Embed Size (px)
Citation preview

Journal of PathologyJ Pathol 2010; 220: 97–107Published online 4 August 2009 in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/path.2609
Original Paper
Identification of a novel 12p13.3 ampliconin nasopharyngeal carcinoma
Yvonne Y-Y Or,1# Grace T-Y Chung,1,2# Ka-Fai To,1,2 Chit Chow,1 Kwong-Wai Choy,3 Carol Y-K Tong,1
Alice W-C Leung,1 Angela B-Y Hui,4 Sai-Wah Tsao,5 Ho-Keung Ng,1,2 Timothy T-C Yip,6 Pierre Busson7
and Kwok-Wai Lo1,2*1Department of Anatomical and Cellular Pathology, State Key Laboratory in Oncology in South China, Prince of Wales Hospital, The ChineseUniversity of Hong Kong, Hong Kong SAR2Li Ka Shing Institute of Health Science, The Chinese University of Hong Kong, Hong Kong SAR3Department of Obstetrics and Gynaecology, Prince of Wales Hospital, The Chinese University of Hong Kong, Hong Kong SAR4Division of Applied Molecular Oncology, Ontario Cancer Institute, University of Toronto, Canada5Department of Anatomy, Li Ka Shing Faculty of Medicine, University of Hong Kong, Hong Kong SAR6Department of Clinical Oncology, Queen Elizabeth Hospital, Hong Kong SAR7Laboratoire de Biologie des Tumeurs Humaines, Institut Gustave Roussy, Villejuif, France
*Correspondence to:Kwok-Wai Lo, Department ofAnatomical and CellularPathology, Prince of WalesHospital, The Chinese Universityof Hong Kong, Shatin, N.T., HongKong SAR.E-mail: [email protected]
#These authors contributedequally to this study.
No conflicts of interest weredeclared.
Received: 21 April 2009Revised: 24 July 2009Accepted: 26 July 2009
AbstractNasopharyngeal carcinoma (NPC) is a distinct type of head and neck cancer commonlyoccurring in southern China. To decipher the molecular basis of this cancer, we performedhigh-resolution array CGH analysis on eight tumour lines and 10 primary tumours toidentify the genes involved in NPC tumorigenesis. In this study, multiple regions of gainwere consistently found at 1q21-q24, 7q11-12, 7q21-22., 11q13, 12p13, 12q13, 19p13 and19q13. Importantly, a 2.1 Mb region at 12p13.31 was highly amplified in a NPC xenograft,xeno-2117. By FISH mapping, we have further delineated the amplicon to a 1.24 regionflanked by RP11-319E16 and RP11-433J6. Copy number gains of this amplicon wereconfirmed in 21/41 (51%) primary tumours, while three cases (7.3%) showed high copynumber amplification. Among the 13 genes within this amplicon, three candidate genes,lymphotoxin beta receptor (LTβR), tumour necrosis factor receptor superfamily memeber1A (TNFRSF1R) and FLJ10665, were specifically over-expressed in the NPC xenograftwith 12p13.3 amplification. However, only LTβR was frequently over-expressed in primarytumours. LTβR is a member of the TNF family of receptors, which can modulate NF-κBsignalling pathways. Over-expression of LTβR in nasopharyngeal epithelial cells resulted inan increase of NF-κB activity and cell proliferation. In vivo study showed that suppressionof LTβR by siRNA led to growth inhibition in the NPC tumour with 12p13.3 amplification.These findings implied that LTβR is a potential NPC-associated oncogene within the 12p13.3amplicon and that its alteration is important in NPC tumorigenesis.Copyright 2009 Pathological Society of Great Britain and Ireland. Published by JohnWiley & Sons, Ltd.
Keywords: nasopharyngeal carcinoma; 12p13.3 amplicon; array CGH; oncogene; LTβR;Epstein–Barr virus
Introduction
Nasopharyngeal carcinoma (NPC) is a highly preva-lent malignant disease in Southern China and South-East Asia. Its incidence in southern Chinese is upto ∼25–30/100 000 persons/year, which is 100-foldhigher than that in Caucasians from Western coun-tries. As a distinct type of head and neck cancer,NPC is closely associated with EBV infection andcommonly appears as poorly differentiated or undif-ferentiated carcinomas [1]. The clonal EBV genomeand latent viral gene products are found in almost allprimary tumours [1,2]. Our previous studies showed
that NPC tumorigenesis is a multi-step process thatinvolved multiple genetic and epigenetic abnormal-ities, resulting in the disruption of various cellularmechanisms [1,3]. Radiation therapy is the mainstayof NPC treatment and the survival rate for patientswith early disease is > 70%. However, most patientsare diagnosed in the later stages and have a lowersurvival rate. Thus, identifying novel biomarkers andtherapeutic targets is important in the control of thisdeadly disease.
