Embed Size (px)
Citation preview

(~) INSTFFUT PASTEUR/ELsEVIER Paris 1996
Res. Microbiol. 1996, 147, 287-296
Identitication of a high-molecular-weight cadmium-binding protein in copper-resistant Bacillus acidocaldarius cells
C. Capasso, F. Nazzaro (*). F. Marulli, A. Capasso, E La Cara and E. Parisi (**)
The CN.R. Institute o f Protein Biochemistry and Enzymology, via Marconi 10, 80125 Napoli (Italy)
SUMMARY
Bacillus acidocaldarius grown in the presence of Cu +" was capable of accumulating the metal in the form of a protease-sensitive high molecular weight (HMW) moiety whose formation was inhibited by actinomycin D. OnFt cells pfeadapted in Cu ++ were able to grow in a Cd ++ -,'ontaining medium. A cell-free extract from c e d m i u m - ~ cells was fractioneted by gel.permeation chromatography. The major~/of cadmium was found associated with a HMW protein fraction which was further purified by anion exchange chromatography and high-performanca liquid chromatography, The molecu- lar weight of the purified protein was estimated to be ,'~t,000 by SDS-PAP:E. Am.;no acid analysis showed a low cysteine content and an abundance of aspmtate .~nd glutamate. It is likely "that the cadmium-binding ~o~ |n ;s an essential component of the mecha- nism mediating recovery from heavy metal toxicity.
Key-words: Copper, Cadmium; Metal-binding proteins, Bacillus acidocaldarius, Heaw metal resistance, Metal toxicity.
~TRODUCTION
Many metal ions are essential for the normal growth of organisms when p~sent at low concen- trations (Gadd, 1990). No biological funcdon is known for other types of metals such as lead, mercury and cadmium that, in contrast, may exert a toxic action towards microbial and other forms of life (Tynecka et al . , 1989; Tynecka and Szczes'niak, 1991).
Due to their long evolutionary history, micro- bial populations have developed various mecha- nisms of metal resistance (reviewed by Summers
and Silver, 1978; Gadd, 1992). These include (i) alteration of the membrane transport system responsible for metal accumulation (Laddaga et aL, 1985), (ii) extracellular precipitation of metal by a number of microbial products such as oxa- late, phosphate, stdphite and polymers (Aiking et al., 1982, 1984; Macaskie and Dean, 1984), (iii) intraceUular sequestration of metal by the cell wall (Mullen et aL, 1989; Andreoni et al., 1991), (iv) an energy-dependent efflux of metal ions (Kawai et al., 1990; Nies, 1992) and (v) enzy- matic reduction of ions to less toxic forms (Izaki et aL, 1974; Bopp and Ehrlich, 1988; Ishibashi
Submitted June 27. 1995, accepted November 7, 1995. (*) Present address: CNR Institute of Fl~)d Science and Technology, Avcllino, Italy. (**) Corresponding author.

288 C. CAPASSO ET AL.
et al., 1990). In a number of cases, metal detoxi- cation is achieved by complexation with metal- binding proteins similar to .mkaryotic metallothi- oneins (Olafson et al., 1979; Higham et al., 1984). Metallothioneins, initially isolated from equire renal cortex, are cadmium-, copper- and zinc-binding low molecular weight (LMW) pro- teins consisting of a single polypeptide chain of 60 amino acids, 25-30% of which are eysteines (Kiigi and Kojima, 1987).
Although metal concentration is generally low in the environment, in certain specific sites such as deep-sea vents, hot springs and volcanic areas, heavy metals are often present at high concentra- tions. Within these sites, microbial cells are natu- ral ly exposed to these compounds and may develop resistance mechanisms (Summers and Silver, 1978). These microorganisms represent - suitable material for the study of heavy metal resistance, both at a physiological and molecular level. In addition, they may also be employed for removal of toxic metals from aqueous wastes.
Bacillus acidocaldarius is an acidophilie ther- mophilic spore-forming eubacterium isolated f rom the the rmal ac id ic e n v i r o n m e n t near Agnano (Naples, Italy). The cells have a Gram- positive to Gram-variable morphology with a pronounced tendency to form filaments when grown in unaerated broth cultures (Darland and Brock, 1971). The purpose of the present study was to evaluate the tolerance of B. acidocaldarius to copper and cadmium exposure, and to study the resistance mechan i sm employed by this organism for metal detoxication.
M A T E R I A L S A N D M E T H O D S
C u l t u r e s
B. acidocaldarius w a s g r o w n at 6 0 ° C in a r a e r o - b ic l iqu id m e d i u m c o n t a i n i n g the f o l l o w i n ~ c o m p o -
nents per litre of distilled water : 3 g sucrose, 1 g yeast extract (Merk), 1 g casamino acids (Sigma), 1.3g (NH4)2SO 4, 0.3g KH2PO 4, 0.6g MgSO4.TH20, 0.1 g CaCI2-2H20. The pH was adjusted to 4.0 by addition of 0 IM H SO Large
" . " 2 . 4 " batch cultures were grown wah an aerauon flux of 200ml/min/1 in a 150-1 stainless steel prototype fer- menter (Bio-Industrie Mantovane s.r.l.). At the end of growth, the broth temperature was brought to 18°C and the cells were harvested by centrifugation at 2%000 rpm in a continuous flow centrifuge (Pad- berg model 7_,41). The cell paste was stored at -80°(: until use. For small scale cultures, cells were grown at 60°C in 500-ml flasks containing 100 ml of medium. Growth was monitored by measuring absorbance at 540 nm. When present, copper and cadmium salts were added at the indicated concen- trations.
