Embed Size (px)
Citation preview

Address correspondence to: Dr Jacopo Moggi-Cecchi, Laboratori di Antropologia, Dipartimento di BiologiaAnimale e Genetica, Universita di Firenze, via del Proconsolo, 12, 50122 Florence, Italy. E-mail: [email protected]
Deano D. StynderDepartment of HumanBiology, University ofCape Town Medical School,Observatory 7925, Capetown,South Africa. E-mail:[email protected]
Jacopo Moggi-CecchiLaboratori di Antropologia,Dipartimento di BiologiaAnimale e Genetica,Universita di Firenze,Florence, Italy. E-mail:[email protected] Palaeo-anthropology Unitfor Research and Exploration,Bernard Price Institute forPalaeontological Research,University of theWitwatersrand, Johannesburg,South Africa
Lee R. BergerPalaeo-anthropology Unit forResearch and ExplorationBernard Price Institute forPalaeontological Research,University of theWitwatersrand, Private Bag3, Wits 2050, Johannesburg,South Africa. E-mail:[email protected]
John E. ParkingtonDepartment of Archaeology,University of Cape Town,Private Bag, Rondebosch7701, Capetown,South Africa. E-mail:[email protected]
Received 30 October 2000Revision received16 March 2001 andaccepted 23 April 2001
Keywords: mandibularincisors, subadult,South Africa, late MiddlePleistocene,hyena accumulation.
Human mandibular incisors from the lateMiddle Pleistocene locality ofHoedjiespunt 1, South Africa
The Hoedjiespunt 1 locality is an archaeological and palaeontologicalsite located on the Hoedjiespunt Peninsula at Saldanha Bay, SouthAfrica. In 1996 two human teeth, a left central mandibular incisorand a left lateral mandibular incisor, were discovered during excava-tions in the late Middle Pleistocene palaeontological layers. Theseteeth are described and are found to belong to a single subadultindividual. Despite their developmental stage, these incisors alreadydisplay early signs of wear. Their crown diameters are larger thanmodern and archaeological African comparative material and aremost closely comparable with crown diameters of an early MiddlePleistocene and late Middle Pleistocene dental sample from Africa,Europe and Asia. In the light of this metrical evidence, data on twopreviously excavated maxillary molars, most probably belongingto the same individual, were re-examined. It was found that theHoedjiespunt 1 hominid possessed dental metrical features (largeanterior teeth and small molars) comparable with other African andEuropean hominids referred to the Middle Pleistocene.
� 2001 Academic Press
Journal of Human Evolution (2001) 41, 369–383doi:10.1006/jhev.2001.0488Available online at http://www.idealibrary.com on
0047–2484/01/100369+15$35.00/0 � 2001 Academic Press

370 . . ET AL.
Hoedjiespunt
SaldanhaBay
Cape Town
1: 1 500 000N
Cape Town
South Africa
Johannesburg
Botswana
Namibia
AtlanticOcean
Saldanha
Sea Harvest
North BayHDP1
Saldanha Bay
Ore
ter
min
al
0 1 2
km
N
Figure 1. Position of the Hoedjiespunt peninsula and the sites of Hoedjiespunt 1 and Sea Harvest.
Introduction
Hoedjiespunt 1 (HDP1) (33�01�45�S,17�57�37�E) is located on the southern edgeof the Hoedjiespunt Peninsula, at SaldanhaBay on the West Coast of South Africa(Figure 1). The peninsula, which projectseastwards from the northwest corner ofSaldanha Bay, is part of a large fossil dunelandscape that houses several archaeologicaland palaeontological sites. Among these isthe important Late Pleistocene site of Sea
Harvest, which, in addition to yielding anextensive mammalian fauna (Klein, 1983),also yielded human skeletal material inthe form of a manual distal phalanx and amaxillary premolar (Grine & Klein, 1993).
Fossils from the Hoedjiespunt Peninsulawere first reported by Klein (1983), who,together with scientists from the SouthAfrican museum, collected bones as theyweathered out of a sandstone cliff face,about 10 m from the present day site ofHDP1. Subsequently this cliff had been
371J
bevelled, thus exposing further archaeo-logical and palaeontological material. In1993, this abundance of exposed materialprompted scientists and students from theUniversity of Cape Town and the Universityof the Witwatersrand to carry out a limitedsurface survey of fossil material on thepeninsula and surrounding area. The dis-covery of several fragments of a human leftmaxillary second molar (HDP1-1) in thecourse of this survey preceded the firstsystematic excavation of HDP1 in 1994.Excavations yielded an additional humantooth, a right maxillary third molar(HDP1-2, field no. 49). According to Berger& Parkington (1995) these molars probablydate from the late Middle Pleistocene andbelong to a single individual that died in itsearly teens.
Subsequent excavations in 1996 have ledto the discovery of two additional humanteeth—a left central mandibular incisor(HDP1-3, field no. 937) and a left lateralmandibular incisor (HDP1-4, field no.3150). A human tibia was recovered in 1998and has recently been described (Churchillet al., 2000). In this paper, we present a
description and analysis of the two incisors.Their possible relation to the molars is alsoinvestigated.
The Hoedjiespunt 1 locality
Shell midden
Dark material (DAMA)
Snail sands
Shelly sands
Consolidated material
Hominid sands (HOMS)
Lower coarse shelly sands
Fine shelly sands
HOMS, the layer thatproduced the hominidteeth.