Our early molecular genetic studies demonstratedthe consistent genetic losses on chromosomes 3p,9, 11q, 13q, 14q and 16q and recurrent gains on
Copyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.www.pathsoc.org.uk

98 Y Y-Y Or et al
chromosome 12 in NPC [1,4,5]. In particular, theinactivation of tumour suppressors on 3p21.3 and 9p21was demonstrated to be an early event during thedevelopment of NPC [6,7]. Although abnormalitiesof multiple critical tumour suppressor genes (eg p16,RASSF1A, TSLC1 ) have been identified, informationabout oncogenes involved in transforming nasopharyn-geal epithelial cells is limited. Recently, using a CGHarray containing 1800 BAC clones, which span thehuman genome at about 1–3 Mb resolution, we haveinvestigated the genomic abnormalities in NPC andrevealed two novel amplicons at 11q13.1–13.3 and12p12.1–13.3. The study further showed that CCND1(cyclin D1 ) is the target oncogene in the 11q13 ampli-con and that its activation plays a role in NPC tumori-genesis [8]. Knockdown of CCND1 by siRNA in NPCcells led to significant inhibition of cell proliferation.However, this BAC array study was not able to fur-ther define the smaller amplicons consistently foundin NPC, due to its low resolution.
In the current study, we aimed to comprehensivelycharacterize the novel amplicon at 12p12.1–13.3 andto identify the candidate oncogenes in a panel ofEBV-positive NPC tumours by high-resolution arrayCGH analysis. A minimal region of gain mapped to12p13.31 was further delineated by FISH analysis. Weidentified the candidate oncogenes in this ampliconby comparing the copy number abnormality (CNA)and its expression in the tumour lines and primarytumours. The role of candidate oncogene in regulat-ing signalling pathways, cell proliferation and tumori-genicity was further demonstrated in nasopharyngealepithelial cells.
Materials and methods
Cell lines, xenografts and tumour specimens
Seven EBV-positive NPC xenografts (xeno-2117,xeno-1915, xeno-8, C15, C17, C18 and C19), a cellline (C666-1) and 10 microdissected primary tumoursfrom our previous BAC array CGH study were sub-jected to the current high-resolulation array CGHanalysis [8–12]. Two immortalized normal nasopha-ryngeal epithelial cell lines, NP69 and NP460, wereused as controls [13,14]. The cell lines were main-tained as described in our previous reports. Forimmunohistochemistry and FISH analysis, a total of 41archival formalin-fixed paraffin-embedded NPCs wererecruited from the Department of Anatomical and Cel-lular Pathology at the Prince of Wales Hospital, TheChinese University of Hong Kong. The study protocolwas approved by the Clinical Research Ethics Com-mittee of the Chinese University of Hong Kong. Allthe tumours were reviewed by two independent pathol-ogists as poorly differentiated or undifferentiated NPC,according to the WHO classification. The tumour sam-ples were also confirmed to be EBV-positive by EBER
Figure 1. Identification of 12p13.3 amplicon in NPC byhigh-resolution array CGH analysis. Copy number changes ofchromosome 12 were examined in 18 NPC tumours. Gainsare shown in red and losses in green. Red arrow indicates the12p13.3 amplicon
in situ hybridization. The clinical parameters are sum-marized in Table S1 (see Supporting information).