For growth in the presence of cadmium, cells preadapted in 0.3 mM CuSO 4 were used as an inoc- ulum. Copper-adapted cells were washed three times with 0.1 M potassium phosphate buffer pH 7A and resuspended in fresh medium without copper. This suspension was used as a 1% inoculum for growth in cadmium-containing medium.
Cell extracts
Cells were grown to mid-log phase and harvested by centrifugation. Frozen cells were resuspended in 3 volumes of 50 mM Tris-HCl pH 8.6 containing 2 mM dithiothreitol (DTT), and disrupted by sonica- tion with a "Branson rood. J 17-A" sonicator using 5 pulses of 10 s each at 50 % power. The extract was centrifuged for 10 min at 17,000 g and the resulting supernatant centrifuged again for i h at 100,000 g.
Gel-filtration chromatography
The 100,000 g supematant, containing the indi- cated amounts of protein, was loaded on a Sephadex-G75 column (1.4 × 32 cm) previously equilibrated with 10 mM Tris-HCl pH 8.6 contain- ing 2 mM DTT. The column was eluted with the same buffer at a flow rate of I ml/min. When indi- cated, gel filtration was carried out on a "Cellufme GH-25" column (Amicon). The column (1.5 x32 cm) was equilibrated and eluted with 10mM Tris-HC1 pH 8.6 containing I mM DTT.
DTI" = dithiothreitol. HMW = high molecular weight. HPLC = high performance liquid chromatography. LMW = low molecular weight.
SDS- PAGE = sodium dodecyl sulphate/polyacrylamide gel ¢lcc-
trophoresis. TFA = triquoroacetic,

CADMIUM-BINDING PROTEIN IN COPPER-RESISTANT B. ACIDOCALDARIUS CELLS 289
Ardon-exchange chromatography
Pooled fractions from a gel filtration step were loaded on a I x l0 cm DEAE-cellulose column "DE- 5T" (Whatman) equilibrated with 10raM Tris-HCl pH 8.6, 2raM DTI'. The column was washed with three volumes of equilibration buffer and eluted with a 400- ml linear gradient from 0 to 1M NaCI in equilibration buffer at a flow rate of 120ml/h. The fractions of interest were pooled, dialysed extensively against 5 m_M Tris-HCl pH 8.6 ~:nd concentrated by polyethy- lene glycol (Fluka, MW 35,000) for further use.
was determined by graphite furnace without any pretreatment. Appropriate amounts of matrix modi- fier were added following the suggestion of the manufacturer.
SDS-PAGE
SDS-PAGE was performed on 12 % acrylamide gels following the method of Laemmli (1970) in the presence of 5 % mercaptoethanol. Gels were stained with silver staining (Otsuka et aL, 1988).
Reverse-phase HPLC
Fractions from the anion-exchange chromatogra- phy step were pooled and concentrated against poly- ethylene glycol. The sample was loaded on a high- performance liquid chromatography (HPLC) column (Pharmacia TSK ODS 1201" CIs, 0.8x30 cm). The column was eluted using as solvent A 0.1% tr;.fluo- roacefie acid (TFA) and as solvent B 90% acetoni- trile:water in 0.1% TFA in the following condi- tions: 0% B for 20 rain followed by a gradient from 1 to 90% of B in 45 rain. The absorbance was mon- itored at 280 nm.
Amino acid composition
Samples were dried under reduced pressure in a "Savant" centrifuge and hydrolysed in 6 N HC! con- taining 1% phenol and 2.5% mereaptoethanol after oxidat ion with performic acid (Behlen and Sehroeder, 1982).
N-terminal sequence
Automated Edman degradation was performed on a pulsed phase sequencer (Applied Biosystem rood. 120A). A sample containing 1 nmol of protein was loaded on a microsarnple cartridge.
Metal determination
Copper and cadmium were determined by atomic absorption spectrometry using a "Model 5100 Per- kin Elmer" apparatus equipped with a "Model HGA-600 Zeeman" graphite furnace. To measure copper accumulatio, in cells, samples were dry- washed at 140°C with 2 ml of 70% HNO 3 and 0.5 ml of 50% 1-120 2 added dropwise. The dry resi- due was dissolved in an appropriate volume of 0.2 % HNO. for flame analysis. For cadmium determina- tton, cells were heated for 5 nun at 70°C m 0.5 ml of 70% HlqO, without ashing. The samples were then diluted and" analysed by the graphite furnace tech- nique. Metal content in the chromatographic eluates
RESULTS
The tolerance of B. acidocaldarius to high concentrations of L-'u ++ was tested because of the high toxicity of this relatively available element. B. acidocaldarius cells showed a marked resis- t ance to Cu ++. No g r o w t h inh ib i t ion was observed from 0.1 to 1 mM CuSO4; a significant inhibition appeared at 2 mM CuSO 4. The amount of copper accumulated by the cells grown in 0. ! M CuSO 4 was estimated to be 0.4 mg per gram of cells.