1 meter
N
Calcrete lip
Calcreteboulders
Road
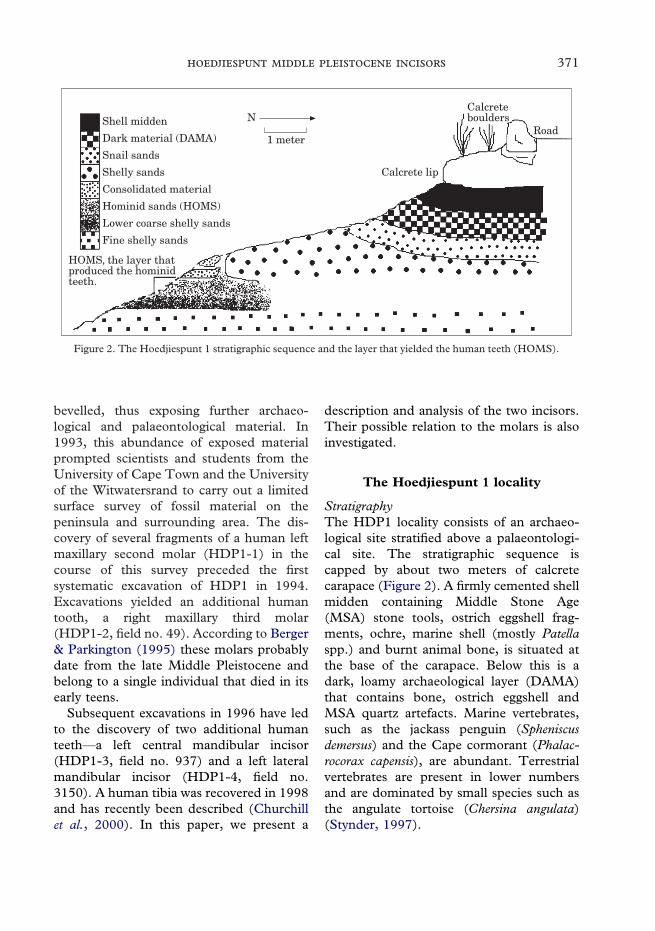
Figure 2. The Hoedjiespunt 1 stratigraphic sequence and the layer that yielded the human teeth (HOMS).
StratigraphyThe HDP1 locality consists of an archaeo-logical site stratified above a palaeontologi-cal site. The stratigraphic sequence iscapped by about two meters of calcretecarapace (Figure 2). A firmly cemented shellmidden containing Middle Stone Age(MSA) stone tools, ostrich eggshell frag-ments, ochre, marine shell (mostly Patellaspp.) and burnt animal bone, is situated atthe base of the carapace. Below this is adark, loamy archaeological layer (DAMA)that contains bone, ostrich eggshell andMSA quartz artefacts. Marine vertebrates,such as the jackass penguin (Spheniscusdemersus) and the Cape cormorant (Phalac-rocorax capensis), are abundant. Terrestrialvertebrates are present in lower numbersand are dominated by small species such asthe angulate tortoise (Chersina angulata)(Stynder, 1997).

372 . . ET AL.
Two shelly sand bodies largely sterile ofmammalian fossils separate the archaeologi-cal site from the palaeontological site below.The main bone-bearing layer of the palaeon-tological site (HOMS) is densely packedwith bone. It is sandwiched between a shelfof consolidated material above and two lessdensely packed bone-bearing layers below.Grazing ungulates such as the black wilde-beest (Connochaetes gnou) and red hartebeest(Alcelaphus buselaphus), and carnivores suchas the black-backed jackal (Canis mesomelas),dominate this assemblage (Stynder, 1997).Marine animals are rare. The lack of anyartefacts or archaeological features in thedeposit, as well as the presence of a con-siderable number of hyena coprolites andgnawed bones, suggest that hyenas, mostprobably the brown hyena (Hyaenabrunnea), accumulated these faunal remains.
AgeThe presence of informal MSA stone tools isconsistent with an age of between 40 ka and240 ka for the archaeological site. Infraredstimulated luminescence (IRSL) and ther-moluminescence (TL) dates on sedimentsfrom DAMA place the age of this horizonat ca. 117 ka, sometime during the LastInterglacial (isotope stage 5e) (Woodborne,2000). The abundance of shellfish andmarine vertebrate remains in these layers isconsistent with accumulation during theLast Interglacial.
The age of the palaeontological site is lesssecure. IRSL and TL dates on sedimentsfrom HOMS suggest accumulation duringthe Middle Pleistocene, with a maximumage of ca. 550 ka (Woodborne, 2000).Analysis of the foraminiferal assemblagefrom HOMS indicates an age of betweenca. 180 ka and ca. 480 ka (McMillan, per-sonal communication). The fauna, most ofwhich derive from the HOMS layer, is typi-cal of the Florisbad Faunal Span. Thisplaces a maximum age of around 250 ka(Grun et al., 1996) on the assemblage.
Based on these dates and our stratigraphicobservations, we suggest that the palaeonto-logical site was most probably depositedbetween 200 ka and 300 ka ago, sometimeduring the late Middle Pleistocene.
Location of teethAll the teeth (HDP1-3 and HDP1-4 as wellas HDP1-1 and HDP1-2) were found in situin the HOMS layer of the palaeontologicalsite. All four teeth were situated in a rela-tively small area of about 1·5 m in length by1 m in breadth. The maximum difference indepth between the highest and lowest toothwas approximately 200 mm.
Anatomical descriptions
HDP1-3 LI1 (Figure 3). This is a complete,well-preserved, permanent left central man-dibular incisor. The corrected mesiodistal(MD) diameter of the crown is 6·4 mm andthe buccolingual (BL) diameter is 7·2 mm.Crown height is ca. 8·1 mm and root lengthis 15·1 mm. Wear is marked with a broadrim of dentine exposed along the incisaledge [Figure 3(a)]. The distal interproximalwear facet (IWF) is placed slightly lingual tothe midline. A clearly defined mesial IWF isnot evident. The crown outline, as seen inlingual view, is asymmetrical, with the weakcervical eminence skewed distally. Very faintmesial and distal marginal ridges create afaint shovelling [Figure 3(b)]. Enamel pol-ishing is evident along its entire length. Thelabial face is slightly convex inciso-cervically(IC). Numerous scratches are evident on theentire surface. An irregular band of hypo-plastic enamel is present on the incisal thirdof the face [Figure 3(c)]. Remnants of den-tal calculus are evident on the cervical thirdof the crown. The root is long and thin, MDcompressed, tapering inferiorly.
