Embed Size (px)
Citation preview

Histochemical Similarities of MucinsProduced by Brunner’s Glands and
Pyloric Glands: A Comparative StudyUDO SCHUMACHER,1* MOSES DUKU,2 MARCUS KATOH,1 JULIA JORNS,1
AND WILLIAM J. KRAUSE3
1Institute for Anatomy II, Experimental Morphology, University HospitalHamburg-Eppendorf, Hamburg, Germany
2Department of Pathology and Anatomical Sciences, University ofMissouri–Columbia, Columbia, Missouri
3Human Morphology, University of Southampton, Southampton, England
ABSTRACTMucins of the gastroduodenal junction are secreted by the mucous surface and mucus-
producing glandular cells in the stomach, and by goblet cells and Brunner’s glands in theduodenum. Developmental studies have demonstrated that Brunner’s glands can arise fromundifferentiated gastric epithelium and/or intestinal epithelium in the proximal duodenum.The aim of this study was to investigate the carbohydrate composition of mucins from thisregion and compare it with that of mucins from Brunner’s glands to evaluate the probableevolution of mucins from these glands. Toward that end, paraffin sections from 13 mamma-lian species were stained by classic carbohydrate histochemistry and treated with 13 lectins.In general, the mucous surface cells of the stomach, pyloric glands, duodenal goblet cells, andBrunner’s glands secretory epithelium had different lectin-binding patterns. However, thelectin-binding profile of the secretory epithelium of Brunner’s glands resembled that ofpyloric glands more closely than that of duodenal goblet cells and mucous surface cells of thestomach. Mucins from Brunner’s glands and pyloric glands showed a greater terminalcarbohydrate residue diversity than those of gastric mucous surface cells or duodenal gobletcells. The lectin-binding profile argues for the evolution of similar mucins from the epitheliaof Brunner’s glands and pyloric glands. The greater diversity of carbohydrate residues inmucins secreted by Brunner’s glands suggests that their mucus is more adaptable. This mayexplain why Brunner’s glands metaplasia rather than goblet cell metaplasia is seen in themucosa adjacent to chronic intestinal ulcers. Anat Rec Part A 278A:540–550, 2004.© 2004 Wiley-Liss, Inc.
Key words: Brunner’s glands; carbohydrate histochemistry; comparativeanatomy; gastroduodenal junction; histogenesis; lectin; mucin
The epithelial cells that line the gastrointestinal tract ofmammals are protected in part from the somewhat harshenvironment of acid, proteolytic enzymes, and abrasives inthe lumen by a mucus layer. In the stomach the mucuslayer, which is several microns thick, is secreted by boththe epithelium lining the gastric mucosa and mucus-pro-ducing cells of the gastric glands, while in the duodenumit is secreted by goblet cells and Brunner’s glands, whichare located in the submucosa. Mucus is a highly hydratedgel that consists of about 95% water, 5% mucins, andminor components, such as electrolytes (Allen, 1981; Neu-tra and Forstner, 1987). Mucins are high-molecular-weight glycoproteins that have gel-forming propertieswhich are highly glycosylated and which in turn are re-sponsible for their protective function.
A particular morphological feature of the duodenal sub-mucosa is the presence of Brunner’s glands. Brunner’sglands are specific to mammals and have been observed inall mammals examined to date (Krause, 1988). They are
*Correspondence to: Udo Schumacher, Institute for AnatomyII, University Hospital Hamburg-Eppendorf, Martinistrasse 52,D-20246 Hamburg, FRG. Fax: (�) 49-40-42803-5427.E-mail: [email protected]
Received 18 September 2002; Accepted 17 February 2004DOI 10.1002/ar.a.20046
THE ANATOMICAL RECORD PART A 278A:540–550 (2004)
© 2004 WILEY-LISS, INC.

mucus-secreting, branched, tubuloalveolar glands locatedin the submucosa of the duodenum beginning at the gas-trointestinal junction and extending for various distancesalong the proximal small intestine. They are not usuallyfound beyond the entrance of the pancreatic duct. Al-though it has been an accepted view that the secretoryproduct of Brunner’s glands, which contains mucus glyco-protein and bicarbonate, protects the duodenal mucosa byneutralising acid in the acid-chyme from the stomach,recent studies showed that no lesions occurred in theduodenum of rats following surgical removal of Brunner’sglands (Krause et al., 1988). The study from Krause et al.(1988) demonstrated that bicarbonate secretion originat-ing from the duodenal epithelium itself may play a greaterrole in neutralising the acid from the stomach compared tothat originating from Brunner’s glands. Hence, the role ofmucin is emphasised over that played by the bicarbonatesecreted by Brunner’s glands.
In humans, the Brunner’s glands begin to differentiatefrom the bases of the crypts of Lieberkuhn just distal tothe gastroduodenal junction after about 3–5 months ofintrauterine life. Comparative studies on other mammalshave shown that Brunner’s glands can originate directlyfrom the duodenal epithelium, from the crypts ofLieberkuhn, or in some cases from both sites (for review,see Krause, 1988). In addition, in marsupials such asDidelphis virginia, some Brunner’s glands also originatefrom the pyloric epithelium lining the distal most extremepart of the stomach. This developmental feature supportsthe observation that mucins of the Brunner’s glands aremore similar to mucins of the pyloric glands than to thoseof the duodenal goblet cells (Schumacher and Krause,1995).
The aim of this study was to compare and contrast thecarbohydrate composition of mucins in mucus-secretingcells on both sides of the gastroduodenal junction. Classicmucin and lectin histochemical techniques were used toachieve this aim. Lectins are carbohydrate-binding pro-teins that can be used for the precise localisation of de-fined terminal carbohydrate residues in tissue sections(Brooks et al., 1997). Lectin histochemistry is thereforeuseful for defining the terminal carbohydrate residue pro-files of mucins from the stomach, duodenum, and Brun-ner’s gland cells. A comparison of the molecular profilesmay subsequently shed some light on the evolution ofmucins produced by epithelial cells on either side of thegastroduodenal junction.
