Embed Size (px)
Citation preview

Molecular and Cellular Endocrinology 383 (2014) 137–146
Contents lists available at ScienceDirect
Molecular and Cellular Endocrinology
journal homepage: www.elsevier .com/locate /mce
Human white adipocytes express the cold receptor TRPM8 whichactivation induces UCP1 expression, mitochondrial activationand heat production
0303-7207/$ - see front matter � 2013 Elsevier Ireland Ltd. All rights reserved.http://dx.doi.org/10.1016/j.mce.2013.12.005
⇑ Corresponding author. Address: Department of Medicine – DIMED, School ofMedicine, University of Padua, Via Giustiniani 2, 35128 Padova, Italy. Tel.: +39 0498218747; fax: +39 049 8213332.
E-mail address: [email protected] (M. Rossato).
Marco Rossato a,⇑, Marnie Granzotto a, Veronica Macchi b, Andrea Porzionato b, Lucia Petrelli b,Alessandra Calcagno a, Juri Vencato c, Diego De Stefani d, Valentina Silvestrin a, Rosario Rizzuto d,Franco Bassetto e, Raffaele De Caro b, Roberto Vettor a
a Department of Medicine – DIMED, Clinica Medica 3, School of Medicine, University of Padova, Padova, Italyb Department of Molecular Medicine, Section of Anatomy, School of Medicine, University of Padova, Padova, Italyc Department of Animal Medicine, Production and Health, University of Padova, Padova, Italyd Department of Biomedical Sciences, School of Medicine, University of Padova, Padova, Italye Department of Neurosciences, Plastic Surgery Clinic, University of Padova, Padova, Italy
a r t i c l e i n f o
Article history:Received 20 June 2013Received in revised form 10 November 2013Accepted 9 December 2013Available online 14 December 2013
Keywords:Human adipocyteTRPM8UCP1MitochondriaMenthol
a b s t r a c t
Mammals possess two types of adipose tissue, white (WAT) and brown (BAT). The uncoupling protein 1(UCP1) is a hallmark of BAT, being the pivotal player for cold-induced thermogenesis. WAT can acquireBAT characteristics with up-regulation of UCP1 after cold exposure or adrenergic stimulation.
In the present study we demonstrated that human white adipocytes express the cold-sensing receptorTRPM8 which activation by menthol and icilin induced a rise in [Ca2+]i and UCP1 expression, increasedmitochondrial membrane potential, glucose uptake and heat production. The induction of ‘‘brown-like’’phenotype in human white adipocytes after TRPM8 activation was supported by ultrastructural morpho-logical changes of mitochondrial morphology and of their intracellular localization, with no modificationsof the genes regulating mitochondrial biogenesis.
In conclusion human white adipocytes express the cold receptor TRPM8 which activation induces their‘‘browning’’ supporting a possible role of this receptor in the control of adipose tissue metabolism andbody energy balance.
� 2013 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
It is well known that mammals possess two distinct adipose tis-sues: white (WAT) and brown (BAT) adipose tissue with the latterexpressing UCP1, a unique protein localized in the inner membraneof mitochondria where it allows protons to leak back into the mito-chondrial matrix from the inner-membrane space resulting in en-ergy dissipation as heat without ATP production (Frontini andCinti, 2010).
Many recent studies have demonstrated that WAT and BATshould not be considered separated organs but intimately relatedtissues within the so called ‘‘adipose organ’’ (Cinti, 2005; Frontiniand Cinti, 2010) that can undergo reciprocal transformation (Bar-batelli et al., 2010; Cinti, 2009). Since it has been shown that
WAT over-expressing the uncoupling protein UCP1 in mouse pro-tects from obesity (Kopecky et al., 1995) and that BAT downregu-lation results in obesity (Bachman et al., 2002; Lowell et al.,1993), the possibility to transdifferentiate WAT in BAT could offera novel strategy to treat obesity and its consequences. The recentevidence of functionally active BAT in adult humans further high-lights the role of BAT as a therapeutical target for studies evaluat-ing the pathophysiology of obesity and its therapy (Cypess et al.,2009; Lim et al., 2012; van Marken Lichtenbelt et al., 2009; Virta-nen et al., 2009).
BAT can be easily and rapidly recruited and activated by adren-ergic stimulation as well as by cold exposure that is ultimately anadrenergic mediated phenomenon (Cinti, 2009; Collins et al., 2004;Lim et al., 2012; Seale et al., 2011). The origin of newly recruitedbrown adipocytes is still debated since they could derive eitherfrom myf-5 positive mesenchymal stem cells giving origin also tomuscle cells (Himms-Hagen et al., 2000) or from white adipocytestransdifferentiation (Barbatelli et al., 2010). Recently a third dis-tinct population of fat cells originating from non myf-5 precursors
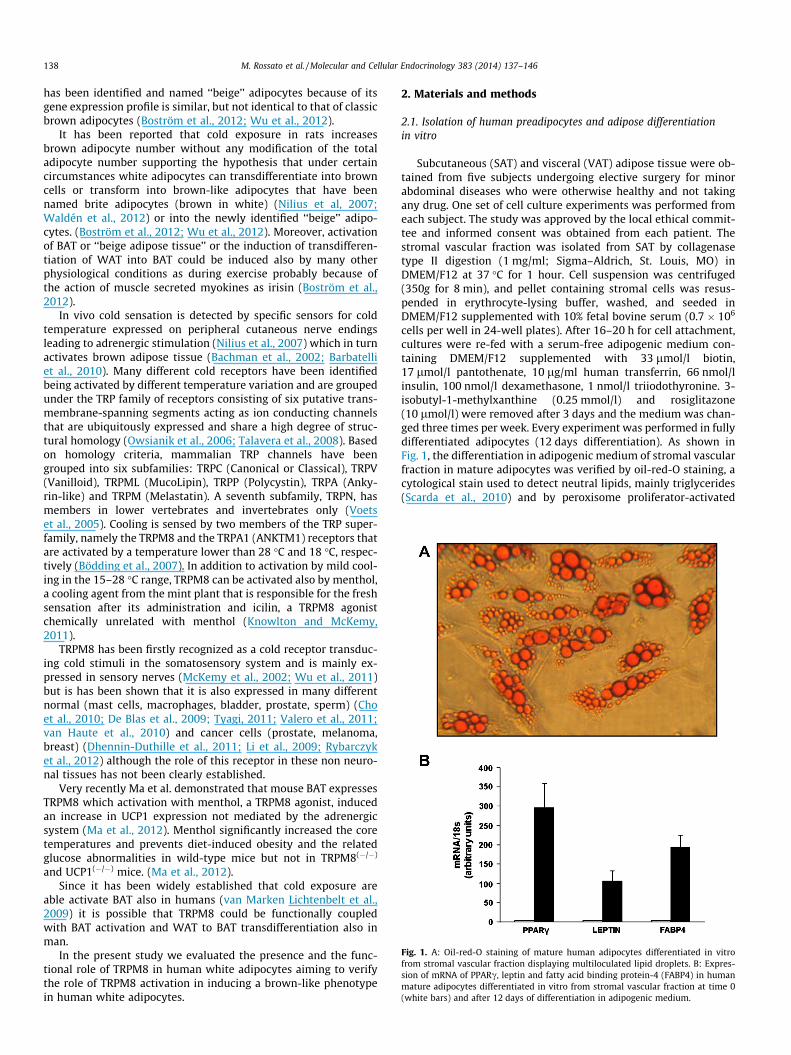
Fig. 1. A: Oil-red-O staining of mature human adipocytes differentiated in vitrofrom stromal vascular fraction displaying multiloculated lipid droplets. B: Expres-sion of mRNA of PPARc, leptin and fatty acid binding protein-4 (FABP4) in humanmature adipocytes differentiated in vitro from stromal vascular fraction at time 0(white bars) and after 12 days of differentiation in adipogenic medium.
138 M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146
has been identified and named ‘‘beige’’ adipocytes because of itsgene expression profile is similar, but not identical to that of classicbrown adipocytes (Boström et al., 2012; Wu et al., 2012).
It has been reported that cold exposure in rats increasesbrown adipocyte number without any modification of the totaladipocyte number supporting the hypothesis that under certaincircumstances white adipocytes can transdifferentiate into browncells or transform into brown-like adipocytes that have beennamed brite adipocytes (brown in white) (Nilius et al, 2007;Waldén et al., 2012) or into the newly identified ‘‘beige’’ adipo-cytes. (Boström et al., 2012; Wu et al., 2012). Moreover, activationof BAT or ‘‘beige adipose tissue’’ or the induction of transdifferen-tiation of WAT into BAT could be induced also by many otherphysiological conditions as during exercise probably because ofthe action of muscle secreted myokines as irisin (Boström et al.,2012).
In vivo cold sensation is detected by specific sensors for coldtemperature expressed on peripheral cutaneous nerve endingsleading to adrenergic stimulation (Nilius et al., 2007) which in turnactivates brown adipose tissue (Bachman et al., 2002; Barbatelliet al., 2010). Many different cold receptors have been identifiedbeing activated by different temperature variation and are groupedunder the TRP family of receptors consisting of six putative trans-membrane-spanning segments acting as ion conducting channelsthat are ubiquitously expressed and share a high degree of struc-tural homology (Owsianik et al., 2006; Talavera et al., 2008). Basedon homology criteria, mammalian TRP channels have beengrouped into six subfamilies: TRPC (Canonical or Classical), TRPV(Vanilloid), TRPML (MucoLipin), TRPP (Polycystin), TRPA (Anky-rin-like) and TRPM (Melastatin). A seventh subfamily, TRPN, hasmembers in lower vertebrates and invertebrates only (Voetset al., 2005). Cooling is sensed by two members of the TRP super-family, namely the TRPM8 and the TRPA1 (ANKTM1) receptors thatare activated by a temperature lower than 28 �C and 18 �C, respec-tively (Bödding et al., 2007). In addition to activation by mild cool-ing in the 15–28 �C range, TRPM8 can be activated also by menthol,a cooling agent from the mint plant that is responsible for the freshsensation after its administration and icilin, a TRPM8 agonistchemically unrelated with menthol (Knowlton and McKemy,2011).