HDP1-4 LI2 (Figure 4). This is a com-plete, well-preserved, permanent left lateralmandibular incisor. The corrected MDdiameter of the crown is 7·7 mm and the BL

373J
diameter is 7·2 mm. Crown height is10·3 mm and root length ca. 13·5. Wear ismarked by a subtriangular rim of dentineexposed on the incisal edge, tapering distally[Figure 4(a)]. No dentine is exposed on thedisto-incisal corner. The mesial IWF has arough surface and it is labially placed to themidline. No clear distal IWF is evident. Thecrown outline, as seen in lingual view, issubtriangular, with the mesio-incisal edgereduced by wear. Marked mesial and distalmarginal ridges merge at the level of the veryfaint cervical eminence [Figure 4(b)]. Onthe labial face, many tiny scratches arepresent, especially near the incisal edge. Alarge area of hypoplastic enamel is presenton the disto-incisal edge of the crown,extending onto the distal face [Figure 4(c)].Smaller areas of enamel hypoplasia are
evident on the mesio-labial edge, near theincisal margin, and on the cervical third ofthe crown. The root is long, strongly com-pressed MD with marked longitudinalgrooves. The root apex is still open, suggest-ing that root development was not yet com-plete [Figure 4(d)].
Figure 3. (a) Incisal edge, (b) lingual view and (c) labialview of HDP1-3, a left central mandibular incisor. Thescale is in centimetres.
Figure 4. (a) Incisal edge, (b) lingual view, (c) labialview and (d) root apex of HDP1-4, a left lateralmandibular incisor. The scale is in centimetres.
Discussion
There is a strong possibility that HDP1-3and HDP1-4 derive from a single individual.Both incisors display similar degrees of wearon their respective incisal edges. There arealso matching interproximal wear facets,situated distolingually on HDP1-3 and

374 . . ET AL.
Figure 5. The Hoedjiespunt 1 molars: (a) occlusal and(b) mesial view of HDP1-1, a left maxillarysecond molar; (c) occlusal and (d) buccal view ofHDP1-2, a right maxillary third molar. The scale is incentimetres.
mesiolabially on HDP1-4. Based on therespective positions of these wear facets oneach tooth, there appears to have been somedegree of crowding in the anterior teeth ofthe mouth(s) from which they derive.
If these teeth do derive from a singleindividual, it is possible, on the basis ofdental development, to give an estimate ofthe age at death of this individual. In thisrespect, only HDP1-4 can be used, since itstill has the root apex open. Following Smith(1991), its stage of development is scored atRc—A 1
2(root complete—apex half open)
resulting in a predicted mean age of between8 and 9 years at death, with a range of 6·5and 10·5 years if we take into account agevariation of formation stages (Moorreeset al., 1963).
The existence of two human molars fromthe site (HDP1-1 with HDP1-2) (Berger &Parkington, 1995) raised the question ofwhether the two sets of teeth derive fromthe same individual. Since HDP1-1 andHDP1-2 were found in the same strati-graphic level as HDP1-3 and HDP1-4, andalso belonged to a subadult individual(Figure 5—see also Figures 2 and 3 inBerger & Parkington, 1995), this was adistinct possibility. In order to address thisissue, the developmental stages of the twomolars have also been assessed. The samestandards as those applied to the incisorswere followed here with an acknowl-edgement that those standards relate tomandibular teeth.
The root of HDP1-1, a LM2, reflects alevel of development comparable to stageA1
2, resulting in a predicted age of approxi-
mately 14 years (range 10–15). The root ofHDP1-2, a RM3, is at a developmentalstage equivalent to stage R1
4to R1
2, resulting
in an age estimate of 15 years (range12–18). Thus on the basis of modernhuman standards of dental development,there appears to be a marked difference inage estimates between the incisor and themolars.
However, some caution is required wheninterpreting this evidence. First, most stan-dards of dental development are derivedfrom modern European populations. This isnot an ideal situation, as marked variabilityexists among living populations in terms oftooth formation (Tompkins, 1996a and ref-erences therein). For example, Tompkins(1996a) has demonstrated that molar teeth(in particular the M3) in African popula-tions display advanced relative developmentcompared with Europeans. Similar conclu-sions were reached by Dean et al. (1993) intheir analysis of incremental markings in theenamel and the dentine of a West Africanindividual. It has been suggested that thisvariability may be due to population differ-ences in the timing of dental and skeletaldevelopment (Tompkins, 1996a). Second,it is well established that the third molar isvery variable in its development (Garn et al.,1962; Anderson et al., 1976).

375J
The scant data presently available on thedental development of Pleistocene hominidsprovide additional information. Bermudezde Castro et al., (1999b) described an essen-tially modern human pattern of dentaldevelopment in the Lower Pleistocenehominids from the site of Atapuerca TD6.However, they also found that the thirdmolar teeth displayed a pattern of advancedrelative development compared to modernpopulations. An advanced relative develop-ment of the upper and lower third molarshas also been described in a Middle Pleisto-cene specimen from Atapuerca-Sima de losHuesos (Bermudez de Castro & Rosas,2001). In addition to these data, Tompkins(1996b), in a study of relative dental devel-opment in Upper Pleistocene hominids(Neanderthals/archaic Homo sapiens andEarly Modern/Upper Paleolithic), has dem-onstrated an advanced relative developmentof M2 and M3 in these fossil groups. At thesame time, there also appeared to havebeen a relative delay in central incisor devel-opment compared with modern humanpopulations.
