Embed Size (px)
Citation preview

Hyaluronan involvement in the changes of mouse interpubic tissueduring late pregnancy and post-partum
Eduardo A. Garcia a, Adriano M. Veridiano a, Jo~ao R.M. Martins b, Helena B. Nader b,M.C. Pinheiro c, Paulo P. Joazeiro a, Olga M.S. Toledo c,*
a Department of Histology and Embryology, Institute of Biology, UNICAMP, Campinas, S~ao Paulo, Brazilb Department of Biochemistry, Division of Molecular Biology, UNIFESP, S~ao Paulo, Brazil
c Department of Morphology, Discipline of Histology and Structural Biology, Federal University of S~ao Paulo e UNIFESP,Rua Botucatu 740, 04023-062 S~ao Paulo, Brazil
Received 19 February 2007; revised 5 February 2008; accepted 2 April 2008
Abstract
The present work quantifies hyaluronan (HA) during the late pregnancy and post-partum in order to provide a better understanding of the roleof HA in the adaptations that occur in the pubic symphysis during this period. HA was quantified in situ (histochemically) and in interpubictissue extracts by fluorimetric assay. Samples were taken from virgin mice and from pregnant animals at various stages of pregnancy: 12the18th days into pregnancy, the day of delivery (D19) and the 3rd and 5th day post-partum. The quantitative fluorimetric analysis indicated a grad-ual increase of HA in the interpubic tissue throughout late pregnancy (2.4e14.6 mg/mg dry weight). This was followed by a decrease beginningon D19 (12.4 mg/mg), reaching close to virgin levels (2.2 mg/mg) on the 5th day post-partum. The same optical density changes could be seen inthe HA staining. Furthermore, the histochemical analysis demonstrated the presence of HA both in the extracellular matrix of the tissue andwithin its cells. Such results indicate that the extracellular presence of HA may contribute to the transformation of the symphysis into a flexiblestructure. In addition, HA’s intracellular presence (until the 18th day of pregnancy) may contribute to cellular proliferation. Finally, during par-turition and on the 5th day post-partum, HA may contribute to the maintenance of the myofibroblastic phenotype of ligament cells, aiding theligament involution after parturition.! 2008 International Federation for Cell Biology. Published by Elsevier Ltd. All rights reserved.
Keywords: Hyaluronan; Pubic symphysis; Histochemistry; HA-biotinylated probe; Pregnancy; Mice; HA-Elisa-like assay
1. Introduction
During mouse pregnancy the articular cartilage and the ex-tremities of the pubic bones are partially absorbed, leading tomodifications in the scarce interpubic connective tissue ob-served in virgin animals. These changes allow the separationof the pelvic bones, permitting an enlargement of the birth ca-nal during parturition to enable safe delivery of the young(Hall, 1947; Storey, 1957; Horn, 1960).
Such changes are noted by the rapid formation and pro-nounced swelling of a fibrous interpubic ligament, producing a
flaccid structure. The process continuously enlarges the pubicsymphysis and promotes rearrangement and increased synthe-sis of the extracellular matrix in this tissue (Storey, 1957;Viell and Struck, 1987; Samuel et al., 1996; Zhao et al.,2000; Ortega et al., 2001).
We have previously reported an extensive remodeling of theinterpubic tissue involving cells and different components ofthe extracellular matrix. Myofibroblast-like cells, for example,are the only cells which are present in the extension of mousepubic ligament during late pregnancy, delivery and post-par-tum (Moraes et al., 2004). It was also demonstrated that theincrease of the symphyseal gap from its original 0.15 mmlength in virgin animals to 2.4 mm in partum is a process inwhich new arrangements of collagen (Pinheiro et al., 2004)
* Corresponding author. Tel.: !55 11 55764268; fax: !55 11 55764271.E-mail address: [email protected] (O.M.S. Toledo).
1065-6995/$ - see front matter ! 2008 International Federation for Cell Biology. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.cellbi.2008.04.006
Cell Biology International 32 (2008) 913e919www.elsevier.com/locate/cellbi

and elastic fibers (Moraes et al., 2003), as well as other differ-ent types of proteoglycans, are involved (Pinheiro et al., 2005).
Another extracellular matrix component that is involved inthe transformations undertaken by the interpubic symphysis ishyaluronan (HA) (Zhao et al., 2000). This molecule is a linearglycosaminoglycan formed of repeating disaccharides of D-glucuronic acid and N-acetylglucosamine (Lee and Spicer,2000). Several functions have been assigned to HA, includingthe stabilization of the extracellular matrix, joint lubrification(Balazs and Denlinger, 1989), water homeostasis (Tammiet al., 2002) and regulation of cellular activities due to its in-teractions with cell surface receptor proteins (Lesley et al.,2000). Most recently, the intracellular presence of HA hasalso been related to the maintenance of the myofibroblasticphenotype of cells (Jenkins et al., 2004), as well as the forma-tion of a facilitated environment for cellular division (Hascallet al., 2004). Finally, other studies have shown that HA, whilecreating a hydrated zone around the cell, facilitates the detach-ment of the cell from its substratum, a necessary step for theprocesses of mitosis and cellular migration (Toole, 2001).