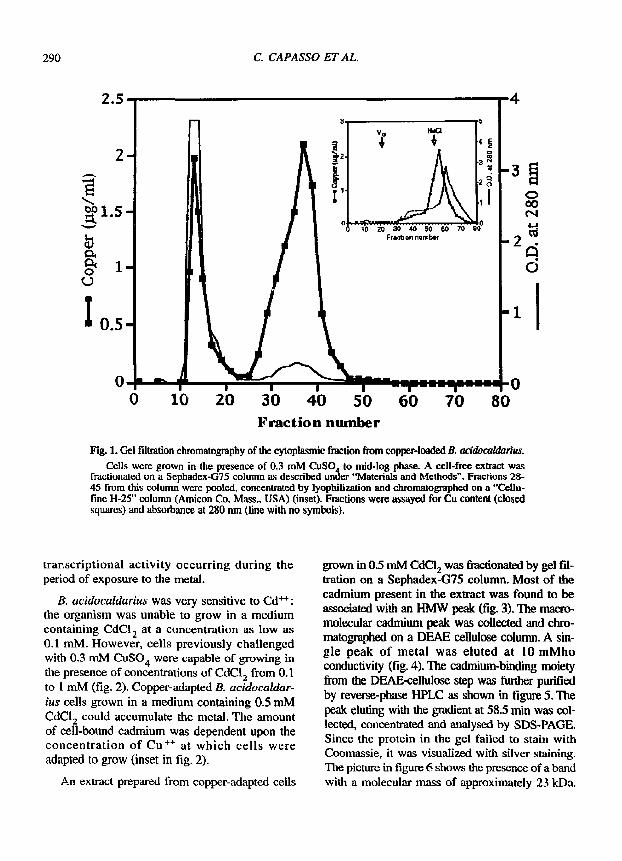
A supernatant extract prepared f rom cells grown in the presence of 0.3 mM CuSO 4 was fractionated by gel permeation chromatography on a Sephadex-G75 column, and the elution pro- file is reported in figure 1. While part of the metal was recovered in the HMW fraction, a significant portion was also found as a LMW peak. In order to better establish the size of the LMW material, the latter was rechromatographed on a "Cellufine GH-25" column with a fractionation range of 0,1- 3 kDa. The profde reported in the inset in figure 1 shows that copper eluted at a Ve/Vo close to that of sodium chloride. This suggests that the LMW peak of copper eluted from the Sephadex-G75 column may be free Cu ++, mainly generated by dissociation of the HMW complex. This, in fact, turned out to be unstable under dialysis, high ionic strength and sonication. The HMW copper- containing peak in figure 1 turned out to be sen- sitive to protease treatment. This indicated that the copper-binding moiety was a protein. In addi- tion, the amount of copper associated with the HMW fraction was markedly decreased in cells t rea ted with 0.05 txg/ml o f a c t i n o m y c i n D. Clearly, the peak of copper-protein is a result of a
290 C. CAPASSO ET AL.
2.5 ,, 4
2- II w ~, ~ ~ '
I I I I I Q6-to" z-'~'~o ~ 5o eo io ~o °
fl t l ,,.o,. . . . . . _ 2 '~
U
I 0 .5 1
0 0 0 1 0 2 0 3 0 4 0 5 0 60 7 0 8 0
F r a c t i o n n u m b e r
Fig. 1, Gel fdtration chromatography of the cytoplasmic fraction from copper-loaded B. acidocaldartus.
Cells were grown in the presence of 0.3 mlVl CuSO 4 to mid-log phase. A cell-free extract was fractionated on a Sephadex-G75 column as described under "Materials and Methods". Fractions 28- 45 from this column were pooled, concentrated by lyophilization and chromatographed on a "Cellu- f'me H-25" column (Amieon Co. Mass., USA) (inset). Fractions were assayed for Cu content (closed squares) and absorbanee at 280 nm (line with no symbols),
trar, scr ip t ional ac t iv i ty occur r ing dur ing the period of exposure to the metal.
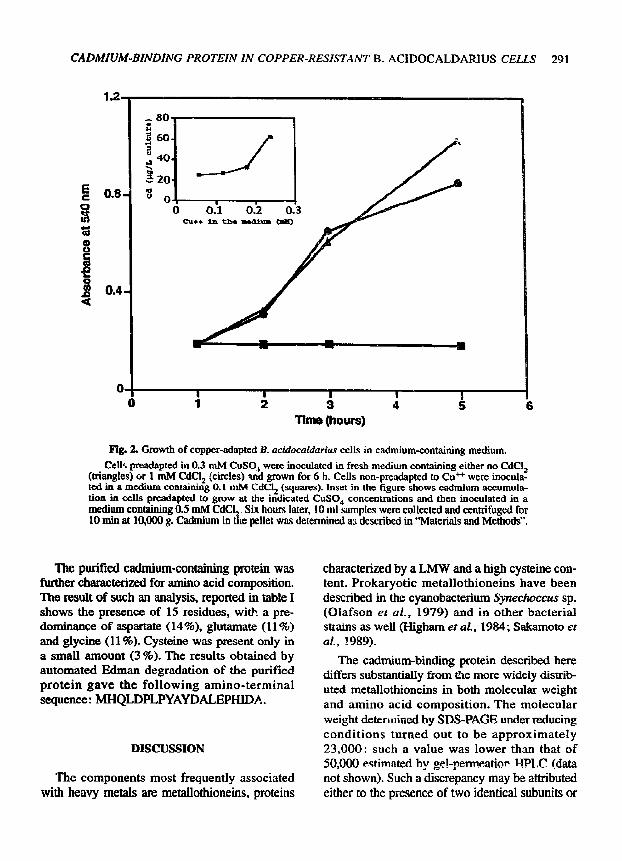
B. acidocaldarius was very sensitive to Cd++: the organism was unable to grow in a medium containing CdC! 2 at a concentration as low as 0.1 mM. However, cells previously chal lenged with 0.3 mM CuSO 4 were capable of growing in the presence of concentrations of CdCl 2 from 0.1 to 1 raM (fig. 2). Copper-adapted B. acidocaldar-
ius cells grown in a medium containing 0.5 mM CdCl 2 could accumulate the metal. The amount of cell-bound cadmium was dependent upon the c o n c e n t r a t i o n of Cu +÷ at w h i c h ce l l s were adapted to grow (inset in fig. 2).
