plusmn
05720 00 7231
plusmn
10 000
plusmn
000 2659
plusmn
003 164
plusmn
020 986
plusmn
0277 7250
plusmn
14 774
plusmn
004 2659
plusmn
003 189
plusmn
021 957
plusmn
05151 7233
plusmn
10 1512
plusmn
009 2650
plusmn
005 179
plusmn
019 960
plusmn
05
Each value represents the mean plusmn SE of data of 40 values from a day for [CO2] and PAR transmittance or 65 d from 6 August to 10 October2001 for UV-B radiation VPD and temperature
Cotton responses to elevated CO2 and UV-B radiation 773
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Plant culture
Seeds of cotton cv NuCOTN 33B were sown on 1 August2001 in fine sand of the SPAR soil bins in 11 rows of fiveplants per row Emergence date was 5 d later The temper-atures in all units were maintained at 3022 infinC (daynight)during the experiment Plants were watered three times aday with half-strength Hoaglands nutrient solution deliv-ered at 0800 1200 and 1700 h to ensure favourable nutrientand water conditions for plant growth Irrigation was pro-vided through an automated and computer-controlled dripsystem Variable-density black shade cloths around theedges of plants were adjusted regularly to match plantheight in order to simulate natural shading in the presenceof other plants
Treatments
The six treatments included two CO2 levels of360 mmol mol-1 (ambient) and 720 mmol mol-1 (elevated)and three daily biologically effective UV-B radiation inten-sities of 0 (control) 77 and 151 kJ m-2 within each CO2
level The [CO2] and UV-B radiation treatments wereimposed from emergence The UV-B doses of 77 and151 kJ m-2 d-1 are designed to simulate the maximum levelof UV-B that would be received on a clear day on thesummer solstice with either ambient levels of ozone or witha 30 ozone depletion in Mississippis cotton productionregion as calculated by an empirical model (Green Crossamp Smith 1980)
Square-wave UV-B supplementation systems were usedto provide respective UV-B radiation in this study undernear ambient PAR The UV-B radiation was delivered toplants for 8 h each day from 0800 to 1600 h by eight fluo-rescent UV-313 lamps (Q-Panel Company Cleveland OHUSA) driven by 40 W dimming ballasts The lamps werewrapped with pre-solarized 008 mm cellulose diacetatefilm to filter UV-C (lt 280 nm) radiation The cellulose diac-etate film was changed at 3ndash4 d intervals The amount ofUV-B energy delivered at the top of the plant canopy waschecked daily at 1000 hours with a UVX digital radiometer(UVP Inc San Gabriel CA USA) and calibrated againstan Optronic Laboratory (Orlando FL USA) Model 754Spectroradiometer which was used to initially quantifylamp output The lamp power was adjusted as needed tomaintain the appropriate UV-B radiation level in eachtreatment and to maintain the distance from lamps to thetop of plants at about 05 m throughout the experiment Themean UV-B dosages measured at three different locationsin each SPAR unit corresponding to the permanent rowsfor all treatments during the experimental period are pre-sented in Table 1 After averaging the radiation dosages forthe plants within UV-B treatments the actual dosages forthe two levels of UV-B treatments were 77 and 151 kJ m-2
d-1 respectively Square-wave UV-B supplemental systemsin controlled environments usually provide higher UV-Bradiation and disproportionate spectral conditions com-pared to ambient condition (Musil et al 2002) and there is
no UV-B supplied in the control treatment This is likely toexaggerate the UV-B response in plants and thus cautionwill be necessary in extrapolating the findings from thisstudy to the field
Measurements
Plant height and the number of mainstem nodes weredetermined from nine plants (three centre plants in eachrow) at 3 d intervals from 10 to 66 d after emergence(DAE) These plants were also used to record dates ofappearance of the first floral bud of 3 mm in length (firstsquare FS) and first flower (FF) and durations in daysfrom emergence to FS and from FS to FF The plants wereharvested at the fourth true leaf stage (14 DAE) earlyfloral bud stage (29 DAE) and finally at 3 weeks after thefirst flower stage (66 DAE) Plants in the odd rows of eachSPAR unit were harvested at 14 DAE The five rows afterthe first sampling were spaced 33 cm apart The two evenrows were harvested 29 DAE and the remaining three rows(with a 67 cm row space) were finally harvested 66 DAEPlants were separated into leaves and stems at the first twoharvests and the leaf area and dry weights of leaves andstems were determined At the final harvest plant heightfruiting branch lengths numbers of mainstem nodes fruit-ing branches fruiting sites squares and bolls on each plantwere recorded The rooting medium was removed and rootswashed over a fine screen dried and weighed Mean inter-node length was calculated by dividing plant height by thenumber of mainstem nodes Fruit (squares + bolls) abscis-sion was calculated as the difference between the numberof fruiting sites and the number of fruits
Net photosynthetic rates (Pn) of the uppermost fullyexpanded mainstem leaf from five plants in each treatmentwere measured between 1000 and 1200 h using a LI-6400portable photosynthesis system (Li-Cor Inc) at 28 44 and64 DAE When measuring leaf Pn the PAR provided by a6400ndash02 LED light source was set to 1500 mmol m-2 s-1 Thetemperature in the leaf cuvette was set to 30 infinC and [CO2]matched CO2 treatments
Chlorophyll a chlorophyll b carotenoid and non-struc-tural carbohydrate (including glucose fructose sucroseand starch) concentrations of the leaves at the same posi-tion were also determined at 45 and 65 DAE For chloro-phyll measurement five leaf discs (0385 cm2 each) werepunched from a leaf and placed in a vial with 4 mL ofdimethyl sulphoxide Three samples were collected fromeach treatment and incubated at room temperature for24 h in the dark to allow for complete extraction of chlo-rophyll into the solution The absorbance of the extract wasmeasured using a Pharmacia UltraSpec Pro UVVIS spec-trophotometer (Pharmacia Cambridge UK) at 470 648and 664 nm and used to calculate chlorophyll a chlorophyllb and carotenoid concentrations (Chappelle Kim ampMcMurtrey 1992)
Five leaf discs from each leaf were collected at 0830 husing the same method described above and placed in a testtube with 2 mL of 80 (vv) cold ethanol Three leaf sam-
774 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
ples were obtained from each treatment and immediatelystored at -80 infinC to determine leaf non-structural carbohy-drates The modified method of Hendrix (1993) was usedto extract and quantify the concentrations of leaf non-structural carbohydrates Briefly soluble sugars wereextracted by incubating leaf tissue with 2 mL of the ethanolin an 80 infinC water bath for 15 min The extraction wasrepeated three times and the supernatants were combinedand brought to a 6 mL volume with 80 ethanol Sixtymilligrams of finely ground activated charcoal was addedto each sample tube covered and shaken by hand After5 min the tubes were centrifuged at 3000 yen g for 15 min toobtain a clear extract Four 40 mL aliquots from each sam-ple were pipetted into separate wells of a microtitrationplate and dried at 50 infinC for 30 min to remove the ethanolEight wells in the first column of each microtitration platewere filled with 20 mL glucose standard solutions of 0 5 1325 50 125 250 and 500 mg L-1 respectively To each samplewell 20 mL deionized water was added and then 100 mLglucose-6P dehydrogenaseiodonitrotetrazolium violet(INT) mixture (glucose kit 115 A Sigma Chemical Com-pany St Louis MO USA) was added under reduced roomillumination Plates were incubated at 37 infinC for 15 min andthe absorbance measured at 490 nm with an Emax preci-sion microplate reader (Emax Sunnyvale CA USA) Aglucose standard curve was obtained for each plate basedon the standard concentrations and absorbance Sampleglucose concentration was calculated according to the stan-dard curve absorbance extract volume and leaf disc areaSubsequently 10 mL phosphoglucose isomerase (PGIenzyme 025 units) and 10 mL invertase (83 units) wereadded to each well separately re-incubated at 37 infinC for15 min each time The absorbance was determined at490 nm to obtain concentrations of fructose and sucrose
After extraction of the soluble sugars 1 mL of 01 MKOH was added to the test tube containing the tissue res-idue and the tubes placed for 1 h in a boiling waterbathAn a-amylase and amyloglucosidase preparation was usedto hydrolyse starch to glucose as described by Hendrix(1993) After centrifuging the hydrolysate the supernatantwas collected and the glucose concentration determined asabove Starch concentration in the sample was calculatedaccording to the glucose concentration multiplied by 09 toaccount for water loss when glucose units are linked Thesum of glucose fructose sucrose and starch was defined astotal non-structural carbohydrate
Data analysis
A uniformity test of the SPAR units conducted in a previ-ous study by determining sorghum growth parametersunder the same controlled environments indicated no sta-tistical differences among all SPAR units (Reddy personalcommunication 2000) Therefore six treatments were ran-domly arranged in six identical SPAR units Except for thetwo treatment factors of [CO2] and UV-B radiation theother growth conditions were the same in all treatments(Table 1) All growth measurements were made on nine
plants and physiological measurements were made oneither three or five plants for each treatment Linear andsigmoid regressions were performed for the number ofmainstem nodes and plant height respectively against daysafter emergence Data were analysed by the analysis ofvariance (ANOVA) procedures in SAS (SAS Institute Inc1997) to determine the main and interactive effects of[CO2] and UV-B radiation on growth dry matter accumu-lation leaf Pn and concentrations of leaf pigments and non-structural carbohydrates If the hypothesis of equal meanswas rejected by the ANOVA test Fisher LSD procedures atP = 005 probability level (SAS Institute Inc 1997) wereemployed to distinguish differences among treatmentmeans for the growth and physiological processesmeasured
RESULTS
Plant growth and development
Plants grown under 720 mmol CO2 mol-1 were typically 25ndash50 cm taller than those grown in 360 mmol CO2 mol-1depending on UV-B exposure (Fig 1) Changes in plantheight from emergence up to 3 weeks after first floweringfollowed a sigmoid growth pattern The maximum main-stem elongation rate was achieved between 35 and 45 DAEfor all treatments but the elongation rates differed substan-tially among the treatments Plants irradiated with151 kJ m-2 d-1 UV-B radiation had significantly lower stemelongation rates in both CO2 levels compared to plants ineither the 0 or 77 kJ m-2 d-1 UV-B treatments At the finalharvest (66 DAE) plants grown in 720 mmol CO2 mol-1