TRPM8 has been firstly recognized as a cold receptor transduc-ing cold stimuli in the somatosensory system and is mainly ex-pressed in sensory nerves (McKemy et al., 2002; Wu et al., 2011)but is has been shown that it is also expressed in many differentnormal (mast cells, macrophages, bladder, prostate, sperm) (Choet al., 2010; De Blas et al., 2009; Tyagi, 2011; Valero et al., 2011;van Haute et al., 2010) and cancer cells (prostate, melanoma,breast) (Dhennin-Duthille et al., 2011; Li et al., 2009; Rybarczyket al., 2012) although the role of this receptor in these non neuro-nal tissues has not been clearly established.
Very recently Ma et al. demonstrated that mouse BAT expressesTRPM8 which activation with menthol, a TRPM8 agonist, inducedan increase in UCP1 expression not mediated by the adrenergicsystem (Ma et al., 2012). Menthol significantly increased the coretemperatures and prevents diet-induced obesity and the relatedglucose abnormalities in wild-type mice but not in TRPM8(�/�)
and UCP1(�/�) mice. (Ma et al., 2012).Since it has been widely established that cold exposure are
able activate BAT also in humans (van Marken Lichtenbelt et al.,2009) it is possible that TRPM8 could be functionally coupledwith BAT activation and WAT to BAT transdifferentiation also inman.
In the present study we evaluated the presence and the func-tional role of TRPM8 in human white adipocytes aiming to verifythe role of TRPM8 activation in inducing a brown-like phenotypein human white adipocytes.
2. Materials and methods
2.1. Isolation of human preadipocytes and adipose differentiationin vitro
Subcutaneous (SAT) and visceral (VAT) adipose tissue were ob-tained from five subjects undergoing elective surgery for minorabdominal diseases who were otherwise healthy and not takingany drug. One set of cell culture experiments was performed fromeach subject. The study was approved by the local ethical commit-tee and informed consent was obtained from each patient. Thestromal vascular fraction was isolated from SAT by collagenasetype II digestion (1 mg/ml; Sigma–Aldrich, St. Louis, MO) inDMEM/F12 at 37 �C for 1 hour. Cell suspension was centrifuged(350g for 8 min), and pellet containing stromal cells was resus-pended in erythrocyte-lysing buffer, washed, and seeded inDMEM/F12 supplemented with 10% fetal bovine serum (0.7 � 106
cells per well in 24-well plates). After 16–20 h for cell attachment,cultures were re-fed with a serum-free adipogenic medium con-taining DMEM/F12 supplemented with 33 lmol/l biotin,17 lmol/l pantothenate, 10 lg/ml human transferrin, 66 nmol/linsulin, 100 nmol/l dexamethasone, 1 nmol/l triiodothyronine. 3-isobutyl-1-methylxanthine (0.25 mmol/l) and rosiglitazone(10 lmol/l) were removed after 3 days and the medium was chan-ged three times per week. Every experiment was performed in fullydifferentiated adipocytes (12 days differentiation). As shown inFig. 1, the differentiation in adipogenic medium of stromal vascularfraction in mature adipocytes was verified by oil-red-O staining, acytological stain used to detect neutral lipids, mainly triglycerides(Scarda et al., 2010) and by peroxisome proliferator-activated

M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146 139
receptor c (PPARc), leptin, and fatty acid binding protein-4 (FABP4)genes that are traditionally expressed in mature adipocytes (Scardaet al., 2010). Adipocytes were stimulated overnight with theTRPM8 agonists menthol (1, 10, 100 lM) and in some experimentswith icilin (1 lM) dissolved in ethanol. Since the highest effectswith menthol were obtained at 100 lM, this was the preferredconcentration utilized for the majority of the experiments. Controlexperiments were performed adding vehicle only. When evaluat-ing the effects of ‘‘cold’’, differentiated adipocytes were incubatedfor 24 h at 26 �C, a temperature that is known to activate TRPM8receptor (Knowlton and McKemy, 2011). Under these experimentalconditions adipocytes viability was not significantly affected asdetermined by trypan blue staining, a well known technique usedto selectively stain dead cells that are easily identifiable as blue un-der light microscope (not shown).
2.2. Glucose uptake in primary human adipocytes
Fully differentiated adipocytes were incubated in the absence orpresence of insulin (2 lM), menthol (1, 10, 100 lM), icilin (1 lM)alone or in combination overnight. When used in combination,insulin was added 30 min before menthol or icilin and thenwashed out. The assay was initiated by the addition of 300 ll ofa solution of 50 lM D-glucose and 1.5 lCi/ml 2-deoxy-D-[1-3H]-glucose (Amersham-Biosciences, Piscataway, NJ, USA) (35). After15 min at 37 �C, the assay was terminated by two rapid washesin ice-cold PBS. Cells were solubilized in 500 ll NaOH 0.5 M andthe radioactivity was measured using a ß-counter (Wallace-Per-kin-Elmer, MA, USA). The glucose uptake was expressed in countsper minute (CPM). Fifty microliters of each sample were used fortotal protein quantification with Coomassie protein assay kit(Pierce, Rockford, IL). Glucose uptake was normalized to total pro-tein content and expressed as percentage of mean basal value.
2.3. RNA isolation, qualitative and quantitative RT-PCR
Total RNA was extracted from adipocytes at 12th day of in vitrodifferentiation using QIAMP mini kit (QIAGEN) following the man-ufacturer’s protocol. First-strand cDNAs were synthesized fromequal amounts of total RNA using random primers and M-MLV re-verse transcriptase (Promega). For qualitative PCR analysis, 5 ll ofcDNA product from 6 and 12 days differentiated adipocytes wereamplified in 10 ll of HotStarTaq Master Mix Kit (QIAGEN) in thepresence of the specific primers for TRPM8 for 30 cycles at 57 �C.The reaction included 5 ll of cDNA from prostate gland as positivecontrol and no sample negative control (saline).
Quantitative real time PCR for UCP1, TRPM8, mitochondrial pro-teins and ‘‘browning’’ genes was performed using Sybr Green flu-orofore. The change in fluorescence at every cycle was monitoredand a threshold cycle above background for each reaction was cal-culated. A melt curve analysis was performed following every runto ensure a single amplified product for every reaction. All reac-tions were carried out in at least duplicated for every sample.18S rRNA was constantly expressed under all experimental condi-tions and was then used as a reference gene for normalization. Ta-ble 1 reports the list and sequence of the primers employed in PCRexperiments.
2.4. Mitochondrial DNA quantification
Total DNA was obtained from adipocytes using the QIAmp DNAMini Kit (QIAGEN) according to the manufacturer’s protocol andstored in buffer AE (QIAGEN). Mitochondrial DNA copy numberwas determined by Sybr Green chemistry from a portion of cyto-chrome B (mtDNA) gene, normalized using a nuclear encoded tem-plate for the ribosomal (18S, nDNA) gene. Each reaction was
optimized and confirmed linear over an appropriate concentrationrange using genomic DNA standard. Samples were analyzed in trip-licate, enabling calculation of the average mtDNA/nDNA ratio.
2.5. TRPM8 detection in human adipocytes by immunofluorescence
Human adipocytes were cultured and grown on glass coverslipsas described above. After differentiation, cells were fixed with ice-cold acetone for 5 min at room temperature and then washedtwice with ice cold PBS. TRPM8 was detected by incubation witha rabbit polyclonal primary antibody (diluted 1:200, Abcam, Cam-bridge, UK) for 1 hour at room temperature using standard proce-dures. After washing with PBS, binding of primary antibodies wasdetected with FITC-conjugated secondary antibodies (MolecularProbes, Invitrogen, Milan, Italy). Adipocytes were then examinedfor localization of fluorescence with a Nikon fluorescence micro-scope. All passages were at room temperature. Negative controlstudies were performed using the same procedure except thatthe primary antibody was omitted.
2.6. Measurement of [Ca2+]i in human adipocytes
Primary cultures of adipocytes were differentiated on glass cov-erslips and incubated with 2 lM fura-2/AM (Molecular Probes,Invitrogen) for 30 min at 37 �C in the dark as previously described(Pagano et al., 2007). After incubation, cells were washed twice inthe standard medium and suspended in 1 ml of a solution contain-ing (mM) 140 NaCl, 5.0 KCl, 1.0 MgCl2, 25 HEPES, and 5 glucose (pH7.4) at 37 �C, with or without the addition of 1.0 mM CaCl2 (forstandard or Ca2+-free medium, respectively). Coverslips weretransferred to the recording chamber, maintained at 37 �C, andplaced on the stage of an inverted epifluorescence microscope.Cells were stimulated with menthol (100 lM) in medium with orwithout calcium. Images of fura-2 loaded adipocytes with the exci-tation wavelength alternating between 340 and 380 nm were cap-tured with a cooled CCD camera. After subtraction of backgroundfluorescence, the ratio of fluorescence intensity at the two wave-lengths was calculated. Ratio levels in groups of 10–15 individualcells per coverslip were analyzed using MetaFluor software pack-age (Universal Imaging Corp., West Chester, PA). All graphs areaveraged responses from groups of 50–100 individual cells fromrepresentative single experiments. All experiments have been re-peated on three separate occasions, and similar results were ob-tained. In parallel experiments fura-2 loaded adipocytes wereincubated in Ca2+-free medium before menthol addition. All exper-iments have been repeated on three separate occasions.