In addition, the increase of the interpubic connective tissuein mice has already been partly explained as a consequence ofmodifications observed both in the amount of HA and waterfound in this tissue (Zhao et al., 2000). Indeed, this moleculeis noted for its great capacity for holding water; it probablyparticipates in the great increase in water content in the inter-pubic ligament during pregnancy.
During pregnancy, this tissue must undergo drastic changesof swelling in order to provide a sufficiently wide and resistantbirth canal for safe delivery of the young, while still ensuringanatomical integrity of the pelvic girdle. The increase of waterwithin the interpubic tissue is crucial for the development ofa flexible structure during pregnancy. The flexibility of the lig-ament is critical during the last few days of pregnancy, whenthe ligament is continuously subject to both tensional andcompressive forces (Sherwood, 1994).
In our previous work, HA was detected in the interpubic tis-sues (fibrocartilage and ligament), notably in the interfibrousspaces and pericellular matrix. Subjected analyses suggesteda large amount of HA on D18, the day when the ‘relaxation’process of the interpubic ligament takes place (Pinheiroet al., 2005). Thus, the present work was undertaken to quan-tify HA during the late pregnancy and post-partum, thus pro-viding a better understating of the role of HA in theadaptations that occur in the symphysis during this period.For this reason, HA was measured both in situ (histochemi-cally) and in interpubic tissue extracts by fluorimetric assay.
2. Materials and methods
2.1. Tissue collection
Adult female virgin Swiss mice (approximately 90 daysold, weighting from 25 to 30 g) were housed at room temper-ature (20" 3 #C) under artificially controlled light from 06:00to 20:00 h. They were provided with laboratory rodent dry pel-lets and water ad libitum. Mating opportunities were provided
by caging the females overnight with breeding males. The vag-inal plug was checked each morning. The first day of preg-nancy was considered as the day (D) on which a vaginalplug was found; birth occurred on day 19.
Pubic symphyses were obtained from pregnant mice (10 an-imals per group) on days 12, 15, 17 and 18; immediately afterdelivery on day 19; and on the 3rd and 5th days post-partum.Six virgin mice were chosen as the control group. All animalexperiments were conducted in accordance with the Guidefor Care and Use of Laboratory Animals, issued by NIH/USA.
The pubic symphyses obtained were fixed in acideformaldehydeeethanol fixative pH 3.2 (Lin et al., 1997) andprocessed according to standard histological techniques forparaffin embedding. The picrosirius method (Montes, 1996)was used to examine the histology of the pubic symphyses.
2.2. HA distribution and measurement
2.2.1. HA-histochemical distributionThe quantification of HA in situ was done in paraffinized
pubic symphysis sections (obtained from specimens of five an-imals per group). All sections underwent the following treat-ment: (a) inhibition of endogenous peroxidase activity with3% hydrogen peroxidase in phosphate buffered saline (PBS)solution (Amersham Life Science, Buckinghamshire, UK);(b) blockage of non-specific binding sites with 1% bovine se-rum albumin (BSA) (Amersham Life Science) in PBS; (c) in-cubation in biotinylated probe solution diluted 1:150 in 1%BSA/PBS for 1 h at 4 #C; (d) elimination of endogenous biotinin tissue sections by incubation in streptavidin peroxidase so-lution (Amersham Life Science); and (e) visualization of per-oxidase activity through incubation with chromogenicdiaminobenzidine (DAB) (Sigma Co., St. Louis, MO) and0.03% hydrogen peroxidase.
In each treatment phase throughout the histochemical pro-cess, the sections were initially washed with PBS and then an-alyzed and photographed using a Nikon Eclipse E800 lightmicroscope. Negative controls were obtained by treatingsome sections with streptomyces hyaluronidase, which specif-ically degrades HA (Ohya and Kaneko, 1970), prior to incuba-tion with the probe. The quantification was done by evaluatingthe brownish color density (resulting from the binding of en-dogenous HA to the biotinylated probe).
The in situ quantification of the HA was done by recordinghistological images with a Sony Exware HAM color videocamera attached to the microscope, using a Plan Fluor 10$ ob-jective (NA% 0.30). The staining measurements were done bycomputer analysis, using Image Pro-Plus 4.1.0.1 software(Media Cybernetics, Silver Spring, MD).