were 22 taller than plants grown under360 mmol CO2 mol-1 averaged across the UV-B treatmentsPlants irradiated with UV-B radiation were significantlyshorter than the controls (except for 77 kJ UV-B treatmentat 360 mmol CO2 mol-1)
Unlike plant height mainstem nodes increased linearly(R2 = 099) as plants aged in all treatments (data notshown) The node appearance rates (slopes of the lines) didnot differ among the UV-B treatments but elevated [CO2]caused a slight increase in node addition rate (Y = 033X - 128 for 360 mmol CO2 mol-1 and Y = 036X - 161 for720 mmol CO2 mol-1 where Y is the number of mainstemnodes and X is days after emergence) Averaged across theUV-B treatments it took about 30 d (slope = 033) to pro-duce a leaf (or node) on the main stem under ambient[CO2] and about 28 d (slope = 036) under elevated [CO2]At the final harvest mean internode lengths of the 0 77and 151 kJ UV-B-treated plants were 85 79 and 45 cmrespectively under ambient [CO2] and 94 83 and 54 cmrespectively under elevated [CO2]
No CO2 yen UV-B radiation interaction effect was detectedfor leaf area development (Table 2) Plants grown in ele-vated [CO2] had significantly greater leaf area than plantsin ambient [CO2] at 14 DAE Leaf area was reduced by UV-B treatments of 77 and 151 kJ m-2 d-1 at 24 DAE At 66DAE only plants exposed to 151 kJ UV-B had significantly
Cotton responses to elevated CO2 and UV-B radiation 775
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
smaller leaf area (47ndash50) than plants in other treatmentsof CO2 and UV-B radiation
Neither elevated [CO2] nor UV-B radiation affected thedates of appearance of the first floral buds and flowers (datanot shown) A significant CO2 yen UV-B interactive effectwas detected for the number of fruiting branches branchlengths and fruit abscission (Table 3) Elevated [CO2] sig-nificantly increased the numbers of fruiting branches fruit-ing sites and squares per plant and mean branch length butdid not affect the number of bolls retained per plant Incontrast to [CO2] UV-B radiation significantly decreasedmost reproductive growth parameters except for the num-
ber of fruiting branches Fruit abscission in UV-B-treatedplants was significantly higher than in control plants in bothambient and elevated CO2 levels Floral bud abscission wasmore than young boll abscission when plants were grownunder UV-B radiation (data not shown)
Dry matter accumulation and partitioning
Elevated [CO2] significantly increased dry matter accumu-lation in leaves and stems at all sampling dates (Tables 4 amp5) Dry weights of leaves and stems did not differ amongthe UV-B radiation treatments at 14 DAE but at 24 DAE
Figure 1 Changes in plant height and stem elongation rate of cotton plants grown under 360 and 720 mmol CO2 mol-1 as affected by UV-B radiation Each data point represents the mean plusmn SE of nine plants The lines are regressions fitted to the data using a sigmoid equation for plant height and a first deriva-tive of the sigmoid equation for stem elon-gation rate
Table 2 Effects of elevated CO2 and UV-B radiation on leaf area (cm2 plant-1) of cotton plant 14 24 and 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Day after emergence
14 24 66
360 00 98 plusmn 6 ab 589 plusmn 48 b 6111 plusmn 627 c77 87 plusmn 3 bc 471 plusmn 50 c 6776 plusmn 331 bc
151 80 plusmn 6 c 455 plusmn 27 c 3064 plusmn 251 e720 00 102 plusmn 6 a 721 plusmn 32 a 8456 plusmn 432 a
77 104 plusmn 4 a 546 plusmn 42 bc 7738 plusmn 440 ab151 110 plusmn 7 a 588 plusmn 31 b 4502 plusmn 354 d
P (CO2) 00002 00011 lt 00001P (UV-B) 05991 00009 lt 00001P (CO2 yen UV-B) 00702 07166 02611
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
776 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
total dry weights of 77 and 151 kJ UV-B-treated plantswere 21 and 30 lower than the control plants respec-tively At the final harvest plants under 720 mmol CO2
mol-1 had a 36 higher total biomass than plants under360 mmol CO2 mol-1 averaged across the three UV-B treat-ments Total biomass produced was lowered by 12 for the77 kJ UV-B treatment and by 69 for the 151 kJ UV-Btreatment at 66 DAE compared to the control (Table 5)Elevated [CO2] increased fruit dry matter accumulationonly in no UV-B control plants because fruit dry weight wassimilar in the ambient or elevated CO2 treatments whenplants were grown under 77 or 151 kJ m-2 d-1 UV-B radi-ation The 77 kJ UV-B radiation did not affect leaf andstem dry weights but significantly decreased fruit dryweight compared to the control The 151 kJ UV-B radia-tion decreased dry matter accumulations of all plant partsand caused the greatest decrease in fruit weight amongplant organs
Leaf photosynthesis
Net photosynthetic rate (Pn) of the uppermost fullyexpanded mainstem leaves of control plants was stimu-
lated 44 by elevated [CO2] (Fig 2) Leaf Pn of plantsgrown under 77 kJ m-2 d-1 of UV-B radiation did not dif-fer from that of the control at either CO2 level at anytime of measurements Averaged across CO2 treatmentsleaf Pn of plants exposed to 151 kJ m-2 d-1 of UV-B radia-tion was decreased 33 at 28 DAE 47 at 44 DAE and42 at 64 DAE in comparison with the control plants Asignificant CO2 yen UV-B interactive effect was detected inleaf Pn at 44 and 64 DAE For instance in comparisonwith the control plants the 151 kJ UV-B-treated plantshad a 38 (44 DAE) or 33 (64 DAE) lower leaf Pn inambient [CO2] but 53 or 49 lower leaf Pn under ele-vated [CO2]
In general plants grown in elevated [CO2] had 11ndash19lower stomatal conductance than plants grown in ambient[CO2] (data not shown) The effect of UV-B radiation onstomatal conductance was similar to the trends observed inleaf Pn Intercellular [CO2] on the other hand did not differamong the UV-B treatments in ambient CO2 level with amean of about 260 mmol mol-1 (data not shown) but thevalues of intercellular [CO2] of 0 77 and 151 kJ UV-B-treated plants were 499 560 and 586 mmol mol-1 respec-tively under elevated CO2 conditions
Table 3 Effect of elevated CO2 and UV-B radiation on fruiting branch length (cm branch-1) the number of fruiting branches fruiting sites bolls floral buds and fruit abscission (no plant-1) 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Fruitingbranches
Branchlength
Fruitingsites Bolls
Floralbuds
Fruitabscission
360 00 141 plusmn 04 c 241 plusmn 15 b 473 plusmn 57 c 187 plusmn 24 ab 243 plusmn 36 c 43 plusmn 06 d77 158 plusmn 03 b 302 plusmn 08 a 558 plusmn 24 bc 147 plusmn 15 bc 341 plusmn 16 b 70 plusmn 08 c
151 142 plusmn 06 c 119 plusmn 13 d 339 plusmn 34 d 52 plusmn 11 d 214 plusmn 31 c 72 plusmn 19 c720 00 162 plusmn 05 b 312 plusmn 13 a 631 plusmn 69 ab 219 plusmn 29 a 344 plusmn 36 b 68 plusmn 09 cd
77 163 plusmn 03 b 310 plusmn 15 a 674 plusmn 30 a 144 plusmn 16 c 412 plusmn 19 a 118 plusmn 09 b151 176 plusmn 05 a 195 plusmn 14 c 514 plusmn 34 c 38 plusmn 12 d 328 plusmn 19 b 149 plusmn 10 a
P (CO2) lt 00001 lt 00001 lt 00001 06681 lt 00001 lt 00001P (UV-B) 00944 lt 00001 lt 00001 lt 00001 lt 00001 lt 00001P (CO2 yen UV-B) 00078 00240 06715 02686 06122 00156
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Table 4 Effect of elevated CO2 and UV-B radiation on above-ground dry matter accumulation (g plant-1) in cotton 14 and 24 d after emergence (DAE)
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
14 DAE 24 DAE
Leaves Stems Total Leaves Stems Total
360 00 050 plusmn 003 b 015 plusmn 001 c 065 plusmn 004 b 227 plusmn 024 b 140 plusmn 010 b 367 plusmn 035 b77 049 plusmn 002 bc 015 plusmn 001 c 064 plusmn 003 bc 190 plusmn 014 bc 111 plusmn 008 c 301 plusmn 022 c
151 040 plusmn 003 c 012 plusmn 001 c 052 plusmn 004 c 152 plusmn 010 c 082 plusmn 006 d 234 plusmn 015 d720 00 070 plusmn 003 a 023 plusmn 001 ab 093 plusmn 004 a 291 plusmn 026 a 192 plusmn 014 a 483 plusmn 039 a
77 065 plusmn 003 a 020 plusmn 001 b 085 plusmn 004 a 232 plusmn 025 b 142 plusmn 015 b 372 plusmn 040 b151 066 plusmn 004 a 024 plusmn 002 a 090 plusmn 006 a 230 plusmn 016 b 133 plusmn 007 bc 363 plusmn 023 b
P (CO2) lt 00001 lt 00001 lt 00001 00003 lt 00001 00002P (UV-B) 00789 02948 01427 00025 lt 00001 00023P (CO2 yen UV-B) 03110 00443 02874 06443 05071 05561
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

Cotton responses to elevated CO2 and UV-B radiation 773
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Plant culture
Seeds of cotton cv NuCOTN 33B were sown on 1 August2001 in fine sand of the SPAR soil bins in 11 rows of fiveplants per row Emergence date was 5 d later The temper-atures in all units were maintained at 3022 infinC (daynight)during the experiment Plants were watered three times aday with half-strength Hoaglands nutrient solution deliv-ered at 0800 1200 and 1700 h to ensure favourable nutrientand water conditions for plant growth Irrigation was pro-vided through an automated and computer-controlled dripsystem Variable-density black shade cloths around theedges of plants were adjusted regularly to match plantheight in order to simulate natural shading in the presenceof other plants
Treatments
The six treatments included two CO2 levels of360 mmol mol-1 (ambient) and 720 mmol mol-1 (elevated)and three daily biologically effective UV-B radiation inten-sities of 0 (control) 77 and 151 kJ m-2 within each CO2
level The [CO2] and UV-B radiation treatments wereimposed from emergence The UV-B doses of 77 and151 kJ m-2 d-1 are designed to simulate the maximum levelof UV-B that would be received on a clear day on thesummer solstice with either ambient levels of ozone or witha 30 ozone depletion in Mississippis cotton productionregion as calculated by an empirical model (Green Crossamp Smith 1980)
Square-wave UV-B supplementation systems were usedto provide respective UV-B radiation in this study undernear ambient PAR The UV-B radiation was delivered toplants for 8 h each day from 0800 to 1600 h by eight fluo-rescent UV-313 lamps (Q-Panel Company Cleveland OHUSA) driven by 40 W dimming ballasts The lamps werewrapped with pre-solarized 008 mm cellulose diacetatefilm to filter UV-C (lt 280 nm) radiation The cellulose diac-etate film was changed at 3ndash4 d intervals The amount ofUV-B energy delivered at the top of the plant canopy waschecked daily at 1000 hours with a UVX digital radiometer(UVP Inc San Gabriel CA USA) and calibrated againstan Optronic Laboratory (Orlando FL USA) Model 754Spectroradiometer which was used to initially quantifylamp output The lamp power was adjusted as needed tomaintain the appropriate UV-B radiation level in eachtreatment and to maintain the distance from lamps to thetop of plants at about 05 m throughout the experiment Themean UV-B dosages measured at three different locationsin each SPAR unit corresponding to the permanent rowsfor all treatments during the experimental period are pre-sented in Table 1 After averaging the radiation dosages forthe plants within UV-B treatments the actual dosages forthe two levels of UV-B treatments were 77 and 151 kJ m-2
d-1 respectively Square-wave UV-B supplemental systemsin controlled environments usually provide higher UV-Bradiation and disproportionate spectral conditions com-pared to ambient condition (Musil et al 2002) and there is