2.7. Western blotting analysis for UCP1 in human adipocytes
Human white adipocytes were cultured and grown in 6 wellplate, homogenized in RIPA Lysis Buffer with protease inhibitors(Sigma–Aldrich, Milan, Italy), centrifuged at 800 g for 10 min at4 �C and then the middle layer was collected. Equal amounts(40 lg) of proteins were separated on 10% SDS-polyacrylamidegel, transferred to a nitrocellulose membrane and detected by pri-mary antibodies, respectively rabbit polyclonal anti-human UCP1(1:1000, Sigma–Aldrich, Milan, Italy) and mouse monoclonalanti-human b-actin (1:5000, Sigma–Aldrich, Milan, Italy). Second-ary antibodies were peroxidase-conjugated anti-rabbit IgG forUCP1 and anti-mouse IgG for b-actin. Western blot analysis wasperformed using Immobilion Western Chemiluminescent HRP Sub-strate (Millipore) and detection was made using photographicfilms. The images have been analyzed by densitometry, whichevaluates the relative amount of protein staining and quantifiesthe results in terms of optical density.
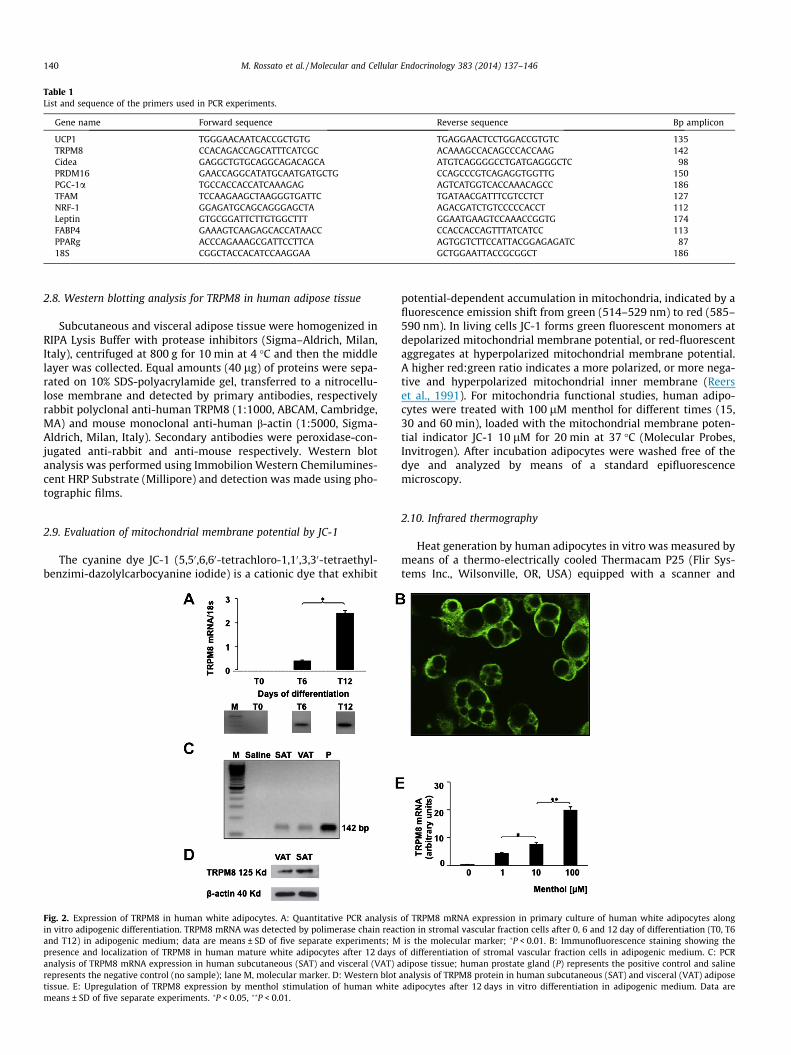
Table 1List and sequence of the primers used in PCR experiments.
Gene name Forward sequence Reverse sequence Bp amplicon
UCP1 TGGGAACAATCACCGCTGTG TGAGGAACTCCTGGACCGTGTC 135TRPM8 CCACAGACCAGCATTTCATCGC ACAAAGCCACAGCCCACCAAG 142Cidea GAGGCTGTGCAGGCAGACAGCA ATGTCAGGGGCCTGATGAGGGCTC 98PRDM16 GAACCAGGCATATGCAATGATGCTG CCAGCCCGTCAGAGGTGGTTG 150PGC-1a TGCCACCACCATCAAAGAG AGTCATGGTCACCAAACAGCC 186TFAM TCCAAGAAGCTAAGGGTGATTC TGATAACGATTTCGTCCTCT 127NRF-1 GGAGATGCAGCAGGGAGCTA AGACGATCTGTCCCCCACCT 112Leptin GTGCGGATTCTTGTGGCTTT GGAATGAAGTCCAAACCGGTG 174FABP4 GAAAGTCAAGAGCACCATAACC CCACCACCAGTTTATCATCC 113PPARg ACCCAGAAAGCGATTCCTTCA AGTGGTCTTCCATTACGGAGAGATC 8718S CGGCTACCACATCCAAGGAA GCTGGAATTACCGCGGCT 186
140 M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146
2.8. Western blotting analysis for TRPM8 in human adipose tissue
Subcutaneous and visceral adipose tissue were homogenized inRIPA Lysis Buffer with protease inhibitors (Sigma–Aldrich, Milan,Italy), centrifuged at 800 g for 10 min at 4 �C and then the middlelayer was collected. Equal amounts (40 lg) of proteins were sepa-rated on 10% SDS-polyacrylamide gel, transferred to a nitrocellu-lose membrane and detected by primary antibodies, respectivelyrabbit polyclonal anti-human TRPM8 (1:1000, ABCAM, Cambridge,MA) and mouse monoclonal anti-human b-actin (1:5000, Sigma-Aldrich, Milan, Italy). Secondary antibodies were peroxidase-con-jugated anti-rabbit and anti-mouse respectively. Western blotanalysis was performed using Immobilion Western Chemilumines-cent HRP Substrate (Millipore) and detection was made using pho-tographic films.
2.9. Evaluation of mitochondrial membrane potential by JC-1
The cyanine dye JC-1 (5,50,6,60-tetrachloro-1,10,3,30-tetraethyl-benzimi-dazolylcarbocyanine iodide) is a cationic dye that exhibit
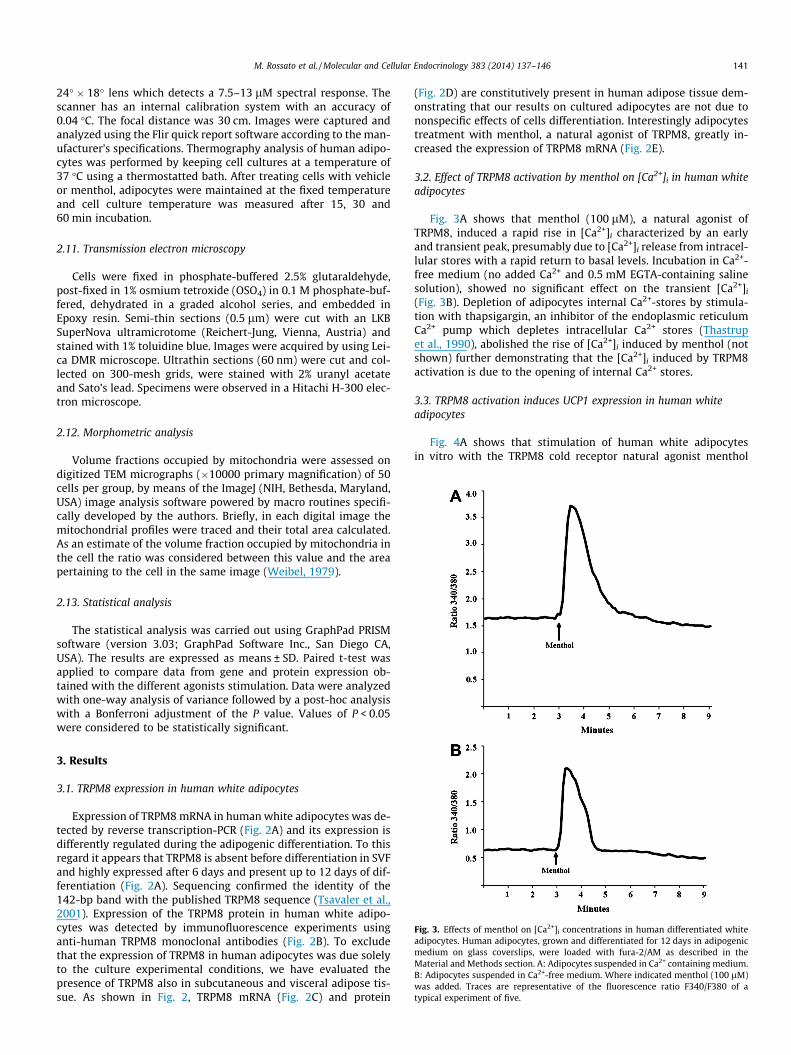
Fig. 2. Expression of TRPM8 in human white adipocytes. A: Quantitative PCR analysisin vitro adipogenic differentiation. TRPM8 mRNA was detected by polimerase chain reacand T12) in adipogenic medium; data are means ± SD of five separate experiments; Mpresence and localization of TRPM8 in human mature white adipocytes after 12 daysanalysis of TRPM8 mRNA expression in human subcutaneous (SAT) and visceral (VAT) arepresents the negative control (no sample); lane M, molecular marker. D: Western blottissue. E: Upregulation of TRPM8 expression by menthol stimulation of human whitemeans ± SD of five separate experiments. �P < 0.05, ��P < 0.01.
potential-dependent accumulation in mitochondria, indicated by afluorescence emission shift from green (514–529 nm) to red (585–590 nm). In living cells JC-1 forms green fluorescent monomers atdepolarized mitochondrial membrane potential, or red-fluorescentaggregates at hyperpolarized mitochondrial membrane potential.A higher red:green ratio indicates a more polarized, or more nega-tive and hyperpolarized mitochondrial inner membrane (Reerset al., 1991). For mitochondria functional studies, human adipo-cytes were treated with 100 lM menthol for different times (15,30 and 60 min), loaded with the mitochondrial membrane poten-tial indicator JC-1 10 lM for 20 min at 37 �C (Molecular Probes,Invitrogen). After incubation adipocytes were washed free of thedye and analyzed by means of a standard epifluorescencemicroscopy.