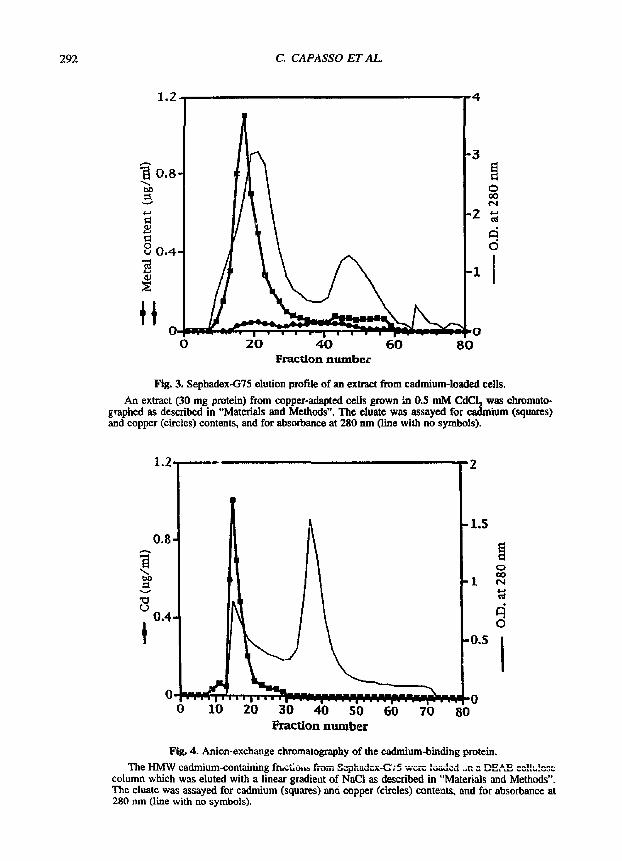
Am extract prepared from copper-adapted cells
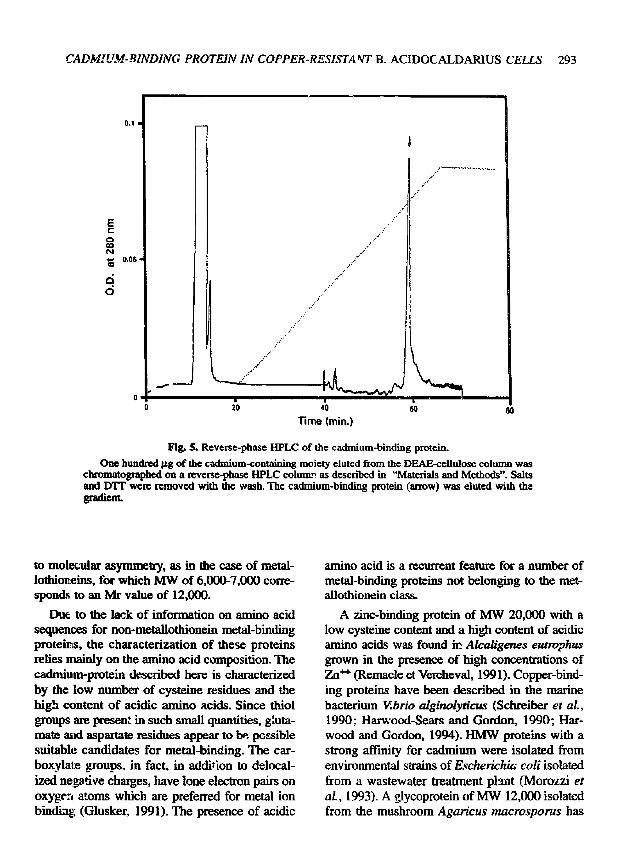
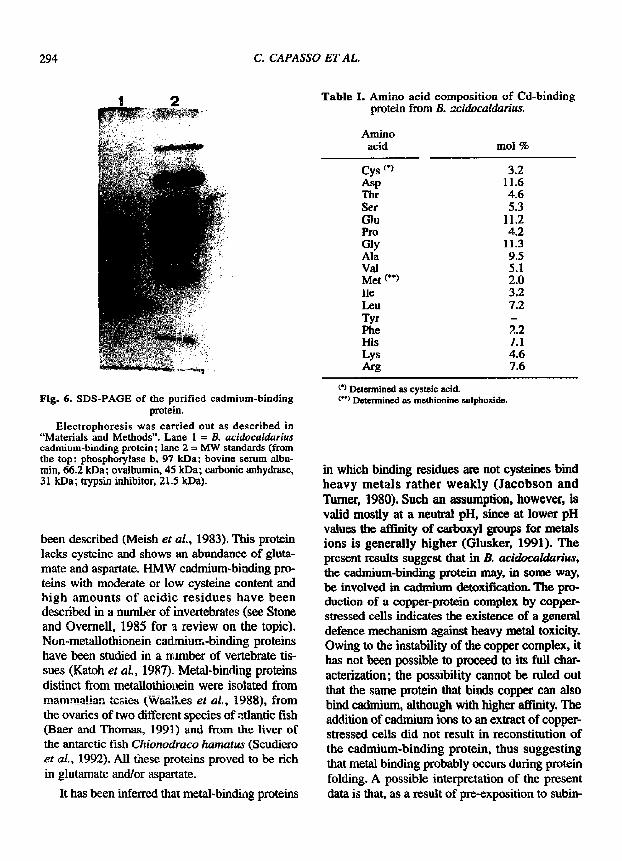
grown in 0.5 mM CdC! 2 was fractionated by gel ill- tration on a Sephadex-G75 column. Most of the cadmium present in the extract was found to be associated with an HMW peak (fig. 3). The macro- molecular cadmium peak was collected and chro- matographed on a DEAE cellulose column. A sin- gle peak of meta l was e luted at 10 m M h o conductivity (fig. 4). The cadmium-binding moiety from the DEAE-ceHulos¢ step was further purified by reverse-phase HPLC as shown in figure 5. The peak eluting with the gradient at 58,5 rain was col- lected, concentrated and analysed by SDS-PAGE. Since the protein in the gel failed to stain with Coomassie, it was visualized with silver staining. The picture in figure 6 shows the presence of a band with a molecular mass of approximately 23 kDa.
CADMIUM-BINDING PROTEIN IN COPPER-RESISTANT B. AC1DOCALDARIUS CELLS 291
1.2
E 0 .8
0 . 4 -
A 8 0 . ,,,
~'~, 60] / * ~,~
0 ,
" ~ ~ ' - ' " , - = . . . . . _
am i i i i l i " " ' - - i
i i i ! | 1 2 3 4 5 6
T i m e (hours )
Fig. 2, Growth of copper-adapted B. acidocaldarius cells in cadmium-containing medium.
Cells preadapted in 0.3 raM CuSO+ were inoculated in fresh medium containing either no CdC! 2 (triangles) or 1 mM CdC! 2 (circles) and grown for 6 h. Cells non-preadapted to Cu ++ were inocula- ted in a medium containin 8 0.1 ~ CdCI 2 (squares). Inset in the figure shows cadmium accumula- tion in cells preadapted to grow at the indicated CuSO 4 concentrations and then inoculated in a medium containing 0.5 mM CdCL~ Six hours later, 10 ml samples were collected and centrifuged for 10 min at 10,000 g. Cadmium in the pellet was determined a3 described in "Materials and Methods".