no UV-B supplied in the control treatment This is likely toexaggerate the UV-B response in plants and thus cautionwill be necessary in extrapolating the findings from thisstudy to the field
Measurements
Plant height and the number of mainstem nodes weredetermined from nine plants (three centre plants in eachrow) at 3 d intervals from 10 to 66 d after emergence(DAE) These plants were also used to record dates ofappearance of the first floral bud of 3 mm in length (firstsquare FS) and first flower (FF) and durations in daysfrom emergence to FS and from FS to FF The plants wereharvested at the fourth true leaf stage (14 DAE) earlyfloral bud stage (29 DAE) and finally at 3 weeks after thefirst flower stage (66 DAE) Plants in the odd rows of eachSPAR unit were harvested at 14 DAE The five rows afterthe first sampling were spaced 33 cm apart The two evenrows were harvested 29 DAE and the remaining three rows(with a 67 cm row space) were finally harvested 66 DAEPlants were separated into leaves and stems at the first twoharvests and the leaf area and dry weights of leaves andstems were determined At the final harvest plant heightfruiting branch lengths numbers of mainstem nodes fruit-ing branches fruiting sites squares and bolls on each plantwere recorded The rooting medium was removed and rootswashed over a fine screen dried and weighed Mean inter-node length was calculated by dividing plant height by thenumber of mainstem nodes Fruit (squares + bolls) abscis-sion was calculated as the difference between the numberof fruiting sites and the number of fruits
Net photosynthetic rates (Pn) of the uppermost fullyexpanded mainstem leaf from five plants in each treatmentwere measured between 1000 and 1200 h using a LI-6400portable photosynthesis system (Li-Cor Inc) at 28 44 and64 DAE When measuring leaf Pn the PAR provided by a6400ndash02 LED light source was set to 1500 mmol m-2 s-1 Thetemperature in the leaf cuvette was set to 30 infinC and [CO2]matched CO2 treatments
Chlorophyll a chlorophyll b carotenoid and non-struc-tural carbohydrate (including glucose fructose sucroseand starch) concentrations of the leaves at the same posi-tion were also determined at 45 and 65 DAE For chloro-phyll measurement five leaf discs (0385 cm2 each) werepunched from a leaf and placed in a vial with 4 mL ofdimethyl sulphoxide Three samples were collected fromeach treatment and incubated at room temperature for24 h in the dark to allow for complete extraction of chlo-rophyll into the solution The absorbance of the extract wasmeasured using a Pharmacia UltraSpec Pro UVVIS spec-trophotometer (Pharmacia Cambridge UK) at 470 648and 664 nm and used to calculate chlorophyll a chlorophyllb and carotenoid concentrations (Chappelle Kim ampMcMurtrey 1992)
Five leaf discs from each leaf were collected at 0830 husing the same method described above and placed in a testtube with 2 mL of 80 (vv) cold ethanol Three leaf sam-
774 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
ples were obtained from each treatment and immediatelystored at -80 infinC to determine leaf non-structural carbohy-drates The modified method of Hendrix (1993) was usedto extract and quantify the concentrations of leaf non-structural carbohydrates Briefly soluble sugars wereextracted by incubating leaf tissue with 2 mL of the ethanolin an 80 infinC water bath for 15 min The extraction wasrepeated three times and the supernatants were combinedand brought to a 6 mL volume with 80 ethanol Sixtymilligrams of finely ground activated charcoal was addedto each sample tube covered and shaken by hand After5 min the tubes were centrifuged at 3000 yen g for 15 min toobtain a clear extract Four 40 mL aliquots from each sam-ple were pipetted into separate wells of a microtitrationplate and dried at 50 infinC for 30 min to remove the ethanolEight wells in the first column of each microtitration platewere filled with 20 mL glucose standard solutions of 0 5 1325 50 125 250 and 500 mg L-1 respectively To each samplewell 20 mL deionized water was added and then 100 mLglucose-6P dehydrogenaseiodonitrotetrazolium violet(INT) mixture (glucose kit 115 A Sigma Chemical Com-pany St Louis MO USA) was added under reduced roomillumination Plates were incubated at 37 infinC for 15 min andthe absorbance measured at 490 nm with an Emax preci-sion microplate reader (Emax Sunnyvale CA USA) Aglucose standard curve was obtained for each plate basedon the standard concentrations and absorbance Sampleglucose concentration was calculated according to the stan-dard curve absorbance extract volume and leaf disc areaSubsequently 10 mL phosphoglucose isomerase (PGIenzyme 025 units) and 10 mL invertase (83 units) wereadded to each well separately re-incubated at 37 infinC for15 min each time The absorbance was determined at490 nm to obtain concentrations of fructose and sucrose
After extraction of the soluble sugars 1 mL of 01 MKOH was added to the test tube containing the tissue res-idue and the tubes placed for 1 h in a boiling waterbathAn a-amylase and amyloglucosidase preparation was usedto hydrolyse starch to glucose as described by Hendrix(1993) After centrifuging the hydrolysate the supernatantwas collected and the glucose concentration determined asabove Starch concentration in the sample was calculatedaccording to the glucose concentration multiplied by 09 toaccount for water loss when glucose units are linked Thesum of glucose fructose sucrose and starch was defined astotal non-structural carbohydrate
Data analysis
A uniformity test of the SPAR units conducted in a previ-ous study by determining sorghum growth parametersunder the same controlled environments indicated no sta-tistical differences among all SPAR units (Reddy personalcommunication 2000) Therefore six treatments were ran-domly arranged in six identical SPAR units Except for thetwo treatment factors of [CO2] and UV-B radiation theother growth conditions were the same in all treatments(Table 1) All growth measurements were made on nine
plants and physiological measurements were made oneither three or five plants for each treatment Linear andsigmoid regressions were performed for the number ofmainstem nodes and plant height respectively against daysafter emergence Data were analysed by the analysis ofvariance (ANOVA) procedures in SAS (SAS Institute Inc1997) to determine the main and interactive effects of[CO2] and UV-B radiation on growth dry matter accumu-lation leaf Pn and concentrations of leaf pigments and non-structural carbohydrates If the hypothesis of equal meanswas rejected by the ANOVA test Fisher LSD procedures atP = 005 probability level (SAS Institute Inc 1997) wereemployed to distinguish differences among treatmentmeans for the growth and physiological processesmeasured
RESULTS
Plant growth and development
Plants grown under 720 mmol CO2 mol-1 were typically 25ndash50 cm taller than those grown in 360 mmol CO2 mol-1depending on UV-B exposure (Fig 1) Changes in plantheight from emergence up to 3 weeks after first floweringfollowed a sigmoid growth pattern The maximum main-stem elongation rate was achieved between 35 and 45 DAEfor all treatments but the elongation rates differed substan-tially among the treatments Plants irradiated with151 kJ m-2 d-1 UV-B radiation had significantly lower stemelongation rates in both CO2 levels compared to plants ineither the 0 or 77 kJ m-2 d-1 UV-B treatments At the finalharvest (66 DAE) plants grown in 720 mmol CO2 mol-1
were 22 taller than plants grown under360 mmol CO2 mol-1 averaged across the UV-B treatmentsPlants irradiated with UV-B radiation were significantlyshorter than the controls (except for 77 kJ UV-B treatmentat 360 mmol CO2 mol-1)
Unlike plant height mainstem nodes increased linearly(R2 = 099) as plants aged in all treatments (data notshown) The node appearance rates (slopes of the lines) didnot differ among the UV-B treatments but elevated [CO2]caused a slight increase in node addition rate (Y = 033X - 128 for 360 mmol CO2 mol-1 and Y = 036X - 161 for720 mmol CO2 mol-1 where Y is the number of mainstemnodes and X is days after emergence) Averaged across theUV-B treatments it took about 30 d (slope = 033) to pro-duce a leaf (or node) on the main stem under ambient[CO2] and about 28 d (slope = 036) under elevated [CO2]At the final harvest mean internode lengths of the 0 77and 151 kJ UV-B-treated plants were 85 79 and 45 cmrespectively under ambient [CO2] and 94 83 and 54 cmrespectively under elevated [CO2]
No CO2 yen UV-B radiation interaction effect was detectedfor leaf area development (Table 2) Plants grown in ele-vated [CO2] had significantly greater leaf area than plantsin ambient [CO2] at 14 DAE Leaf area was reduced by UV-B treatments of 77 and 151 kJ m-2 d-1 at 24 DAE At 66DAE only plants exposed to 151 kJ UV-B had significantly
Cotton responses to elevated CO2 and UV-B radiation 775
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
smaller leaf area (47ndash50) than plants in other treatmentsof CO2 and UV-B radiation
Neither elevated [CO2] nor UV-B radiation affected thedates of appearance of the first floral buds and flowers (datanot shown) A significant CO2 yen UV-B interactive effectwas detected for the number of fruiting branches branchlengths and fruit abscission (Table 3) Elevated [CO2] sig-nificantly increased the numbers of fruiting branches fruit-ing sites and squares per plant and mean branch length butdid not affect the number of bolls retained per plant Incontrast to [CO2] UV-B radiation significantly decreasedmost reproductive growth parameters except for the num-
ber of fruiting branches Fruit abscission in UV-B-treatedplants was significantly higher than in control plants in bothambient and elevated CO2 levels Floral bud abscission wasmore than young boll abscission when plants were grownunder UV-B radiation (data not shown)
Dry matter accumulation and partitioning
Elevated [CO2] significantly increased dry matter accumu-lation in leaves and stems at all sampling dates (Tables 4 amp5) Dry weights of leaves and stems did not differ amongthe UV-B radiation treatments at 14 DAE but at 24 DAE
Figure 1 Changes in plant height and stem elongation rate of cotton plants grown under 360 and 720 mmol CO2 mol-1 as affected by UV-B radiation Each data point represents the mean plusmn SE of nine plants The lines are regressions fitted to the data using a sigmoid equation for plant height and a first deriva-tive of the sigmoid equation for stem elon-gation rate
Table 2 Effects of elevated CO2 and UV-B radiation on leaf area (cm2 plant-1) of cotton plant 14 24 and 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Day after emergence
14 24 66
360 00 98 plusmn 6 ab 589 plusmn 48 b 6111 plusmn 627 c77 87 plusmn 3 bc 471 plusmn 50 c 6776 plusmn 331 bc
151 80 plusmn 6 c 455 plusmn 27 c 3064 plusmn 251 e720 00 102 plusmn 6 a 721 plusmn 32 a 8456 plusmn 432 a
