Embed Size (px)
Citation preview
Editor-in-ChiefManel Esteller
Cancer Epigenetics and Biology ProgramBarcelona, Spain
Volume 6 • Issue 7 • July 2011 Special Issue on Environmental Epigenetics

Editor-in-ChiEf
ManEl EstEllEr
Ca nCEr EpigEnEtiCs a nd Biology progr a M
Ba rCElona, spain
VoluME 6 issuE 7 July 2011
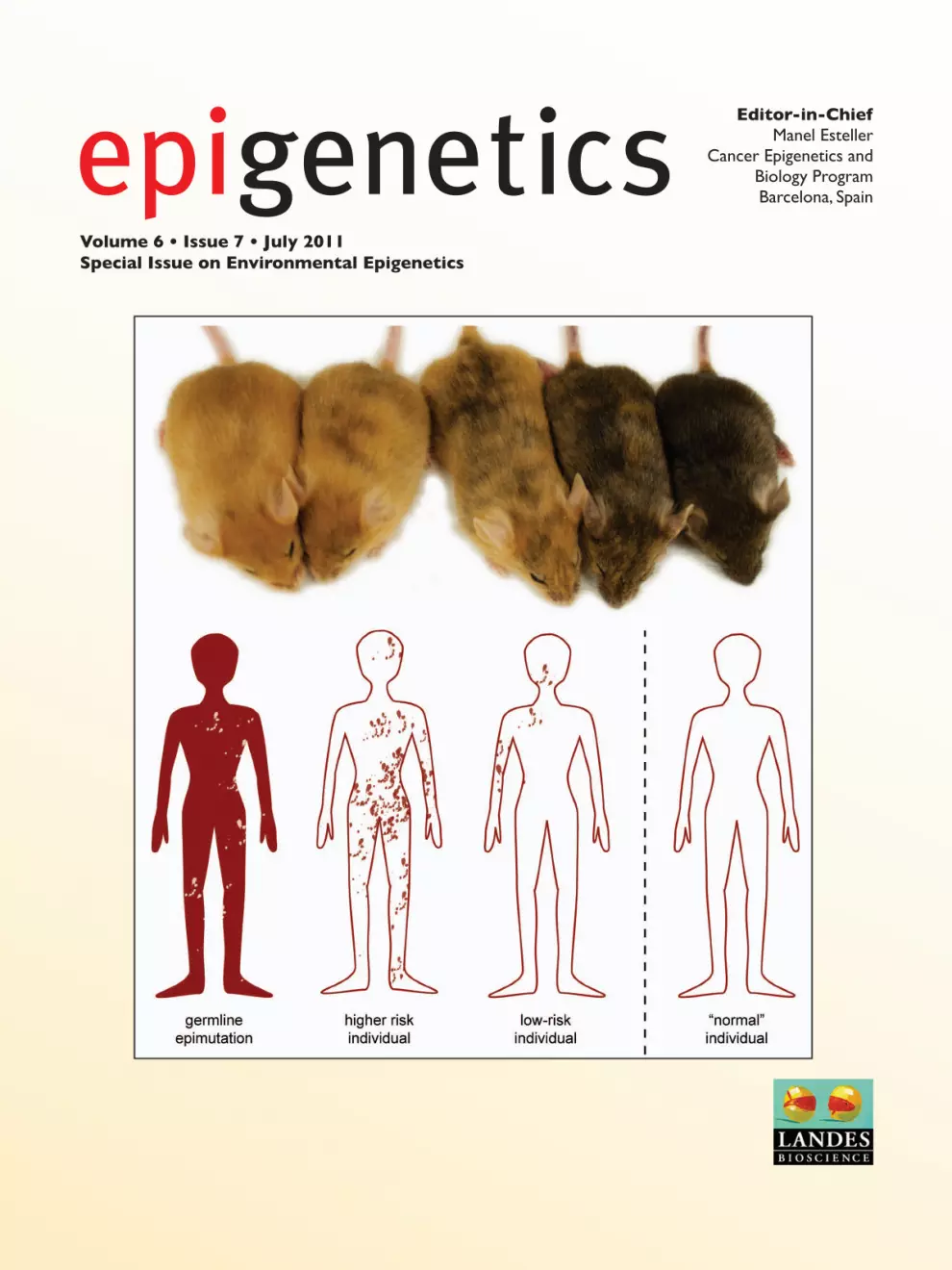
About the cover Epigenetic mosaicism may modulate
the risk and severity of epigenetic diseases. This is illustrated by the Avy
mouse (top). The range of coat colors of these isogenic littermates is caused
by varying epigenetic mosaicism of the Avy allele, which also leads to
variable expressivity of the associated obese diabetic phenotype. In humans
(bottom), the somatic burden of epi-mutation at disease-associated genes could impart a phenotype or disease risk relative to the proportion of cells
affected.For more information, see Martin et al., pp. 843–8.