Given this evidence, the observed discrep-ancy in age estimates between the HDP1lateral incisor and the molars may well berelated to a difference in the dental develop-mental patterns of this specific populationand that of modern humans. Thus it seemsreasonable to suggest that the two molars,HDP1-1 and HDP1-2, and the two incisors,HDP1-3 and HDP1-4, belonged to thesame individual.
One of the most remarkable features ofHDP1-4 is the simultaneous presence ofa developing root and of marked incisalattrition comparable to wear stage 4 inthe Murphy system [as modified bySmith (1984)] [Figure 4(a)]. A similarstage of wear is also evident on HDP1-3[Figure 3(a)]. Enamel polishing, as well asnumerous scratch marks more commonlyassociated with the incisal edge occur on thelabial surfaces of both teeth. Heavy wear of
the anterior teeth with a tendency for thedevelopment of labial rounding, is oftenseen in people pursuing a hunter-gathererlifestyle, and has been attributed to thehabitual nonmasticatory utilization ofanterior teeth (Hinton, 1981). It is possiblethat the HDP1 incisors are displaying theearly stages of habitual nonmasticatoryutilization. To shed light on this issue, adetailed analysis of the microwear patternson these teeth is presently underway.
A comparative analysis of the metricalfeatures of the HDP1 incisors providesadditional information. Tables 1 and 2report the MD and BL diameters of centraland lateral mandibular incisors respectively,in selected recent, archaeological and fossilhuman specimens. Crown shape index (CSI=BL/MD*100) values are also provided.
The MD and BL diameters of HDP1-3are larger than the mean values of a sampleof recent and archaeological African centralincisors. The MD diameter of HDP1-3is within the range of Homo ergaster(KNM-WT 15000 and KNM-ER 820) andHomo erectus (the Zhoukoudian sampleand Sangiran 11), and only slightly smallerthan Homo habilis (OH 7 and OH 16). TheBL diameter of HDP1-3 is exceeded onlyby European specimens from FontanaRanuccio (an approximate value by Segre& Ascenzi, 1984) and Atapuerca-TD6-52,the latter attributed to the species Homoantecessor by Bermudez de Castro et al.(1999a), and with a suggested age of greaterthan 780 ka.
The MD and BL diameters of HDP1-4are larger than the mean values of a sampleof modern and archaeological African lateralincisors. The MD diameter of HDP1-4 isclose to the values of H. habilis and amongthe largest in the sample of Early and Mid-dle Pleistocene hominid lateral incisors fromAfrica, Europe and Asia. The BL diameterof HDP1-4 is larger than the mean value forthe H. erectus specimens from Zhoukoudian,as well as the individual dimensions of the

376 . . ET AL.
Tab
le1
Mes
iod
ista
l(co
rrec
ted
)an
db
ucc
olin
gual
dia
met
ers
(in
mm
)an
dcr
own
shap
ein
dex
(CS
I)of
HD
P1-
3an
dof
cen
tral
man
dib
ula
rin
ciso
rsin
sele
cted
rece
nt,
arch
aeol
ogic
alan
dfo
ssil
hu
man
sfr
omA
fric
a,E
uro
pe
and
Asi
a
nM
DS
.D.
nB
LS
.D.
Cro
wn
heig
htR
oot
leng
thC
SI
Ref
eren
ces
HD
P1-
36·
47·
2(8
·1)
wor
n15
·111
2·5
Per
sona
lda
taO
H7
6·8
6·6
9·2
97·1
Tob
ias
(199
1)O
H16
6·5
710
·610
7·7
Tob
ias
(199
1)S
KX
2355
6—
Per
sona
lda
taK
NM
-WT
-150
006·
66·
810
·119
·910
3·0
Bro
wn
&W
alke
r(1
993)
KN
M-E
R-8
206·
16·
310
3·3
Woo
d(1
991)
Dm
anis
i6·
25·
995
·2G
abun
ia&
Vek
ua(1
995)
San
gira
n11
7·4
6·5
87·8
Woo
d(1
991)
San
gira
nB
s97
066·
05·
89·
996
·7B
aba
etal
.(2
000)
Zho
ukou
dian
56·
30·
265
6·4
0·42
101·
6W
ood
(199
1)T
erni
fine
5·7
6·6
9·0
115·
8B
erm
udez
deC
astr
o(1
986)
Rab
at6
711
6·7
Ber
mud
ezde
Cas
tro
(198
6)A
tapu
erca
-TD
AT
D6-
52—
7·6
Ber
mud
ezde
Cas
tro
etal
.(1
999a
)M
auer
5·5
7·1
129·
1B
erm
udez
deC
astr
o(1
986)
Fon
tana
Ran
ucci
o6·
5ca
.7·
5ca
.9·
5ca
.19
·0S
egre
&A
scen
zi(1
984)
Ata
puer
ca-S
H5
5·5
0·08
56·
50·
4311
8·2
Ber
mud
ezde
Cas
tro
(198
6,19
88,
1993
)B
orde
rC
ave
5—
6·6
Per
sona
lda
taN
ubia
,‘‘
Mes
olit
hic’
’,m
ales
85·
800·
3511
6·43
0·39
110·
9C
alca
gno
(198
9)N
ubia
,‘‘
Mes
olit
hic’
’,fe
mal
es12
5·48
0·34
146·
310·
3911
5·1
Cal
cagn
o(1
989)
San
,m
ales
65·
40·
47—
5·3
0·27
98·1
van
Ree
nen
(198
2)S
an,
fem
ales
55·
40·
84—
5·6
0·61
103·
7va
nR
eene
n(1
982)
San
—5
——
5·2
—6·
812
104·
0D
renn
an(1
929)
S.
A.
blac
ks,
mal
es18
15·
330·
42—
5·83
0·39
109·
4Ja
cobs
on(1
982)
S.