2.2.2. Quantification of HA by fluorescence-based assayAn accurate fluorassay method (Martins et al., 2003; Lebl
et al., 2007) was used to quantify HA in the interpubic tissues(fibrocartilage and ligament). Tissue samples were collectedfrom four mice per group and processed as a pool to ensuresufficient amount of HA (at least 5 mg/dry tissue) for analysis.
914 E.A. Garcia et al. / Cell Biology International 32 (2008) 913e919

2.2.3. HA tissue extractionHA extraction was done in the following steps: (a) removal
of lipid residues by acetone; (b) drying and weighting of thedry tissue (ketonic powder); (c) proteic digestion with alkalineprotease (maxatalase, Biocon do Brasil industrial, RJ, Brasil)for 24 h at 60 #C; (d) inactivation of the protease (20 min at100 #C); and (e) removal of the supernatant, produced by cen-trifugation of the ketonic powder (tissue extract), for biochem-ical HA quantification.
2.2.4. HA quantificationThe biotin-conjugated probes were adsorbed into enzyme-
linked immunosorbent assay (ELISA) plates. In sequence,standard solution (0e500 mg/l) of HA of human umbilicalcord (Sigma Chemical Co., St. Louis, MO, USA) was addedto the wells, together with solutions of interpubic tissueextract, in triplicate. After incubation for 12 h at 4 #C andwashing three times with TriseHCl 0.05 M pH 8.0, 100 mlof HA-probe biotinylated (1 mg/ml) was added to each well.After a new wash, europium-labeled streptavidin (Wallac eTurku, Finland) was added to the plate for 30 min. Finally, af-ter washing, 200 ml of enhancement solution (PerkineElmerLife Sciences, Wallac Oy, Turku, Finland) was added to re-lease the europium bound to the streptavidin (which has an af-finity for the biotinylated HA-probe). The solution was thenshaken for 10 min and the released fluorescence was measuredin a time-resolved fluorometer. The data (counts/s) were pro-cessed automatically in the MultiCalc software programfrom PerkineElmer Life Sciences, Wallac Oy.
2.3. Statistical analysis
The optical density measurements from five mice in eachgroup were expressed as a mean" standard deviation. Statis-tical analyses were performed using the ManneWhitney testfor standard assumption of normality and constant variance.Differences between the groups were further assessed by theNewmaneKeuls test. P values <0.05 were considered statisti-cally significant.
3. Results
3.1. HA-histochemical analysis (HA-probe)
The presence of brownish HA staining was observed in theinterpubic tissue of virgin animals at D12, D15, D17 and D18of pregnancy; on D19 (the day of delivery); and in the 3dppand 5dpp animals. The HA staining presented a gradual in-crease from mid-pregnancy up to the last day preceding partu-rition (D18). Beginning at parturition (D19), a decline overa few days was observed.
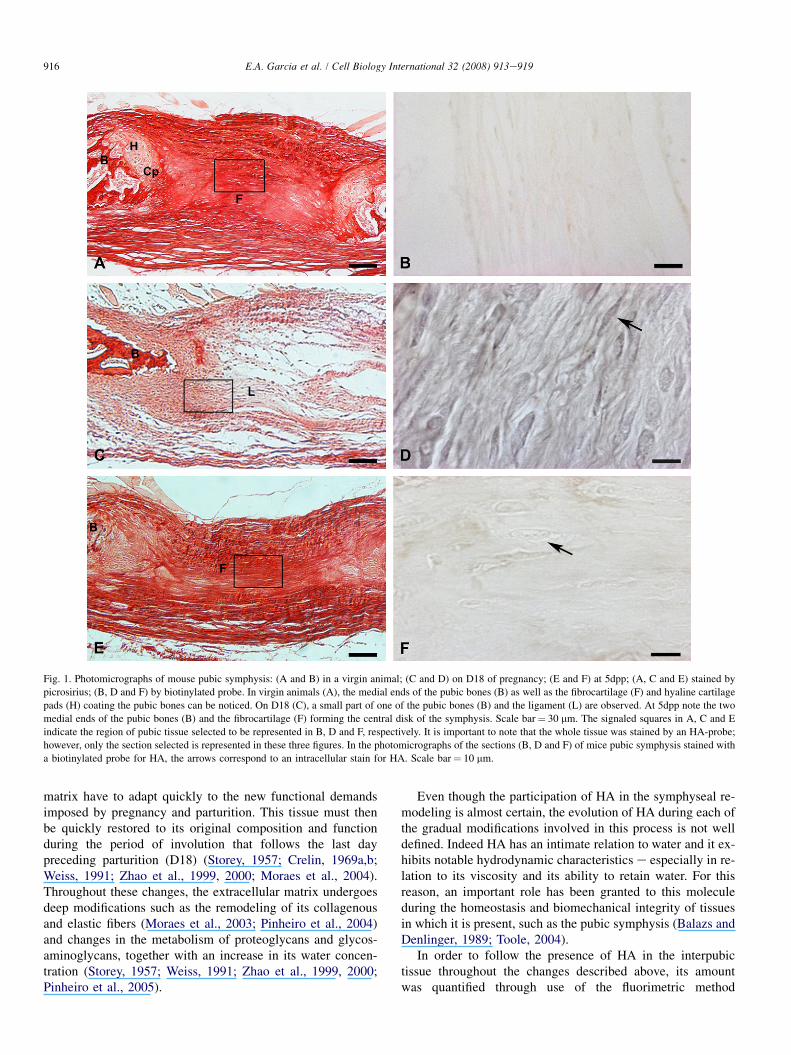
In the interpubic tissue of the virgin animals (Fig. 1A), itwas possible to observe only a slight brownish extracellularmatrix staining (Fig. 1B).