EDITOR’S PREVIEW789 In this issue of Epigenetics: Special focus on environmental epigenetics Barbara P. Rattner
REVIEWS791 Timing is everything: The when and how of environmentally induced changes in the epigenome of animals Christopher Faulk and Dana C. Dolinoy
798 Biological memories of past environments: Epigenetic pathways to health disparities Zaneta M. Thayer and Christopher W. Kuzawa
804 Epigenetic mechanisms and cancer: An interface between the environment and the genome Zdenko Herceg and Thomas Vaissière
820 Environmental epigenetics in metal exposure Ricardo Martinez-Zamudio and Hyo Chol Ha
828 DNA methylation in white blood cells: Association with risk factors in epidemiologic studies Mary Beth Terry, Lissette Delgado-Cruzata, Neomi Vin-Raviv, Hui Chen Wu and Regina M. Santella
POINTS-OF-VIEW838 Environmental epigenetic transgenerational inheritance and somatic epigenetic mitotic stability Michael K. Skinner
843 Epigenetics in disease: Leader or follower? David I.K. Martin, Jennifer E. Cropley and Catherine M. Suter
849 Epigenetic modifications of metastasis suppressor genes in colon cancer metastasis Qian Li and Hong Chen
853 Hypothesis: Environmental regulation of 5-hydroxymethyl-cytosine by oxidative stress Nancy Chia, Luan Wang, Xiangyi Lu, Mary-Claude Senut, Carol Brenner and Douglas M. Ruden
857 Sex differences in epigenetic mechanisms may underlie risk and resilience for mental health disorders Heather M. Jessen and Anthony P. Auger
862 A genomic point-of-view on environmental factors influencing the human brain methylome Janine M. LaSalle
870 The effects of the dietary polyphenol resveratrol on human healthy aging and lifespan Agustín F. Fernández and Mario F. Fraga
875 The emerging role of epigenetic mechanisms in the etiology of neural tube defects Nicholas D.E. Greene, Philip Stanier and Gudrun E. Moore
884 Remodeling is at the heart of chromatin: The heartaches of chromatin Assam El-Osta
888 Genistein, an epigenome modifier during cancer prevention Yukun Zhang and Hong Chen
POINTS-OF-VIEW (CONTINUED)892 The role of holocarboxylase synthetase in genome stability is mediated partly by epigenomic synergies between methylation and biotinylation events Janos Zempleni, Yong Li, Jing Xue and Elizabeth L. Cordonier
BRIEF REPORT895 Heritability and role for the environment in DNA methylation in AXL receptor tyrosine kinase Carrie V. Breton, Muhammad T. Salam and Frank D. Gilliland
RESEARCH PAPERS899 Maternal dietary protein affects transcriptional regulation of myostatin gene distinctively at weaning and finishing stages in skeletal muscle of Meishan pigs Xiujuan Liu, Jinquan Wang, Runsheng Li, Xiaojing Yang, Qinwei Sun, Elke Albrecht and Ruqian Zhao
908 The influence of aging, environmental exposures and local sequence features on the variation of DNA methylation in blood Scott M. Langevin, E. Andres Houseman, Brock C. Christensen, John K. Wiencke, Heather H. Nelson, Margaret R. Karagas, Carmen J. Marsit and Karl T. Kelsey
920 Infant growth restriction is associated with distinct patterns of DNA methylation in human placentas Carolyn E. Banister, Devin C. Koestler, Matthew A. Maccani, James F. Padbury, E. Andres Houseman and Carmen J. Marsit
928 Methylation variation at IGF2 differentially methylated regions and maternal folic acid use before and during pregnancy Cathrine Hoyo, Amy P. Murtha, Joellen M. Schildkraut, Randy Jirtle, Wendy Demark-Wahnefried, Michele R. Forman, Edwin S. Iversen, Joanne Kurtzberg, Francine Overcash, Zhiqing Huang and Susan K. Murphy
937 Effects of endocrine disruptors on imprinted gene expression in the mouse embryo Eun-Rim Kang, Khursheed Iqbal, Diana A. Tran, Guillermo E. Rivas, Purnima Singh, Gerd P. Pfeifer and Piroska E. Szabó