2.10. Infrared thermography
Heat generation by human adipocytes in vitro was measured bymeans of a thermo-electrically cooled Thermacam P25 (Flir Sys-tems Inc., Wilsonville, OR, USA) equipped with a scanner and
of TRPM8 mRNA expression in primary culture of human white adipocytes alongtion in stromal vascular fraction cells after 0, 6 and 12 day of differentiation (T0, T6
is the molecular marker; �P < 0.01. B: Immunofluorescence staining showing theof differentiation of stromal vascular fraction cells in adipogenic medium. C: PCRdipose tissue; human prostate gland (P) represents the positive control and saline
analysis of TRPM8 protein in human subcutaneous (SAT) and visceral (VAT) adiposeadipocytes after 12 days in vitro differentiation in adipogenic medium. Data are
M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146 141
24� � 18� lens which detects a 7.5–13 lM spectral response. Thescanner has an internal calibration system with an accuracy of0.04 �C. The focal distance was 30 cm. Images were captured andanalyzed using the Flir quick report software according to the man-ufacturer’s specifications. Thermography analysis of human adipo-cytes was performed by keeping cell cultures at a temperature of37 �C using a thermostatted bath. After treating cells with vehicleor menthol, adipocytes were maintained at the fixed temperatureand cell culture temperature was measured after 15, 30 and60 min incubation.
2.11. Transmission electron microscopy
Cells were fixed in phosphate-buffered 2.5% glutaraldehyde,post-fixed in 1% osmium tetroxide (OSO4) in 0.1 M phosphate-buf-fered, dehydrated in a graded alcohol series, and embedded inEpoxy resin. Semi-thin sections (0.5 lm) were cut with an LKBSuperNova ultramicrotome (Reichert-Jung, Vienna, Austria) andstained with 1% toluidine blue. Images were acquired by using Lei-ca DMR microscope. Ultrathin sections (60 nm) were cut and col-lected on 300-mesh grids, were stained with 2% uranyl acetateand Sato’s lead. Specimens were observed in a Hitachi H-300 elec-tron microscope.
2.12. Morphometric analysis
Volume fractions occupied by mitochondria were assessed ondigitized TEM micrographs (�10000 primary magnification) of 50cells per group, by means of the ImageJ (NIH, Bethesda, Maryland,USA) image analysis software powered by macro routines specifi-cally developed by the authors. Briefly, in each digital image themitochondrial profiles were traced and their total area calculated.As an estimate of the volume fraction occupied by mitochondria inthe cell the ratio was considered between this value and the areapertaining to the cell in the same image (Weibel, 1979).
2.13. Statistical analysis
The statistical analysis was carried out using GraphPad PRISMsoftware (version 3.03; GraphPad Software Inc., San Diego CA,USA). The results are expressed as means ± SD. Paired t-test wasapplied to compare data from gene and protein expression ob-tained with the different agonists stimulation. Data were analyzedwith one-way analysis of variance followed by a post-hoc analysiswith a Bonferroni adjustment of the P value. Values of P < 0.05were considered to be statistically significant.
Fig. 3. Effects of menthol on [Ca2+]i concentrations in human differentiated whiteadipocytes. Human adipocytes, grown and differentiated for 12 days in adipogenicmedium on glass coverslips, were loaded with fura-2/AM as described in theMaterial and Methods section. A: Adipocytes suspended in Ca2+ containing medium.B: Adipocytes suspended in Ca2+-free medium. Where indicated menthol (100 lM)was added. Traces are representative of the fluorescence ratio F340/F380 of atypical experiment of five.
3. Results
3.1. TRPM8 expression in human white adipocytes
Expression of TRPM8 mRNA in human white adipocytes was de-tected by reverse transcription-PCR (Fig. 2A) and its expression isdifferently regulated during the adipogenic differentiation. To thisregard it appears that TRPM8 is absent before differentiation in SVFand highly expressed after 6 days and present up to 12 days of dif-ferentiation (Fig. 2A). Sequencing confirmed the identity of the142-bp band with the published TRPM8 sequence (Tsavaler et al.,2001). Expression of the TRPM8 protein in human white adipo-cytes was detected by immunofluorescence experiments usinganti-human TRPM8 monoclonal antibodies (Fig. 2B). To excludethat the expression of TRPM8 in human adipocytes was due solelyto the culture experimental conditions, we have evaluated thepresence of TRPM8 also in subcutaneous and visceral adipose tis-sue. As shown in Fig. 2, TRPM8 mRNA (Fig. 2C) and protein
(Fig. 2D) are constitutively present in human adipose tissue dem-onstrating that our results on cultured adipocytes are not due tononspecific effects of cells differentiation. Interestingly adipocytestreatment with menthol, a natural agonist of TRPM8, greatly in-creased the expression of TRPM8 mRNA (Fig. 2E).
3.2. Effect of TRPM8 activation by menthol on [Ca2+]i in human whiteadipocytes
Fig. 3A shows that menthol (100 lM), a natural agonist ofTRPM8, induced a rapid rise in [Ca2+]i characterized by an earlyand transient peak, presumably due to [Ca2+]i release from intracel-lular stores with a rapid return to basal levels. Incubation in Ca2+-free medium (no added Ca2+ and 0.5 mM EGTA-containing salinesolution), showed no significant effect on the transient [Ca2+]i
(Fig. 3B). Depletion of adipocytes internal Ca2+-stores by stimula-tion with thapsigargin, an inhibitor of the endoplasmic reticulumCa2+ pump which depletes intracellular Ca2+ stores (Thastrupet al., 1990), abolished the rise of [Ca2+]i induced by menthol (notshown) further demonstrating that the [Ca2+]i induced by TRPM8activation is due to the opening of internal Ca2+ stores.
3.3. TRPM8 activation induces UCP1 expression in human whiteadipocytes
Fig. 4A shows that stimulation of human white adipocytesin vitro with the TRPM8 cold receptor natural agonist menthol
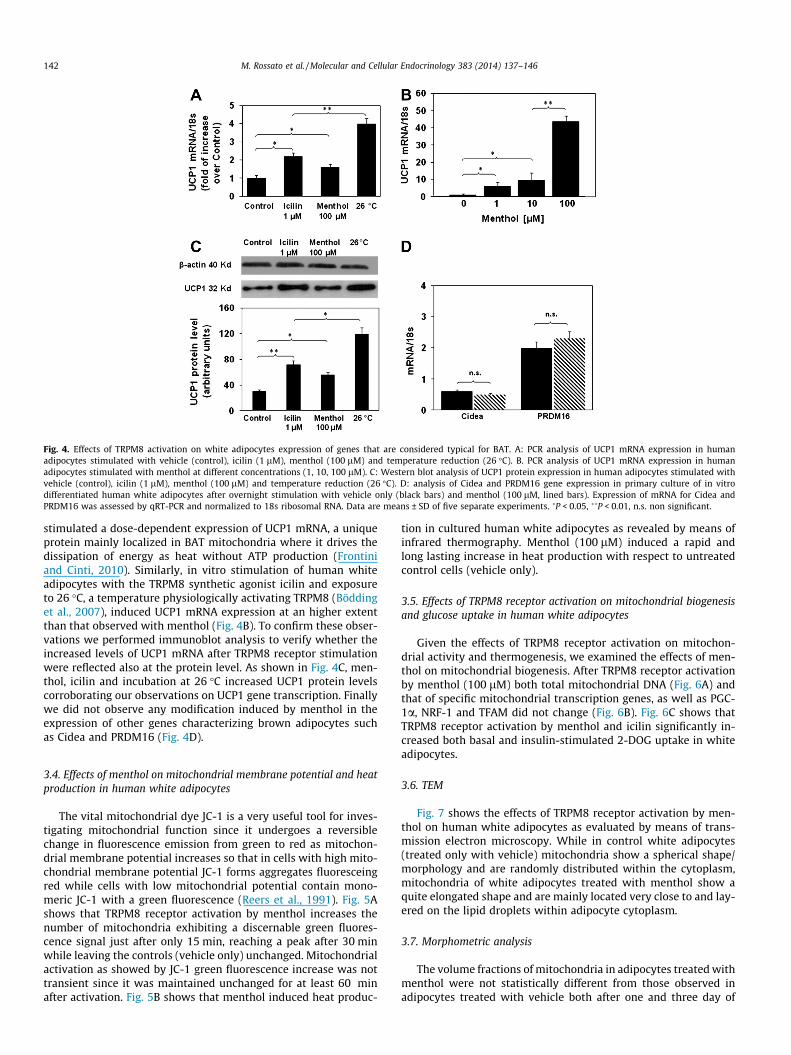
Fig. 4. Effects of TRPM8 activation on white adipocytes expression of genes that are considered typical for BAT. A: PCR analysis of UCP1 mRNA expression in humanadipocytes stimulated with vehicle (control), icilin (1 lM), menthol (100 lM) and temperature reduction (26 �C). B. PCR analysis of UCP1 mRNA expression in humanadipocytes stimulated with menthol at different concentrations (1, 10, 100 lM). C: Western blot analysis of UCP1 protein expression in human adipocytes stimulated withvehicle (control), icilin (1 lM), menthol (100 lM) and temperature reduction (26 �C). D: analysis of Cidea and PRDM16 gene expression in primary culture of in vitrodifferentiated human white adipocytes after overnight stimulation with vehicle only (black bars) and menthol (100 lM, lined bars). Expression of mRNA for Cidea andPRDM16 was assessed by qRT-PCR and normalized to 18s ribosomal RNA. Data are means ± SD of five separate experiments. �P < 0.05, ��P < 0.01, n.s. non significant.