On the 17th and 18th days of pregnancy, hyaluronan stain-ing was found both in the extracellular matrix and within thetissue cells. Meanwhile, the greatest extracellular matrix
staining was observed on D18, preceding parturition.Fig. 1C and D shows the morphology and HA staining onD18, respectively. On post-partum (3dpp and 5dpp) samples,the extracellular matrix stains became less notable. However,even with the slighter staining found during the post-partum period, there is still evidence of intracellular HA(Fig. 1F).
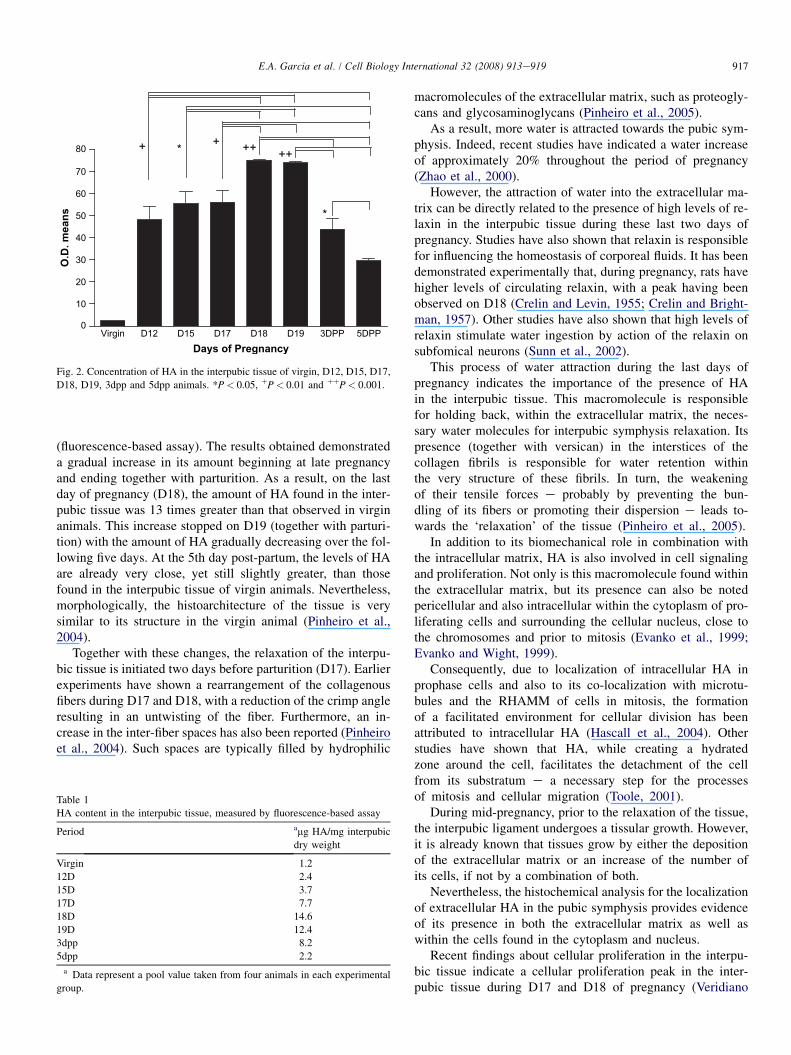
The optical density values obtained from virgin sampleswhen compared with samples from days D12, D15, D17,D18, D19 and post-partum days 3 and 5 were statistically sig-nificant (P< 0.001). The samples which presented the leastdifference when compared to virgin animals were five daysfollowing parturition (5dpp). Comparisons of the valuesfrom D12 samples with the other groups e D15, D17, D18and D19 and 3dpp and 5dpp e showed a statistically signifi-cant difference only for samples from D18, D19, and 5dpp in-terpubic tissue sections (P< 0.01). Fig. 2 demonstrates HAconcentration and standard deviation.