ERRATA951 Erratum to: An S/MAR-based L1 retrotransposition cassette mediates sustained levels of insertional mutagenesis without suffering from epigenetic silencing of DNA methylation Danny Rangasamy
952 Erratum to: Ubiquitin-dependent distribution of the transcriptional coactivator p300 in cytoplasmic inclusion bodies Jihong Chen, Sabina Halappanavar, John P.H. Th’ ng, Qiao Li

EDITORIAL BOARDPierre-Olivier AngrandInterdisciplinary Research Institute (IRI)
Stephen BeckUCL Cancer Institute
Xiaodong ChengEmory University
David CrewsUniversity of Texas
Walter DoerflerUniversity of Cologne and Erlangen University
Melanie EhrlichTulane University
Assam El-OstaBaker Medical Research Institute
Nina FedoroffPennsylvania State University
Robert FeilInstitute of Molecular Genetics, CNRS
Patrizia FileticiUniversita La Sapienza
Francois FuksFree University of Brussels
Dennis R. GraysonPsychiatric Institute
Ueli GrossniklausUniversity of Zürich
Alessandro GuidottiUniversity of Illinois at Chicago
Lin HeShanghai Jiao Tong University
Randy JirtleDuke University
Victor LobanekovNIAID
Michael MeaneyMcGill University
Barbara R. MigeonJohns Hopkins University
Rolf OhlssonUppsala University
Alexander OlekEpigenomics AG
Arturas PetronisUniversity of Toronto
Aharon RazinHebrew University
Norbert ReichUniversity of California at Santa Barbara
Arthur D. RiggsBeckman Research Institute
Keith RobertsonUniversity of Florida
Carmen SapienzaTemple University
Paolo Sassone-CorsiUniversity of California, Irvine
Dirk SchübelerFriedrich-Miescher-Institute for Biomedical Research
Jonathan SecklUniversity of Edinburgh
Michael K. SkinnerWashington State University
Irina StanchevaUniversity of Edinburgh
Azim SuraniUniversity of Cambridge
Moshe SzyfMcGill University
Paul WadeNIEHS
Anton WutzResearch Institute of Molecular Pathology
Moshe YanivInstitut Pasteur
Dau-Xiu ZhouUniversity Paris XI
EXECUTIVE EDITORBarbara P. RattnerLandes Bioscience; Austin, TX USA
Abstracted/Indexed in Medline/PubMed and Index Medicus.
Print ISSN: 1559-2294; Online ISSN: 1559-2308
EDITORIAL POLICIES AND GUIDELINESOur Editorial Policies and guidelines for submitting manuscripts may be found here:http://www.landesbioscience.com/journals/epigenetics/guidelines
SUBMISSION OF MANUSCRIPTSManuscripts should be submitted online: http://epigenetics.msubmit.net
COPYRIGHT AND COPYRIGHT CLEARANCE CENTERThe Copyright Revision Act (PL 94-553), which became effective January 1, 1978, states that the copyright of a work is vested in the author from the moment of creation. Therefore, all authors who wish to publish in Epigenetics must grant an exclusive license to Landes Bioscience. It is understood that the authors grant Landes Bioscience an exclusive license to publish the work and also grant rights of reproduction, derivation, distribution, sale and display.
Authors who prepared their articles as part of their official duties as employees of the US Federal Government are not required to transfer copyright to Landes Bioscience, since these articles are considered to be in the public domain. However, it is necessary for these authors to sign the License to Publish form. In the case of articles supported by federal grants or contracts, a License to Publish is also required. The federal government may retain a nonexclusive license to publish or republish such material.
SUBSCRIPTION CLAIMSClaims for undelivered copies must be made no later than 6 months following month of publication. The publisher will supply missing copies when losses have been sustained in transit and when the reserve stock will permit.