Array comparative genomic hybridization (CGH)analysis
The Agilent Human Genome CGH Microarray 44B(Agilent Technologies Inc., Palo Alto, CA, USA),which contained about 44 000 oligonucleotide probes,was used in this study. Array CGH analysis wasperformed according to the manufacturer’s protocol.In each array, 1 µg genomic male DNA was used
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

12p13.3 amplicon in NPC 99
Figure 2. Characterization of 12p13.3 amplicon in NPC. (a) The 12p13.3 locus amplified in NPCs. The regions of gains wereshown as grey bars. Black bars represent regions with high-level amplification. A 2.05 Mb minimal common region of gains wasdelineated. (b) Illustration of 12p13.3 amplicon in xeno-2117. The genomic location is shown on the x axis and the copy numberchange is shown on the y axis. The position and the copy number of the probes contain on the microarray are indicated byred (gains) or green (losses) dots. Two distinct regions of gains (regions 1 and 2) were found. The 2.1 Mb amplicon core withthreshold > 7.5 at region 1 was defined
as reference and labelled with Cy3–dUTP (Perkin-Elmer), while the target genomic DNA was labelledwith Cy5–dUTP. After scanning, data extraction andnormalization, the results were analysed using CGHAnalytics 3.3 software (Human Genome Build 17,Agilent Technologies). The log2 ratio measured for allprobes of the chromosome was smoothed by using2 Mb moving average. The Z-score algorithm wasapplied for calculating the chromosomal aberration,with a threshold set at 2 for defining regions withsignificant copy number gains and losses, while athreshold of 7.5 was used for amplification.
cDNA expression array
The Whole Human Genome Oligo Microarray, withover 41 000 probes (Agilent Technologies), was used
for expression analysis on NPC tumour lines (xeno-2117, C666-1 and C15), with NP69 as the reference.The expression analysis was carried out accordingto the manufacturer’s protocol. NPC sample cRNAwas labelled with 5-CTP (Cy5), while NP69 refer-ence cRNA was labelled with cyanine 3-CTP (Cy-3). Microarrays were scanned on Agilent’s dual-lasermicroarray scanner. The data were normalized andanalysed using GeneSpring GX version 9.0.5 software(Agilent Technologies).
Fluorescence in situ hybridization (FISH)
FISH analysis was performed to confirm the amplifica-tion of 12p13.31 in NPC tumour lines and 41 paraffin-embedded primary tumours as described [8]. FiveBAC clones located at 12p13.32–p13.31 were labelled
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

100 Y Y-Y Or et al
Figure 3. FISH analysis of 12p13.3 amplicon in NPC tumour lines. (a) Representative images from the FISH validation of the12p13.3 amplicon in xeno-2117. Clusters of RP11–451H11 signals (red) and two reference signals (green, chromosome 18centromeric probe) are shown in the xeno-2117 tumour cells. Only 24% of the tumour cells showed the cluster signals ofRP11–277E18 in region 2. (b) Illustration of FISH analysis of NPC tumour lines (xeno-2117, xeno-1915 and C666-1) with six BACclones on chromosome 12p13.3. The maxima located at RP11–451H11 are shown in xeno-2117
with Spectrum Orange–dUTP (Vysis, Downers Grove,IL, USA) by nick translation as the test probes.Centromere 18 labelled with Spectrum Green–dUTP(Vysis) was used as the reference probe. Chromosomallocations of these clones were validated in metaphasesof a normal individual. Amplifications and chromo-somal gains were scored when the ratios of test toreference probes were > 2 and 1.2, respectively.
Quantitative reverse transcription polymerasechain reaction (qRT–PCR)
Transcriptional expression of the candidate oncogenesat 12p13.3 in NPC samples and normal nasopharyn-geal epithelial cells were evaluated by quantitativeRT–PCR as described [8,12]. The expression levelsof β-actin of each sample were determined for nor-malization. The primers used are shown in Table S2(see Supporting information).
Western blotting and immunohistochemistryanalysis
The expression of TNFRSF1A and LTβR in NPCsamples was investigated by western blotting andimmunohistochemistry analysis. Total cell protein ofthe xenografts and cell lines was extracted and sub-jected to western blotting analysis as described [8].Primary antibodies used in this study were anti-LTβRgoat polyclonal antibody (N-15), anti-TNFRSF1Agoat polyclonal antibody (G-20) and anti-actin goatpolyclonal antibody (I-19; Santa Cruz). By immuno-histochemistry analysis, expression of LTβR andTNFRSF1A in the panel of 41 paraffin-embedded pri-mary tumours was also examined. Expression scoreswere defined with positive staining according to the
percentage of cells (<20%, score 1; 20–79%, score 2;80–100%, score 3). Scores 2 and 3 were assigned asover-expression.