77 104 plusmn 4 a 546 plusmn 42 bc 7738 plusmn 440 ab151 110 plusmn 7 a 588 plusmn 31 b 4502 plusmn 354 d
P (CO2) 00002 00011 lt 00001P (UV-B) 05991 00009 lt 00001P (CO2 yen UV-B) 00702 07166 02611
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
776 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
total dry weights of 77 and 151 kJ UV-B-treated plantswere 21 and 30 lower than the control plants respec-tively At the final harvest plants under 720 mmol CO2
mol-1 had a 36 higher total biomass than plants under360 mmol CO2 mol-1 averaged across the three UV-B treat-ments Total biomass produced was lowered by 12 for the77 kJ UV-B treatment and by 69 for the 151 kJ UV-Btreatment at 66 DAE compared to the control (Table 5)Elevated [CO2] increased fruit dry matter accumulationonly in no UV-B control plants because fruit dry weight wassimilar in the ambient or elevated CO2 treatments whenplants were grown under 77 or 151 kJ m-2 d-1 UV-B radi-ation The 77 kJ UV-B radiation did not affect leaf andstem dry weights but significantly decreased fruit dryweight compared to the control The 151 kJ UV-B radia-tion decreased dry matter accumulations of all plant partsand caused the greatest decrease in fruit weight amongplant organs
Leaf photosynthesis
Net photosynthetic rate (Pn) of the uppermost fullyexpanded mainstem leaves of control plants was stimu-
lated 44 by elevated [CO2] (Fig 2) Leaf Pn of plantsgrown under 77 kJ m-2 d-1 of UV-B radiation did not dif-fer from that of the control at either CO2 level at anytime of measurements Averaged across CO2 treatmentsleaf Pn of plants exposed to 151 kJ m-2 d-1 of UV-B radia-tion was decreased 33 at 28 DAE 47 at 44 DAE and42 at 64 DAE in comparison with the control plants Asignificant CO2 yen UV-B interactive effect was detected inleaf Pn at 44 and 64 DAE For instance in comparisonwith the control plants the 151 kJ UV-B-treated plantshad a 38 (44 DAE) or 33 (64 DAE) lower leaf Pn inambient [CO2] but 53 or 49 lower leaf Pn under ele-vated [CO2]
In general plants grown in elevated [CO2] had 11ndash19lower stomatal conductance than plants grown in ambient[CO2] (data not shown) The effect of UV-B radiation onstomatal conductance was similar to the trends observed inleaf Pn Intercellular [CO2] on the other hand did not differamong the UV-B treatments in ambient CO2 level with amean of about 260 mmol mol-1 (data not shown) but thevalues of intercellular [CO2] of 0 77 and 151 kJ UV-B-treated plants were 499 560 and 586 mmol mol-1 respec-tively under elevated CO2 conditions
Table 3 Effect of elevated CO2 and UV-B radiation on fruiting branch length (cm branch-1) the number of fruiting branches fruiting sites bolls floral buds and fruit abscission (no plant-1) 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Fruitingbranches
Branchlength
Fruitingsites Bolls
Floralbuds
Fruitabscission
360 00 141 plusmn 04 c 241 plusmn 15 b 473 plusmn 57 c 187 plusmn 24 ab 243 plusmn 36 c 43 plusmn 06 d77 158 plusmn 03 b 302 plusmn 08 a 558 plusmn 24 bc 147 plusmn 15 bc 341 plusmn 16 b 70 plusmn 08 c
151 142 plusmn 06 c 119 plusmn 13 d 339 plusmn 34 d 52 plusmn 11 d 214 plusmn 31 c 72 plusmn 19 c720 00 162 plusmn 05 b 312 plusmn 13 a 631 plusmn 69 ab 219 plusmn 29 a 344 plusmn 36 b 68 plusmn 09 cd
77 163 plusmn 03 b 310 plusmn 15 a 674 plusmn 30 a 144 plusmn 16 c 412 plusmn 19 a 118 plusmn 09 b151 176 plusmn 05 a 195 plusmn 14 c 514 plusmn 34 c 38 plusmn 12 d 328 plusmn 19 b 149 plusmn 10 a
P (CO2) lt 00001 lt 00001 lt 00001 06681 lt 00001 lt 00001P (UV-B) 00944 lt 00001 lt 00001 lt 00001 lt 00001 lt 00001P (CO2 yen UV-B) 00078 00240 06715 02686 06122 00156
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Table 4 Effect of elevated CO2 and UV-B radiation on above-ground dry matter accumulation (g plant-1) in cotton 14 and 24 d after emergence (DAE)
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
14 DAE 24 DAE
Leaves Stems Total Leaves Stems Total
360 00 050 plusmn 003 b 015 plusmn 001 c 065 plusmn 004 b 227 plusmn 024 b 140 plusmn 010 b 367 plusmn 035 b77 049 plusmn 002 bc 015 plusmn 001 c 064 plusmn 003 bc 190 plusmn 014 bc 111 plusmn 008 c 301 plusmn 022 c
151 040 plusmn 003 c 012 plusmn 001 c 052 plusmn 004 c 152 plusmn 010 c 082 plusmn 006 d 234 plusmn 015 d720 00 070 plusmn 003 a 023 plusmn 001 ab 093 plusmn 004 a 291 plusmn 026 a 192 plusmn 014 a 483 plusmn 039 a
77 065 plusmn 003 a 020 plusmn 001 b 085 plusmn 004 a 232 plusmn 025 b 142 plusmn 015 b 372 plusmn 040 b151 066 plusmn 004 a 024 plusmn 002 a 090 plusmn 006 a 230 plusmn 016 b 133 plusmn 007 bc 363 plusmn 023 b
P (CO2) lt 00001 lt 00001 lt 00001 00003 lt 00001 00002P (UV-B) 00789 02948 01427 00025 lt 00001 00023P (CO2 yen UV-B) 03110 00443 02874 06443 05071 05561
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

774 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
ples were obtained from each treatment and immediatelystored at -80 infinC to determine leaf non-structural carbohy-drates The modified method of Hendrix (1993) was usedto extract and quantify the concentrations of leaf non-structural carbohydrates Briefly soluble sugars wereextracted by incubating leaf tissue with 2 mL of the ethanolin an 80 infinC water bath for 15 min The extraction wasrepeated three times and the supernatants were combinedand brought to a 6 mL volume with 80 ethanol Sixtymilligrams of finely ground activated charcoal was addedto each sample tube covered and shaken by hand After5 min the tubes were centrifuged at 3000 yen g for 15 min toobtain a clear extract Four 40 mL aliquots from each sam-ple were pipetted into separate wells of a microtitrationplate and dried at 50 infinC for 30 min to remove the ethanolEight wells in the first column of each microtitration platewere filled with 20 mL glucose standard solutions of 0 5 1325 50 125 250 and 500 mg L-1 respectively To each samplewell 20 mL deionized water was added and then 100 mLglucose-6P dehydrogenaseiodonitrotetrazolium violet(INT) mixture (glucose kit 115 A Sigma Chemical Com-pany St Louis MO USA) was added under reduced roomillumination Plates were incubated at 37 infinC for 15 min andthe absorbance measured at 490 nm with an Emax preci-sion microplate reader (Emax Sunnyvale CA USA) Aglucose standard curve was obtained for each plate basedon the standard concentrations and absorbance Sampleglucose concentration was calculated according to the stan-dard curve absorbance extract volume and leaf disc areaSubsequently 10 mL phosphoglucose isomerase (PGIenzyme 025 units) and 10 mL invertase (83 units) wereadded to each well separately re-incubated at 37 infinC for15 min each time The absorbance was determined at490 nm to obtain concentrations of fructose and sucrose
After extraction of the soluble sugars 1 mL of 01 MKOH was added to the test tube containing the tissue res-idue and the tubes placed for 1 h in a boiling waterbathAn a-amylase and amyloglucosidase preparation was usedto hydrolyse starch to glucose as described by Hendrix(1993) After centrifuging the hydrolysate the supernatantwas collected and the glucose concentration determined asabove Starch concentration in the sample was calculatedaccording to the glucose concentration multiplied by 09 toaccount for water loss when glucose units are linked Thesum of glucose fructose sucrose and starch was defined astotal non-structural carbohydrate
Data analysis
A uniformity test of the SPAR units conducted in a previ-ous study by determining sorghum growth parametersunder the same controlled environments indicated no sta-tistical differences among all SPAR units (Reddy personalcommunication 2000) Therefore six treatments were ran-domly arranged in six identical SPAR units Except for thetwo treatment factors of [CO2] and UV-B radiation theother growth conditions were the same in all treatments(Table 1) All growth measurements were made on nine
plants and physiological measurements were made oneither three or five plants for each treatment Linear andsigmoid regressions were performed for the number ofmainstem nodes and plant height respectively against daysafter emergence Data were analysed by the analysis ofvariance (ANOVA) procedures in SAS (SAS Institute Inc1997) to determine the main and interactive effects of[CO2] and UV-B radiation on growth dry matter accumu-lation leaf Pn and concentrations of leaf pigments and non-structural carbohydrates If the hypothesis of equal meanswas rejected by the ANOVA test Fisher LSD procedures atP = 005 probability level (SAS Institute Inc 1997) wereemployed to distinguish differences among treatmentmeans for the growth and physiological processesmeasured
RESULTS
Plant growth and development
Plants grown under 720 mmol CO2 mol-1 were typically 25ndash50 cm taller than those grown in 360 mmol CO2 mol-1depending on UV-B exposure (Fig 1) Changes in plantheight from emergence up to 3 weeks after first floweringfollowed a sigmoid growth pattern The maximum main-stem elongation rate was achieved between 35 and 45 DAEfor all treatments but the elongation rates differed substan-tially among the treatments Plants irradiated with151 kJ m-2 d-1 UV-B radiation had significantly lower stemelongation rates in both CO2 levels compared to plants ineither the 0 or 77 kJ m-2 d-1 UV-B treatments At the finalharvest (66 DAE) plants grown in 720 mmol CO2 mol-1
were 22 taller than plants grown under360 mmol CO2 mol-1 averaged across the UV-B treatmentsPlants irradiated with UV-B radiation were significantlyshorter than the controls (except for 77 kJ UV-B treatmentat 360 mmol CO2 mol-1)
Unlike plant height mainstem nodes increased linearly(R2 = 099) as plants aged in all treatments (data notshown) The node appearance rates (slopes of the lines) didnot differ among the UV-B treatments but elevated [CO2]caused a slight increase in node addition rate (Y = 033X - 128 for 360 mmol CO2 mol-1 and Y = 036X - 161 for720 mmol CO2 mol-1 where Y is the number of mainstemnodes and X is days after emergence) Averaged across theUV-B treatments it took about 30 d (slope = 033) to pro-duce a leaf (or node) on the main stem under ambient[CO2] and about 28 d (slope = 036) under elevated [CO2]At the final harvest mean internode lengths of the 0 77and 151 kJ UV-B-treated plants were 85 79 and 45 cmrespectively under ambient [CO2] and 94 83 and 54 cmrespectively under elevated [CO2]
No CO2 yen UV-B radiation interaction effect was detectedfor leaf area development (Table 2) Plants grown in ele-vated [CO2] had significantly greater leaf area than plantsin ambient [CO2] at 14 DAE Leaf area was reduced by UV-B treatments of 77 and 151 kJ m-2 d-1 at 24 DAE At 66DAE only plants exposed to 151 kJ UV-B had significantly
Cotton responses to elevated CO2 and UV-B radiation 775
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
smaller leaf area (47ndash50) than plants in other treatmentsof CO2 and UV-B radiation