Printed on acid-free paper effective with Volume 1, Issue 1, 2006.
EDITORIAL OFFICE1806 Rio Grande St.
Austin, Texas 78701 USA512.637.6050 phone
512.637.6079 fax
Senior Managing EditorBetsy Granger
Managing EditorNeil Kahn
Production AssistantKendell Richmond
Art Direction/Production DirectorKathryn Sauceda
Production and Print ManagerNicole Todd
Journal Publications DirectorKimberly Mitchell

www.landesbioscience.com Epigenetics 853
Epigenetics 6:7, 853-856; July 2011; © 2011 Landes Bioscience
POINT-OF-VIEW POINT-OF-VIEW
Key words: 5-methyl-cytosine, 5-hydroxymethyl-cytosine, metabolism, isocitrate dehydrogenase, TET
Abbreviations: 5-hmC, 5-hydroxymethyl-cytosine; 5-mC, 5-methyl-cytosine; IDH, isocitrate dehydrogenase; SDHD, succinate dehydrogenase; FH, fumarate hydratase; TET, ten-eleven-translocation; α-KG, α ketoglutarate; 2-HG, 2-hydroxyglutarate; HIF1α, hypoxia inducible factor 1alpha; DNMT, DNA nucleotide methyltransferase; ESC, embryonic stem cell; AML, acute myeloid leukemia; SAM, S-adenosylmethionine
Submitted: 05/10/11
Accepted: 05/15/11
DOI: 10.4161/epi.6.7.16461
*Correspondence to: Douglas M. Ruden; Email: [email protected]
Many environmental toxins, such as heavy metals, air particles
and ozone, induce oxidative stress and decrease the levels of NADH and NADPH, cofactors that drive anabolic biochemical reactions and provide reducing capacity to combat oxida-tive stress. Recently, it was found that the Ten-eleven translocation (TET) protein family members, which oxi-dize 5-methyl-cytosine (5-mC) to 5-hydroxymethyl-cytosine (5-hmC) in the DNA, were found to be activated under high oxygen conditions by alpha ketoglutarate (α-KG), a cofactor pro-duced by aerobic metabolism in the citric acid cycle. TET, Jumonji-family histone demethylases and prolylhydrox-ylase, a repressor of HIF1α under high oxygen conditions, all require α-KG as a cofactor for their activation. HIF1α and TET proteins, which appear to have opposing functions, impact several aspects of human life, including cell growth regulation, embryonic stem cell maintenance, cell differentiation and tumorigenesis. The role of metabolism on the regulation of global DNA meth-ylation and chromatin organization has recently gained greater attention from the biomedical research community. This article will discuss the possible role of TET activation and the regulation of 5-hmC and 5-mC levels in response to environmental stress. We will also dis-cuss how 5-hmC and 5-mC levels at the promoters of specific genes might be a useful biomarker for exposure to envi-ronmental toxins.
HypothesisEnvironmental regulation of 5-hydroxymethyl-cytosine by oxidative stress
Nancy Chia, Luan Wang, Xiangyi Lu, Mary-Claude Senut, Carol Brenner and Douglas M. Ruden*Department of Obstetrics and Gynecology; Institute of Environmental Health Sciences; C. S. Mott Center for Human Health and Development;
Wayne State University; Detroit, MI USA
Introduction
Environmental factors can alter the way our genes are expressed via epigenetic alter-ations. These epigenetic alterations dictate heritable patterns of gene expression that arise in the absence of DNA sequence modification and are largely regulated by changes in chromatin structure, mediated primarily by post-replicative methylation of DNA and post-translational modifica-tions of histones.1,2
The best-characterized epigenetic mark is a methyl group at the 5-position of cytosine bases known as 5-methyl-cytosine (5-mC) (Fig. 1). 5-mC has been studied extensively, and its role in gene regulation, X-chromosome inactivation, genomic imprinting, long-term silenc-ing of transposons and cancer develop-ment is well described.2 Cytosine exists as a free nucleotide that is incorporated into DNA during replication. The pat-tern of 5-mC in the genome is accurately preserved by mitotic inheritance through the action of DNA methyltransferases (DNMTs), specifically the maintenance DNA methyltransferase DNMT1, which catalyze the covalent addition of methyl groups to cytosine in newly synthesized DNA.3 5-mC is formed by post-replica-tive addition of a methyl group to cyto-sine through the action DNMTs, which use S-adenosylmethionine (SAM) as the methyl donor.3