MATERIALS AND METHODSThe gastroduodenal junction in 13 mammalian species
(for the number and order of the mammals, see Table 1)was studied. The tissues were fixed in 4% neutral bufferedformalin and routinely processed to paraffin wax. Sectionswere cut at 5 �m thickness, deparaffinized in xylene, andrehydrated through a series of graded ethanols.
The sections were mounted on L-polylysine-coatedslides and stained with haematoxylin and eosin (H&E),periodic acid-Schiff (PAS), and defined Alcian blue stainsfor carboxylated (pH 2.5) and sulphated (pH 1.0) residues(Schumacher and Adam, 1994).
PAS TechniqueThe sections were oxidized for 5 min in 1% aqueous
periodic acid, washed under running tap water for 5 min,rinsed in distilled water, and then treated with Schiff’sreagent for 15 min. Afterwards, the sections were washedunder running tap water for 10 min and counterstainedwith haematoxylin. Finally, the sections were dehydratedin alcohol, cleared in xylene, and mounted in a resinousmountant.
Alcian Blue Method (pH 2.5 and 1.0)For the pH 2.5 method, the sections were stained in
freshly filtered 1% Alcian blue 8 GX in 3% acetic acid (pH2.5) for 30 min and washed in water. For the pH 1.0method, the sections were stained in 1% Alcian blue 8 GXin 0.1 N hydrochloric acid for 30 min and rinsed briefly in0.1 N HCl. The sections were dried with fine filter paper.In both methods, the sections were dehydrated in alcohol,cleared in xylene, and mounted in a resinous mountant.
Lectin HistochemistryWe used 13 biotin-conjugated lectins (all obtained from
Sigma, Poole, Dorset; for their origin, abbreviations, andnominal sugar specificity, see Table 2) in an indirect tech-nique with the avidin-biotin peroxidase/DAB method. Thesections were incubated in Tris-buffered saline (TBS) con-taining 50 mM Tris, 150 mM NaCl, and 1 mM MgCl2,adjusted to pH 7.6. They were then treated with 0.1 %trypsin (Sigma, St. Louis, MO) for 15 min at 37°C. Thesections were again transferred to TBS, washed threetimes, and incubated with the biotinylated lectins at aconcentration of 10 �g ml–1 for 60 min. After a further
TABLE 1. Taxonomic name, number and order of the mammals used in this study
Taxonomic name Number Order
Homo sapiens (human) 4 PrimatesMacaca fuscata (Japanese macaque) 5 PrimatesMacaca mulatta (rhesus macaque) 6 PrimatesRattus rattus (rat, Sprague-Dawley) 3 RodentiaMicrotis ochrogaster (prairie vole) 3 RodentiaCavia porcellus (Guinea pig) 3 RodentiaOdocoileus virginianus (white-tailed deer) 4 ArtiodactylaBison bison (bison) 4 ArtiodactylaOryctolagus cuniculus (domestic rabbit, Flemish giant) 6 LagomorphaSylvilagus floridanus (eastern cottontailed rabbit) 5 LagomorphaFelis catus (domestic cat, feral) 4 CarnivoraProcyon lotor (common raccoon) 4 CarnivoraDidelphis marsupialis (southern opossum) 5 Marsupialia
541BRUNNER’S GLAND MUCINS

wash in TBS, the sections were treated with an avidin-horseradish peroxidase complex followed by a wash inTBS. For visualisation, the sections were incubated withDAB-hydrogen peroxidase for 10 min. A light haematoxy-lin stain was used as a counterstain after the sectionswere washed under running tap water for approximately 5min. Finally, the sections were dehydrated in alcohol,cleared in xylene, and mounted in a resinous mountant(Brooks et al., 1997).
Control Inhibition With InhibitoryMonosaccharides
All of the sugars (galactose, mannose, N-acetyl-glu-cosamine (glcNAc), and N-acetyl-galactosamine (galNAc))were prepared as a 200-mM solution, and the lectins weredissolved at a concentration of 20 �g ml–1. The solution ofsugars and lectins was mixed in a ratio of 1:1, resulting ina final concentration of 0.1 mol sugar and 10 �g ml–1
lectin, and the solution was incubated on the vortexer for1 hr. The preincubated lectins were subsequently used forthe lectin histochemistry as described above.
Control Inhibition With Bovine Thyroglobulinin the Case of PHA-L
Bovine thyroglobulin and lectins were prepared as a 20�g ml–1 solution. Again, one part of lectin solution wasmixed with one part of the thyroglobulin solution. Thismixture was incubated for 1 hr before it was applied to thetissue sections.
Removal of Neuraminic Acid in the Case ofMAA and SNA-I
Ten units of neuraminidase were dissolved in 70 ml 0.5M sodium acetate buffer (pH 5.5). The slides were placedin a humid chamber and incubated overnight at 37°C.After the slides were washed in lectin buffer, regular MAAand SNA-I staining was carried out as described above.
Photographs were taken with Ilford PANF 50 black andwhite film on a Zeiss Axiophot 2 photomicroscope.
RESULTSThe general histology of the stomach and duodenum
was essentially similar to that described in the literature,and the results of the conventional histology are men-tioned below only if they differ significantly from previous
findings. The results of the classic carbohydrate histo-chemistry are summarised in Table 3.
The vast majority of the inhibition experiments showeda total or at least very definite inhibition of lectin binding.The brush border was the only structure in the mamma-lian sections that was not completely inhibited even if thelectins were applied with their nominal inhibitorymonosaccharide.