The purified cadmium-containing protein was further characterized for amino acid composition. The result of such an analysis, reported in table I shows the presence of 15 residues, wit/-, a pre- dominance of aspartate (14%), glutamate (11%) and glycine (11%). Cysteine was present only in a small amount (3 %). The results obtained by automated Edman degradat ion of the purif ied p r o t e i n g a v e the f o l l o w i n g a m i n o - t e r m i n a l sequence: MHQLDPLPYAYDALEPHIDA.
DISCUSSION
The components most frequently associated with heavy metals are metallothioneins, proteins
characterized by a L M W and a high cysteine con- tent. P rokaryo t i c me ta l lo th ione ins have been described in the cyanobacterium Synechoccus sp. (O ia f son et al. , 1979) and in o ther bac te r ia l sRains as well (Higham et aL, 1984; Sakamoto ¢t
aL, 1989).
The cadmium-binding protein described here differs substantially from t.he more widely distrib- uted metaUothioneins in both molecular weight and a m i n o acid compos i t i on . The mo lecu la r weight deten;fined by SDS-PAGE under reducing c o n d i t i o n s t u r n e d ou t to be a p p r o x i m a t e l y 23 ,000 : such a value was lower than that of 50,000 eqtimated hy ge!-permeafior T-!P(,C (data not shown). Such a discrepancy may be attributed either to the presence of two identical subunits or
292 C. CAPASSO ET A L
I l l l r l l l l W r
0 20 60
1.2
t ~
/_a
c~ o 0 . 4
tu
t+ 0
4 O Fraction n u m b e c
O 8O
"3
O eo
- 2
O
]
Fig. 3. Sephadex-G75 elufion profile of an extract from cadmium-lolled cells.
An extract (30 mg protein) from copper-adapted cells grown m 0.5 mM CdCL 2 was chromato- graphed as described in "Materials and Methods". The eluate was assayed for cadmium (squares) and copper (circles) contents, and for absorbance at 280 nm (line with no symbols). 1.2]
I 2
0
- 1 . 5
-0 .5 1
- 0 10 20 30 40 50 60 70 80
Fraction number
Fig. 4. Anion-exchange chromatography of the c~lmium-binding protein.
The HMW cadmium-containing frac~oti., from Sephadcx-C't5 wcrc ,~,,~,.~'-- ~ ~'~ on a DE,A~ ,.~,~..,,~,,~'" ' .... column which was eluted with a linear gradient of NaCI as described in "Materials and Methods". The eluate was assayed for cadmium (squares) ane copper (circles) contents, and for absorbance at 280 nm (line with no symbols).