A.
blac
ks,
fem
ales
615·
30·
35—
5·71
0·42
107·
7Ja
cobs
on(1
982)
S.
A.
blac
ks,
mal
es56
5·43
0·45
—6·
20·
3211
4·2
Kie
ser
etal
.(1
987)
S.
A.
blac
ks,
fem
ales
495·
480·
56—
5·96
0·49
108·
8K
iese
ret
al.
(198
7)S
.A
.bl
acks
—5·
9—
—6
—7·
812
·410
1·7
Sha
w(1
931)

377J
Tab
le2
Mes
iod
ista
l(co
rrec
ted
)an
db
ucc
olin
gual
dia
met
ers
(in
mm
)an
dcr
own
shap
ein
dex
(CS
I)of
HD
P1-
4an
dof
late
ralm
and
ibu
lar
inci
sors
inse
lect
edre
cen
t,ar
chae
olog
ical
and
foss
ilh
um
ans
from
Afr
ica,
Eu
rop
ean
dA
sia
nM
DS
.D.
nB
LS
.D.
Cro
wn
heig
htR
oot
leng
thC
SI
Ref
eren
ces
HD
P1-
47·
77·
210
·313
·5in
c.93
·5P
erso
nal
data
OH
77·
77·
410
·196
·1T
obia
s(1
991)
OH
167·
87·
511
·796
·2T
obia
s(1
991)
SK
X23
546·
9—
Per
sona
lda
taS
KX
2120
47·
1—
Per
sona
lda
taK
NM
-WT
1500
07·
58·
311
·120
·511
0·7
Bro
wn
&W
alke
r(1
993)
KN
M-E
R82
07·
36·
910
·894
·5W
ood
(199
1)K
NM
-ER
992
7·2
797
·2W
ood
(199
1)K
NM
-ER
1812
7·1
6·6
93·0
Woo
d(1
991)
Dm
anis
i6·
46·
398
·4G
abun
ia&
Vek
ua(1
995)
Zho
ukou
dian
76·
80·
4—
6·9
0·3
101·
5B
erm
udez
deC
astr
oet
al.
(199
9a)
Ter
nifin
e3
6·3
7·3
115·
9B
erm
udez
deC
astr
oet
al.
(199
9a)
Rab
at7·
07·
510
7·1
Ber
mud
ezde
Cas
tro
(198
6)A
tapu
erca
-TD
AT
D6-
27·
07·
817
·811
1·4
Ber
mud
ezde
Cas
tro
etal
.(1
999a
)A
tapu
erca
-TD
AT
D6-
487·
67·
710
1·3
Ber
mud
ezde
Cas
tro
etal
.(1
999a
)M
auer
6·0
7·8
130·
0B
erm
udez
deC
astr
o(1
986)
Ata
puer
ca-S
H14
6·6
0·3
—7·
30·
411
0·6
Ber
mud
ezde
Cas
tro
etal
.(1
999a
)Je
bel
Irho
ud3
—7·
2D
ay(1
986)
Nub
ia,
‘‘M
esol
ithi
c’’,
mal
es14
6·31
0·53
156·
860·
2610
8·7
Cal
cagn
o(1
989)
Nub
ia,
‘‘M
esol
ithi
c’’,
fem
ales
155·
950·
4416
6·62
0·51
111·
3C
alca
gno
(198
9)S
an,
mal
es8
5·9
0·5
—5·
60·
3294
·9va
nR
eene
n(1
982)
San
,fe
mal
es7
5·7
0·25
—5·
70·
5410
0·0
van
Ree
nen
(198
2)S
an—
5·6
——
5·6
—7·
113
·410
0·0
Dre
nnan
(192
9)S
.A
.bl
acks
,m
ales
223
6·06
0·47
—6·
270·
3710
3·5
Jaco
bson
(198
2)S
.A
.bl
acks
,fe
mal
es83
5·99
0·42
—6·
120·
410
2·2
Jaco
bson
(198
2)S
.A
.bl
acks
,m
ales
526·
430·
51—
6·56
0·46
102·
0K
iese
ret
al.
(198
7)S
.A
.bl
acks
,fe
mal
es48
6·11
0·55
—6·
450·
2810
5·6
Kie
ser
etal
.(1
987)
S.
A.
blac
ks—
6—
—6
—8·
413
·210
0·0
Sha
w(1
931)

378 . . ET AL.
sample of H. ergaster specimens (excludingKNM-WT 15000). The BL dimensions aresimilar to other African Middle Pleistocenehominids (Tighenif 3 and Rabat) andwithin the range of the Homo heidelbergensissample from Atapuerca-SH. Buccolingualdimensions are smaller than H. habilis andspecimens from Atapuerca-TD6.
The large size of HDP1-3 and HDP1-4relative to the other samples is also evidentin bivariate plots of crown measurements[Figure 6(a) and (b)]. Both HDP1-3 andHDP1-4 fall outside the distribution of H.erectus and H. heidelbergensis. Their positionsin the plots compare most closely tospecimens of H. habilis and Atapuerca-TDATD6-48 (in the case of HDP1-4).
When the overall shape of the incisors (asexpressed by the CSI) is considered,HDP1-3 shows a marked degree of BLexpansion, whereas this doesn’t seem to bethe case for HDP1-4 (Tables 1 and 2).Bermudez de Castro et al. (1999a) describeBL expansion of the mandibular incisors asone of the evolutionary trends of hominidevolution from the Lower Pleistocenethrough the Middle Pleistocene. This trendis exemplified by the Atapuerca TD6hominids and later in time, by the Mauerspecimen and the Atapuerca SH hominids.They also hypothesized that the AfricanMiddle Pleistocene hominids may haveincreased the BL dimensions of the man-dibular incisors. The evidence from HDP1provides contrasting results in this respect.In terms of absolute size, both HDP1-3 andHDP1-4 display large BL size. In terms ofrelative BL expansion, the value of the CSIfor HDP1-3 seems to follow a trend alsoevident in other African Middle Pleistocenehominids (Tighenif and Rabat). However,despite its large absolute size, HDP1-4seems to retain a primitive condition incontrast to other African Middle Pleistocenespecimens.