142 M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146
stimulated a dose-dependent expression of UCP1 mRNA, a uniqueprotein mainly localized in BAT mitochondria where it drives thedissipation of energy as heat without ATP production (Frontiniand Cinti, 2010). Similarly, in vitro stimulation of human whiteadipocytes with the TRPM8 synthetic agonist icilin and exposureto 26 �C, a temperature physiologically activating TRPM8 (Böddinget al., 2007), induced UCP1 mRNA expression at an higher extentthan that observed with menthol (Fig. 4B). To confirm these obser-vations we performed immunoblot analysis to verify whether theincreased levels of UCP1 mRNA after TRPM8 receptor stimulationwere reflected also at the protein level. As shown in Fig. 4C, men-thol, icilin and incubation at 26 �C increased UCP1 protein levelscorroborating our observations on UCP1 gene transcription. Finallywe did not observe any modification induced by menthol in theexpression of other genes characterizing brown adipocytes suchas Cidea and PRDM16 (Fig. 4D).
3.4. Effects of menthol on mitochondrial membrane potential and heatproduction in human white adipocytes
The vital mitochondrial dye JC-1 is a very useful tool for inves-tigating mitochondrial function since it undergoes a reversiblechange in fluorescence emission from green to red as mitochon-drial membrane potential increases so that in cells with high mito-chondrial membrane potential JC-1 forms aggregates fluoresceingred while cells with low mitochondrial potential contain mono-meric JC-1 with a green fluorescence (Reers et al., 1991). Fig. 5Ashows that TRPM8 receptor activation by menthol increases thenumber of mitochondria exhibiting a discernable green fluores-cence signal just after only 15 min, reaching a peak after 30 minwhile leaving the controls (vehicle only) unchanged. Mitochondrialactivation as showed by JC-1 green fluorescence increase was nottransient since it was maintained unchanged for at least 60 minafter activation. Fig. 5B shows that menthol induced heat produc-
tion in cultured human white adipocytes as revealed by means ofinfrared thermography. Menthol (100 lM) induced a rapid andlong lasting increase in heat production with respect to untreatedcontrol cells (vehicle only).
3.5. Effects of TRPM8 receptor activation on mitochondrial biogenesisand glucose uptake in human white adipocytes
Given the effects of TRPM8 receptor activation on mitochon-drial activity and thermogenesis, we examined the effects of men-thol on mitochondrial biogenesis. After TRPM8 receptor activationby menthol (100 lM) both total mitochondrial DNA (Fig. 6A) andthat of specific mitochondrial transcription genes, as well as PGC-1a, NRF-1 and TFAM did not change (Fig. 6B). Fig. 6C shows thatTRPM8 receptor activation by menthol and icilin significantly in-creased both basal and insulin-stimulated 2-DOG uptake in whiteadipocytes.
3.6. TEM
Fig. 7 shows the effects of TRPM8 receptor activation by men-thol on human white adipocytes as evaluated by means of trans-mission electron microscopy. While in control white adipocytes(treated only with vehicle) mitochondria show a spherical shape/morphology and are randomly distributed within the cytoplasm,mitochondria of white adipocytes treated with menthol show aquite elongated shape and are mainly located very close to and lay-ered on the lipid droplets within adipocyte cytoplasm.
3.7. Morphometric analysis
The volume fractions of mitochondria in adipocytes treated withmenthol were not statistically different from those observed inadipocytes treated with vehicle both after one and three day of
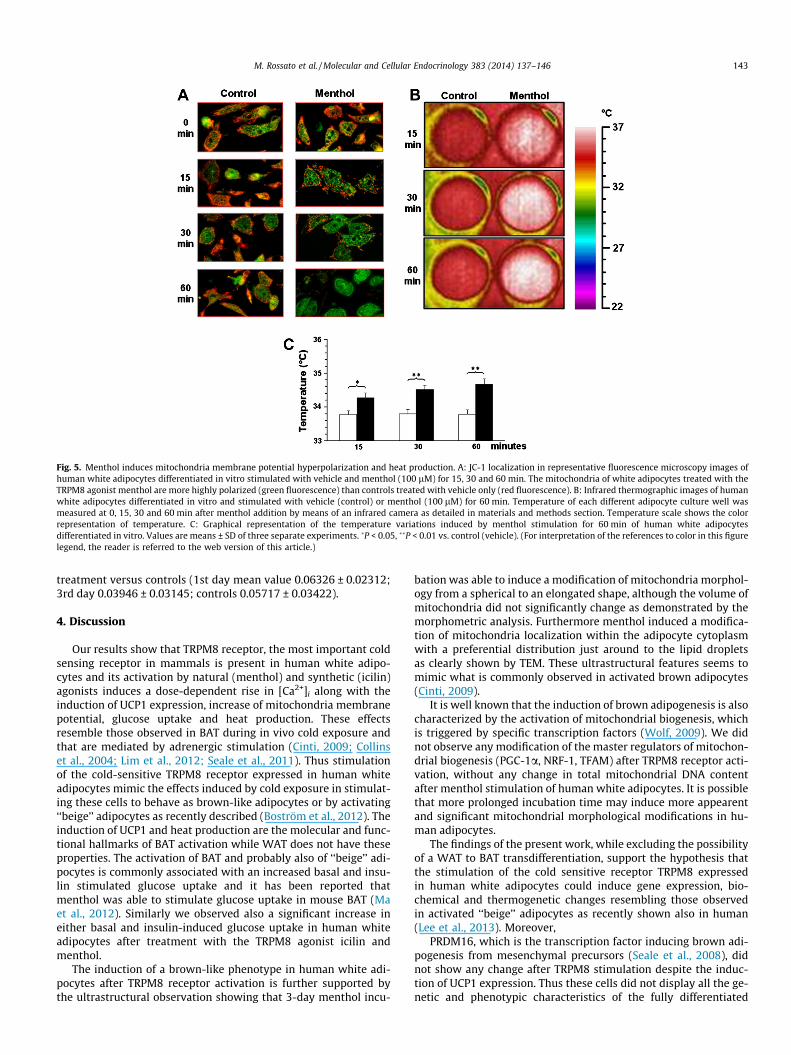
Fig. 5. Menthol induces mitochondria membrane potential hyperpolarization and heat production. A: JC-1 localization in representative fluorescence microscopy images ofhuman white adipocytes differentiated in vitro stimulated with vehicle and menthol (100 lM) for 15, 30 and 60 min. The mitochondria of white adipocytes treated with theTRPM8 agonist menthol are more highly polarized (green fluorescence) than controls treated with vehicle only (red fluorescence). B: Infrared thermographic images of humanwhite adipocytes differentiated in vitro and stimulated with vehicle (control) or menthol (100 lM) for 60 min. Temperature of each different adipocyte culture well wasmeasured at 0, 15, 30 and 60 min after menthol addition by means of an infrared camera as detailed in materials and methods section. Temperature scale shows the colorrepresentation of temperature. C: Graphical representation of the temperature variations induced by menthol stimulation for 60 min of human white adipocytesdifferentiated in vitro. Values are means ± SD of three separate experiments. �P < 0.05, ��P < 0.01 vs. control (vehicle). (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146 143
treatment versus controls (1st day mean value 0.06326 ± 0.02312;3rd day 0.03946 ± 0.03145; controls 0.05717 ± 0.03422).
4. Discussion
Our results show that TRPM8 receptor, the most important coldsensing receptor in mammals is present in human white adipo-cytes and its activation by natural (menthol) and synthetic (icilin)agonists induces a dose-dependent rise in [Ca2+]i along with theinduction of UCP1 expression, increase of mitochondria membranepotential, glucose uptake and heat production. These effectsresemble those observed in BAT during in vivo cold exposure andthat are mediated by adrenergic stimulation (Cinti, 2009; Collinset al., 2004; Lim et al., 2012; Seale et al., 2011). Thus stimulationof the cold-sensitive TRPM8 receptor expressed in human whiteadipocytes mimic the effects induced by cold exposure in stimulat-ing these cells to behave as brown-like adipocytes or by activating‘‘beige’’ adipocytes as recently described (Boström et al., 2012). Theinduction of UCP1 and heat production are the molecular and func-tional hallmarks of BAT activation while WAT does not have theseproperties. The activation of BAT and probably also of ‘‘beige’’ adi-pocytes is commonly associated with an increased basal and insu-lin stimulated glucose uptake and it has been reported thatmenthol was able to stimulate glucose uptake in mouse BAT (Maet al., 2012). Similarly we observed also a significant increase ineither basal and insulin-induced glucose uptake in human whiteadipocytes after treatment with the TRPM8 agonist icilin andmenthol.