Neither elevated [CO2] nor UV-B radiation affected thedates of appearance of the first floral buds and flowers (datanot shown) A significant CO2 yen UV-B interactive effectwas detected for the number of fruiting branches branchlengths and fruit abscission (Table 3) Elevated [CO2] sig-nificantly increased the numbers of fruiting branches fruit-ing sites and squares per plant and mean branch length butdid not affect the number of bolls retained per plant Incontrast to [CO2] UV-B radiation significantly decreasedmost reproductive growth parameters except for the num-
ber of fruiting branches Fruit abscission in UV-B-treatedplants was significantly higher than in control plants in bothambient and elevated CO2 levels Floral bud abscission wasmore than young boll abscission when plants were grownunder UV-B radiation (data not shown)
Dry matter accumulation and partitioning
Elevated [CO2] significantly increased dry matter accumu-lation in leaves and stems at all sampling dates (Tables 4 amp5) Dry weights of leaves and stems did not differ amongthe UV-B radiation treatments at 14 DAE but at 24 DAE
Figure 1 Changes in plant height and stem elongation rate of cotton plants grown under 360 and 720 mmol CO2 mol-1 as affected by UV-B radiation Each data point represents the mean plusmn SE of nine plants The lines are regressions fitted to the data using a sigmoid equation for plant height and a first deriva-tive of the sigmoid equation for stem elon-gation rate
Table 2 Effects of elevated CO2 and UV-B radiation on leaf area (cm2 plant-1) of cotton plant 14 24 and 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Day after emergence
14 24 66
360 00 98 plusmn 6 ab 589 plusmn 48 b 6111 plusmn 627 c77 87 plusmn 3 bc 471 plusmn 50 c 6776 plusmn 331 bc
151 80 plusmn 6 c 455 plusmn 27 c 3064 plusmn 251 e720 00 102 plusmn 6 a 721 plusmn 32 a 8456 plusmn 432 a
77 104 plusmn 4 a 546 plusmn 42 bc 7738 plusmn 440 ab151 110 plusmn 7 a 588 plusmn 31 b 4502 plusmn 354 d
P (CO2) 00002 00011 lt 00001P (UV-B) 05991 00009 lt 00001P (CO2 yen UV-B) 00702 07166 02611
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
776 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
total dry weights of 77 and 151 kJ UV-B-treated plantswere 21 and 30 lower than the control plants respec-tively At the final harvest plants under 720 mmol CO2
mol-1 had a 36 higher total biomass than plants under360 mmol CO2 mol-1 averaged across the three UV-B treat-ments Total biomass produced was lowered by 12 for the77 kJ UV-B treatment and by 69 for the 151 kJ UV-Btreatment at 66 DAE compared to the control (Table 5)Elevated [CO2] increased fruit dry matter accumulationonly in no UV-B control plants because fruit dry weight wassimilar in the ambient or elevated CO2 treatments whenplants were grown under 77 or 151 kJ m-2 d-1 UV-B radi-ation The 77 kJ UV-B radiation did not affect leaf andstem dry weights but significantly decreased fruit dryweight compared to the control The 151 kJ UV-B radia-tion decreased dry matter accumulations of all plant partsand caused the greatest decrease in fruit weight amongplant organs
Leaf photosynthesis
Net photosynthetic rate (Pn) of the uppermost fullyexpanded mainstem leaves of control plants was stimu-
lated 44 by elevated [CO2] (Fig 2) Leaf Pn of plantsgrown under 77 kJ m-2 d-1 of UV-B radiation did not dif-fer from that of the control at either CO2 level at anytime of measurements Averaged across CO2 treatmentsleaf Pn of plants exposed to 151 kJ m-2 d-1 of UV-B radia-tion was decreased 33 at 28 DAE 47 at 44 DAE and42 at 64 DAE in comparison with the control plants Asignificant CO2 yen UV-B interactive effect was detected inleaf Pn at 44 and 64 DAE For instance in comparisonwith the control plants the 151 kJ UV-B-treated plantshad a 38 (44 DAE) or 33 (64 DAE) lower leaf Pn inambient [CO2] but 53 or 49 lower leaf Pn under ele-vated [CO2]
In general plants grown in elevated [CO2] had 11ndash19lower stomatal conductance than plants grown in ambient[CO2] (data not shown) The effect of UV-B radiation onstomatal conductance was similar to the trends observed inleaf Pn Intercellular [CO2] on the other hand did not differamong the UV-B treatments in ambient CO2 level with amean of about 260 mmol mol-1 (data not shown) but thevalues of intercellular [CO2] of 0 77 and 151 kJ UV-B-treated plants were 499 560 and 586 mmol mol-1 respec-tively under elevated CO2 conditions
Table 3 Effect of elevated CO2 and UV-B radiation on fruiting branch length (cm branch-1) the number of fruiting branches fruiting sites bolls floral buds and fruit abscission (no plant-1) 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Fruitingbranches
Branchlength
Fruitingsites Bolls
Floralbuds
Fruitabscission
360 00 141 plusmn 04 c 241 plusmn 15 b 473 plusmn 57 c 187 plusmn 24 ab 243 plusmn 36 c 43 plusmn 06 d77 158 plusmn 03 b 302 plusmn 08 a 558 plusmn 24 bc 147 plusmn 15 bc 341 plusmn 16 b 70 plusmn 08 c
151 142 plusmn 06 c 119 plusmn 13 d 339 plusmn 34 d 52 plusmn 11 d 214 plusmn 31 c 72 plusmn 19 c720 00 162 plusmn 05 b 312 plusmn 13 a 631 plusmn 69 ab 219 plusmn 29 a 344 plusmn 36 b 68 plusmn 09 cd
77 163 plusmn 03 b 310 plusmn 15 a 674 plusmn 30 a 144 plusmn 16 c 412 plusmn 19 a 118 plusmn 09 b151 176 plusmn 05 a 195 plusmn 14 c 514 plusmn 34 c 38 plusmn 12 d 328 plusmn 19 b 149 plusmn 10 a
P (CO2) lt 00001 lt 00001 lt 00001 06681 lt 00001 lt 00001P (UV-B) 00944 lt 00001 lt 00001 lt 00001 lt 00001 lt 00001P (CO2 yen UV-B) 00078 00240 06715 02686 06122 00156
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Table 4 Effect of elevated CO2 and UV-B radiation on above-ground dry matter accumulation (g plant-1) in cotton 14 and 24 d after emergence (DAE)
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
14 DAE 24 DAE
Leaves Stems Total Leaves Stems Total
360 00 050 plusmn 003 b 015 plusmn 001 c 065 plusmn 004 b 227 plusmn 024 b 140 plusmn 010 b 367 plusmn 035 b77 049 plusmn 002 bc 015 plusmn 001 c 064 plusmn 003 bc 190 plusmn 014 bc 111 plusmn 008 c 301 plusmn 022 c
151 040 plusmn 003 c 012 plusmn 001 c 052 plusmn 004 c 152 plusmn 010 c 082 plusmn 006 d 234 plusmn 015 d720 00 070 plusmn 003 a 023 plusmn 001 ab 093 plusmn 004 a 291 plusmn 026 a 192 plusmn 014 a 483 plusmn 039 a
77 065 plusmn 003 a 020 plusmn 001 b 085 plusmn 004 a 232 plusmn 025 b 142 plusmn 015 b 372 plusmn 040 b151 066 plusmn 004 a 024 plusmn 002 a 090 plusmn 006 a 230 plusmn 016 b 133 plusmn 007 bc 363 plusmn 023 b
P (CO2) lt 00001 lt 00001 lt 00001 00003 lt 00001 00002P (UV-B) 00789 02948 01427 00025 lt 00001 00023P (CO2 yen UV-B) 03110 00443 02874 06443 05071 05561
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

Cotton responses to elevated CO2 and UV-B radiation 775
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
smaller leaf area (47ndash50) than plants in other treatmentsof CO2 and UV-B radiation
Neither elevated [CO2] nor UV-B radiation affected thedates of appearance of the first floral buds and flowers (datanot shown) A significant CO2 yen UV-B interactive effectwas detected for the number of fruiting branches branchlengths and fruit abscission (Table 3) Elevated [CO2] sig-nificantly increased the numbers of fruiting branches fruit-ing sites and squares per plant and mean branch length butdid not affect the number of bolls retained per plant Incontrast to [CO2] UV-B radiation significantly decreasedmost reproductive growth parameters except for the num-
ber of fruiting branches Fruit abscission in UV-B-treatedplants was significantly higher than in control plants in bothambient and elevated CO2 levels Floral bud abscission wasmore than young boll abscission when plants were grownunder UV-B radiation (data not shown)
Dry matter accumulation and partitioning
Elevated [CO2] significantly increased dry matter accumu-lation in leaves and stems at all sampling dates (Tables 4 amp5) Dry weights of leaves and stems did not differ amongthe UV-B radiation treatments at 14 DAE but at 24 DAE
Figure 1 Changes in plant height and stem elongation rate of cotton plants grown under 360 and 720 mmol CO2 mol-1 as affected by UV-B radiation Each data point represents the mean plusmn SE of nine plants The lines are regressions fitted to the data using a sigmoid equation for plant height and a first deriva-tive of the sigmoid equation for stem elon-gation rate
Table 2 Effects of elevated CO2 and UV-B radiation on leaf area (cm2 plant-1) of cotton plant 14 24 and 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Day after emergence
14 24 66
360 00 98 plusmn 6 ab 589 plusmn 48 b 6111 plusmn 627 c77 87 plusmn 3 bc 471 plusmn 50 c 6776 plusmn 331 bc
151 80 plusmn 6 c 455 plusmn 27 c 3064 plusmn 251 e720 00 102 plusmn 6 a 721 plusmn 32 a 8456 plusmn 432 a
77 104 plusmn 4 a 546 plusmn 42 bc 7738 plusmn 440 ab151 110 plusmn 7 a 588 plusmn 31 b 4502 plusmn 354 d
P (CO2) 00002 00011 lt 00001P (UV-B) 05991 00009 lt 00001P (CO2 yen UV-B) 00702 07166 02611
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
776 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
total dry weights of 77 and 151 kJ UV-B-treated plantswere 21 and 30 lower than the control plants respec-tively At the final harvest plants under 720 mmol CO2
mol-1 had a 36 higher total biomass than plants under360 mmol CO2 mol-1 averaged across the three UV-B treat-ments Total biomass produced was lowered by 12 for the77 kJ UV-B treatment and by 69 for the 151 kJ UV-Btreatment at 66 DAE compared to the control (Table 5)Elevated [CO2] increased fruit dry matter accumulationonly in no UV-B control plants because fruit dry weight wassimilar in the ambient or elevated CO2 treatments whenplants were grown under 77 or 151 kJ m-2 d-1 UV-B radi-ation The 77 kJ UV-B radiation did not affect leaf andstem dry weights but significantly decreased fruit dryweight compared to the control The 151 kJ UV-B radia-tion decreased dry matter accumulations of all plant partsand caused the greatest decrease in fruit weight amongplant organs
Leaf photosynthesis
Net photosynthetic rate (Pn) of the uppermost fullyexpanded mainstem leaves of control plants was stimu-
lated 44 by elevated [CO2] (Fig 2) Leaf Pn of plantsgrown under 77 kJ m-2 d-1 of UV-B radiation did not dif-fer from that of the control at either CO2 level at anytime of measurements Averaged across CO2 treatmentsleaf Pn of plants exposed to 151 kJ m-2 d-1 of UV-B radia-tion was decreased 33 at 28 DAE 47 at 44 DAE and42 at 64 DAE in comparison with the control plants Asignificant CO2 yen UV-B interactive effect was detected inleaf Pn at 44 and 64 DAE For instance in comparisonwith the control plants the 151 kJ UV-B-treated plantshad a 38 (44 DAE) or 33 (64 DAE) lower leaf Pn inambient [CO2] but 53 or 49 lower leaf Pn under ele-vated [CO2]
In general plants grown in elevated [CO2] had 11ndash19lower stomatal conductance than plants grown in ambient[CO2] (data not shown) The effect of UV-B radiation onstomatal conductance was similar to the trends observed inleaf Pn Intercellular [CO2] on the other hand did not differamong the UV-B treatments in ambient CO2 level with amean of about 260 mmol mol-1 (data not shown) but thevalues of intercellular [CO2] of 0 77 and 151 kJ UV-B-treated plants were 499 560 and 586 mmol mol-1 respec-tively under elevated CO2 conditions