The results of the metrical analysis ofHDP1-3 and HDP1-4, as well as the avail-
ability of a more expanded data set than thatavailable to Berger & Parkington (1995),prompted us to re-examine the crowndimensions of the two molars, HDP1-1 andHDP1-2. Bivariate plots of the dentaldimensions of HDP1-1 and HDP1-2 aredisplayed in Figure 7(a) and (b). Specimensand samples analysed are listed in thefootnotes.
A different pattern appears to emerge inthis analysis. In the case of HDP1-1 (LM2),the dimensions fall well inside the distribu-tion of both H. erectus and H. heidelbergensis,and are smaller than specimens of H. habilisand H. ergaster. In the case of HDP1-2(RM3), the dimensions are also smaller thanspecimens of H. habilis and H. ergaster andfall just outside the distribution of H. erectusand H. heidelbergensis. Both HDP1-1 andHDP1-2 fall towards the lower end of thedistribution for African Middle Pleistocenespecimens. Overall, both molars appear tobe relatively small when compared with theother fossil samples examined in this study.Size reduction of the posterior teeth seemsto be a common trend in Lower and MiddlePleistocene hominids (Bermudez de Castro& Nicolas, 1995; Bermudez de Castro et al.,1999a), and the molars from HDP1 appearto fit this trend.
The results of the metrical analyses of theHDP1 incisors and molars suggest that weare looking at an individual that displays apattern of dental metrical features (largeanterior teeth and small molars) that is com-parable with that seen in other African andEuropean Middle Pleistocene hominids,e.g. Atapuerca (Bermudez de Castro, 1993)and Fontana Ranuccio (Ascenzi & Segre,1996).
A taxonomic attribution based on a fewdental remains is more problematic,especially when dealing with specimensfrom the African Middle Pleistocene, thatare presently subject to debate (seeMcBrearty & Brooks, 2000 for a review).Churchill et al. (2000) suggested that

379J
5.58.0
8.5
MD
(b)
BL
5.5
8.0
7.5
7.0
6.5
6.0
6.0 6.5 7.0 7.5
H. heidelbergensis
SA BlacksH. erectus
San
5.07.5
7.5
MD
(a)B
L
5.0
7.0
6.5
5.5
5.5 6.0 6.5 7.0
H. heidelbergensis
SA BlacksH. erectus
San
6.0
HDP1–3NubiaAFMPDmanisiH.ergasterH.habilis
HDP1–4Nubia
AFMPDmanisiH.ergasterH.habilis
ATD6
Figure 6. Bivariate plots comparing the dental dimensions of (a) HDP1-3 and (b) HDP1-4 to those ofselected archaeological and fossil humans from Africa, Europe and Asia. Specimens included andreferences are listed in Tables 1 and 2. H. habilis includes OH 7, OH 16. H. ergaster includes KNM-WT15,000, KNM-ER 820, KNM-ER 992, KNM-ER 1812. H. erectus includes the Khoukoudian sample andSangiran 11. H. heidelbergensis includes the Atapuerca sample and the Mauer mandible. The AfricanMiddle Pleistocene sample (AFMP) includes Tighenif and Rabat. ATD6 indicates Atapuerca TD6hominids. Fontana Ranuccio has not been included because measurements given are approximate.
the tibia recovered from HDP1 might beattributed to H. heidelbergenesis in view of itsmorphological similarities to the Boxgrove
tibia. This view lends support to the idea byRightmire (1998) of a geographically broaddistribution of this Middle Pleistocene

380 . . ET AL.
9.514.0
17.0
MD
(b)
BL
7.5
H. heidelbergensis
H. erectus
13.513.012.512.011.511.010.510.09.59.08.58.0
16.5
16.0
15.5
15.0
14.5
14.0
13.5
13.0
12.5
12.0
11.5
11.0
10.5
10.0
11.014.0
16.5
MD
(a)
BL
9.5
H. heidelbergensis
H. erectus
13.513.012.512.011.511.010.510.0
16.0
15.5
15.0
14.5
14.0
13.5
13.0
12.5
12.0
11.5
HDP1–2SA BlacksFlorisbadAFMPearly HomoH.ergaster
HDP1–2SA Blacks
D2282AFMPearly HomoH.ergaster
ATD6–12
Figure 7. Bivariate plots comparing the dental dimensions of (a) HDP1-1 and (b) HDP1-2 to those ofselected recent and fossil humans from Africa, Europe and Asia. Specimens included: South AfricanBlacks (Jacobson, 1982). Atapuerca TD6 (Bermudez de Castro et al., 1999a,b). Dmanisi D2282(Gabunia et al., 2000). African Middle Pleistocene (AFMP): Kabwe (Day, 1986): Rabat (Bermudez deCastro, 1986); Thomas III (Bermudez de Castro, 1986); Sale’ (Day, 1986); Lainyamok (Shipman et al.,1983); Dagadle’ (de Bonis et al., 1984); Laetoli H 18 (Day et al., 1980). Florisbad, personal data. H.heidelbergensis: Atapuerca SH (Bermudez de Castro, 1986, 1988, 1993); Petralona (Bermudez de Castro,1986); Arago XXI (Bermudez de Castro, 1986). H. erectus: Zhoukoudian (Wood, 1991); Sangiran(Wood, 1991). H. ergaster: (KNM ER 807, ER 1808, ER 3733) (Wood, 1991); KNM WT 15,000 (Brown& Walker, 1993). Early Homo: H. habilis (OH 13, OH 15, OH 16, OH 24, OH 39, ER 1813) (Tobias,1991; Wood & Richmond, 2000); South African early Homo (SE 1508, SK 27, SK 847) personal data.