5-hydroxymethyl-cytosine (5-hmC) was discovered in mammalian DNA in 1972 (Fig. 1),4 but it was thought to be an oxidative-damage product of DNA of

854 Epigenetics Volume 6 Issue 7
catalytic activity to produce 2-hydroxygu-latrate (2-HG) (Fig. 2).20,21
IDH1R132MUT is a gain of function mutation in isocitrate dehydrogenase that produces the “cancer metabolite” 2-HG, which inactivates TET1 (Fig. 2). 2-HG is a competitive inhibitor of multiple α-KG-dependent dioxygenases, including histone demethylases and the TET fam-ily of 5-mC hydroxylases. The NADP+-dependent isocitrate dehydrogenase genes IDH1 and IDH2 are mutated in >75% of low-grade gliomas and secondary glioblas-toma multiforme (GBM) and in ~20% of AML leukemia.20,21 When its normal catalytic activity is lost, mutant IDH1 and IDH2 also gained the function of cata-lyzing the reduction of α-KG to produce 2-HG, resulting in an accumulation of 2-HG in IDH1 or IDH2 mutated glio-mas and AML. In IDH1 mutated glioma, 2-HG accumulated to extraordinarily high levels of 5–35 mmol/g of GBM,25 which could be equivalent to 5–35 mM assuming the tissue density of 1 g/ml.
IDH1 and IDH2 mutations were subsequently observed in myeloid malig-nancies including de novo and second-ary AML (15%–30%) and pre-leukemic clonal malignancies including myelodys-plasia and myeloproliferative neoplasms (5% of chronic phase and 20% of trans-formed cases).26,27 The precise genetic con-text in which IDH1/2 mutations occur is not known, nor is the mechanism through which they contribute to the malignant phenotype. The most common IDH1/2 mutations in AML and brain tumors, affecting R132 of IDH1 or R140 and R172 of IDH2, have the common feature of acquiring a neomorphic enzymatic activ-ity catalyzing the NADPH-dependent reduction of αKG to R(-)-2HG.
and repressing (H3K27me3) chromatin marks.18 This suggests that 5-hmC is asso-ciated with poised chromatin in ESCs that become activated upon differentiation into specific developmental pathways.18
One of the most exciting recent find-ings in the cancer field is that mutations in metabolic genes are associated with many malignancies.19 Recent genomic sequencing efforts in acute myeloid leukemia (AML) and in other malignancies have identified new classes of oncogenic disease alleles. One recently identified class of genes mutated in cancer is that of genes coding for enzymes involved in citrate metabolism. The most prevalent of such mutations iden-tified to date affects the genes for cytosolic isocitrate dehydrogenase 1 (IDH1) and its mitochondrial homolog IDH2.20,21 Other metabolic mutants associated with cancer are succinate dehydrogenase (SDHD),23 and fumarate hydratase (FH).24 These findings have highlighted the metabolism, and drugs that affect metabolism, as one of the most important areas of cancer research and drug discovery.19
IDH1 and IDH2 are NADP+-dependent enzymes that normally cata-lyze the interconversion of isocitrate and alpha-ketoglutarate (α-KG; also known as 2-oxoglutarate). A dehydrogenase (also called DHO in the literature) is an enzyme that oxidizes a substrate by transferring one or more hydrides (H) to an acceptor. An oxygenase is any enzyme that oxidizes a substrate by transferring the oxygen from molecular oxygen O
2 (as in air) to it.
Dioxygenase is an oxygenase that transfers both oxygen atoms in O
2 to the substrate.