NF-κB activity
The NF-κB activity of LTβR-transfected NP69 cellsand controls was determined by electrophoretic moli-bity shift assay (EMSA) and p50 ELISA assay. NP69cells were transfected with vector pReceiver–M03 orpReceiver–-LTβR (GeneCopoeia), using Fugene HD(Roche Diagnostics). After 24 h, the culture mediumwas aspirated and replaced with medium supplementedwith lymphotoxin α1β2 at concentrations of 0, 10, 50and 100 ng/ml, respectively. After 20 h, the cells wereharvested and nuclear protein was extracted. EMSAassay was performed as described previously [19]. p50binding activity was measured using a NF-κB p50ELISA Kit (Panomics).
Cell proliferation analysis
The cell proliferation of LTβR-transfected NP69 cellswas determined by WST-1 assay. In brief, NP69 cellswere seeded at 4 × 103 cells/well in a 96-well plateand transfected with pReceiver–M03 (vector) or pRe-ceiver–LTβR (GeneCopoeia). After 24 h, the culturemedium was replaced with medium supplemented withlymphotoxin α1β2 at concentrations of 0, 10, 50 and100 ng/ml, respectively. Six culture wells were setup for each treatment. WST-1 assay was performedaccording to the manufacturer’s protocol after 44 h.WST-1 (10 µl; Roche Diagnostics) and 100 µl mediumwas added to replace the old medium, incubated at37 ◦C for 30 min and UV absorbance was measuredat 450 and 690 nm.
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

12p13.3 amplicon in NPC 101
In vivo siRNA treatment
For in vivo siRNA treatment, xeno-2117 was inoc-ulated into athymic nude mice and allowed togrow to about 150 mm3. The treatment was doneusing Invivofectamine, according to the manufac-turer’s protocol (Invitrogen, Carlsbad, CA, USA).siRNA (10 mg) against LTβR (5′-GCACCUAUGUCUCAGCUAA) or control siRNA (Silencer NegativeControl No. 2) were mixed with 1 ml Invivofectamineand diluted in 15 ml 5% glucose solution. After con-centration using Amicon Ultra-15 Centrifugal Devicewith Ultracel-50 membrane, the concentrated complexwas collected and brought up to 1 ml with 5% glucosesolution. Each mouse was injected with 75 µl Invivo-fectamine complex intratumorally. The injection wasrepeated at day 4 after the first injection. The size ofthe tumour was measured every 2 days until day 8.The mice were sacrificed at day 8 and the xenograftswere collected for histological examination.
Results
Identification of 12p13.3 amplicon in NPC by arrayCGH
Using array CGH analysis, a high-resolution genomicprofile of each NPC sample was generated. Con-sistent with our previous studies, frequent chromo-some gains at 1q21-q24, 7q11-12, 7q21-22., 11q13,12p13, 12q13, 19p13 and 19q13, and chromosomelosses at 11q13, 11q23, 13q12-13, 14q21, 20p11 and22q13, were detected [4,8]. All these copy numberchanges were found in > 40% NPC samples (seeSupporting information, Figure S1). Interestingly, anovel region of gain or amplification on chromosome12p13 was identified in 50% NPC samples, includingfive xenografts and four primary tumours (Figure 1).A minimal region of gain mapped to 2.05 Mb at12p13.31 delineated by these tumours is shown inFigure 2a. Importantly, high copy number amplifica-tion of 12p13.31 was found in xeno-2117 and primarytumour T1 (Figure 2a). Figure 2b shows the xeno-2117 array CGH profile at 12p13.31. Two distinctamplification regions, regions 1 and 2, were noted.Region 1 spans a 2.6 Mb region flanked by the probesat the location of 4515530–7135185. Furthermore,an amplicon core was found within region 1 whenthe threshold was increased to 7.5. This maximumis about 2.1 Mb in size, located between 5025464and 7135185. Region 2 is about 0.4 Mb in size andis located at the centrometric part of 12p13.31. Theregion is flanked by the probes at 7708418–8129280.