381J
species. Other authors, however, contendthat the species H. heidelbergensis is part ofan exclusively European lineage leading toHomo neanderthalensis, whereas anotherlineage with an African distribution rootedin the species H. antecessor would havegiven rise to H. sapiens (Bermudez deCastro et al., 1997, 1999a). In this respect,it is interesting to note that the dentalremains from HDP1 seem to display someof the traits described by Bermudez deCastro et al. (1999a) as characteristic of agroup of hominids which include, amongothers, the ATD6 human remains. Thesetraits include size reduction of the thirdmolars and increase of the BL dimensionsof the mandibular incisors. However, theavailable evidence is still insufficient topoint to closer affinities of the HDP1sample with either the African or EuropeanMiddle Pleistocene hominids.
Conclusion
The human remains from HDP1 (twomaxillary molars, two mandibular incisorsand a tibia) probably date from the lateMiddle Pleistocene. The analysis presentedhere suggests that the two incisors and thetwo molars belong to a single individual.The incisors display early signs of a wearpattern often seen in people pursuing ahunter-gatherer lifestyle. These incisors arealso larger than a sample of modern andarchaeological African dental remains. Theyare comparable in size with dentition fromAfrican, European and Asian early Middleand late Middle Pleistocene humans. Themetrical evidence from these incisors inconjunction with a re-examination ofmetrical data from the molars, suggest thatthe Hoedjiespunt 1 hominid possesseddental metrical features (large anterior teethand small molars) comparable with otherAfrican and European Middle Pleistocenehominids.
Acknowledgements
We thank the National Monuments Council(now the South African Heritage ResourcesAgency) for issuing the necessary permits forexcavation; Dr S. Woodborne of the CSIRfor running IRSL and TL analysis of sedi-ments from Hoedjiespunt; Dr I. McMillanof De Beers Marine for the examination ofthe foraminiferal assemblage; Professor J. C.Sealy, Dr J. A. Lee-Thorp and Miss K. L.Van Niekerk of the Department ofArchaeology at the University of Cape Townfor their useful comments on drafts ofthe manuscript; Mr C. Poggenpoel of theDepartment of Archaeology at theUniversity of Cape Town for help withthe excavations; and Mr P. Faugust of theDepartment of Anatomical Sciences at theUniversity of the Witwatersrand for photo-graphic assistance. Comments from ananonymous referee vastly improved themanuscript. The research was funded bygrants from the Foundation for ResearchDevelopment (now the National ResearchFoundation) (grant number 2034546),the Palaeo-Anthropology Scientific Trust,the National Geographic Society and thePalaeo-anthropology Unit for Researchand Exploration of the University of theWitwatersrand.
References
Anderson, D. L., Thompson, G. W. & Popovich, F.(1976). Age of attainment of mineralization stagesof the permanent dentition. J. Forensic Sci. 21,191–200.
Ascenzi, A. & Segre, A. G. (1996). Artefacts andhuman teeth at the Fontana Ranuccio MiddlePleistocene site (Central Italy). Anthropologie (Brno)34, 39–46.
Baba, H., Aziz, F., Narasaki, S., Sudijono, Kaifu, Y.,Suprijo, A., Hyodo, M., Susanto, E. E. & Jacob, T.(2000). A new hominid incisor from Sangiran,Central Java. J. hum. Evol. 38, 855–862.
Berger, L. R. & Parkington, J. E. (1995). Brief com-munication: A new Pleistocene hominid-bearinglocality at Hoedjiespunt, South Africa. Am. J. phys.Anthrop. 98, 601–609.

382 . . ET AL.
Bermudez de Castro, J. M. (1986). Dental remainsfrom Atapuerca (Spain). I. Metrics. J. hum. Evol. 15,265–287.
Bermudez de Castro, J. M. (1988). Dental remainsfrom Atapuerca/Ibeas (Spain) II. Morphology.J. hum. Evol. 17, 279–304.
Bermudez de Castro, J. M. (1993). The Atapuercadental remains. New evidence (1987–1991 excava-tions) and interpretations. J. hum. Evol. 24, 339–371.
Bermudez de Castro, J. M. & Nicolas, M. E. (1995).Posterior dental size reduction in hominids: theAtapuerca evidence. Am. J. phys. Anthrop. 96,335–356.
Bermudez de Castro, J. M. & Rosas, A. (2001). Patternof dental development in Hominid XVIII from theMiddle Pleistocene Atapuerca-Sima de los Huesossite (Spain). Am. J. phys. Anthrop. 114, 325–330.
Bermudez de Castro, J. M., Arsuaga, J. L., Carbonell,E., Rosas, A., Martinez, I. & Mosquera, M. (1997).A Hominid from the Lower Pleistocene Atapuerca,Spain: possible ancestor to Neandertals and modernhumans. Science 276, 1392–1395.
Bermudez de Castro, J. M., Rosas, A. & Nicolas, M. E.(1999a). Dental remains from Atapuerca-TD6(Gran Dolina site, Burgos, Spain). J. hum. Evol. 37,523–566.
Bermudez de Castro, J. M., Rosas, A., Carbonell, E.,Nicolas, M. E., Rodriguez, J. & Arsuaga, J. L.(1999b). A modern human pattern of dental devel-opment in Lower Pleistocene hominids fromAtapuerca-TD6 (Spain). Proc. Natn. Acad. Sci. USA96, 4210–4213.
Brown, B. & Walker, A. (1993). The dentition. In (A.Walker & R. E. Leakey, Eds) The NariokotomeHome erectus skeleton, pp. 161–192. Berlin: SpringerVerlag.
Calcagno, J. M. (1989). Mechanisms of human dentalreduction. A case study from Post-PleistoceneNubia. University of Kansas Publications inAnthropology 18. Lawrence: University of Kansas.