Two distinct alterations are caused by the tumor-derived mutations in IDH1 or IDH2: loss of its normal catalytic activity in the production of α-KG and gain of the
little importance and was therefore largely ignored. 5-hmC was re-discovered in 2009 by the laboratories of Nat Heintz5 at Rockefeller University and Anjana Rao at Harvard Medical School,6 and became of interest when it was shown that the TET family of Fe(II) and α-KG-dependent dioxygenases utilize molecular oxygen to transfer a hydroxyl group to 5-mC in the catalytic conversion of 5-mC to 5-hmC, and that one of these enzymes, TET2, is frequently mutated in myeloid neoplasms7,8 and both TET1 and TET2 are frequently mutated in glioma.9-14 The formation of 5-hmC can lead to demeth-ylation of DNA, which may contribute to the dynamics of DNA methylation.15 Recent evidence linking TET to aberrant DNA methylation has revealed that oxida-tion of 5-mC occurs in human cells and is catalyzed by TET proteins; the resulting 5-hmC may have opposite or alternative functions as 5-mC. It is possible that other yet undiscovered modifications of cytosine exist with yet-to-be-determined epigenetic effects during development (Fig. 1 and we refer to them as 5-xC or “excytosine”).
Dense methylation found around gene promoters in so-called CpG islands is asso-ciated with gene silencing; thus, the dis-tribution of methyl groups in the genome defines regions of varying transcriptional potential.16 The DNA methylation pat-tern during embryonic development is tissue specific, which may explain the connection to important aspects of cellu-lar differentiation.17 The role of 5-hmC in the promoters and gene bodies of genes is less well characterized, but a recent paper from Pastor and colleagues has shown that 5-hmC correlates with “bivalent” chromatin marks in embryonic stem cells (ESCs), with both activating (H3K4me3)
Figure 1. Cytosine base modifications. The “excytidine” modification refers to yet undiscovered modification(s) to cytosine with unknown functions.

www.landesbioscience.com Epigenetics 855
global chromatin and DNA methylation and hydroxymethylation patterns.
References1. Bird A. Perceptions of epigenetics. Nature 2007;
447:396-8.2. Portela A, Esteller M. Epigenetic modifications and
human disease. Nat Biotechnol 2010; 28:1057-68.3. Goll MG, Bestor TH. Eukaryotic cytosine methyl-
transferases. Annu Rev Biochem 2005; 74:481-514.4. Penn NW, Suwalski R, O’Riley C, Bojanowski K,
Yura R. The presence of 5-hydroxymethylcytosine in animal deoxyribonucleic acid. Biochem J 1972; 126:781-90.
be a key regulator of caloric restriction, which is thought to also increase NAD+ levels, because Sirt3 is required to pre-vent age-related hearing loss under caloric restriction.34
How might 5-hmC and 5-mC levels at the promoters of specific genes be a use-ful biomarker for exposure to early envi-ronmental toxins? We mentioned that developmental exposure to Pb alters DNA methylation patterns in human28 and in mammalian models.29-32 We speculate that this altered methylation pattern is mediated in part by oxidative stress-mediated altera-tions in metabolism, and thereby activated TET and other global chromatin modi-fying proteins (Fig. 3). We also speculate that each environmental toxin might affect metabolism and chromatin organization in a different manner and, consequently, each toxin has a unique “fingerprint” that reflects both the dose of the toxin and the type of environmental toxin.
If one can determine a genetic “finger-print” for the dose and type of prenatal toxin exposure, it might then be possible to perform whole genome DNA meth-ylation and hydroxymethylation assays on newborn dried blood spots to determine whether a newborn was exposed to an environmental toxin. These global DNA methylation and hydroxymethylation assays would potentially be much more sensitive than spectroscopic assays for environmental toxins, especially polyaro-matic hydrocarbons and other organic environmental toxins, which are difficult to detect. Consequently, global DNA methylation and hydroxymethylation assays are potentially extremely useful future tools for environmental toxicolo-gists to determine the long-term effects of early exposure to environmental toxins on human health.
In summary, we believe that we can combine an understanding of metabolism gleaned from cancer and caloric restric-tion studies to better understand how early exposures to oxidative-stress inducing envi-ronmental toxins might have lifelong effects on a person’s epigenome. We hope that some of the ideas in this review will stimu-late further studies on the metabolic effects of environmental toxins and thereby lead to a better understanding of their effects on
The genetic connection has given a major boost to the field of cancer metab-olism. Normal cells primarily gener-ate energy aerobically in mitochondria, whereas most tumor cells rely more heavily on glycolysis (the anaerobic con-version of glucose to lactate) to generate energy. Tumors also display other unique metabolic features. But the discovery of a mutated metabolic enzyme was strong evi-dence that metabolic abnormalities play an important role in oncogenesis (and it set off a frantic effort to understand what the mutation was doing in such common and lethal cancers).