Characterization of 12p13.3 amplicon by FISHanalysis
To verify the array CGH findings, we have furtherexamined the amplification of 12p13.31 in NPC sam-ples by FISH analysis, using multiple BAC clones
Figure 4. FISH analysis of 12p13.3 amplicon in primary NPCs.(a) Copy number aberrations of the five BAC clones are plottedfor 41 primary tumours. Red spots represented the cases withcopy number gain or amplification. RP11–451H11 shows thehighest frequency and level of gain and amplification in primarytumours. (b) Examples of 12p13.3 amplification in primarytumours. Multiple copies of RP11–451H11 were detected in thecases A10, A12 and A25. In case A10, no gains of RP11–319E6and RP11–433J6 were detected
(Figure 3a). Copy number gains and amplificationof the BAC clones within the two distinct ampli-cons were detected in xeno-1915 and xeno-2117(Figure 3a, b). In xeno-2117, high-level amplificationwas detected in RP11–451H11, which locates withinthe core region but not in the two flanking BACclones, RP11–319E16 and RP11–433J6. Figure 3ashows examples of FISH analysis on xeno-2117,showing clusters of RP11–451H11 signals in thetumour cells. This RP11–451H11 amplifcation wasdetected in almost all tumour cells of xeno-2117. Theresult further refined the amplicon core to a 1.24 Mbregion flanked by RP11–319E16 and RP11–433J6.However, only 24% xeno-2117 tumour cells showedcluster signals of RP11–277E18 in region 2. This find-ing suggested that the amplification of region 2 onlyoccurred in a subset of the tumour cells during cancerdevelopment.
Copy number gains of the 12p13.3 ampliconwere further examined in 41 undifferentiated primary
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

102 Y Y-Y Or et al
Figure 5. Expression of 12p13.3 candidate oncogenes in NPC. (a) cDNA expression arrray revealed the expression of 17 candidategenes on 12p13.3 amplicon in NPC tumour lines, xeno-2117, C666-1 and C15. NP69 was used as reference. (b) QuantitativeRT–PCR analysis of the transcription level of seven candidate genes in NPC tumour lines (xeno-2117, C15 and C666-1) andimmortalized nasopharyngeal epithelial cells (NP460 and NP69). β-Actin was used for normalization. Over-expression of FLJ10665,TNFRSF1A and LTβR were only detected in xeno-2117, but not in C666-1, C15 and NP460
Figure 6. Expression of LTβR in NPC tumour lines. Westernblot analysis of LTβR and TNFRSF1A in NPC tumour lines.Over-expression of LTβR and TNFRSF1A was found inxeno-2117
tumours to delineate the critical amplified region byFISH analysis. Figure 4a shows the copy numberchanges of five BAC clones in these tumours. Therewere 21 cases (51.2%) with copy number gains, andthree of them (7.3%) were high-level amplifications ofRP11–451H11 (Figure 4a, b). However, only 12.2%and 14.6% of the cases were detected with copy num-ber gains of RP11–433J6 and RP11–319E16, respec-tively. The results demonstrated that the 1.24 Mbregion flanked by RP11–319E16 and RP11–433J6was also commonly amplified in primary NPCs. Since
all 41 primary tumours are EBV-positive, no correla-tion between 12p13.3 amplifications and EBV infec-tion was identified.
Transcription of candidate oncogenes on 12p13.3amplicon
Although 12p13.3 amplification was reported in sev-eral human cancers including basal breast cancer,information about candidate oncogenes in this regionis rare. In this study, we focused on searchingfor candidate oncogene(s) in the region flankedby RP11–433J6 and RP11–319E16. This 1.24 Mbamplicon contains 17 known genes and transcripts.In order to identify the target oncogene(s), we inves-tigated the transcriptional level of these genes fromthe gene expression profiles of NPC tumour lines.The Whole Human Genome Oligo Microarray (Agi-lent Technologies) was used for gene expression pro-filing on xeno-2117, with the immortalized nasopha-ryngeal epithelial cell line, NP69, as reference. TheNPC tumour lines without 12p13.3 amplicon, C666-1 and C15, were also included as negative controls todefine the target(s). Among these 17 genes, only seven(NTF3, CD9, FLJ10665, TNFRSF1A, LTβR, TAPBPLand MRPL51 ) were significantly over-expressed inxeno-2117 (Figure 5a). More than two-fold increases
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

12p13.3 amplicon in NPC 103
Figure 7. Expression of LTβR in primary NPCs. (a) Detection of LTβR and TNFRSF1A expression in primary NPCs byimmunohistochemical staining. No or rare LTβR- or TNFRSF1A-expressing cells were found in the normal nasopharyngealepithelia. LTβR were highly expressed in the representative tumours (A10, A12 and A23). TNFRSF1A over-expression was onlydetected in a case (A9) without 12p13.3 amplification. (b) Results of LTβR copy number gains and expression in 41 primary NPCs.(c) A scattered plot shows the copy number changes of RP11–451H11 in the primary tumours with low, moderate and high LTβRexpression. Red spots represented the cases with copy number gain or amplification
in expression of these seven genes were found. Quan-titative RT–PCR analysis was performed to confirmthe expression array results in xeno-2117. Two immor-talized nasopharyngeal epithelial cell lines (NP69 andNP460) and samples without this amplicon, C666-1and C15 (negative control), were also included. Threecandidate genes, FLJ10665, TNFRSF1A and LTβR,were highly expressed in xeno-2117 but not in the neg-ative controls (C666-1 and C15) and normal nasopha-ryngeal epithelial cells (NP69 and NP460) (Figure 5b).