Human (Fig. 1).In the stomach, mucous surface cellsand pyloric glands stained with PAS only. In the duode-num, goblet cells stained with PAS and Alcian blue at bothpH 1.0 and 2.5, whereas the Brunner’s glands, like themucous surface cells and pyloric glands, stained only withPAS.
The mucous surface cells reacted with four lectins only(VVA, MPA, JAC, and PNA), while the pyloric glandsreacted with nine lectins (Con A, WGA, HPA, MPA, JAC,PNA, SNA, LTA, and UEA-I (Fig. 14)). In the duodenum,goblet cells reacted only with HPA, MPA (Fig. 16), andJAC. In addition to reacting with VVA, the Brunner’sglands reacted with the same lectins as did the pyloricglands (Con A, WGA (Fig. 21), HPA, MPA (Fig. 24), JAC,PNA, SNA, LTA, and UEA-I (Fig. 13)).
Rhesus macaque (Fig. 2).The mucous surface cellsof the stomach stained with PAS and Alcian blue at bothpH 1.0 and 2.5, whereas the pyloric glands stained withPAS and Alcian blue only at pH 2.5. In the duodenum, thegoblet cells, like the mucous surface cells, stained withPAS and Alcian blue at both pH 1.0 and 2.5, while Brun-ner’s glands stained only with PAS.
In the stomach, mucous surface cells reacted with WGA,HPA, MPA, JAC, and PNA, while pyloric glands reactedwith Con A WGA, HPA, MPA, JAC, PNA, LTA, andUEA-I. In the duodenum, goblet cells reacted only withHPA, MPA, and JAC. Brunner’s glands reacted with all ofthe lectins that reacted with the pyloric glands (Con AWGA, HPA, MPA, JAC, PNA, LTA, and UEA-I). The re-action with LTA and UEA-I demonstrated the same in-tensity and cytoplasmic pattern in both the Brunner’s andpyloric glands.
Japanese macaque (Fig. 3).In the stomach, mucoussurface cells stained with PAS and Alcian blue at both pH1.0 and 2.5, whereas pyloric glands stained with PAS only.In the duodenum, goblet cells stained with PAS and Al-
TABLE 2. Source, abbreviation, and sugar specificity of the lectins used
Source species Abbreviation Major sugar specificity
Canavalia ensiformis Con A �-D-Man, �-D-GlcGalanthus nivalis GNA �-D-ManTriticum vulgaris WGA D-GlcNAcHelix pomatia HPA D-GalNAcVicia villosa VVA D-GalNAcArachis hypogaea PNA �-D-Gal1,�-D-GalNAcMaclura pomifera MPA �-D-Gal, �-D-GalNacArtocarpus integrifolia JAC �-D-GalLotus tetragonolobus LTA �-L-FucUlex europaeus UEA-I �-FucSambucus nigra SNA-I Neu5Ac�2,6Gal/ GalNAcMaackia amurensis MAA Neu5Ac�2,3Gal/ GalNAcPhaseolus vulgaris PHA-L Complex type carbohydrates
542 SCHUMACHER ET AL.

cian blue at both pH 1.0 and 2.5, while Brunner’s glands,like the pyloric glands, stained with PAS only.
The mucous surface cells of the stomach reacted withWGA, HPA, MPA, and JAC, while the cells of the pyloricglands reacted with Con A, WGA, HPA, MPA, JAC, PNA,LTA, and UEA-I. In the duodenum, the goblet cells re-acted only with WGA, MPA, and JAC. Brunner’s glandsreacted with almost the same intensity to all of the lectins(Con A, WGA, HPA, MPA, JAC, PNA, LTA, and UEA-I)that reacted with the pyloric glands.
Rat (Fig. 4).The specimens of rat, guinea pig, and voleduodenum examined lacked plicae circulares, and had avery thin submucosa and a poorly developed muscularismucosae.
In rats, the mucous surface cells stained only with PAS,whereas the pyloric glands and goblet cells stained with PASand Alcian blue at both pH 1.0 and 2.5. The Brunner’sglands, like the mucous surface cells, stained only with PAS.
In the stomach, mucous surface cells reacted with WGA,HPA (Fig. 26), and VVA (Fig. 23), while pyloric glandsreacted with Con A, WGA, HPA, VVA, MPA, JAC (Fig.
18), PNA, UEA-I (Fig. 15), and PHA-L. The duodenalgoblet cells reacted only with HPA and VVA (Fig. 22). Inaddition to reacting with LTA, the Brunner’s glands re-acted with all of the lectins that reacted with the pyloricglands (Con A, WGA, HPA (Fig. 25), MPA, JAC, PNA (Fig.20), UEA-I, LTA, and PHA-L).
Vole (Fig. 5).In voles, the mucous surface cells stainedonly with PAS, the pyloric glands stained with PAS andAlcian blue at pH 1.0, and the goblet cells and Brunner’sglands stained with PAS and Alcian blue at both pH 1.0and 2.5.
Mucous surface cells reacted with Con A, WGA, VVA,MPA, PNA, UEA-I, and PHA-L, while pyloric glands re-acted with Con A, WGA, HPA, VVA, MPA, JAC, PNA,LTA, UEA-I, and PHA-L. In the duodenum, goblet cellsreacted only with one lectin (UEA-I), while Brunner’sglands reacted with Con A, WGA, HPA, VVA, MPA, JAC,PNA, SNA, and UEA-I.