We speculate, based on the insights gleamed from studying the effects of metabolic mutations on cancer epi-genetics and the seemingly unrelated field of caloric restriction and longevity, that environmental toxins might also affect global epigenetic patterns by interfering with metabolism (Fig. 3). Developmental exposure to the heavy metal lead (Pb), for instance, alters DNA methylation patterns in human28 and in mammalian models.29-32 Furthermore, the studies in the mammalian models have shown that these DNA methylation changes can last several years into the lifespan of the ani-mal (reviewed in ref. 32). The long-lasting effects of Pb on DNA methylation at spe-cific sites might explain the early origins of adult disease observations suggesting that exposure to Pb can have effects on human development decades later.33
In our model, oxidative stress decreases NADPH and NADH levels, which, in addition to glutathione (GSH), are cofac-tors that drive anabolic biochemical reac-tions and provide reducing capacity to combat oxidative stress. Consequently, NAD+ levels increase in the mitochon-dria under oxidative stress conditions, and this leads to the activation of the Sirtuin NAD+-dependent family of pro-tein deacetylases.34 Sirt3, a Sirtuin-family member in mammals, was recently found to deacetylase IDH2, the mitochon-drial isoform of isocitrate dehydroge-nase, and thereby activate it. Activated IDH2, in our model, can then produce increased amounts of α-KG and lead to changes in 5-hmC and 5-mC patterns in the genome (Fig. 3). Sirt3 was found to
Figure 2. The Yin-Yang of Isocitrate Dehy-drogenase. The wild-type IDH1 produces a-ketoglutarate, which is used by TET. The IDH1MUT produces 2-HG, which inhibits TET and thereby decreases global 5-hmC levels.
Figure 3. Regulation of TET by environmen-tal toxins. Environmental toxins produce oxidative stress, which leads to an increase in the ratio of NAD+ to NADH. Sirt3 is a NAD+ de-pendent protein deacetylase, which activates IDH2. IDH2 produces α-KG from isocitrate and α-KG activates TET, which leads to global DNA hydroxymethylation.

856 Epigenetics Volume 6 Issue 7
26. Marcucci G, Maharry K, Wu YZ, Radmacher MD, Mrozek K, Margeson D, et al. IDH1 and IDH2 gene mutations identify novel molecular subsets within de novo cytogenetically normal acute myeloid leukemia: a Cancer and Leukemia Group B study. J Clin Oncol 2010; 28:2348-55.
27. Ward PS, Patel J, Wise DR, Abdel-Wahab O, Bennett BD, Coller HA, et al. The common feature of leukemia-associated IDH1 and IDH2 mutations is a neomorphic enzyme activity converting alpha-ketoglutarate to 2-hydroxyglutarate. Cancer Cell 2010; 17:225-34.
28. Pilsner JR, Hu H, Ettinger A, Sanchez BN, Wright RO, Cantonwine D, et al. Influence of prenatal lead exposure on genomic methylation of cord blood DNA. Environ Health Persp 2009; 117:1466-71.
29. Zawia NH, Harry GJ. Correlations between devel-opmental ornithine decarboxylase gene expression and enzyme activity in the rat brain. Dev Brain Res 1993; 71:53-7.
30. Zawia NH, Harry GJ. Exposure to lead-acetate modulates the developmental expression of myelin genes in the rat frontal lobe. Int J Dev Neurosci 1995; 13:639-44.
31. Zawia NH, Harry GJ. Developmental exposure to lead interferes with glial and neuronal differential gene expression in the rat cerebellum. Toxicol Appl Pharm 1996; 138:43-7.
32. Zawia NH, Lahiri DK, Cardozo-Pelaez F. Epigenetics, oxidative stress and Alzheimer disease. Free Radical Bio Med 2009; 46:1241-9.
33. Brubaker CJ, Dietrich KN, Lanphear BP, Cecil KM. The influence of age of lead exposure on adult gray matter volume. Neurotoxicology 2010; 31:259-66.
34. Someya S, Yu W, Hallows WC, Xu J, Vann JM, Leeuwenburgh C, et al. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell 2010; 143:802-12.