Over-expression of LTβR in NPCSince antibody for the protein encoded by FLJ10665 isnot available, we only examined the protein expressionof TNFRSF1A and LTβR in NPC tumours. Interest-ingly, TNFRSF1A and LTβR are critical componentsof the NF-κB signalling pathways which are consis-tently activated in NPC. By Western blotting, we con-firmed the over-expression of TNFRSF1A and LTβRin xeno-2117, but not in the cells without 12p13.3amplification (Figure 6). Expression of TNFRSF1Aand LTβR were also examined in the panel of 41 pri-mary tumours by immunohistochemical staining. Highfrequency (29/41, 70.7%) of LTβR over-expressionwas found in the primary tumours (Figure 7a-c). More-over, over-expression of LTβR was found in 16cases (76.2%) among those with 12p13 copy num-ber gain. High LTβR expression was detected in
all the three cases with high-level amplification of12p13.3 (Figure 7a, c). Surprisingly, TNFRSF1A wasonly expressed in 1/41 primary tumours (0.02%). Inthe cases with high-level amplification of 12p13.3, noTNFRSF1A over-expression was detected (Figure 7a).These findings strongly suggested that LTβR, but notTNFRSF1A, is one of the target oncogenes of 12p13.3amplicon in NPC.
Ectopic expression of LTβR-activated NF-κBsignals in nasopharyngeal epithelial cells
To confirm that up-regulation of LTβR induces NF-κBactivity in nasopharyngeal epithelia cells, we over-expressed LTβR in NP69 cells by transient transfec-tion of full-length LTβR cDNA (Figure 8a). By elec-trophoretic molibity shift (EMSA) assay, we foundthat NF-κB activity was increased in the LTβR-transfected NP69 cells. The NF-κB activity was highlyinduced in the LTβR-transfected NP69 cells treatedwith 10 or 50 ng/ml lymphotoxin α1β2 when com-pared with the controls (Figure 8b). A similar findingwas also detected by the ELISA assay for deter-mining p50-binding activity (Figure 8c). Significanthigher p50-binding activity was detected in the LTβR-transfected NP69 cells treated with various concen-tration of lymphotoxin α1β2. Furthermore, cell pro-liferation was also determined in LTβR-transfected
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

104 Y Y-Y Or et al
NP69 cells and controls with or without lympho-toxin α1β2 treatment. Figure 8d demonstrates thatthe growth rate showed 1.5–1.8-fold increase in theLTβR-transfected NP69 cells treated with 0–50 ng/mllymphotoxin α1β2. The findings suggested that LTβRover-expression contributes to aberrant activation ofNF-κB signals and cell proliferation in nasopharyngealepithelial cells.
Knockdown of LTβR inhibits in vivo tumourgrowth
The effect of LTβR on NPC tumour growth wasfurther investigated by siRNA-mediated suppression.Since xeno-2117, the tumour with LTBR amplification,is unable to grow in vitro, we performed in vivo LTβRsuppression by local administration of the siRNA tothe tumours established in athymic nude mice. Thespecific knockdown of LTβR by siRNA was firstconfirmed in HeLa cells (Figure 9). LTβR mRNAand protein expression were highly reduced in cellstreated with siRNA against LTβR. In vivo knockdownof LTβR was then carried out by intratumoral injectionof mixtures of Invivofectamine and siRNA againstLTβR into mice with xeno-2117. After 2 days, wedetected significant LTβR suppression in siRNA-treated xeno-2117 by immunohistochemistry. Mostof tumour cells in the xenografts showed absent orweak LTβR expression (Figure 9). For determiningthe effect of LTβR suppression on xeno-2117, weinjected LTβR siRNA into tumours at days 0 and 4and measured the tumour size for 8 days. As shownin Figure 10, significant inhibition of tumour growthwas observed in nude mice treated with LTβR siRNAwhen compared with the controls. Under microscopicexamination, massive cell death was observed in allfour LTβR siRNA-treated tumours, but not in tumourstreated with non-specific siRNA and reagent controls(Figure 10). The in vivo study provided evidence thatLTβR over-expression is critical for the growth ofNPC tumours with 12p13.3 amplification.