The induction of a brown-like phenotype in human white adi-pocytes after TRPM8 receptor activation is further supported bythe ultrastructural observation showing that 3-day menthol incu-
bation was able to induce a modification of mitochondria morphol-ogy from a spherical to an elongated shape, although the volume ofmitochondria did not significantly change as demonstrated by themorphometric analysis. Furthermore menthol induced a modifica-tion of mitochondria localization within the adipocyte cytoplasmwith a preferential distribution just around to the lipid dropletsas clearly shown by TEM. These ultrastructural features seems tomimic what is commonly observed in activated brown adipocytes(Cinti, 2009).
It is well known that the induction of brown adipogenesis is alsocharacterized by the activation of mitochondrial biogenesis, whichis triggered by specific transcription factors (Wolf, 2009). We didnot observe any modification of the master regulators of mitochon-drial biogenesis (PGC-1a, NRF-1, TFAM) after TRPM8 receptor acti-vation, without any change in total mitochondrial DNA contentafter menthol stimulation of human white adipocytes. It is possiblethat more prolonged incubation time may induce more appearentand significant mitochondrial morphological modifications in hu-man adipocytes.
The findings of the present work, while excluding the possibilityof a WAT to BAT transdifferentiation, support the hypothesis thatthe stimulation of the cold sensitive receptor TRPM8 expressedin human white adipocytes could induce gene expression, bio-chemical and thermogenetic changes resembling those observedin activated ‘‘beige’’ adipocytes as recently shown also in human(Lee et al., 2013). Moreover,
PRDM16, which is the transcription factor inducing brown adi-pogenesis from mesenchymal precursors (Seale et al., 2008), didnot show any change after TRPM8 stimulation despite the induc-tion of UCP1 expression. Thus these cells did not display all the ge-netic and phenotypic characteristics of the fully differentiated
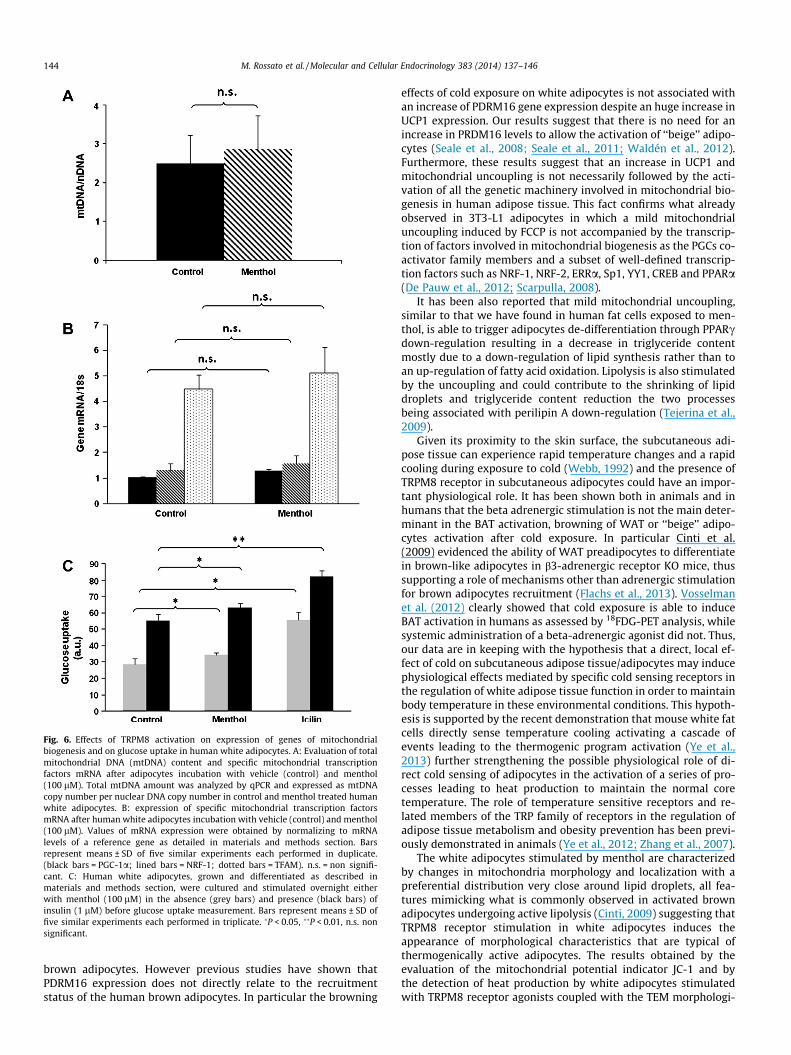
Fig. 6. Effects of TRPM8 activation on expression of genes of mitochondrialbiogenesis and on glucose uptake in human white adipocytes. A: Evaluation of totalmitochondrial DNA (mtDNA) content and specific mitochondrial transcriptionfactors mRNA after adipocytes incubation with vehicle (control) and menthol(100 lM). Total mtDNA amount was analyzed by qPCR and expressed as mtDNAcopy number per nuclear DNA copy number in control and menthol treated humanwhite adipocytes. B: expression of specific mitochondrial transcription factorsmRNA after human white adipocytes incubation with vehicle (control) and menthol(100 lM). Values of mRNA expression were obtained by normalizing to mRNAlevels of a reference gene as detailed in materials and methods section. Barsrepresent means ± SD of five similar experiments each performed in duplicate.(black bars = PGC-1a; lined bars = NRF-1; dotted bars = TFAM). n.s. = non signifi-cant. C: Human white adipocytes, grown and differentiated as described inmaterials and methods section, were cultured and stimulated overnight eitherwith menthol (100 lM) in the absence (grey bars) and presence (black bars) ofinsulin (1 lM) before glucose uptake measurement. Bars represent means ± SD offive similar experiments each performed in triplicate. �P < 0.05, ��P < 0.01, n.s. nonsignificant.
144 M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146
brown adipocytes. However previous studies have shown thatPDRM16 expression does not directly relate to the recruitmentstatus of the human brown adipocytes. In particular the browning
effects of cold exposure on white adipocytes is not associated withan increase of PDRM16 gene expression despite an huge increase inUCP1 expression. Our results suggest that there is no need for anincrease in PRDM16 levels to allow the activation of ‘‘beige’’ adipo-cytes (Seale et al., 2008; Seale et al., 2011; Waldén et al., 2012).Furthermore, these results suggest that an increase in UCP1 andmitochondrial uncoupling is not necessarily followed by the acti-vation of all the genetic machinery involved in mitochondrial bio-genesis in human adipose tissue. This fact confirms what alreadyobserved in 3T3-L1 adipocytes in which a mild mitochondrialuncoupling induced by FCCP is not accompanied by the transcrip-tion of factors involved in mitochondrial biogenesis as the PGCs co-activator family members and a subset of well-defined transcrip-tion factors such as NRF-1, NRF-2, ERRa, Sp1, YY1, CREB and PPARa(De Pauw et al., 2012; Scarpulla, 2008).
It has been also reported that mild mitochondrial uncoupling,similar to that we have found in human fat cells exposed to men-thol, is able to trigger adipocytes de-differentiation through PPARcdown-regulation resulting in a decrease in triglyceride contentmostly due to a down-regulation of lipid synthesis rather than toan up-regulation of fatty acid oxidation. Lipolysis is also stimulatedby the uncoupling and could contribute to the shrinking of lipiddroplets and triglyceride content reduction the two processesbeing associated with perilipin A down-regulation (Tejerina et al.,2009).
Given its proximity to the skin surface, the subcutaneous adi-pose tissue can experience rapid temperature changes and a rapidcooling during exposure to cold (Webb, 1992) and the presence ofTRPM8 receptor in subcutaneous adipocytes could have an impor-tant physiological role. It has been shown both in animals and inhumans that the beta adrenergic stimulation is not the main deter-minant in the BAT activation, browning of WAT or ‘‘beige’’ adipo-cytes activation after cold exposure. In particular Cinti et al.(2009) evidenced the ability of WAT preadipocytes to differentiatein brown-like adipocytes in b3-adrenergic receptor KO mice, thussupporting a role of mechanisms other than adrenergic stimulationfor brown adipocytes recruitment (Flachs et al., 2013). Vosselmanet al. (2012) clearly showed that cold exposure is able to induceBAT activation in humans as assessed by 18FDG-PET analysis, whilesystemic administration of a beta-adrenergic agonist did not. Thus,our data are in keeping with the hypothesis that a direct, local ef-fect of cold on subcutaneous adipose tissue/adipocytes may inducephysiological effects mediated by specific cold sensing receptors inthe regulation of white adipose tissue function in order to maintainbody temperature in these environmental conditions. This hypoth-esis is supported by the recent demonstration that mouse white fatcells directly sense temperature cooling activating a cascade ofevents leading to the thermogenic program activation (Ye et al.,2013) further strengthening the possible physiological role of di-rect cold sensing of adipocytes in the activation of a series of pro-cesses leading to heat production to maintain the normal coretemperature. The role of temperature sensitive receptors and re-lated members of the TRP family of receptors in the regulation ofadipose tissue metabolism and obesity prevention has been previ-ously demonstrated in animals (Ye et al., 2012; Zhang et al., 2007).