CADM!UM-BINDING PROTEIN IN COPPER-RESISTA~rF B. ACIDOCALDARIUS CELLS 293
E c o
0.05 ,m ¢O
d 6
., .:;"
...,"
, j ."" ., .."
./" y'J
.,:
£.,"
,/.,:J"
0 • •
0 20 40
Time (rain.)
~?"
1 6O 80
Fig. 5. Reverse-phase HPLC of the cadmium-binding protein. Ono hundred pg of the cadmium-containing moiety elutcd from the DEAE-cellulose column was
chromatographed on a reverse-phase HPLC colum" as described in "Materials and Methods". Salts and DTF were removed with the wash. The cadmium-binding protein (arrow) was eluted with the, gradicnL
to molecular asymmetry, as in the case of metal- lothior~eins, for which M W of 6,000-7,000 corre- slxmds to an Mr value of 12,000.
Due to the lack of information on amino acid sequences for non-metallothionein metal-binding proteins, the characterization of these proteins relies mainly on the amino acid composition. The cadmium-protein described here is characterized by the low number of cysteine residues and the high content of acidic amino acids, Since thiol groups are presen: in such small quantities, gluta- mate and aspartate residues appear to be, po~ible suitable candidates for metal-binding. The car- boxylate groups, in fact, in addition to delocal- ized negative charges, have lone electron pairs on oxygen a:oms which are prefened for metal ion binding (Glusker, 1991). The presence of acidic
amino acid is a recurrent feature for a number of metal-binding proteins not belonging to the met- allothionein class.
A zinc-binding protein of MW 20,000 with a low cysteine content and a high content of acidic amino acids was found ~= Alcaligenes eutror~hus grown in the presence of high concentrations of Za~ ++ (Rcmaclc ct Vercheval, 1991). Copper-bind- ing proteins have been described in the marine bacterimn V, brio alginolyticus (Schreiber et al., 1990; Harwood-Sears and Gordon, 1990; Har- wood and Gordon, 1994). HMW proteins with a strong affinity for cadmium were isolated from environmental strains of Escherichia coli isolated from a wastewater treatment plant (Morozzi et al., 1993). A glycoprotein of MW 12,000 isolated from the mushroom Agaricus macrosporus has
294 C. CAPASSO E T AL.
! 2
Fig. 6. SDS-PAGE of the purified cadmium-binding protein.
Electrophores."s was carried out as described in "Materials and Methods". Lane 1 = B. acidocaldarius cadmium-binding protein; lane 2 = MW standards (from the top: phosphorylase b, 97 kDa; bovine serum albu- min, 66.2 kDa; ovalbumin, 45 kDa; carbonic anhydrase, 31 kDa; trypsin inhibitor, 21.5 kDa).
been described (Meish et al., 1983). This protein lacks cysteine and shows an abundance of gluta- mate and aspartate. HMW cadmium-binding pro- reins with moderate or low eysteine content and high amounts of acidic residues have been described in a number of invertebrates (see Stone and Ovemell, 1985 for a review on the topic). Non-metallothionein cadmium-binding proteins have been studied in a nmnber of vertebrate tis- sues (Katoh et al., 1987). Metal-binding proteins distinct from metaUothio~ein were isolated from mamma3ian testes (,Waalkes et aL, 1988), from the ovaries of two different species of ,',.tlantic fish (Baer and Thomas, 1991) and from the liver of the antarctic fish Chionodraco hamatus (Scudiero et aL, 1992). All these proteins proved to be rich in glutamate and/or aspartate.
It has been inferred that metal-binding proteins
Table I. Amino acid composition of Cd-binding protein from B, acidocaldarius.
Amino acid mol %
Cysr-) 3.2 Asp 11.6 Thr 4.6 Set 5.3 Glu 11.2 Pro 4.2 Gly 11.3 Ala 9.5 Val 5.1 Met (**) 2.0 lie 3.2 Leu 7.2 Tyr Phe 2.2 His 1.1 Lys 4.6 Arg 7.6
(') Determined as cystcic acid. ( " ) Determined as methionine sulphoxide.
in which binding residues are not cysteines bind heavy metals rather weakly (Jacobson and Turner, 1980). Such an assumption, however, is valid mostly at a neutral pH, since at lower pH values the affinity of carboxyl groups for metals ions is generally higher (Glusker, 1991). The present results suggest that in B. ac/doca/dar/us, the cadmium-binding protein may, in some way, be involved in cadmium detoxification. The pro- duction of a copper-protein complex by copper- stressed cells indicates the existence of a general defenee mechanism against heavy metal toxicity. Owing to the instability of the copper complex, it has not been possible to proceed to its full char- acterization; the possibility cannot be ruled out that the same protein that binds copper can also bind cadmium, although with higher affinity, The addition of cadmium ions to an extract of copper- stressed cells did not result in reconstitution of the cadmium-binding protein, thus suggesting that metal binding probably occurs during protein folding. A possible interpretation of the present data is that, as a result of pre-exposition to subin-

C A D M I U M - B I N D I N G P R O T E I N IN C O P P E R - R E S I S T A N T B. ACIDOCALDARIUS CELLS 295
hibitory levels o f C u ++, copper-resis tant variants we re selected. These variants cou ld be const i tu- t ive p roduce r s o f a m e t a l - b i n d i n g pro te in , thus
increasing resistance to c a d m i u m toxicity.