Guinea pig (Fig. 6).In guinea pigs, the mucous sur-face cells of the stomach stained only with PAS, while the
TABLE 3. Results of the classical carbohydrate histochemistry of the gastroduodenal junction
Mucussurface cells Pyloric glands Brunner’s glands Goblet cells
Human PAS ��� �� ��� ���AB pH 1.0 – – – ���AB pH 2.5 – – – ���
Rhesus macaque PAS ��� ��� ��� ���AB pH 1.0 � – – ��AB pH 2.5 �� �� – ���
Japanese macaque PAS ��� ��� ��� ���AB pH 1.0 � – – �AB pH 2.5 ��� – – ���
Rat PAS ��� � ��� ���AB pH 1.0 – �� – ���AB pH 2.5 – � – ���
Prairie vole PAS ��� �� ��� ���AB pH 1.0 – �� �� ��AB pH 2.5 – – �� ��
Guinea pig PAS ��� ��� ��� ���AB pH 1.0 – ��� ��� ��AB pH 2.5 – ��� ��� ��
Bison PAS ��� ��� ��� ���AB pH 1.0 �� �� �� ��AB pH 2.5 �� ��� ��� ���
White-tailed deer PAS ��� ��� �� ���AB pH 1.0 – �� �� ��AB pH 2.5 – �� �� ��
Rabbit PAS ��� �� ���/� ���AB pH 1.0 – �� �� ���AB pH 2.5 – � ��� ���
Domestic cat PAS ��� ��� ��� ���AB pH 1.0 – – – ���AB pH 2.5 ��� – – ���
Common raccoon PAS ��� ��� ��� ���AB pH 1.0 �� – – ���AB pH 2.5 �� – – ���
Southern opossum PAS ��� ��� ��� ���AB pH 1.0 – – – ���AB pH 2.5 – – – ���
PAS stains neutral carbohydrate residues, while AB at pH 2.5 stains carboxylateted and AB at pH 1.0 stains sulphatedmucins. If differences in the staining pattern between Brunner’s glands and duodenal goblet cells and Pyloric glands exists,the staining pattern of Brunner’s gland cells resembles that of the Pyloric glands more often than that of the duodenal gobletcells. PAS, periodic acid-Schiff; AB, Alcian blue; –, no staining; �, weak staining; ��, moderate staining; ���, intensestaining.
543BRUNNER’S GLAND MUCINS

pyloric glands, goblet cells, and Brunner’s glands stainedwith PAS and Alcian blue at both pH 1.0 and 2.5.
In the stomach, mucous surface cells reacted with ConA, WGA, HPA, VVA, MPA, JAC, PNA, and UEA-I, whilepyloric glands reacted with WGA, HPA, VVA, JAC, SNA,UEA-I, PHA-L, and MAA. In the duodenum, goblet cellsreacted only with one lectin (JAC), while Brunner’s glandsreacted with WGA, HPA, VVA, MPA, JAC, PNA, SNA,and UEA-I.
A striking histochemical similarity among the rat, vole,and guinea pig was the moderately intense to very intensereaction of enteroendocrine cells in the duodenum withSNA and PNA.
Bison (Fig. 8).Mucous surface cells of the stomach,pyloric glands, goblet cells, and Brunner’s gland cellsall stained with PAS and Alcian blue at both pH 1.0and 2.5.
Fig. 1. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in humans (0 � nostaining, 1 � weak staining, 2 � moderate staining, 3 � intense staining,and 4 � very intense staining). The staining intensity of every single indi-vidual was scored in this semiquantitative manner. The intermediate valuesindicate when staining intensity differences were noted between severalindividuals in each group. These represent the average values.
Fig. 2. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in rhesus ma-caques (for scoring, see Fig. 1).
Fig. 3. Summary of the lectin histochemical findings regarding the
mucus-producing cells of the gastroduodenal junction in Japanese ma-caques (for scoring, see Fig. 1).
Fig. 4. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in the rat (forscoring, see Fig. 1).
Fig. 5. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in voles (for scor-ing, see Fig. 1).
Fig. 6. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in guinea pigs (forscoring, see Fig. 1).
544 SCHUMACHER ET AL.

In the stomach, mucous surface cells reacted with WGA,HPA, VVA, SNA, and UEA-I, while cells of the pyloricglands reacted with Con A, WGA, HPA, VVA, MPA, JAC,SNA, LTA, and UEA-I. In the duodenum, goblet cellsreacted only with WGA, HPA, VVA, and SNA. In additionto reacting with PNA, Brunner’s glands reacted with all ofthe lectins (Con A, WGA, HPA, VVA, MPA (Fig. 17), JAC,SNA, LTA, and UEA-I) that reacted with the pyloricglands. However, LTA and PNA showed a positive reac-tion in only about 60% of the acini of Brunner’s glands.
White-tailed deer (Fig. 7).Mucous surface cells ofthe stomach stained only with PAS. Pyloric glands, duo-denal goblet cells, and Brunner’s glands all stained withPAS and Alcian blue at both pH 1.0 and 2.5.
All of the mucus-producing cells of both the stomach andthe duodenum reacted with HPA and MPA. In the stom-ach, mucous surface cells reacted with WGA, HPA, andMPA, while pyloric glands reacted with Con A, WGA,HPA, VVA, MPA, JAC, SNA, and UEA-I. In the duode-num, goblet cells reacted with HPA, MPA, VVA, and MPA.
Fig. 7. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in bison (for scor-ing, see Fig. 1).
Fig. 8. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in white-taileddeer (for scoring, see Fig. 1).
Fig. 9. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in cotton-tailedand domestic rabbits (for scoring, see Fig. 1).
Fig. 10. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in cats (for scor-ing, see Fig. 1).
Fig. 11. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in raccoons (forscoring, see Fig. 1).
Fig. 12. Summary of the lectin histochemical findings regarding themucus-producing cells of the gastroduodenal junction in opossums (forscoring, see Fig. 1).