Table 3 Effect of elevated CO2 and UV-B radiation on fruiting branch length (cm branch-1) the number of fruiting branches fruiting sites bolls floral buds and fruit abscission (no plant-1) 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Fruitingbranches
Branchlength
Fruitingsites Bolls
Floralbuds
Fruitabscission
360 00 141 plusmn 04 c 241 plusmn 15 b 473 plusmn 57 c 187 plusmn 24 ab 243 plusmn 36 c 43 plusmn 06 d77 158 plusmn 03 b 302 plusmn 08 a 558 plusmn 24 bc 147 plusmn 15 bc 341 plusmn 16 b 70 plusmn 08 c
151 142 plusmn 06 c 119 plusmn 13 d 339 plusmn 34 d 52 plusmn 11 d 214 plusmn 31 c 72 plusmn 19 c720 00 162 plusmn 05 b 312 plusmn 13 a 631 plusmn 69 ab 219 plusmn 29 a 344 plusmn 36 b 68 plusmn 09 cd
77 163 plusmn 03 b 310 plusmn 15 a 674 plusmn 30 a 144 plusmn 16 c 412 plusmn 19 a 118 plusmn 09 b151 176 plusmn 05 a 195 plusmn 14 c 514 plusmn 34 c 38 plusmn 12 d 328 plusmn 19 b 149 plusmn 10 a
P (CO2) lt 00001 lt 00001 lt 00001 06681 lt 00001 lt 00001P (UV-B) 00944 lt 00001 lt 00001 lt 00001 lt 00001 lt 00001P (CO2 yen UV-B) 00078 00240 06715 02686 06122 00156
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Table 4 Effect of elevated CO2 and UV-B radiation on above-ground dry matter accumulation (g plant-1) in cotton 14 and 24 d after emergence (DAE)
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
14 DAE 24 DAE
Leaves Stems Total Leaves Stems Total
360 00 050 plusmn 003 b 015 plusmn 001 c 065 plusmn 004 b 227 plusmn 024 b 140 plusmn 010 b 367 plusmn 035 b77 049 plusmn 002 bc 015 plusmn 001 c 064 plusmn 003 bc 190 plusmn 014 bc 111 plusmn 008 c 301 plusmn 022 c
151 040 plusmn 003 c 012 plusmn 001 c 052 plusmn 004 c 152 plusmn 010 c 082 plusmn 006 d 234 plusmn 015 d720 00 070 plusmn 003 a 023 plusmn 001 ab 093 plusmn 004 a 291 plusmn 026 a 192 plusmn 014 a 483 plusmn 039 a
77 065 plusmn 003 a 020 plusmn 001 b 085 plusmn 004 a 232 plusmn 025 b 142 plusmn 015 b 372 plusmn 040 b151 066 plusmn 004 a 024 plusmn 002 a 090 plusmn 006 a 230 plusmn 016 b 133 plusmn 007 bc 363 plusmn 023 b
P (CO2) lt 00001 lt 00001 lt 00001 00003 lt 00001 00002P (UV-B) 00789 02948 01427 00025 lt 00001 00023P (CO2 yen UV-B) 03110 00443 02874 06443 05071 05561
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

776 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
total dry weights of 77 and 151 kJ UV-B-treated plantswere 21 and 30 lower than the control plants respec-tively At the final harvest plants under 720 mmol CO2
mol-1 had a 36 higher total biomass than plants under360 mmol CO2 mol-1 averaged across the three UV-B treat-ments Total biomass produced was lowered by 12 for the77 kJ UV-B treatment and by 69 for the 151 kJ UV-Btreatment at 66 DAE compared to the control (Table 5)Elevated [CO2] increased fruit dry matter accumulationonly in no UV-B control plants because fruit dry weight wassimilar in the ambient or elevated CO2 treatments whenplants were grown under 77 or 151 kJ m-2 d-1 UV-B radi-ation The 77 kJ UV-B radiation did not affect leaf andstem dry weights but significantly decreased fruit dryweight compared to the control The 151 kJ UV-B radia-tion decreased dry matter accumulations of all plant partsand caused the greatest decrease in fruit weight amongplant organs
Leaf photosynthesis
Net photosynthetic rate (Pn) of the uppermost fullyexpanded mainstem leaves of control plants was stimu-
lated 44 by elevated [CO2] (Fig 2) Leaf Pn of plantsgrown under 77 kJ m-2 d-1 of UV-B radiation did not dif-fer from that of the control at either CO2 level at anytime of measurements Averaged across CO2 treatmentsleaf Pn of plants exposed to 151 kJ m-2 d-1 of UV-B radia-tion was decreased 33 at 28 DAE 47 at 44 DAE and42 at 64 DAE in comparison with the control plants Asignificant CO2 yen UV-B interactive effect was detected inleaf Pn at 44 and 64 DAE For instance in comparisonwith the control plants the 151 kJ UV-B-treated plantshad a 38 (44 DAE) or 33 (64 DAE) lower leaf Pn inambient [CO2] but 53 or 49 lower leaf Pn under ele-vated [CO2]
In general plants grown in elevated [CO2] had 11ndash19lower stomatal conductance than plants grown in ambient[CO2] (data not shown) The effect of UV-B radiation onstomatal conductance was similar to the trends observed inleaf Pn Intercellular [CO2] on the other hand did not differamong the UV-B treatments in ambient CO2 level with amean of about 260 mmol mol-1 (data not shown) but thevalues of intercellular [CO2] of 0 77 and 151 kJ UV-B-treated plants were 499 560 and 586 mmol mol-1 respec-tively under elevated CO2 conditions
Table 3 Effect of elevated CO2 and UV-B radiation on fruiting branch length (cm branch-1) the number of fruiting branches fruiting sites bolls floral buds and fruit abscission (no plant-1) 66 d after emergence
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Fruitingbranches
Branchlength
Fruitingsites Bolls
Floralbuds
Fruitabscission
360 00 141 plusmn 04 c 241 plusmn 15 b 473 plusmn 57 c 187 plusmn 24 ab 243 plusmn 36 c 43 plusmn 06 d77 158 plusmn 03 b 302 plusmn 08 a 558 plusmn 24 bc 147 plusmn 15 bc 341 plusmn 16 b 70 plusmn 08 c
151 142 plusmn 06 c 119 plusmn 13 d 339 plusmn 34 d 52 plusmn 11 d 214 plusmn 31 c 72 plusmn 19 c720 00 162 plusmn 05 b 312 plusmn 13 a 631 plusmn 69 ab 219 plusmn 29 a 344 plusmn 36 b 68 plusmn 09 cd
77 163 plusmn 03 b 310 plusmn 15 a 674 plusmn 30 a 144 plusmn 16 c 412 plusmn 19 a 118 plusmn 09 b151 176 plusmn 05 a 195 plusmn 14 c 514 plusmn 34 c 38 plusmn 12 d 328 plusmn 19 b 149 plusmn 10 a
P (CO2) lt 00001 lt 00001 lt 00001 06681 lt 00001 lt 00001P (UV-B) 00944 lt 00001 lt 00001 lt 00001 lt 00001 lt 00001P (CO2 yen UV-B) 00078 00240 06715 02686 06122 00156
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Table 4 Effect of elevated CO2 and UV-B radiation on above-ground dry matter accumulation (g plant-1) in cotton 14 and 24 d after emergence (DAE)
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
14 DAE 24 DAE
Leaves Stems Total Leaves Stems Total
360 00 050 plusmn 003 b 015 plusmn 001 c 065 plusmn 004 b 227 plusmn 024 b 140 plusmn 010 b 367 plusmn 035 b77 049 plusmn 002 bc 015 plusmn 001 c 064 plusmn 003 bc 190 plusmn 014 bc 111 plusmn 008 c 301 plusmn 022 c
151 040 plusmn 003 c 012 plusmn 001 c 052 plusmn 004 c 152 plusmn 010 c 082 plusmn 006 d 234 plusmn 015 d720 00 070 plusmn 003 a 023 plusmn 001 ab 093 plusmn 004 a 291 plusmn 026 a 192 plusmn 014 a 483 plusmn 039 a
77 065 plusmn 003 a 020 plusmn 001 b 085 plusmn 004 a 232 plusmn 025 b 142 plusmn 015 b 372 plusmn 040 b151 066 plusmn 004 a 024 plusmn 002 a 090 plusmn 006 a 230 plusmn 016 b 133 plusmn 007 bc 363 plusmn 023 b
P (CO2) lt 00001 lt 00001 lt 00001 00003 lt 00001 00002P (UV-B) 00789 02948 01427 00025 lt 00001 00023P (CO2 yen UV-B) 03110 00443 02874 06443 05071 05561
Each value is the mean plusmn SE of nine plantsMeans followed by the same letter within a column are not significantly different (P gt 005)
Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

Cotton responses to elevated CO2 and UV-B radiation 777
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Leaf pigments
Elevated [CO2] did not affect leaf total chlorophyll caro-tenoid concentrations or the chlorophyll a b ratio (Fig 3)There were no interactive effects on the pigment compo-nents between CO2 and UV-B radiation treatments Totalchlorophyll concentration decreased and chlorophyll a bratio increased with the increase in UV-B radiation Aver-aged across the two CO2 levels chlorophyll concentrationsof 0 77 and 151 kJ UV-B-treated plants were 402 368 and326 mg m-2 respectively chlorophyll a b ratios were 409438 and 447 respectively Carotenoid concentration didnot differ between the control and 77 kJ UV-B-treatedplants but was 16 lower in plants exposed to 151 kJm-2 d-1 UV-B radiation
Leaf non-structural carbohydrates
Because leaf non-structural carbohydrate concentrationsdid not differ statistically between the two measurementdates the data were averaged across sampling dates(Fig 4) Elevated [CO2] significantly increased leaf glucose(P lt 005) sucrose (P lt 001) and starch (P lt 00001) con-centrations but did not affect fructose concentration As aresult plants grown in elevated [CO2] had 118 highertotal non-structural carbohydrates 28 higher glucose andsucrose and 191 more starch than plants grown in ambi-ent [CO2] No statistical differences in the concentrationsof non-structural carbohydrates were detected among theUV-B treatments at ambient [CO2] Under elevated CO2
condition however leaf sucrose and starch decreased asUV-B radiation increased the 77 kJ m-2 d-1 UV-B radia-tion did not affect leaf glucose and fructose but the151 kJ m-2 d-1 UV-B radiation decreased the concentrationof both sugars compared to the controls or 77 kJ UV-B-treated plants
DISCUSSION
Effects of elevated CO2
The results of elevated [CO2] stimulating growth and drymatter accumulation of cotton plants grown under no UV-B and 77 kJ m-2 d-1 UV-B conditions are similar to earlierreports (Mauney et al 1994 Reddy et al 2000) Elevated[CO2] affected neither cotton leaf chlorophyll concentra-tions nor chlorophyll a b ratio in the present study (Fig 3)This is in agreement with the findings in wheat by Monjeamp Bugbee (1998) but inconsistent with Radoglou amp Jarvis(1992) who reported elevated CO2 decreased leaf chloro-phyll concentration of Phaseolus vulgaris Numerous stud-ies have shown that elevated [CO2] increases Pn in C3 plantsbecause higher [CO2] can suppress RuBP oxygenase activ-ity decrease photorespiration and increase carbon assimi-lates for plant growth and development (Lawlor amp Mitchell2000) Our results also indicate that elevated [CO2] signifi-
Table 5 Effect of elevated CO2 and UV-B radiation on dry matter accumulation (g plant-1) of leaves fruits (bolls + flowers + floral buds) stems and roots at the final harvest 66 d after emergence Each value is the mean plusmn SE of nine plants
CO2
(mmol mol-1)UV-B(kJ m-2 d-1)
Plant parts
Leaves Fruits Stems Roots Total
360 00 387 plusmn 40 b 203 plusmn 23 b 507 plusmn 47 c 98 1196 plusmn 103 c77 364 plusmn 18 b 164 plusmn 15 c 520 plusmn 24 c 82 1130 plusmn 51 c
151 162 plusmn 13 c 30 plusmn 09 d 147 plusmn 18 e 32 371 plusmn 39 d720 00 540 plusmn 28 a 261 plusmn 20 a 789 plusmn 32 a 121 1711 plusmn 71 a
77 484 plusmn 28 a 151 plusmn 16 c 668 plusmn 36 b 126 1429 plusmn 73 b151 199 plusmn 16 c 22 plusmn 05 d 272 plusmn 23 d 41 534 plusmn 42 d
P (CO2) lt 00001 06934 lt 00001 ndash dagger lt 00001P (UV-B) lt 00001 lt 00001 lt 00001 ndash lt 00001P (CO2 yen UV-B) 00695 00345 0 1863 ndash 00364
Means followed by the same letter within a column are not significantly different (P gt 005)daggerStatistical analysis was not carried out because roots were harvested from entire soil bin and not for each plant within each SPAR unit