Discussion
Since recurrent gains of chromosome 12p were consis-tently found in NPC in multiple studies, activation ofoncogene(s) in this region is suspected to be a crit-ical event in NPC tumorigenesis [8,9,15,16]. Earlycytogenetic study of a NPC xenograft, xeno-2117, hasdemonstrated homogeneously staining regions (HSR)in a metacentric chromosome containing chromosome
Figure 8. Ectopic expression of LTβR activated NF-κBsignals in nasopharyngeal epithelial cells. (a) Detection of LTβRexpression in NP69 transient transfected with LTβR by Westernblotting. Arrow, endogenous LTβR; triangle, ectopic LTβR.(b) Increased NF-κB activity in NP69 cells transient transfectedwith LTβR. NP69 was transiently transfect with LTβR or vectorcontrol for 24 h and treated with various concentration of ligandlymphotoxin α1β2 for 20 h. Arrows indicate the increasedsignals for NF-κB in LTβR-transfected NP69 cells. (c) IncreasedNF-κB p50 binding activity in NP69 cells after transienttransfection of LTβR. NP69 was transfected with LTβR or vectoronly and treated with the range of LTα1β2 indicated. The p50binding activity was significantly enhanced in the lymphotoxinα1β2-treated LTβR-transfected NP69 cells. (d) Increased cellproliferation was found in the LTβR-transfected NP69 cells. Thecell proliferation of transfected and control cells was determinedby WST-1 assay. The growth rate of LTβR-transfected NP69cells treated with 0, 10 and 50 ng/ml lymphotoxin α1β2 wasapproximate 1.5-fold higher than that of controls
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
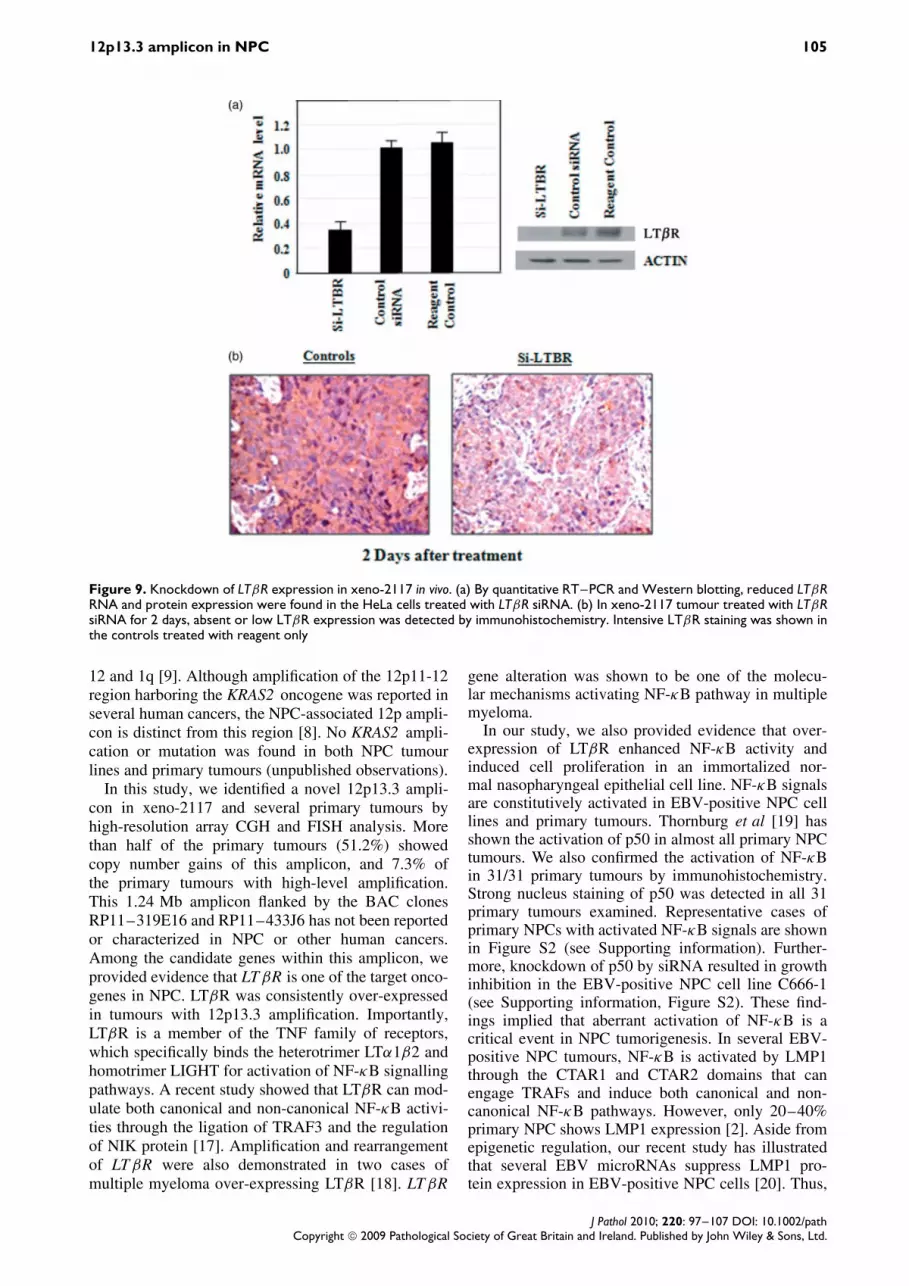
12p13.3 amplicon in NPC 105
Figure 9. Knockdown of LTβR expression in xeno-2117 in vivo. (a) By quantitative RT–PCR and Western blotting, reduced LTβRRNA and protein expression were found in the HeLa cells treated with LTβR siRNA. (b) In xeno-2117 tumour treated with LTβRsiRNA for 2 days, absent or low LTβR expression was detected by immunohistochemistry. Intensive LTβR staining was shown inthe controls treated with reagent only