545BRUNNER’S GLAND MUCINS
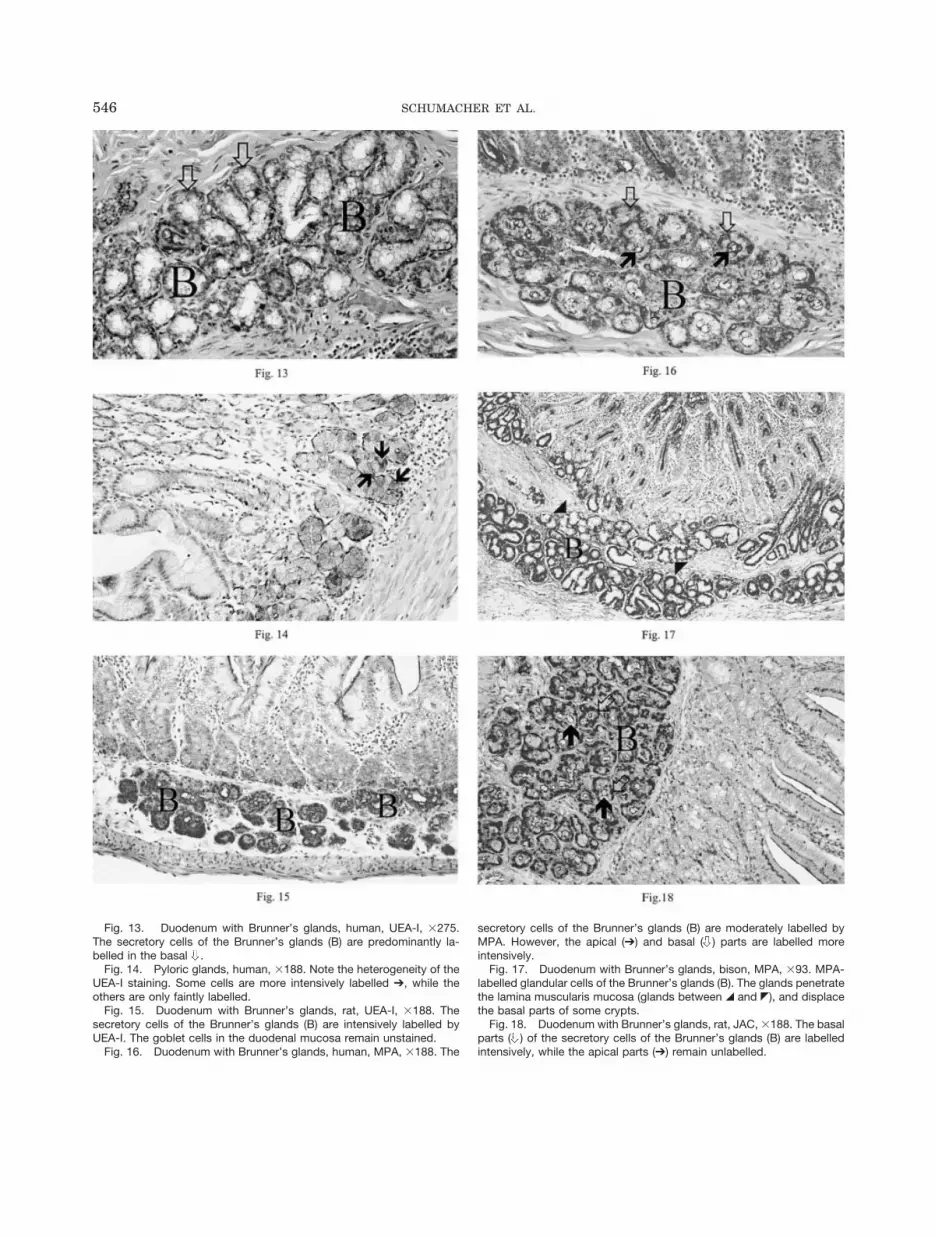
Fig. 13. Duodenum with Brunner’s glands, human, UEA-I, �275.The secretory cells of the Brunner’s glands (B) are predominantly la-belled in the basal .
Fig. 14. Pyloric glands, human, �188. Note the heterogeneity of theUEA-I staining. Some cells are more intensively labelled ➔, while theothers are only faintly labelled.
Fig. 15. Duodenum with Brunner’s glands, rat, UEA-I, �188. Thesecretory cells of the Brunner’s glands (B) are intensively labelled byUEA-I. The goblet cells in the duodenal mucosa remain unstained.
Fig. 16. Duodenum with Brunner’s glands, human, MPA, �188. The
secretory cells of the Brunner’s glands (B) are moderately labelled byMPA. However, the apical (➔) and basal ( ) parts are labelled moreintensively.
Fig. 17. Duodenum with Brunner’s glands, bison, MPA, �93. MPA-labelled glandular cells of the Brunner’s glands (B). The glands penetratethe lamina muscularis mucosa (glands between 7 and 6), and displacethe basal parts of some crypts.
Fig. 18. Duodenum with Brunner’s glands, rat, JAC, �188. The basalparts ( ) of the secretory cells of the Brunner’s glands (B) are labelledintensively, while the apical parts (➔) remain unlabelled.
546 SCHUMACHER ET AL.

Fig. 19. Duodenum with Brunner’s glands, raccoon, JAC, �188. Ex-cept for small granules within the secretory cells of the Brunner’s glands (B),they remain unstained. Note the labelling of the brush border (1) by JAC.
Fig. 20. Duodenum with Brunner’s glands, rat, PNA, �188. Note theintense labelling of the basal part ( ) of the secretory epithelia of Brun-ner’s glands (B) with PNA, while the apical parts (➔) remain unlabelled.
Fig. 21. Duodenum with Brunner’s glands, human, WGA, �188.Note that both the basal ( ) and apical (➔) parts of the secretory cells ofthe Brunner’s glands (B) are most intensively demarcated by WGA. Basalcells of the crypts, which presumably represent Paneth cells (1), arealso labelled.
Fig. 22. Duodenum with Brunner’s glands, rat, VVA, �93. The basalparts ( ) of the secretory cells of the Brunner’s glands (B) are labelledintensively, while the apical parts (➔) remain unlabelled.