3.2. Quantification of HA by fluorescence-based assay
The quantitative results demonstrated the presence of1.2 mg/mg of dry weight of HA in the interpubic tissue of thevirgin samples. This amount increases gradually to 2.4 mg/mgdry weight of HA on D12, reaching 3.7 and 7.7 mg/mg dryweight on D15 and D17, respectively. The greatest amount ofHA, however, can be found on D18: 14.6 mg/mg dry weight.
Following parturition (D19), a small reduction in HA con-centration can be seen. The D19 samples, for example, present12.4 mg/mg dry weight of HA. The 3dpp samples also presentreduced amounts (8.2 mg/mg dry weight of HA). Even thoughthe 5dpp samples present an HA concentration 60% greaterthan that found in virgin animals, both these periods presentconcentrations that are relatively smaller when compared tothe 14.6 mg/mg dry weight peak on D18. The virgin samplescontain only 8% of the HA found on D18, while the 5dpp sam-ples present 15% of this peak concentration. Overall, there wasa 13-fold increase in HA amounts in D18 of pregnancy whencompared with virgin animals. Finally, the pubic symphysealHA amounts in 5dpp were similar to those of D12 animals.Table 1 presents these results.
4. Discussion
It is well-known that the pubic symphysis plays an impor-tant role in the transformation of the sexual organs duringpregnancy. In some species, a successful parturition greatlydepends on these hormonally regulated adaptations, whichare determined mainly by the size of the embryo and themother’s pelvis (Steinetz et al., 1957, 1965; Sherwood,1994). For this reason, the pubic symphysis is today a classicmodel of study of the hormonally induced transformationtaking part in the connective tissue in species such as miceand guinea pigs (Steinetz et al., 1957, 1965; Sherwood,1994).
Similar to the behavior observed in the uterine cervix, theinterpubic tissue cells and the components of its extracellular
915E.A. Garcia et al. / Cell Biology International 32 (2008) 913e919
matrix have to adapt quickly to the new functional demandsimposed by pregnancy and parturition. This tissue must thenbe quickly restored to its original composition and functionduring the period of involution that follows the last daypreceding parturition (D18) (Storey, 1957; Crelin, 1969a,b;Weiss, 1991; Zhao et al., 1999, 2000; Moraes et al., 2004).Throughout these changes, the extracellular matrix undergoesdeep modifications such as the remodeling of its collagenousand elastic fibers (Moraes et al., 2003; Pinheiro et al., 2004)and changes in the metabolism of proteoglycans and glycos-aminoglycans, together with an increase in its water concen-tration (Storey, 1957; Weiss, 1991; Zhao et al., 1999, 2000;Pinheiro et al., 2005).
Even though the participation of HA in the symphyseal re-modeling is almost certain, the evolution of HA during each ofthe gradual modifications involved in this process is not welldefined. Indeed HA has an intimate relation to water and it ex-hibits notable hydrodynamic characteristics e especially in re-lation to its viscosity and its ability to retain water. For thisreason, an important role has been granted to this moleculeduring the homeostasis and biomechanical integrity of tissuesin which it is present, such as the pubic symphysis (Balazs andDenlinger, 1989; Toole, 2004).
In order to follow the presence of HA in the interpubictissue throughout the changes described above, its amountwas quantified through use of the fluorimetric method
Fig. 1. Photomicrographs of mouse pubic symphysis: (A and B) in a virgin animal; (C and D) on D18 of pregnancy; (E and F) at 5dpp; (A, C and E) stained bypicrosirius; (B, D and F) by biotinylated probe. In virgin animals (A), the medial ends of the pubic bones (B) as well as the fibrocartilage (F) and hyaline cartilagepads (H) coating the pubic bones can be noticed. On D18 (C), a small part of one of the pubic bones (B) and the ligament (L) are observed. At 5dpp note the twomedial ends of the pubic bones (B) and the fibrocartilage (F) forming the central disk of the symphysis. Scale bar% 30 mm. The signaled squares in A, C and Eindicate the region of pubic tissue selected to be represented in B, D and F, respectively. It is important to note that the whole tissue was stained by an HA-probe;however, only the section selected is represented in these three figures. In the photomicrographs of the sections (B, D and F) of mice pubic symphysis stained witha biotinylated probe for HA, the arrows correspond to an intracellular stain for HA. Scale bar% 10 mm.
916 E.A. Garcia et al. / Cell Biology International 32 (2008) 913e919
(fluorescence-based assay). The results obtained demonstrateda gradual increase in its amount beginning at late pregnancyand ending together with parturition. As a result, on the lastday of pregnancy (D18), the amount of HA found in the inter-pubic tissue was 13 times greater than that observed in virginanimals. This increase stopped on D19 (together with parturi-tion) with the amount of HA gradually decreasing over the fol-lowing five days. At the 5th day post-partum, the levels of HAare already very close, yet still slightly greater, than thosefound in the interpubic tissue of virgin animals. Nevertheless,morphologically, the histoarchitecture of the tissue is verysimilar to its structure in the virgin animal (Pinheiro et al.,2004).