16. Law JA, Jacobsen SE. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet 2010; 11:204-20.
17. Feng S, Jacobsen SE, Reik W. Epigenetic reprogram-ming in plant and animal development. Science 2010; 330:622-7.
18. Pastor WA, Pape UJ, Huang Y, Henderson HR, Lister R, Ko M, et al. Genome-wide mapping of 5-hydroxy-methylcytosine in embryonic stem cells. Nature 2011; 473:394-7.
19. Cairns RA, Harris IS, Mak TW. Regulation of cancer cell metabolism. Nat Rev Cancer 2011; 11:85-95.
20. Chowdhury R, Yeoh KK, Tian YM, Hillringhaus L, Bagg EA, Rose NR, et al. The oncometabolite 2-hydroxyglutarate inhibits histone lysine demethyl-ases. EMBO Reports 2011; 12:463-9.
21. Xu W, Yang H, Liu Y, Yang Y, Wang P, Kim SH, et al. Oncometabolite 2-hydroxyglutarate is a competitive inhibitor of alpha-ketoglutarate-dependent dioxygen-ases. Cancer Cell 2011; 19:17-30.
22. Baysal BE, Ferrell RE, Willett-Brozick JE, Lawrence EC, Myssiorek D, Bosch A, et al. Mutations in SDHD, a mitochondrial complex II gene, in heredi-tary paraganglioma. Science 2000; 287:848-51.
23. Ricketts CJ, Forman JR, Rattenberry E, Bradshaw N, Lalloo F, Izatt L, et al. Tumor risks and genotype-phenotype-proteotype analysis in 358 patients with germline mutations in SDHB and SDHD. Hum Mutat 2010; 31:41-51.
24. Tomlinson IP, Alam NA, Rowan AJ, Barclay E, Jaeger EE, Kelsell D, et al. Germline mutations in FH predispose to dominantly inherited uterine fibroids, skin leiomyomata and papillary renal cell cancer. Nat Genet 2002; 30:406-10.
25. Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM, et al. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2010; 465:966.
5. Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neu-rons and the brain. Science 2009; 324:929-30.
6. Tahiliani M, Koh KP, Shen Y, Pastor WA, Bandukwala H, Brudno Y, et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 2009; 324:930-5.
7. Dahl C, Gronbaek K, Guldberg P. Advances in DNA methylation: 5-hydroxymethylcytosine revisited. Clin Chim Acta 2011; 412:831-6.
8. Abdel-Wahab O. Genetics of the myeloproliferative neoplasms. Curr Opin Hematol 2011; 18:117-23.
9. Xu Y, Wu F, Tan L, Kong L, Xiong L, Deng J, et al. Genome-wide regulation of 5-hmC, 5-mC and gene expression by Tet1 hydroxylase in mouse embryonic stem cells. Mol Cell 2011; 42:451-64.
10. Guo JU, Su Y, Zhong C, Ming GL, Song H. Hydroxylation of 5-Methylcytosine by TET1 pro-motes active DNA demethylation in the adult brain. Cell 2011; 145:423-34.
11. Williams K, Christensen J, Pedersen MT, Johansen JV, Cloos PA, Rappsilber J, et al. TET1 and hydroxy-methylcytosine in transcription and DNA methyla-tion fidelity. Nature 2011; 473:343-8.
12. Wu H, D’Alessio AC, Ito S, Xia K, Wang Z, Cui K, et al. Dual functions of Tet1 in transcriptional regu-lation in mouse embryonic stem cells. Nature 2011 ; 473:389-93.
13. Frauer C, Rottach A, Meilinger D, Bultmann S, Fellinger K, Hasenoder S, et al. Different bind-ing properties and function of CXXC zinc finger domains in Dnmt1 and Tet1. PLoS One 2011; 6:16627.
14. Koh KP, Yabuuchi A, Rao S, Huang Y, Cunniff K, Nardone J, et al. Tet1 and Tet2 regulate 5-hydroxy-methylcytosine production and cell lineage specifica-tion in mouse embryonic stem cells. Cell Stem Cell 2011; 8:200-13.
15. Ndlovu MN, Denis H, Fuks F. Exposing the DNA methylome iceberg. Trends Biochem Sci 2011; In press.





