The white adipocytes stimulated by menthol are characterizedby changes in mitochondria morphology and localization with apreferential distribution very close around lipid droplets, all fea-tures mimicking what is commonly observed in activated brownadipocytes undergoing active lipolysis (Cinti, 2009) suggesting thatTRPM8 receptor stimulation in white adipocytes induces theappearance of morphological characteristics that are typical ofthermogenically active adipocytes. The results obtained by theevaluation of the mitochondrial potential indicator JC-1 and bythe detection of heat production by white adipocytes stimulatedwith TRPM8 receptor agonists coupled with the TEM morphologi-
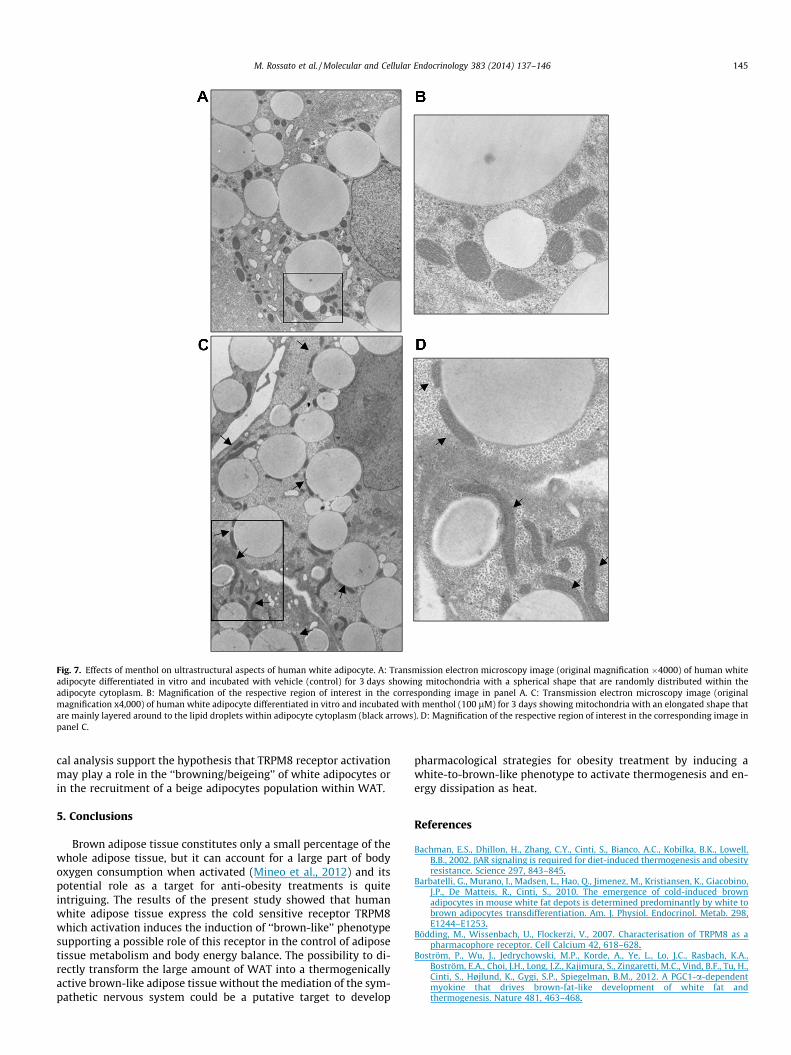
Fig. 7. Effects of menthol on ultrastructural aspects of human white adipocyte. A: Transmission electron microscopy image (original magnification �4000) of human whiteadipocyte differentiated in vitro and incubated with vehicle (control) for 3 days showing mitochondria with a spherical shape that are randomly distributed within theadipocyte cytoplasm. B: Magnification of the respective region of interest in the corresponding image in panel A. C: Transmission electron microscopy image (originalmagnification x4,000) of human white adipocyte differentiated in vitro and incubated with menthol (100 lM) for 3 days showing mitochondria with an elongated shape thatare mainly layered around to the lipid droplets within adipocyte cytoplasm (black arrows). D: Magnification of the respective region of interest in the corresponding image inpanel C.
M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146 145
cal analysis support the hypothesis that TRPM8 receptor activationmay play a role in the ‘‘browning/beigeing’’ of white adipocytes orin the recruitment of a beige adipocytes population within WAT.
5. Conclusions
Brown adipose tissue constitutes only a small percentage of thewhole adipose tissue, but it can account for a large part of bodyoxygen consumption when activated (Mineo et al., 2012) and itspotential role as a target for anti-obesity treatments is quiteintriguing. The results of the present study showed that humanwhite adipose tissue express the cold sensitive receptor TRPM8which activation induces the induction of ‘‘brown-like’’ phenotypesupporting a possible role of this receptor in the control of adiposetissue metabolism and body energy balance. The possibility to di-rectly transform the large amount of WAT into a thermogenicallyactive brown-like adipose tissue without the mediation of the sym-pathetic nervous system could be a putative target to develop
pharmacological strategies for obesity treatment by inducing awhite-to-brown-like phenotype to activate thermogenesis and en-ergy dissipation as heat.
References
Bachman, E.S., Dhillon, H., Zhang, C.Y., Cinti, S., Bianco, A.C., Kobilka, B.K., Lowell,B.B., 2002. bAR signaling is required for diet-induced thermogenesis and obesityresistance. Science 297, 843–845.
Barbatelli, G., Murano, I., Madsen, L., Hao, Q., Jimenez, M., Kristiansen, K., Giacobino,J.P., De Matteis, R., Cinti, S., 2010. The emergence of cold-induced brownadipocytes in mouse white fat depots is determined predominantly by white tobrown adipocytes transdifferentiation. Am. J. Physiol. Endocrinol. Metab. 298,E1244–E1253.
Bödding, M., Wissenbach, U., Flockerzi, V., 2007. Characterisation of TRPM8 as apharmacophore receptor. Cell Calcium 42, 618–628.
Boström, P., Wu, J., Jedrychowski, M.P., Korde, A., Ye, L., Lo, J.C., Rasbach, K.A.,Boström, E.A., Choi, J.H., Long, J.Z., Kajimura, S., Zingaretti, M.C., Vind, B.F., Tu, H.,Cinti, S., Højlund, K., Gygi, S.P., Spiegelman, B.M., 2012. A PGC1-a-dependentmyokine that drives brown-fat-like development of white fat andthermogenesis. Nature 481, 463–468.

146 M. Rossato et al. / Molecular and Cellular Endocrinology 383 (2014) 137–146
Cho, Y., Jang, Y., Yang, Y.D., Lee, C.H., Lee, Y., Oh, U., 2010. TRPM8 mediates cold andmenthol allergies associated with mast cell activation. Cell Calcium 48, 202–208.
Cinti, S., 2005. The adipose organ. Prostaglandins Leukot. Essent. Fatty Acids 73, 9–15.
Cinti, S., 2009. Transdifferentiation properties of adipocytes in the adipose organ.Am. J. Physiol. Endocrinol. Metab. 297, E977–E986.
Collins, S., Cao, W., Robidoux, J., 2004. Learning new tricks from old dogs: beta-adrenergic receptors teach new lessons on firing up adipose tissue metabolism.Mol. Endocrinol. 18, 2123–2131.
Cypess, A.M., Lehman, S., Williams, G., Tal, I., Rodman, D., Goldfine, A.B., Kuo, F.C.,Palmer, E.L., Tseng, Y.H., Doria, A., Kolodny, G.M., Kahn, C.R., 2009. Identificationand importance of brown adipose tissue in adult humans. New Engl. J. Med.360, 1509–1517.
De Blas, G.A., Darszon, A., Ocampo, A.Y., Serrano, C.J., Castellano, L.E., Hernández-González, E.O., Chirinos, M., Larrea, F., Beltrán, C., Treviño, C.L., 2009. TRPM8, aversatile channel in human sperm. PLoS ONE 4, e6095.
De Pauw, A., Demine, S., Tejerina, S., Dieu, M., Delaive, E., Kel, A., Renard, P., Raes, M.,Arnould, T., 2012. Mild mitochondrial uncoupling does not affect mitochondrialbiogenesis but downregulates pyruvate carboxylase in adipocytes: role fortriglyceride content reduction. Am. J. Physiol. Endocrinol. Metab. 302, E1123–E1141.
Dhennin-Duthille, I., Gautier, M., Faouzi, M., Guilbert, A., Brevet, M., Vaudry, D.,Ahidouch, A., Sevestre, H., Ouadid-Ahidouch, H., 2011. High expression oftransient receptor potential channels in human breast cancer epithelial cellsand tissues: correlation with pathological parameters. Cell. Physiol. Biochem.28, 813–822.
Flachs, P., Rossmeisl, M., Kuda, O., Kopecky, J., 2013. Stimulation of mitochondrialoxidative capacity in white fat independent of UCP1: a key to lean phenotype.Biochim. Biophys. Acta 1831, 986–1003.
Frontini, A., Cinti, S., 2010. Distribution and development of brown adipocytes in themurine and human adipose organ. Cell Metab. 11, 253–256.
Himms-Hagen, J., Melnyk, A., Zingaretti, M.C., Ceresi, E., Barbatelli, G., Cinti, S., 2000.Multilocular fat cells in WAT of CL-316243-treated rats derive directly fromwhite adipocytes. Am. J. Physiol. Cell Physiol. 279, C670–C681.
Knowlton, W.M., McKemy, D.D., 2011. TRPM8: from cold to cancer, peppermint topain. Curr. Pharm. Biotechnol. 12, 68–77.
Kopecky, J., Clarke, G., Enerback, S., Spiegelman, B., Kozak, L.P., 1995. Expression ofthe mitochondrial uncoupling protein gene from the aP2 gene promoterprevents genetic obesity. J. Clin. Invest. 96, 2914–2923.
Lee, P., Werner, C.D., Kebebew, E., Celi, F.S., 2013. Functional thermogenic beigeadipogenesis is inducible in human neck fat. Int. J. Obes. Int. J. Obes., http://dx.doi.org/10.1038/ijo.2013.82.
Li, Q., Wang, X., Yang, Z., Wang, B., Li, S., 2009. Menthol induces cell death via theTRPM8 channel in the human bladder cancer cell line T24. Oncology 77, 335–341.