Figure 2 Effects of elevated CO2 and UV-B radiation on net photosynthetic rate of uppermost fully expanded mainstem leaves at 28 44 and 64 d after emergence Each data point is the mean plusmn SE of five plants
778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

778 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
cantly enhanced cotton leaf Pn under either no UV-B or77 kJ m-2 d-1 UV-B radiation
Leaf non-structural carbohydrate concentrations areclosely related to leaf Pn Our results of elevated [CO2]increasing cotton leaf non-structural carbohydrate (espe-cially sucrose and starch) concentrations are similar to thefindings in faba bean (Visser et al 1997) and in Ricinuscommunis (Grimmer Bachfischer amp Komor 1999) Sucrosecontrols the expression of ADP-glucose pyrophosphory-lase leading to a shift of carbohydrate partitioning intostarch when sucrose synthesis is more rapid than consump-
tion andor export at elevated [CO2] (Grimmer et al 1999)Although leaf photosynthetic acclimation may exist whenplants are grown under elevated [CO2] (Visser et al 1997)due to accumulation of non-structural carbohydrates inleaves our results clearly indicated that elevated [CO2]stimulated leaf Pn (Fig 2) and increased photo-assimilatesupply resulting in rapid growth and biomass accumulationin cotton (Tables 4 amp 5) These results are consistent withthe findings of Begonia et al (1996)
Effects of UV-B radiation
Plant growth physiological and yield responses to UV-Bradiation differ considerably among crop species or culti-vars (Corlett et al 1997 Krupa et al 1998) Our studyrevealed that UV-B radiation of 77 kJ m-2 d-1 the amountessentially equivalent to that found on summer clear daysin todays ambient environment in the study site (TOMS2002) did not affect cotton growth and development buthigher levels of UV-B radiation (151 kJ m-2 d-1) theamount expected to occur with a 30 depletion in thestratospheric ozone significantly reduced stem elongationrate leaf area and dry matter accumulation Decreasedplant height due to high UV-B radiation was closely relatedto shorter internodes rather than a fewer number of nodesThe results of UV-B radiation effect on cotton growth inour study agree with the earlier reports in pea (Nogues et al1998) and many other crops (Krupa et al 1998)
In terms of the components of biomass UV-B radiationhad the greatest effect on fruit dry weight Although totalbiomass and leaf Pn of the 77 kJ UV-B-treated plants didnot differ from that of 0 kJ UV-B-treated control plantsunder ambient CO2 level the fruit dry weight was signifi-cantly lower as UV-B levels increased (Table 5) Reductionin fruit dry weight was related to a higher fruit abscissionor fewer bolls retained per plant Furthermore floral budabscission was more than boll abscission in both the 77 and151 kJ UV-B treatments The increase in floral bud abscis-sion by 77 kJ UV-B radiation was probably associated withsome other causes rather than photo-assimilate supplybecause either leaf Pn or leaf non-structural carbohydrateswere not different between the 77 kJ UV-B-treated plantsand the control plants Demchik amp Day (1996) demon-strated that enhanced UV-B radiation decreased pollenquantity and quality of Brassica rapa We expect pollensterility or a reduction in viable pollen production may beone of factors increasing young boll abscission at high UV-B radiation The mechanisms of UV-B radiation effect oncotton fruit abscission still need further investigation In arecent study Song et al (1999) found that a daily supple-ment of 92 kJ m-2 UV (280ndash400 nm) radiation for 169 ddecreased dry matter production of field-grown cottonOur study further indicated that decreased total biomassproduction due to high UV-B radiation was closelyrelated to both small leaf area and low leaf Pn duringsquaring and fruiting (R2 = 073ndash097 Tables 2 amp 5 Fig 2)The result of elevated UV-B causing the greatest decreasein the fruit dry weight among plant parts due to high fruit
Figure 3 Effects of elevated CO2 and UV-B radiation on con-centrations of leaf chlorophyll and carotenoids and chlorophyll a b ratio Data represent means + 1 SE of six leaf samples mea-sured at first flower and 3 weeks after the first flower
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002
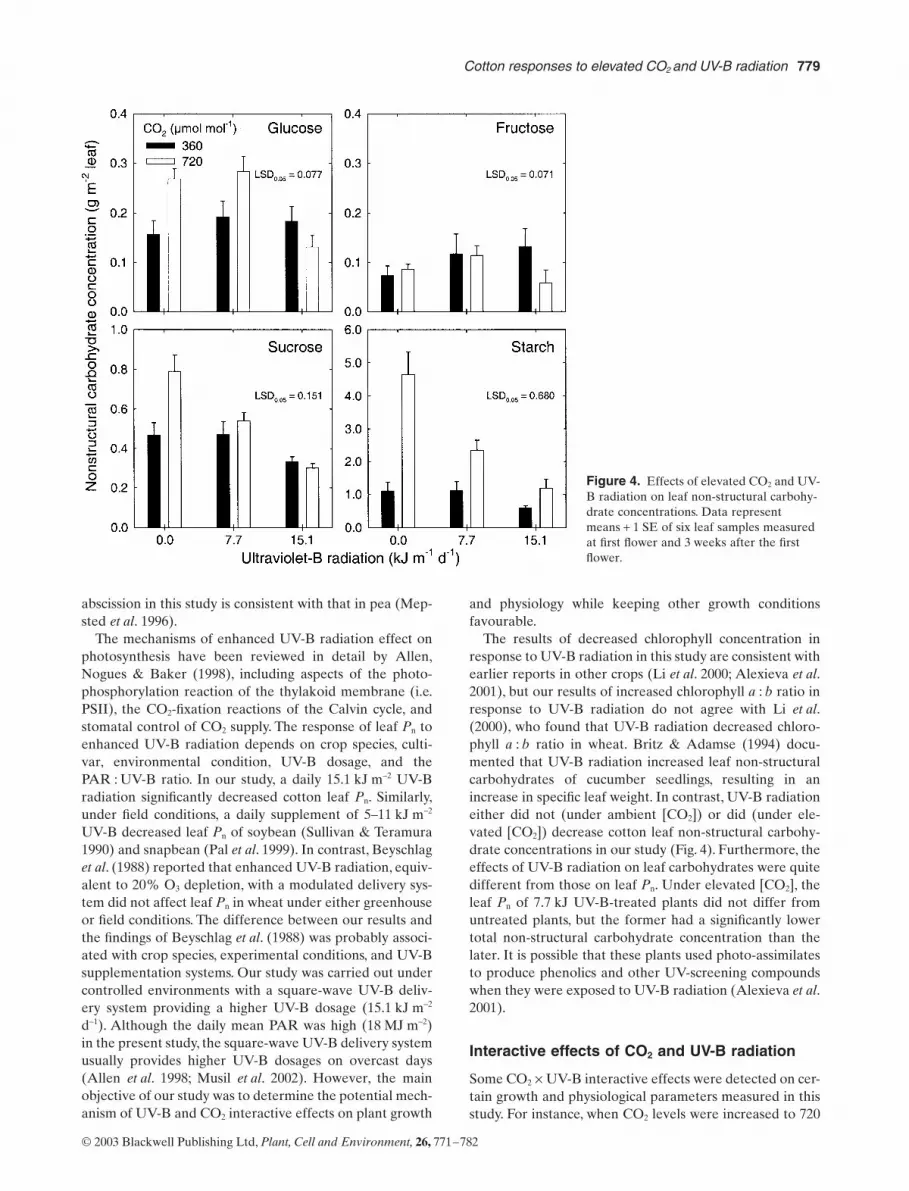
Cotton responses to elevated CO2 and UV-B radiation 779
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
abscission in this study is consistent with that in pea (Mep-sted et al 1996)
The mechanisms of enhanced UV-B radiation effect onphotosynthesis have been reviewed in detail by AllenNogues amp Baker (1998) including aspects of the photo-phosphorylation reaction of the thylakoid membrane (iePSII) the CO2-fixation reactions of the Calvin cycle andstomatal control of CO2 supply The response of leaf Pn toenhanced UV-B radiation depends on crop species culti-var environmental condition UV-B dosage and thePAR UV-B ratio In our study a daily 151 kJ m-2 UV-Bradiation significantly decreased cotton leaf Pn Similarlyunder field conditions a daily supplement of 5ndash11 kJ m-2
UV-B decreased leaf Pn of soybean (Sullivan amp Teramura1990) and snapbean (Pal et al 1999) In contrast Beyschlaget al (1988) reported that enhanced UV-B radiation equiv-alent to 20 O3 depletion with a modulated delivery sys-tem did not affect leaf Pn in wheat under either greenhouseor field conditions The difference between our results andthe findings of Beyschlag et al (1988) was probably associ-ated with crop species experimental conditions and UV-Bsupplementation systems Our study was carried out undercontrolled environments with a square-wave UV-B deliv-ery system providing a higher UV-B dosage (151 kJ m-2
d-1) Although the daily mean PAR was high (18 MJ m-2)in the present study the square-wave UV-B delivery systemusually provides higher UV-B dosages on overcast days(Allen et al 1998 Musil et al 2002) However the mainobjective of our study was to determine the potential mech-anism of UV-B and CO2 interactive effects on plant growth
and physiology while keeping other growth conditionsfavourable
The results of decreased chlorophyll concentration inresponse to UV-B radiation in this study are consistent withearlier reports in other crops (Li et al 2000 Alexieva et al2001) but our results of increased chlorophyll a b ratio inresponse to UV-B radiation do not agree with Li et al(2000) who found that UV-B radiation decreased chloro-phyll a b ratio in wheat Britz amp Adamse (1994) docu-mented that UV-B radiation increased leaf non-structuralcarbohydrates of cucumber seedlings resulting in anincrease in specific leaf weight In contrast UV-B radiationeither did not (under ambient [CO2]) or did (under ele-vated [CO2]) decrease cotton leaf non-structural carbohy-drate concentrations in our study (Fig 4) Furthermore theeffects of UV-B radiation on leaf carbohydrates were quitedifferent from those on leaf Pn Under elevated [CO2] theleaf Pn of 77 kJ UV-B-treated plants did not differ fromuntreated plants but the former had a significantly lowertotal non-structural carbohydrate concentration than thelater It is possible that these plants used photo-assimilatesto produce phenolics and other UV-screening compoundswhen they were exposed to UV-B radiation (Alexieva et al2001)
Interactive effects of CO2 and UV-B radiation
Some CO2 yen UV-B interactive effects were detected on cer-tain growth and physiological parameters measured in thisstudy For instance when CO2 levels were increased to 720
Figure 4 Effects of elevated CO2 and UV-B radiation on leaf non-structural carbohy-drate concentrations Data represent means + 1 SE of six leaf samples measured at first flower and 3 weeks after the first flower
780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

780 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
from 360 mmol mol-1 cotton leaf Pn increased by about40 under no UV-B or 77 kJ m-2 d-1 UV-B radiationwhereas leaf Pn was almost unchanged by elevated [CO2]under 151 kJ UV-B radiation during flowering and fruiting(44 and 64 DAE) In an earlier study Adamse amp Britz(1992) found that UV-B-induced reductions in plant growthwere less severe when elevated [CO2] was used to increasePn Rozema et al (1997) also reported a significantCO2 yen UVndashB interaction for plant biomass production inElymus athericus as plant growth was much less reducedby enhanced UV-B radiation at 720 mmol CO2 mol-1 thanat ambient atmospheric [CO2] However the evidence ofelevated [CO2] mitigating UV-B detrimental effect on cot-ton dry matter accumulation was not detected in thepresent study The interactive effect of [CO2] and UV-Bradiation on fruit dry weight was detected such that plantsgrown in elevated [CO2] appeared to be more sensitive tohigh UV-B radiation in comparison with plants grown inambient CO2 level (Table 5) Under ambient [CO2] fruitdry weights of the 77 and 151 kJ UV-B-treated plants were81 and 15 of the control plants respectively In contrastfruit dry weights of the two UV-B treatments were only 58and 11 of the control under elevated [CO2] Thereforean important finding of this study is that elevated [CO2] didnot mitigate the detrimental effects of high UV-B radiationon cotton growth and physiology especially fruit growth
A recent study (Smith Burritt amp Bannister 2000)reported that dry matter accumulation is a good indicatorof screening plant sensitivity to UV-B radiation among veg-etable crop species and a rapid growth rate renders plantsmore sensitive to the injurious effects of UV-B radiationBased on the findings of Smith et al (2000) and our resultswe speculate that the sensitivity of cotton to UV-B radia-tion may change with growth stages and be most sensitiveto high UV-B radiation during squaring and early floweringwhen plants are growing rapidly Cotton growth is oftenaccelerated by CO2 enrichment as shown in Fig 1 and inReddy Hodges amp McKinion (1997) Thus we observedmore injurious effects from the UV-B radiation in plantsgrown in elevated [CO2] which was supported from mea-surements of Pn and soluble sugars in the uppermost fullyexpanded leaves (Figs 2 and 4)
As mentioned earlier this experiment was conducted insunlit chambers under controlled environments and usinga square-wave UV-B delivery system The square-waveUV-B system usually overestimates UV-B dosages on over-cast days (Musil et al 2002) Data for each treatment werecollected from nine plants in a SPAR unit due to limitednumbers of chambers Although evidence indicated no sys-tematic differences among the chambers (Table 1) restric-tion of sufficient numbers of the chambers for individualtreatments and square-wave UV-B system might limit ourstrong conclusions in this study
In summary levels near the current maximum solar UV-B radiation (77 kJ m-2 d-1) observed in United States mid-south cotton production area on sunny days between Mayand July did not affect cotton leaf Pn and total biomassproduction but significantly increased fruit abscission and