Together with these changes, the relaxation of the interpu-bic tissue is initiated two days before parturition (D17). Earlierexperiments have shown a rearrangement of the collagenousfibers during D17 and D18, with a reduction of the crimp angleresulting in an untwisting of the fiber. Furthermore, an in-crease in the inter-fiber spaces has also been reported (Pinheiroet al., 2004). Such spaces are typically filled by hydrophilic
macromolecules of the extracellular matrix, such as proteogly-cans and glycosaminoglycans (Pinheiro et al., 2005).
As a result, more water is attracted towards the pubic sym-physis. Indeed, recent studies have indicated a water increaseof approximately 20% throughout the period of pregnancy(Zhao et al., 2000).
However, the attraction of water into the extracellular ma-trix can be directly related to the presence of high levels of re-laxin in the interpubic tissue during these last two days ofpregnancy. Studies have also shown that relaxin is responsiblefor influencing the homeostasis of corporeal fluids. It has beendemonstrated experimentally that, during pregnancy, rats havehigher levels of circulating relaxin, with a peak having beenobserved on D18 (Crelin and Levin, 1955; Crelin and Bright-man, 1957). Other studies have also shown that high levels ofrelaxin stimulate water ingestion by action of the relaxin onsubfomical neurons (Sunn et al., 2002).
This process of water attraction during the last days ofpregnancy indicates the importance of the presence of HAin the interpubic tissue. This macromolecule is responsiblefor holding back, within the extracellular matrix, the neces-sary water molecules for interpubic symphysis relaxation. Itspresence (together with versican) in the interstices of thecollagen fibrils is responsible for water retention withinthe very structure of these fibrils. In turn, the weakeningof their tensile forces e probably by preventing the bun-dling of its fibers or promoting their dispersion e leads to-wards the ‘relaxation’ of the tissue (Pinheiro et al., 2005).
In addition to its biomechanical role in combination withthe intracellular matrix, HA is also involved in cell signalingand proliferation. Not only is this macromolecule found withinthe extracellular matrix, but its presence can also be notedpericellular and also intracellular within the cytoplasm of pro-liferating cells and surrounding the cellular nucleus, close tothe chromosomes and prior to mitosis (Evanko et al., 1999;Evanko and Wight, 1999).
Consequently, due to localization of intracellular HA inprophase cells and also to its co-localization with microtu-bules and the RHAMM of cells in mitosis, the formationof a facilitated environment for cellular division has beenattributed to intracellular HA (Hascall et al., 2004). Otherstudies have shown that HA, while creating a hydratedzone around the cell, facilitates the detachment of the cellfrom its substratum e a necessary step for the processesof mitosis and cellular migration (Toole, 2001).
During mid-pregnancy, prior to the relaxation of the tissue,the interpubic ligament undergoes a tissular growth. However,it is already known that tissues grow by either the depositionof the extracellular matrix or an increase of the number ofits cells, if not by a combination of both.
Nevertheless, the histochemical analysis for the localizationof extracellular HA in the pubic symphysis provides evidenceof its presence in both the extracellular matrix as well aswithin the cells found in the cytoplasm and nucleus.
Recent findings about cellular proliferation in the interpu-bic tissue indicate a cellular proliferation peak in the inter-pubic tissue during D17 and D18 of pregnancy (Veridiano
Virgin D12 D15 D17 D18 D19 3DPP 5DPP0
10
20
30
40
50
60
70
80 + *
*
+ ++ ++
Days of Pregnancy
O.D
. m
ea
ns
Fig. 2. Concentration of HA in the interpubic tissue of virgin, D12, D15, D17,D18, D19, 3dpp and 5dpp animals. *P< 0.05, !P< 0.01 and !!P< 0.001.
Table 1HA content in the interpubic tissue, measured by fluorescence-based assay
Period amg HA/mg interpubicdry weight
Virgin 1.212D 2.415D 3.717D 7.718D 14.619D 12.43dpp 8.25dpp 2.2
a Data represent a pool value taken from four animals in each experimentalgroup.
917E.A. Garcia et al. / Cell Biology International 32 (2008) 913e919

et al., 2007), which in turn coincides with the intracellularpresence of HA found in the present study. Such resultssuggest that the intracellular HA found on these two daysparticipates in the process of cellular proliferation.
Nonetheless, from days D19 to 5dpp it is still possible tonotice an intracellular staining of HA through histochemicalanalysis. Furthermore, in the days following parturition thereis both a reduction in the length of the interpubic ligament(Pinheiro et al., 2004) together with low cellular proliferation(Veridiano et al., 2007).