Lim, S., Honek, J., Xue, Y., Seki, T., Cao, Z., Andersson, P., Yang, X., Hosaka, K., Cao, Y.,2012. Cold-induced activation of brown adipose tissue and adiposeangiogenesis in mice. Nat. Protoc. 7, 606–615.
Lowell, B.B., S-Susulic, V., Hamann, A., Lawitts, J.A., Himms-Hagen, J., Boyer, B.B.,Kozak, L.P., Flier, J.S., 1993. Development of obesity in transgenic mice aftergenetic ablation of brown adipose tissue. Nature 366, 740–742.
Ma, S., Yu, H., Zhao, Z., Luo, Z., Chen, J., Ni, Y., Jin, R., Ma, L., Wang, P., Zhu, Z., Li, L.,Zhong, J., Liu, D., Nilius, B., Zhu, Z., 2012. Activation of the cold-sensing TRPM8channel triggers UCP1 dependent thermogenesis and prevents obesity. J. Mol.Cell. Biol. 4, 88–96.
McKemy, D.D., Neuhausser, W.M., Julius, D., 2002. Identification of a coldreceptor reveals a general role for TRP channels in thermosensation. Nature416, 52–58.
Mineo, P.M., Cassell, E.A., Roberts, M.E., Schaeffer, P.J., 2012. Chronic coldacclimation increases thermogenic capacity, non-shivering thermogenesis andmuscle citrate synthase activity in both wild-type and brown adipose tissuedeficient mice. Comp. Biochem. Physiol. A: Mol. Integr. Physiol. 161, 395–400.
Nilius, B., Owsianik, G., Voets, T., Peters, J.A., 2007. Transient receptor potentialcation channels in disease. Physiol. Rev. 87, 165–217.
Owsianik, G., D’Hoedt, D., Voets, T., Nilius, B., 2006. Structure-function relationshipof the TRP channel superfamily. Rev. Physiol. Biochem. Pharmacol. 156, 61–90.
Pagano, C., Pilon, C., Calcagno, A., Urbanet, R., Rossato, M., Milan, G., Bianchi, K.,Rizzuto, R., Bernante, P., Federspil, G., Vettor, R., 2007. The endogenouscannabinoid system stimulates glucose uptake in human fat cells viaphosphatidylinositol 3-kinase and calcium-dependent mechanisms. J. Clin.Endocrinol. Metab. 92, 4810–4819.
Reers, M., Smith, T.W., Chen, L.B., 1991. J-aggregate formation of a carbocyanine as aquantitative fluorescent indicator of membrane potential. Biochemistry 30,4480–4486.
Rybarczyk, P., Gautier, M., Hague, F., Dhennin-Duthille, I., Chatelain, D., Kerr-Conte,J., Pattou, F., Regimbeau, J.M., Sevestre, H., Ouadid-Ahidouch, H., 2012. Transientreceptor potential melastatin related 7 channel is overexpressed in human
pancreatic ductal adenocarcinomas and regulates human pancreatic cancer cellmigration. Int. J. Cancer 131, E851–E861.
Scarda, A., Franzin, C., Milan, G., Sanna, M., Dalprà, C., Pagano, C., Boldrin, L., Piccoli,M., Trevellin, E., Granzotto, M., Gamba, P., Federspil, G., De Coppi, P., Vettor, R.,2010. Increased adipogenic conversion of muscle satellite cells in obese Zuckerrats. Int. J. Obes. 34, 1319–1327.
Scarpulla, R.C., 2008. Transcriptional paradigms in mammalian mitochondrialbiogenesis and function. Physiol. Rev. 88, 611–638.
Seale, P., Bjork, B., Yang, W., Kajimura, S., Chin, S., Kuang, S., Scimè, A., Devarakonda,S., Conroe, H.M., Erdjument-Bromage, H., Tempst, P., Rudnicki, M.A., Beier, D.R.,Spiegelman, B.M., 2008. PRDM16 controls a brown fat/skeletal muscle switch.Nature 454, 961–967.
Seale, P., Conroe, H.M., Estall, J., Kajimura, S., Frontini, A., Ishibashi, J., Cohen, P.,Cinti, S., Spiegelman, B.M., 2011. PRDM16 determines the thermogenic programof subcutaneous white adipose tissue in mice. J. Clin. Invest. 121, 96–105.
Talavera, K., Nilius, B., Voets, T., 2008. Neuronal TRP channels: thermometers,pathfinders and life-savers. Trends Neurosci. 31, 287–295.
Tejerina, S., De Pauw, A., Vankoningsloo, S., Houbion, A., Renard, P., De Longueville,F., Raes, M., Arnould, T., 2009. Mild mitochondrial uncoupling induces 3T3-L1adipocyte de-differentiation by a PPARc-independent mechanism, whereasTNFa-induced de-differentiation is PPARc dependent. J. Cell Sci. 122, 145–155.
Thastrup, O., Cullen, P.J., Drobak, B.K., Hanley, M.R., Dawson, A.P., 1990.Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores byspecific inhibition of the endoplasmic reticulum Ca2+-ATPase. Proc. Natl. Acad.Sci. USA 87, 2466–2470.
Tsavaler, L., Shapero, M.H., Morkowski, S., Laus, R., 2001. Trp-p8, a novel prostate-specific gene, is up-regulated in prostate cancer and other malignancies andshares high homology with transient receptor potential calcium channelproteins. Cancer Res. 61, 3760–3769.
Tyagi, P., 2011. Pathophysiology of the urothelium and detrusor. Can. Urol. Assoc. J.5 (Suppl 2), S128–S130.
Valero, M., Morenilla-Palao, C., Belmonte, C., Viana, F., 2011. Pharmacological andfunctional properties of TRPM8 channels in prostate tumor cells. Pflugers Arch.461, 99–114.
van Haute, C., De Ridder, D., Nilius, B., 2010. TRP channels in human prostate. Sci.World J. 10, 1597–1611.
van Marken Lichtenbelt, W.D., Vanhommerig, J.W., Smulders, N.M., Drossaerts, J.M.,Kemerink, G.J., Bouvy, N.D., Schrauwen, P., Teule, G.J., 2009. Cold-activatedbrown adipose tissue in healthy men. New Engl. J. Med. 360, 1500–1508.
Virtanen, K.A., Lidell, M.E., Orava, J., Heglind, M., Westergren, R., Niemi, T., Taittonen,M., Laine, J., Savisto, N.J., Enerbäck, S., Nuutila, P., 2009. Functional brownadipose tissue in healthy adults. N. Engl. J. Med. 360, 1518–1525.
Voets, T., Talavera, K., Owsianik, G., Nilius, B., 2005. Sensing with TRP channels. Nat.Chem. Biol. 1, 85–92.
Vosselman, M.J., van der Lans, A.A., Brans, B., Wierts, R., van Baak, M.A., Schrauwen,P., Lichtenbelt, W.D., 2012. Systemic b-adrenergic stimulation of thermogenesisis not accompanied by brown adipose tissue activity in humans. Diabetes 61,3106–3113.
Waldén, T.B., Hansen, I.R., Timmons, J.A., Cannon, B., Nedergaard, J., 2012. Recruitedvs. nonrecruited molecular signatures of brown, ‘‘brite’’, and white adiposetissues. Am. J. Physiol. Endocrinol. Metab. 302, E19–E31.
Webb, P., 1992. Temperatures of skin, subcutaneous tissue, muscle and core inresting men in cold, comfortable and hot conditions. Eur. J. Appl. Physiol. Occup.Physiol. 64, 471–476.
Weibel, E.R., 1979. Stereological Methods: Practical Methods for BiologicalMorphometry, vol. 1. Academic Press, London.
Wolf, G., 2009. Brown adipose tissue: the molecular mechanism of its formation.Nutr. Rev. 67, 167–171.
Wu, S.N., Wu, P.Y., Tsai, M.L., 2011. Characterization of TRPM8-like channelsactivated by the cooling agent icilin in the macrophage cell line RAW 264.7. J.Membr. Biol. 241, 11–20.
Wu, J., Boström, P., Sparks, L.M., Ye, L., Choi, J.H., Giang, A.H., Khandekar, M.,Virtanen, K.A., Nuutila, P., Schaart, G., Huang, K., Tu, H., van Marken Lichtenbelt,W.D., Hoeks, J., Enerbäck, S., Schrauwen, P., Spiegelman, B.M., 2012. Beigeadipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell150, 366–376.
Ye, L., Kleiner, S., Wu, J., Sah, R., Gupta, R.K., Banks, A.S., Cohen, P., Khandekar, M.J.,Boström, P., Mepani, R.J., Laznik, D., Kamenecka, T.M., Song, X., Liedtke, W.,Mootha, V.K., Puigserver, P., Griffin, P.R., Clapham, D.E., Spiegelman, B.M., 2012.TRPV4 is a regulator of adipose oxidative metabolism, inflammation, and energyhomeostasis. Cell 151, 96–110.
Ye, L., Wu, J., Cohen, P., Kazak, L., Khandekar, M.J., Jedrychowski, M.P., Zeng, X., Gygi,S.P., Spiegelman, B.M., 2013. Fat cells directly sense temperature to activatethermogenesis. Proc. Natl. Acad. Sci. USA 110, 12480–12485.
Zhang, L.L., Yan Liu, D., Ma, L.Q., Luo, Z.D., Cao, T.B., Zhong, J., Yan, Z.C., Wang, L.J.,Zhao, Z.G., Zhu, S.J., Schrader, M., Thilo, F., Zhu, Z.M., Tepel, M., 2007. Activationof transient receptor potential vanilloid type-1 channel prevents adipogenesisand obesity. Circ. Res. 100, 1063–1070.





