Churchill, S. E., Berger, L. R. & Parkington, J. E.(2000). A Middle Pleistocene human tibia fromHoedjiespunt, Western Cape, South Africa. S. Afr. J.Sci. 96, 367–368.
Day, M. H. (1986). Guide to Fossil Man. 4th edn.London: Cassell.
Day, M. H., Leakey, M. D. & Magori, C. (1980). Anew hominid fossil skull from the Ngaloba Beds,Laetoli, northern Tanzania. Nature 284, 55–56.
Dean, M. C., Beynon, A. D., Reid, D. J. & Whittaker,D. K. (1993). A longitudinal study of tooth growth ina single individual based on long- and short-periodincremental markings in dentine and enamel. Int. J.Osteoarch. 3, 249–264.
de Bonis, L., Geraads, D., Guerin, G., Haga, A.,Jaeger, J. J. & Sen, S. (1984). Decouverte d’unHominide fossile dans le Pleistocene de laRepublique de Djibouti. C. r. Acad. Sci. Paris 299,1097–1100.
Drennan, M. R. (1929). The dentition of a Bushmantribe. Ann. S. Afr. Mus. 24, 61–87.
Gabunia, L. & Vekua, A. (1995). A Plio-Pleistocenehominid from Dmanisi, East Georgia, Caucasus.Nature 373, 509–512.
Gabunia, L., Vekua, A., Lordkipanidze, D., SwisherIII, C. C., Ferring, R., Justus, A., Nioradze,M., Tvalchrelidze, M., Anton, S. C., Bosinski, G.,Joris, O., de Lumley, M. A., Majsuradze, G. &Mouskhelishvili, A. (2000). Earliest Pleistocenehominid cranial remains from Dmanisi, Republicof Georgia; taxonomy, geological setting, and age.Science 288, 1019–1025.
Garn, S. M., Lewis, A. B. & Bonne, B. (1962). Thirdmolar formation and its development course. AngleOrthodontist 32, 270–279.
Grine, F. E. & Klein, R. G. (1993). Late Pleistocenehuman remains from the Sea Harvest site, SaldanhaBay, South Africa. S. Afr. J. Sci. 89, 145–152.
Grun, R., Brink, J. S., Spooner, N. A., Taylor, L.,Stringer, C. B., Franciscus, R. G. & Murray, A. S.(1996). Direct dating of Florisbad hominid. Nature382, 500–501.
Hinton, J. (1981). Form and patterning of anteriortooth wear among aboriginal human groups. Am. J.phys. Anthrop. 54, 557–564.
Jacobson, A. (1982). The Dentition of the South AfricanNegro. Anniston: Higginbotham.
Kieser, J. A., Groeneveld, H. T. & Cameron, N.(1987). Evidence for a secular trend in the Negrodentition. Ann. Hum. Biol. 14, 517–532.
Klein, R. G. (1983). Palaeoenvironmental implicationsof Quaternary large mammals in the Fynbos Biome.South African National Science Progress Report 75,116–138.
McBrearty, S. & Brooks, A. S. (2000). The revolutionthat wasn’t: a new interpretation of the origin ofmodern human behavior. J. hum. Evol. 39, 453–563.
Moorrees, C. F. A., Fanning, E. A. & Hunt, E. E.(1963). Age variation of formation stages for tenpermanent teeth. J. Dent. Res. 42, 1490–1502.
Rightmire, G. P. (1998). Human evolution in theMiddle Pleistocene: the role of Homo heidelbergensis.Evol. Anthrop. 6, 218–227.
Segre, A. G. & Ascenzi, A. (1984). Fontana Ranuccio:Italy’s earliest Middle Pleistocene hominid site. Curr.Anthrop. 25, 230–233.
Shaw, J. C. M. (1931). The Teeth, the Bony Palate andthe Mandible in Bantu races of South Africa. London:John Bull, Sons & Danielsson.
Shipman, P., Potts, R. & Pickford, M. (1983).Lainyamok, a new middle Pleistocene hominid site.Nature 306, 365–368.
Smith, B. H. (1984). Patterns of molar wear in hunter-gatherers and agriculturalists. Am. J. phys. Anthrop.63, 39–56.
Smith, B. H. (1991). Standards of human tooth forma-tion and dental age assessment. In (M. Kelley &C. S. Larsen, Eds) Advances in Dental Anthropology,pp. 143–168. New York: Wiley-Liss.
Stynder, D. D. (1997). The use of faunal evidence toreconstruct site history at Hoedjiespunt 1 (HDP1),Western Cape. M.A. Thesis. Department ofArchaeology, University of Cape Town.

383J
Tobias, P. V. (1991). Olduvai Gorge, Volume IV.The Skulls, Endocasts and Teeth of Homo habilis.Cambridge: Cambridge University Press.
Tompkins, R. L. (1996a). Human population varia-bility in relative dental development. Am. J. phys.Anthrop. 99, 79–102.
Tompkins, R. L. (1996b). Relative dental developmentof Upper Pleistocene hominids compared to humanpopulation variation. Am. J. phys. Anthrop. 99,103–118.
van Reenen, J. F. (1982). The effects of attritionon tooth dimensions of San (Bushmen). In (B.
Kurten, Ed.) Teeth: Form, Function and Evolution,pp. 182–203. New York: Columbia University Press.
Wood, B. A. (1991). Koobi Fora Research Project IV:Hominid Cranial Remains from Koobi Fora. Oxford:Clarendon Press.
Wood, B. A. & Richmond, B. G. (2000). Humanevolution: taxonomy and paleobiology. J. Anat. 196,19–60.
Woodborne, S. (2000). Luminescence dating of theMiddle Stone Age in South Africa. Report to theNational Science Foundation.





