However, the intracellular presence of HA during this pe-riod may be attributed to the endocytosis of extracellular HAfor its degradation within the cell. The degradation processof HA occurs through its interaction with the CD44 receptor,promoting its endocytosis and posterior degradation by hyal-uronidases (Underhill, 1992). As previously shown, beginningwith parturition (D19), there is a significant reduction in theamount of HA in the interpubic tissue. On the 3rd day post-partum its levels are close to those observed on D17, whileon the 5th day post-partum it is reduced to levels close to thoseof D12.
For these reasons, we believe that intracellular HA doesparticipate in cellular proliferation found in the interpubic lig-ament from D17 to D18, whereas its continued intracellularpresence from D19 to 5dpp is a consequence of its necessarydegradation from the extracellular matrix.
During the last week of pregnancy and in the post-partumdays, cells of the interpubic ligament e particularly fibro-blasts e acquire structural characteristics of myofibroblasts.The high presence of this component during late pregnancyand post-partum has been considered a necessary factor inclosing the late pregnancy and the post-partum canal afterbirth (Moraes et al., 2004).
In addition, there is evidence that shows, when the myofi-broblast is differentiated, it has a reduced capacity for degrad-ing HA, which in turn allows HA to accumulate within thecell. Consequently, the accumulated HA within these cellscontributes to the maintenance of their myofibroblastic pheno-types (Jenkins et al., 2004). For this reason, it is believed thatperhaps the delay found in the reduction of HA levels in theinterpubic ligament following birth (3dpp) helps maintainthe myofibroblastic phenotype of its cells. After all, these my-ofibroblasts must keep together the pelvis bones in the pubicjoint, which helps to close the birth canal after delivery.
Therefore, the present biochemical and morphological re-sults, together with results of other studies, suggest the in-volvement of HA in mice pubic symphysis during latepregnancy, partum and post-partum in three distinct processes:(1) in the formation of a flexible structure of rigid consistencythat allows the passage of the embryo through the birth canal;(2) in the process of cellular proliferation, through possible in-teractions with intracellular components of mitotic cells, andalso through the formation of a hydrated zone around the cells,facilitating its detachment from the substratum; and (3) in themaintenance of the myofibroblastic phenotype of these cells,aiding in the process of involution in the interpubic tissue fol-lowing parturition.
Acknowledgements
EA Garcia and AM Veridiano were supported by CAPES(Coordenac~ao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and Pinheiro MC by CNPq (Conselho Nacional de De-senvolvimento Cientıfico e Tecnologico).
References
Balazs EA, Denlinger JL. Clinical uses of hyaluronan. Ciba Found Symp1989;143:265e80.
Crelin ES. Interpubic ligament, elasticity in pregnant free-tailed bat. Science1969a;164:81e2.
Crelin ES. The development of the bony pelvis and its changes during preg-nancy and parturition. Trans NY Acad Sci 1969b;31:1049e59.
Crelin ES, Brightman MW. The pelvis of the rat in response to estrogen andrelaxin. Anat Rec 1957;128:467e83.
Crelin ES, Levin J. The prepuberal pubic symphysis and uterus in the mouse:their response to estrogen and relaxin. Endocrinology 1955;57:730e47.
Evanko SP, Wight TN. Intracellular localization of hyaluronan in proliferatingcells. J Histochem Cytochem 1999;47:1331e41.
Evanko SP, Angello JC, Wight TN. Formation of hyaluronan and versican richpericellular matrix is required for proliferation and migration of vascularsmooth muscle cells. Arterioscler Thromb Vasc Biol 1999;19:1004e13.
Hall K. The effects of pregnancy and relaxin on the histology of the pubicsymphysis in the mouse. J Endocrinol 1947;5:174e82.
Hascall VC, Majors AK, De La Motte CA, Evanko SP, Wang A, Drazba JA,et al. Intracellular hyaluronan: a new frontier for inflammation. BiochimBiophys Acta 2004;1673:3e12.
Horn EH. Interpubic ligament regression in relaxed-treated virgin and primip-arous, post-partum mice. Endocrinology 1960;67:668e73.
Jenkins RH, Thomas GJ, Williams JD, Steadman R. Myofibroblastic differen-tiation leads to hyaluronan accumulation through reduced hyaluronan turn-over. J Biol Chem 2004;279:41453e60.
Lebl MDA, Martins JRM, Nader HB, Sim~oes MJ, De Biase N. Laryngoscope2007;117:595e9.
Lee JY, Spicer AP. Hyaluronan: multifunctional, megaDalton, stealth mole-cule. Curr Opin Cell Biol 2000;12:581e6.
Lesley J, Hascall VC, Tammi M, Hyman R. Hyaluronan binding by cell sur-face CD44. J Biol Chem 2000;275:26967e75.
Lin W, Shuster S, Maibach HI, Stern R. Patterns of hyaluronan stainingare modified by fixation techniques. J Histochem Cytochem 1997;45:1157e63.