Acknowledgement s
This work was carried out in the framework of a exchange programme between the Italian CNR and the French CNRS.
We thank Dr. Christian Jeanthon for helpful discus- sion and Mr. Vitale Carrotore for sequence analysis.
Identification d'une prot~ine de haut poids mol~culaire f lxant le c a d m i u m darts les ceUules de Bacillus acidoealdarius r~ i s tantes au cuivre
Les cellules de Bacillus acidocaldarius cultiv~es en presence de Cu ++ sent capables d 'accumuler ce m6tal sous la forme d ' u n complexe qui se r~v~le sensible h l 'aet ion d ' une prot~ase et h l ' inhibit ion par rae t inomyein D. Scales les cellules adapt~es au cuivre ont montr6 la capacit6 de crot"tre en p~sence de cadmium. Un extrait de cellules de B. acidocal- t/at/us edtivg~,s en pr6senee de Cd ++ a 6t6 fracfionn~ par filtration sur gel. La plus forte part de cadmium se t r o u v e l i6e i~ u n e p r o t 6 i n e de h a u t po ids mol~culairc, qui a 6t~ purifi6e pax chromatographic anionic et HPLC. Le poids mol~eu la i re de cet te p ro tS ine , ~va lu6 p a r f i l t r a t i o n su r ge l , es t de 50.000 kDa, L ' ana ly se des acides amin6s entrant darts la compos i t ion de cet te prot6ine r6vi~le une teneur mod6t£'¢ en eyst~ine et des hautes teneurs en aeides aspartique et glutamique. On peut conelure clue eett¢ prot~ine est usa constituant important pour la d6fense centre la toxicit6 des m6taux lourds,
Mots-clds: Cuivre, Cadmium; M~talloprot6ines, Baci l lus ac idoca ldar ius , R6sis tance aux m6taux lourds, M6tallotoxicit6.
References
Aiking, H., Kok, K., Van Hecrikhuizen, H. & Van't Riet, J. (1982), Adaptation to eadmiuia by KlebsieUa aero- genes growing in cont inuous culture proceeds maintly via formation of cadmium sulfide. Appl. Environ. Microbiol., 44, 938-944.
Aiking, H., Stijnraan, A., Van Garderen, C., Van Heerik- huizen, H. & Van't Riet, J. (1984), Inorganic phos- phate accumulation and cadmium detoxification in
Nlebsieiia aerogenes NCTC418 growing in continu- ous culture. Appl. Environ. MicrobioL, 47, 374-377.
Andreoni, V., Finoli, C., Manfrin, P., Pelosi, M. & Vec- chio, A. (1991), Studies on the accumulation of cad- mium by a strain of Proteus mirabilis. FEMS Micro- biol. Ecol., 85, 183-192.
Baer, K.N. & Thomas, P. (1991), Isolation of novel metal- binding proteins distinct from metallothionein from spotted seatrout (Cynoscion nebulosus) and atlantic croaker (Micropogonia nndulatus) ovaires. Mar. BioL, 108, 31-37.
Bcblen, P. & Schroeder, R. (1982), High-sensitivity amino acid analysis: metodology for the determination of amino acid compositions with less than 100 pied- moles of pepddes. Anal. Biochem, 126, 144-152.
Bopp, L.H. & Ehrlich, H.L. (1988), Chromate resistance and reduction in Pseudomonas fluorescens strain LB300. ArclL MicrobioL, 150, 426-431.
Darland, G. & Brock, I".I). (1971), Bacillus acidocaldar- ius sp, nov., an aeidophilic thormophiUc spore-form- ing bacterium. & Gem MicrobioL, 67, %15.
Gadd, G.M. (1990). Metal tolerance, in: "Microbiology of extreme environments". (C., Edwards) (pp, 178-210). Open University Press, Cambridge.
Gadd, G.M. (1992), Molecular biology and bioteehnology of microbial interactions with organic and inorganic heavy met,'d compounds, in "'Molecular biology and biotechnology of extremophiles" (R.A. Herbert & R.L Sharp) (pp. 225-257). Blackie & Son Ltd, Glas- gow-London.
Glusker, J.P. (1991), Structural aspect of metal liganding to functional groups in proteins. Adv. Prot. Chenz, 42, 1-76.
Harwood, V. & Gordon, A.S. (1994), Regulation of extra- cellular copper-binding proteins and copper-sensitive mutants of Vibrio alginolyticus. Appl. Environ. Microbiol., 60, 1749-1753.
Harwood-Sears, V. & Gordon, A.S. (1990), Copper- induced production of copper-binding supernatant proteins by the marine bacterium Vibrio alginolyti- cus. AppL Environ. MicrobioL, 56, 1327-1332.
Higham, D.P., Sadler, P.J. & Scawen, M.D. (1984), Cad- injure-resistant Pseudomonas putida synthesizes novel cadmium protein. ~c~ence, 225, 1043-1046.
Ishibashi, Y., Cervantes, C. & Silver, S. (1990), Chro- mium reduction in Pseudomonas putida. Appl. Envi- ron. Microbiol., 56, 2268-2270.