Fig. 23. Pyloric glands, rat, VVA, �275. Only a supranuclear field( ➔), which presumably represents the Golgi apparatus, is labelled by
➔
VVA.Fig. 24. Duodenum with Brunner’s glands, human, MPA, �188. The
staining pattern of the glandular cells of the Brunner’s glands (B) is againbasal-apical. Note the penetration of Brunner’s gland cells into the basalpart of two crypts (C) (glands between ‹ and Š).
547BRUNNER’S GLAND MUCINS

In addition to reacting with PNA, the Brunner’s glandsalso reacted with all of the lectins (Con A, WGA, HPA,VVA, MPA, JAC, SNA, and UEA-I) that reacted with thepyloric glands.
Cotton-tailed and domestic rabbits (Fig. 9).Inthe stomach of both cotton-tailed and domestic rabbits,mucous surface cells stained only with PAS, while pyloricglands stained with PAS and Alcian blue at both pH 1.0and 2.5. In the duodenum, goblet cells of both speciesstained with both PAS and Alcian blue (pH 1.0 and 2.5).The Brunner’s glands of the cotton-tailed rabbit reactedmore intensely with PAS than those of the domestic rab-bit. Alcian blue (pH 1.0 and 2.5) stained Brunner’s glandsof both species with the same intensity.
The gastroduodenal junctions in both species were sim-ilar in terms of lectin histochemistry. Mucous surface cellsreacted with only one lectin (Con A), while pyloric glandsreacted with eight lectins (Con A, WGA, HPA, VVA, MPA,JAC, PNA, and SNA). In the duodenum, goblet cells re-acted with four lectins (HPA, VVA, MPA, and JAC), whileBrunner’s glands reacted with six lectins (Con A, WGA,HPA, MPA, JAC, and SNA). A few Brunner’s gland ducts,located in the submucosa, reacted with LTA and UEA-I.
Cat (Fig. 10).The duodenum of the cat had a relativelythin muscular layer, and no plicae circularis.
In the stomach, mucous surface cells stained with PASand Alcian blue at pH 2.5, while pyloric glands stainedonly with PAS. In the duodenum, goblet cells stained withboth PAS and Alcian blue (pH 1.0 and 2.5), while Brun-ner’s glands stained with PAS only.
The mucous surface cells of the stomach reacted withonly one lectin (HPA), which also reacted with all othermucus-producing cells of both the stomach and the duo-denum. Pyloric glands reacted with WGA, HPA, VVA,JAC, and PNA. In the duodenum, goblet cells reacted withWGA, HPA, VVA, MPA, JAC, LTA, and UEA-I, whileBrunner’s glands reacted with Con A (Fig. 4), WGA, HPA,VVA, MPA, JAC, PNA, LTA, and UEA-I.
Raccoon (Fig. 11).The acini of Brunner’s glands inthe raccoon were relatively dilated in comparison withthese of the other mammals.
In the stomach, mucous surface cells stained with bothPAS and Alcian blue at both pH 1.0 and 2.5, while pyloricglands stained only with PAS. In the duodenum, gobletcells stained with PAS and Alcian blue at both pH 1.0 and2.5, while Brunner’s glands stained only with PAS.
In the stomach, mucous surface cells reacted with WGA,MPA, JAC, PNA, and LTA, while pyloric glands reactedwith Con A, WGA, MPA, JAC (Fig. 19), LTA, and UEA-I.In the duodenum, goblet cells reacted with GNA, MPA,JAC, LTA, and UEA-I. In addition to reacting with HPAand VVA, Brunner’s glands reacted with all of the lectinsthat reacted with the pyloric glands (Con A, WGA, MPA,JAC, LTA, and UEA-I).
Opossum (Fig. 12).The histology of the gastroduode-nal junction of the opossum differed from that describedabove in that the Brunner’s glands were concentrated in avery short portion of the submucosa, extending from thedistal part of the pyloric region of the stomach to theproximal duodenum.
Mucous surface cells, pyloric glands, and Brunner’sglands all stained with PAS but did not stain with Alcian
blue at either pH 1.0 or 2.5, whereas goblet cells stainedwith both PAS and Alcian blue (pH 1.0 and 2.5).
In the stomach, mucous surface cells reacted with ConA, WGA, MPA, JAC, PNA, LTA, and UEA-I, and pyloricglands reacted with Con A, WGA, HPA, MPA, JAC, andPNA. In the duodenum, goblet cells reacted only with PNAand UEA-I, while Brunner’s glands reacted with sevenlectins (Con A, WGA, HPA, MPA, JAC, LTA, and UEA-I).
DISCUSSIONThe general morphology of the duodenum of all species
examined in this study was in accordance with that de-scribed for mammals in general. However, in a few speciesthe distribution and morphology of the Brunner’s glandsdeviated from the general pattern. In the South Americanopossum, as has also been reported for Didelphis virginia(Schumacher and Krause, 1995), the Brunner’s glandswere confined to a very narrow region, immediately distalto the pyloric sphincter. In comparison with the othermammals examined, the secretory units of Brunner’sglands in the raccoon were more dilated and lined withflatter cells. Similar observations regarding the luminalsize of Brunner’s glands in the mouse have been reported(Krause, 1981).