Martins JR, Passerotti CC, Maciel RM, Sampaio LO, Dietrich CP, Nader HB.Practical determination of hyaluronan by a new noncompetitive fluores-cence-based assay on serum of normal and cirrhotic patients. Anal Bio-chem 2003;319:65e72.
Montes GS. Structure biology of the fibers of the collagenous and elastic sys-tem. Cell Biol Int 1996;20:17e27.
Moraes GS, Pinheiro MC, Yamada AT, Toledo OMS, Joazeiro PP. Differentialdistribution of elastic system fibers in the pubic symphysis of mice duringpregnancy, partum and post-partum. Braz J Morphol Sci 2003;20:85e92.
Moraes SG, Pinheiro MC, Toledo OMS, Joazeiro PP. Phenotypic modulationof fibroblastic cells in mice pubic symphysis during pregnancy, partumand postpartum. Cell Tissue Res 2004;315:223e31.
Ohya T, Kaneko Y. Novel hyaluronidase from streptomuces. Biochim BiophysActa 1970;198:607e9.
Ortega HH, Joazeiro PP, Mu~noz-De-Toro MM, Luque Eh, Montes GS. Differ-ential distribution of the fibres of the collagenous and elastic systems andof glycosaminoglycans in the rat pubic joint. J Submicrosc Cytol Pathol2001;33:463e72.
Pinheiro MC, Moraes SG, Battlehner CN, Caldini EG, Toledo OMS,Joazeiro PP. Histochemical and ultrastructural study of collagen fibers inmouse pubic symphysis during late pregnancy. Micron 2004;35:685e93.
Pinheiro MC, Mora OA, Caldini EG, Battlhener CN, Joazeiro PP,Toledo OMS. Ultrastructural, immunohistochemical and biochemical
918 E.A. Garcia et al. / Cell Biology International 32 (2008) 913e919

analysis of glycosaminoglycans and proteoglycans in the mouse pubicsymphysis during pregnancy. Cell Biol Int 2005;29:458e71.
Samuel CS, Butkus A, Coghlan JP, Bateman JF. The effect of relaxin on col-lagen metabolism in the nonpregnant rat pubic symphysis: the influence ofestrogen and progesterone in regulating relaxin activity. Endocrinology1996;137:3884e90.
Sherwood OD. Relaxin. In: Knobil E, Neill JD, editors. The physiology of re-production, vol.2. New York: Raven Press; 1994. p. 861e1009.
Steinetz BG, Beach VL, Kroc RL. The influence of progesterone, relaxin andestrogen on some structural and functional changes in the pre-parturientmouse. Endocrinology 1957;61:271e80.
Steinetz BG, Manning JP, Butler MT, Beach V. Relationships of growth hor-mone, steroids and relaxin in the transformation of pubic joint cartilageto ligament in hypophysectomized mice. Endocrinology 1965;76:876e82.
Storey E. Relaxation in the pubic symphysis of the mouse during pregnancyand after relaxin administration, with special reference to the behaviorof collagen. J Pathol Bacteriol 1957;74:147e62.
Sunn N, Egli M, Burazin TCD, Burns P, Colvill L, Davern P, et al. Circulatingrelaxin acts on subfornical organ neurons to stimulate water drinking in therat. Proc Natl Acad Sci 2002;99:1701e6.
Tammi MI, Day AJ, Turley EA. Hyaluronan and homeostasis: a balancing act.J Biol Chem 2002;277:4581e4.
Toole BP. Hyaluronan in morphogenesis. Semin Cell Dev Biol 2001;12:79e87.Toole BP. Hyaluronan from extracellular glue to pericellular cue. Nature 2004;
4:528e39.Underhill C. CD44: the hyaluronan receptor. J Cell Sci 1992;103:293e8.Veridiano AM, Garcia EA, Pinheiro MC, Nishimori FY, Toledo OM, Joazeiro
PP. The mouse pubic symphysis as a remodeling system: morphometricalanalysis of proliferation and cell death during pregnancy, partum and post-partum. 2007; 161e7.
Viell B, Struck H. Effects of the hormone relaxin on the metabolism of glycos-aminoglycans in the mouse symphysis pubis. Horm Metabol Res 1987;19:415e8.
Weiss G. The physiology of human relaxin. Contrib Gynecol Obstet 1991;18:130e46.
Zhao L, Roche PJ, Gunnersen JM, Hammond VE, Tregear GW, Wintour EM,et al. Mice without a functional relaxin gene are unable to deliver milk totheir pups. Endocrinology 1999;140:445e53.
Zhao L, Samuel CS, Tregear GW, Beck F, Wintour EM. Collagen studies inlate pregnant relaxin null mice. Biol Reprod 2000;63:697e703.
919E.A. Garcia et al. / Cell Biology International 32 (2008) 913e919





