Izaki, K., Tashiro, Y. & Funaba, T. (1974), Mechanism of mercuric chloride resistance in microorganisms. - - HI. Purification and properties of mercuric ion reducing enzyme from Escherichia coli bearing R factor. J. Biochem., 75, 591-599.
Jacobsen, K.B. & Turner, J.E. (1980), The interaction of cadmium and certain other metal ions with proteins and nucleic acids. Toxicol. (Amsterdam), 16, 1-38.
K~igi, J.H.R. & Kojima, Y. (1987), Reviews on metalloth- ionein, in "Metallothionein Ir" (J.H.R. Kiigi & Y. Kojima) (pp. 25-61). Birkh~iuser Verlage, Basel-Bos- ton.
Katoh, M., Aono, S., Keino, H. & Kashiwamata, S. (1987), High-molecular-weight cadmium-binding proteins in rat liver cytosol: isolation and partial charactedzat;on of an approximately 50 kDa protein. J. Biochem., 192, 1459-!468.
Kawai, K., Horitsu, H., Hamada, K. & Watanabe, M.

296 C. C A P A S S O E T AL.
(1990), Induction of cadmium resistance of Pseudo- monas putida GAM-1. Aerie. Biol. Chem., 54, 1553- 1555.
Laddaga, R.A., Bessen, R. & Silver, S. (1985), Cadmium- resistant mutant of Bacillus subtilis 168 with reduced cadmium transport. J. Bacteriol., 162, 1 I06-1110.
Laemmli, U.K. (t970), Cleavage of structural protein dur- ing the assembly of the head of bacteriophage T4. Nature (Lond.), 227, 680-687.
Macaskie, L.E. & Dean, A.C.R. (1984), Cadmium accu- mulation by a Citrobacter sp. J. Gen, Microbiol., 130, 53-62.
Meish, H.U.. Bechmann, I. & Smitt, J.A. (1983), A new cadmium-binding phosphoglycoprotein, cadmium- mycophosphatin, from the mushroom, Asaricus mac- rosporus. Biochym. Byophys. Acta, 745, 259-266.
Morozzi, G., Di Marco, L., Contend, S., Mangi',~ne, C. & Scardazza, E (1993), The presence of high-molec- ular weight proteins with a strong affinity for cad- mium in environmental Escherichia coli strains. Microbios, 75, 7-16.
Mullen, M.D., Wolf, D C., Ferris, F.G., Beveridge, TJ. , Fleming, C.A & Bailey, G.W. (1 °' )), Bacterial sorption of heavy metals. Appl. Environ. MicrobioL, 55, 3143-3149.
Nies, D.H. (1992), Resistance to cadmium, cobalt, zinc, and nickel in microbes. Piasmid. 27. 17-28.
Olafson, R.W., Abel, R.S. & Sire. R.S. (1979), Procaryotic mctallothionein. Preliminary characterization of a blue-green algae heavy-metal binding protein. Bio- chem. Biophys. Res. Commun., 89, 36-43.
Otsuka, F., Koizumi, S., Kimura, M,, & Oshawa, M, (1988), Silver staining for carboxymethylated metal- lothioneins in polyacrylamide gels. Analyt. Biochem., 168, 184-192.
Remacle, J. & Vercheval, C. (1991), A zinc-binding pro-
rein in a metal-resistant strain, Alcaligenes eutrophus CH34. Can. J. Microbiol., 37, 875-877.
Sakamoto, K., Yagasaki, M., Kirimura, K. & Usami, S. (1989), Resistance acquisition of ThiobaciUus thioox- idans upon cadmium and zinc ion addition and for- marion of cadmium ion-binding and zinc ion-binding proteins exhibiting metallothionein-like properties. J. Ferment. Bioeng., 67, 266-273.
Schreiber, D.R., Millero, FJ. & Gordon, A.S. (1990), Pro- duction of an extracellular copper-binding compound by the heterotrophic marine bacterium Vibrio algino- lyticus. Mar. Chem., 28, 275-284.
Scudiero, R., De Prisco, P., Camardella, L., D'Avino, R., di Prisco, G. & Parisi, E. (1992), Apparent deficiency of metallothionein in the liver of tim antarctic ic¢fish Chionodraco hamatus. Identification and isolation of a zinc-containing protein unlike metallothionein. Comp. Biochem. Physiot , 103B, 201-207.
Stone, H. & Ovem¢ll, $. (1985), Non-metallothionein cad- mium binding proteins. Comp. Biochem. Physiol., 8(82, 9-14.
Summers, A.O. & Silver, S. (1978), Microbial transforma- tion of metals. Ann. Rev. Microbiol., 32, 637-672.
Tynecka, Z., Maim, A. & Skwarek, T. (1989), Effect of Cd2 ÷ on growth of the cadmium-resistant and -sensi- tive Staphylococcus aureus. Acta Microbiol. Polon,, 38, 117-129.
Tynecka, Z. & Szczes'niak, Z. (1991), Efft~zt of Cd 2÷ on phosphate uptake by cadmium-resistant and cad- mium-sensitive Staphylococcus aureus. Microbios., 68, 53-63.
Waalkes, M.P., Perantoni, A. & Paln~r, A.E. (1988), Iso- lation and partial charact©rization of tim low-molecu- la," mass zinc/cadmium-binding protein from the testes of the patas monkey (Erythrocebus paras). Bio- chem. J., 256, 131-137.





