decreased fruit dry matter accumulation in comparisonwith plants that received no UV-B radiation A doubling ofthe current UV-B radiation to 151 kJ m-2 d-1 furtherreduced not only fruit production but also total biomassDecreased biomass from high UV-B radiation was closelyrelated to both smaller leaf area and lower leaf Pn Elevated[CO2] significantly increased Pn growth and dry matteraccumulation in cotton under no UV-B or 77 kJ m-2 d-1
UV-B conditions However elevated atmospheric [CO2]could not alleviate the detrimental effects of high UV-Bradiation on cotton Pn and growth particularly on repro-ductive growth
ACKNOWLEDGMENTS
The project was funded in part by the USDA UV-B Mon-itoring Program and the RSTC-NASA We thank Dr DGitz for his help in the initial UV-B facility setup Dr M FBalla for the use of their laboratory for non-structural car-bohydrate measurements W Ladner D Brand and KGourley for technical support and Drs D J Allen T MBrennan and D J Burritt for reviewing the manuscript andtheir helpful comments Contribution from the Departmentof Plant and Soil Sciences Mississippi State University Mis-sissippi Agricultural and Forestry Experiment Stationpaper no J10128
REFERENCES
Adamse P amp Britz SJ (1992) Amelioration of UV-B damageunder high irradiance I Role of photosynthesis Photochemistryand Photobiology 56 645ndash650
Alexieva V Sergiev I Mapelli S amp Karanov E (2001) The effectof drought and ultraviolet radiation on growth and stress mark-ers in pea and wheat Plant Cell and Environment 24 1337ndash1344
Allen DJ Nogues S amp Baker NR (1998) Ozone depletion andincreased UV-B radiation is there a real threat to photosynthe-sis Journal of Experimental Botany 49 1775ndash1788
Balakumar T Hani BVV amp Paliwal K (1993) On the interactionof UV-B radiation (280ndash315 nm) with water stress in crop plantsPhysiologia Plantarum 87 217ndash222
Begonia GB Russ B Cunningham SG amp Moore R (1996)Evidence of no photosynthetic acclimation in cotton exposed tostage-specific CO2 enrichment Journal of the Mississippi Acad-emy of Science 41 95ndash98
Beyschlag W Barnes PW Flint SD amp Caldwell MM (1988)Enhanced UV-B irradiation has no effect on photosyntheticcharacteristics of wheat (Triticum aestvum L) and wild oat(Avena fatua L) under greenhouse and field conditions Photo-synthetica 22 516ndash525
Britz SJ amp Adamse P (1994) UV-B-induced increase in specificleaf weight of cucumber as a consequence of increased starchcontent Photochemistry and Photobiology 60 116ndash119
Chappelle EW Kim MS amp McMurtrey JE III (1992) Ratioanalysis of reflectance spectra (RARS) An algorithm for theremote estimation of the concentrations of chlorophyll a chlo-rophyll b and carotenoids in soybean leaves Remote Sensing ofthe Environment 39 239ndash247
Christiansen MN (1986) Influence of atmospheric parameters ongrowth and development In Cotton Physiology (eds JRMauney amp JM Stewart) pp 39ndash46 The Cotton FoundationPublisher Memphis TN USA
Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

Cotton responses to elevated CO2 and UV-B radiation 781
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Corlett JE Stephen J Jones HG Woodfin R Mepsted R ampPaul ND (1997) Assessing the impact of UV-B radiation on thegrowth and yield of field crops In Plants and UV-B Responsesto Environmental Change (ed PJ Lumsden) pp 195ndash211 Cam-bridge University Press Cambridge UK
Dai Q Coronal VP Vergara BS Barnes PW amp Quintos AT(1992) Ultraviolet-B radiation effects on growth and physiologyof four rice cultivars Crop Science 32 1269ndash1274
Demchik SM amp Day TA (1996) Effect of enhanced UV-B radi-ation on pollen quantity quality and seed yield in Brassica rapaAmerican Journal of Botany 83 573ndash579
Gerik TJ Oosterhuis DM amp Torbet HA (1998) Managing cot-ton nitrogen supply Advances in Agronomy 64 115ndash147
Gonzalez R Paul ND Percy K Ambrose M McLaughlinCK Barnes JD Areses M amp Wellburn AR (1996)Responses to ultraviolet-B (280ndash315 nm) of pea (Pisum sativumL) lines differing in leaf surface wax Physiologia Plantarum98 852ndash860
Green AES Cross KR amp Smith LA (1980) Improved analyt-ical characterization of ultraviolet skylight Photochemistry andPhotobiology 31 59ndash65
Grimmer C Bachfischer T amp Komor E (1999) Carbohydratepartitioning into starch in leaves of Ricinus communis L grownunder elevated CO2 is controlled by sucrose Plant Cell andEnvironment 22 1275ndash1280
Hendrix DL (1993) Rapid extraction and analysis of nonstruc-tural carbohydrates in plant tissues Crop Science 33 1306ndash1311
Houghton JT Ding D Griggs DJ Noguer M van der LindenPJ Dai X Maskell K amp Johnson CA (2001) The ScientificBasis Contribution of Working Group I to the Third AssessmentReport of the Intergovernmental Panel on Climatic Change(IPCC) Cambridge University Press Cambridge UK
Jordan BR Chow WS amp Anderson JM (1992) Changes inmRNA levels and polypeptide subunits of ribulose 1 5-bispho-sphate carboxylase in response to supplementary ultraviolet-Bradiation Plant Cell and Environment 15 91ndash98
Kimball BA (1983) Carbon dioxide and agricultural yield anassemblage and analysis of 430 prior observations AgronomyJournal 75 779ndash788
Kimball BA amp Mauney JR (1993) Response of cotton to varyingCO2 irrigation and nitrogen yield and growth Agronomy Jour-nal 85 706ndash712
Krupa SV Kickert RN amp Jaumlger HJ (1998) Elevated UV-Bradiation and Crops In Elevated Ultraviolet (UV)-B Radiationand Agriculture (eds SV Krupa RN Kickert amp HJ Jaumlger)pp 104ndash131 Springer-Veriag Berlin Heidelberg and LandesBioscience Georgetown TX USA
Lawlor DW amp Mitchell RAC (2000) Crop ecosystem responsesto climatic change Wheat In Climate Change and Global CropProductivity (eds KR Reddy amp HF Hodges) pp 57ndash80 CABInternational Wallingford UK
Li Y Zu Y Chen J Yang J amp Hu Z (2000) Intraspecific differ-ences in physiological response of 20 wheat cultivars toenhanced ultraviolet-B radiation under field conditions Envi-ronmental and Experimental Botany 44 95ndash103
Madronich S McKenzie RL Bjorn LO amp Caldwell MM(1998) Changes in biologically active ultraviolet radiation reach-ing the Earths surface Journal of Photochemistry and Photobi-ology B Biology 46 5ndash19
Mark U amp Tevini M (1996) Combination effect of UV-B radiationand temperature on sunflower and maize seedlings Journal ofPlant Physiology 148 49ndash56
Mauney JR Kimball BA Pinter PJ Jr LaMorte RL LewinKF Nagy J amp Hendrey GR (1994) Growth and yield ofcotton in response to free-air carbon dioxide enrichment
(FACE) environment Agricultural and Forestry Meterology 7049ndash68
Mepsted R Paul ND Stephen J Corlett JE Nogues S BakerNR Jones HG amp Ayres PG (1996) Effect of enhanced UV-B radiation on pea (Pisum sativum L) grown under field condi-tions in the UK Global Change Biology 2 325ndash334
Monje O amp Bugbee B (1998) Adaptation to high CO2 concentra-tion in an optimal environment radiation capture canopy quan-tum yield and carbon use efficiency Plant Cell and Environment21 315ndash324
Murali NS amp Teramura AH (1985) Effect of UV-B radiation onsoybean VI Influence of phosphorus nutrition of growth andflavonoid content Plant Physiology 63 413ndash416
Musil CF Bjoumlrn LO Scourfield MWJ amp Bodeker GE (2002)How substantial are ultraviolet-B supplementation inaccuraciesin experimental square-wave delivery systems Environmentaland Experimental Botany 47 25ndash38
Nogues S Allen DJ Morison JIL amp Baker NR (1998) Ultra-violet-B radiation effects on water relations leaf developmentand photosynthesis in droughted pea plants Plant Physiology17 173ndash181
Nogues S Allen DJ Morison JIL amp Baker NR (1999) Char-acterization of stomatal closure caused by ultraviolet-B radia-tion Plant Physiology 121 489ndash496
Pal M Sengupta UK Srivastava AC Jain V amp Meena RC(1999) Changes in growth and photosynthesis of mungbeaninduced by UV-B radiation Indian Journal of Plant Physiology4 79ndash84
Radoglou KM amp Jarvis PG (1992) The effects of CO2 enrich-ment and nutrient supply on growth morphology and anatomyof Phaseolus vulgaris L seedlings Annals of Botany 70 245ndash256
Reddy KR Hodges HF amp Kimball BA (2000) Crop ecosystemresponses to global climate change cotton In Climate Changeand Global Crop Productivity (eds KR Reddy amp HF Hodges)pp 162ndash187 CAB International Wallingford UK
Reddy KR Hodges HF amp McKinion JM (1997) A comparisonof scenarios for the effect of global climate change on cottongrowth and yield Australian Journal of Plant Physiology 24707ndash713
Reddy KR Read JJ Baker JT McKinion JM Tarpley LHodges HF amp Reddy VR (2001) Soil-Plant-Atmosphere-Research (SPAR) facility ndash a tool for plant research and mod-eling Biotronics 30 27ndash50
Rozema J Lenssen GM Staaij JWM Tosserams M VisserAJ amp Broekman RA (1997) Effects of UV-B radiation onterrestrial plants and ecosystems interaction with CO2 enrich-ment Plant Ecology 128 182ndash191
SAS Institute Inc (1997) SAS Users Guide Statistics SAS Insti-tute Inc Cary NC USA
Smith JL Burritt DJ amp Bannister P (2000) Shoot dry weightchlorophyll and UV-B-absorbing compounds as indicators of aplants sensitivity to UV-B radiation Annals of Botany 86 1057ndash1063
Song YZ Zhang YF Wan CJ Wang CH amp Zhang FC (1999)Impact of intensified ultraviolet radiation on cotton growthJournal of Nanjing Institute of Meteorology 22 269ndash273
Stapleton AE (1992) Ultraviolet radiation and plants burningquestions Plant Cell 105 881ndash889
Sullivan JH (1997) Effects of increasing UV-B radiation and CO2
on photosynthesis and growth implications for terrestrial eco-systems Plant Ecology 128 194ndash206
Sullivan JH amp Teramura AH (1990) Field study of the interac-tion between solar ultraviolet-B radiation and drought on pho-tosynthesis and growth in soybean Plant Physiology 92 141ndash146
782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002

782 D Zhao et al
copy 2003 Blackwell Publishing Ltd Plant Cell and Environment 26 771ndash782
Teramura AH Sullivan JH amp Ziska LH (1990) Interaction ofelevated ultraviolet-B radiation and CO2 in productivity andphotosynthetic characteristics in wheat rice and soybean PlantPhysiology 94 470ndash475
TOMS (2002) Total Ozone Mapping Spectrometer Data productErythemal UV exposure [online] Available at httptomsgsfcnasagovery_uveuvhtml (verified 04 September2002)
Visser AJ Tosserams M Groen MW Magendans GWH ampRozema J (1997) The combined effects of CO2 concentration
and solar UV-B radiation on faba bean grown in open-top cham-bers Plant Cell and Environment 20 189ndash199
WMO (1995) Scientific assessment of ozone depletion 1994 WorldMeteorological Organization Global Ozone Research and Mon-itoring Project Report No 37 World Meteorological Organiza-tion Geneva Switzerland
Received 5 September 2002 received in revised form 2 December2002 accepted for publication 5 December 2002





