In classic carbohydrate histochemistry, a positive PASreaction indicates the presence of neutral carbohydrates,while positive Alcian blue reactions at pH 1.0 and 2.5indicate the presence of acidic sulphated and acidic car-boxylated residues, respectively (Spicer and Schulte,1992). In the present study, the results of classic carbohy-drate histochemistry demonstrated variation in the stain-ing properties of the different mucus-producing cell typesat the gastroduodenal junction in most of the mammalsinvestigated. In addition, the results showed that the gob-let cells in all of the mammals investigated containedacidic carboxylated and acidic sulphated mucins. Theseobservations suggest that the mucins secreted by gobletcells have been conserved with regard to their chargeduring the evolution of mammals (Ota et al., 1998). Thesefindings are in agreement with studies of single mamma-lian species, such as that by Poddar and Jakob (1979),which showed that duodenal goblet cells in the ferretcontain both sialomucins and sulphated mucosubstances.However, they disagree with the results of Filipe andFenger (1979), who observed sialomucins but no sulphatedmucins in the goblet cells of the human duodenum, jeju-num, and ileum. This difference in observation could bedue to the use of different materials or to differences in themethods employed. Our sections were directly cut fromthe gastroduodenal junction, whereas Filipe and Fenger(1979) may have used material from a more distal region,which might account for the difference. In addition, weused a strictly tested Alcian blue, because a great diver-sity in staining results has been observed with the use ofdifferent batches of Alcian blue (Schumacher and Adam,1994). Since these methodological differences were un-known at the time of Filipe and Fenger’s publication(1979), we assume that this is the most likely cause of thedifferences between their study and ours.
A rather striking contrast is seen in the staining resultsof Alcian blue at pH 2.5 and lectin histochemistry withSNA-I and MAA. However, this contrast is not surprising,because the Alcian blue reaction at pH 2.5 is very generaland nonspecific, and at this pH, Alcian blue will stain allacidic carboxylated residues. Thus, the goblet cells of all of
548 SCHUMACHER ET AL.

the mammals, and the mucus surface cells, pyloric glands,and Brunner’s glands of half of the investigated mammalswere labelled. On the other hand, the lectins SNA-I andMAA are extremely specific: SNA-I binds only toNeu5Ac�2,6Gal (Shibuya et al., 1987), and MAA bindsonly to Neu5Ac�2,3Gal (Alvarez et al., 1999). In a study oflymphoid organs in chickens, Jorns et al. (2003) differen-tiated between these two linkages by showing that hardlyany structures in the spleen reacted with MAA, whileSNA-I labelled the endothelium, lymphocytes in the PALSand PELS, and cells with dendritic morphology in the redpulp. Because of the extreme specificity of these two lec-tins, it is not surprising that major differences in theSNA-I and MAA labelling patterns were found, and thatthese patterns also differ significantly from that obtainedwith Alcian blue at pH 2.5.
Despite their similar morphological appearance in theH&E sections, the PAS and Alcian blue (pH 1.0 and 2.5)staining properties of Brunner’s glands showed markeddifferences in our series of mammals. In bison, deer,guinea pigs, voles, and cotton-tailed and domestic rabbits,they contain acidic sulphated and carboxylated mucins,whereas in humans, rhesus and Japanese macaques, cats,raccoons, rats, and opossums, they contain neutral mu-cins. This variation could not be attributed to either theorder or the diet of the mammals. For example, althoughrats and voles are both rodents, Brunner’s glands in therat produce only neutral mucins, while those in the voleproduce acidic mucins. This observation is in agreementwith earlier studies on Brunner’s glands in 27 species ofbats that differed markedly in diet (insect-, fish-, blood-,nectar-, plant-, or animal-feeding species) but producedonly neutral mucins (Forman, 1971).
The lectins used in this study were typical of the termi-nal carbohydrate residues found in animal cells. MPA andJAC (galactose-specific lectins) reacted with Brunner’sglands in all of the mammals investigated. The carbohy-drate residue is typical of O-linked mucus-type glycopro-teins (Strous and Dekker, 1992), and thus its presence isnot surprising. In contrast, PHA-L, a bi- and tri-anten-nary complex-type carbohydrate-specific lectin, reactedonly with the mucin granules of some of the mucus-pro-ducing cells at the gastroduodenal junction in the rodents,rats, voles, and guinea pigs. The presence of complex-typecarbohydrates in mucins is rather unusual; however, thisrather unusual histochemical reaction pattern of rat gas-tric mucin has been reported previously (Ota et al., 1991).Our findings, therefore, appear to emphasize the special-isation of rodent mucins with respect to the presence ofcomplex-type carbohydrates.
The lectin histochemical results of the duodenumshowed that the mucins from one mammalian speciesgenerally differ from those of another species, providingfurther evidence for the diversity of terminal carbohydrateresidues in animal cells. Despite this interspecies diver-sity, a comparison of the lectin-binding profiles of mucus-producing cells in the stomach with those in the duode-num reveals that Brunner’s glands in one species are moreclosely related to the pyloric glands than to the mucoussurface epithelium of the stomach or to goblet cells of theduodenum. In addition, both Brunner’s gland epitheliaand pyloric glands have a greater carbohydrate diversitythan mucous surface cells of the stomach and goblet cellsin the duodenum, except in the cat. The significance of thedeviation from the general pattern seen in the cat is un-
known. The lectin histochemical observations again em-phasise the fact that mucins of the Brunner’s and pyloricglands are more closely related to each other than to anyother mucin configuration.
It is perhaps not surprising that the carbohydrate res-idues of the Brunner’s glands resembled those of the py-loric glands, because the content of the lumen in thepyloric region of the stomach is similar to that of theproximal duodenum with regard to potential epithelium-damaging factors. In addition, both mucins are secretedfrom a glandular lumen that secondarily opens onto thesurface of the duodenum or the stomach, respectively,while goblet cells and mucous surface cells secrete directlyinto the lumen of the gastrointestinal tract. The greaterdiversity of the carbohydrate residues in the mucins ofBrunner’s and pyloric glands as compared to goblet andmucous surface cells may simply reflect a different phys-iological property of these mucins, which are modified bythe carbohydrate side chains on the mucin protein back-bone.
It has long been recognised that following chronic ulcer-ation of the intestine, new glandular structures developadjacent to the lesion (Kawel and Tesluk, 1955; Lee, 1964;Kushima et al., 1999). Recent studies have shown that
Fig. 25. Duodenum with Brunner’s glands, rat, �188. The secretorycells of the Brunner’s glands (B) are basally ( ) stained by HPA.
Fig. 26. Pyloric glands, rat, HPA, �275. Note that HPA labels theperinuclear zone of the gland cells (➔).
549BRUNNER’S GLAND MUCINS

these glands develop in a manner very similar to that ofBrunner’s glands, and, like Brunner’s glands, they secreteepidermal growth factor in addition to mucus (Ahnen etal., 1994). From our comparative observations, it appearsthat mucus from Brunner’s glands is more heterogeneousthan the other mucins. This diversity could aid in theadaptability of the mucus to various activities, includingwound-healing and protecting areas adjacent to an ulcer.Extensive lectin-binding studies have not yet been per-formed on these pathologic glands; however, the currentstudy indicates that the carbohydrate residues of the mu-cins secreted by these glands may be as diverse as those ofthe pyloric and Brunner’s glands. Thus, in mammals,glands appear to have evolved on both sides of the gastro-intestinal junction. These glands produce similar mucinsthat are involved primarily in protecting the mucosa fromthe erosive effects of luminal contents in this region.
LITERATURE CITEDAhnen DJ, Poulsom R, Stamp GWH, Elia G, Pike C, Jeffery R,
Longcroft J, Rio M-C, Chambon P, Wright NA. 1994. The ulcer-ation-associated cell lineage (UACL) reiterates the Brunner’sglands differentiation programme but acquires the proliferativeorganisation of the gastric glands. J Pathol 173:317–326.
Allen A. 1981. Structure and function of gastrointestinal mucus. In:Johnson L-R, editor. Physiology of the gastrointestinal tract. NewYork: Raven. p 617–639.
Alvarez G, Lascurain R, Perez A, Degand P, Montano LF, Martinez-Cairo S, Zenteno E. 1999. Relevance of sialoglycoconjugates inmurine thymocytes during maturation and selection in the thymus.Immunol Invest 28:9–18.
Brooks SA, Leathem AJC, Schumacher U. 1997. Lectin histochemis-try, a concise practical handbook. Oxford: BIOS Scientific Publish-ers.
Filipe MI, Fenger C. 1979. Histochemical characteristics of mucins inthe small intestine. A comparative study of normal mucosa, benignepithelial tumours and carcinoma. Histochem J 11:277–287.
Forman GL. 1971. Histochemical differences in gastric mucus of bats.J Mammal 52:191–193.
Jorns J, Mangold U, Neumann U, Van Damme EJM, Peumans WJ,Pfuller U, Schumacher U. 2003. Lectin histochemistry of the lym-phoid organs of the chicken. Anat Embryol 207:85–94.
Kawel CA, Tesluk H. 1955. Brunner-type glands in regional enteritis.Gastroenterol 28:210–221.
Krause WJ. 1981. Morphological and histochemical observations onthe duodenal glands of eight wild ungulate species native to NorthAmerica. J Anat 162:167–181.
Krause WJ. 1988. Biology of duodenal (Brunner’s) glands. In: MottaPM, Fujita H, editors. Ultrastructure of the digestive tract. Boston:Martinius Nijhoff Publishing. p 67–84.
Krause WJ, Steven R, Palmiter P. 1988. The effect of surgical ablationof Brunner’s glands on the proximal duodenal mucosa. Anat Anz166:111–115.
Kushima R, Manabe R, Hattori T, Borchard F. 1999. Histogenesis ofgastric foveolar metaplasia following duodenal ulcer: a definite re-parative lineage of Brunner’s gland. Histopathology 35:38–43.
Lee F. 1964. Pyloric metaplasia in the small intestine. J PatholBacteriol 87:267–279.
Neutra MR, Forstner JF. 1987. Gastrointestinal mucus: synthesis,secretions, and function. In: Johnson LR, editor. Physiology of thegastrointestinal tract. 2nd ed. New York: Raven Press. p 975–1009.
Ota H, Katsuyama T, Ishii K, Nakayama I, Shiozawa T, Tsukahara Y.1991. A dual staining method for identifying mucins of differentgastric epithelial mucous cells. Histochem J 23:22–28.
Ota H, Nakayama J, Momose M, Kurihara M, Ishihara K, Hotta K,Katsuyama T. 1998. New monoclonal antibodies against gastricgland mucous cell-type mucins: a comparative immunohistochemi-cal study. Histochem Cell Biol 110:113–119.
Poddar S, Jakob S. 1979. Mucosubstance histochemistry of Brunner’sglands, pyloric glands and duodenal goblet cells in the ferret. His-tochemistry 65:459–472.
Schumacher U, Adam E. 1994. Standardization of staining in glycos-aminoglycan histochemistry: Alcian blue, its analogues, and dia-mine methods. Biotechnol Histochem 69:18–24.
Schumacher U, Krause WJ. 1995. Molecular anatomy of an endoder-mal gland: investigations on mucus glycoproteins and cell turnoverin Brunner’s glands of Didelphis virginiana using lectins and PCNAimmunoreactivity. Cell Biochem 58:56–64.
Shibuya N, Goldstein IJ, Broekaert WF, Nsimba-Lubaki M, PeetersB, Peumans WJ. 1987. The elderberry (Sambucus nigra L.) barklectin recognizes the Neu5Ac(alpha 2-6)Gal/GalNAc sequence.J Biol Chem 262:1596–1601.
Spicer SS, Schulte BA. 1992. Diversity of cell glycoconjugates shownhistochemically: a perspective. J Histochem Cytochem 40:1–38.
Strous GJ, Dekker J. 1992. Mucin-type glycoproteins. Crit Rev Bio-chem Mol Biol 27:57–92.
550 SCHUMACHER ET AL.





