12 and 1q [9]. Although amplification of the 12p11-12region harboring the KRAS2 oncogene was reported inseveral human cancers, the NPC-associated 12p ampli-con is distinct from this region [8]. No KRAS2 ampli-cation or mutation was found in both NPC tumourlines and primary tumours (unpublished observations).
In this study, we identified a novel 12p13.3 ampli-con in xeno-2117 and several primary tumours byhigh-resolution array CGH and FISH analysis. Morethan half of the primary tumours (51.2%) showedcopy number gains of this amplicon, and 7.3% ofthe primary tumours with high-level amplification.This 1.24 Mb amplicon flanked by the BAC clonesRP11–319E16 and RP11–433J6 has not been reportedor characterized in NPC or other human cancers.Among the candidate genes within this amplicon, weprovided evidence that LTβR is one of the target onco-genes in NPC. LTβR was consistently over-expressedin tumours with 12p13.3 amplification. Importantly,LTβR is a member of the TNF family of receptors,which specifically binds the heterotrimer LTα1β2 andhomotrimer LIGHT for activation of NF-κB signallingpathways. A recent study showed that LTβR can mod-ulate both canonical and non-canonical NF-κB activi-ties through the ligation of TRAF3 and the regulationof NIK protein [17]. Amplification and rearrangementof LTβR were also demonstrated in two cases ofmultiple myeloma over-expressing LTβR [18]. LTβR
gene alteration was shown to be one of the molecu-lar mechanisms activating NF-κB pathway in multiplemyeloma.
In our study, we also provided evidence that over-expression of LTβR enhanced NF-κB activity andinduced cell proliferation in an immortalized nor-mal nasopharyngeal epithelial cell line. NF-κB signalsare constitutively activated in EBV-positive NPC celllines and primary tumours. Thornburg et al [19] hasshown the activation of p50 in almost all primary NPCtumours. We also confirmed the activation of NF-κBin 31/31 primary tumours by immunohistochemistry.Strong nucleus staining of p50 was detected in all 31primary tumours examined. Representative cases ofprimary NPCs with activated NF-κB signals are shownin Figure S2 (see Supporting information). Further-more, knockdown of p50 by siRNA resulted in growthinhibition in the EBV-positive NPC cell line C666-1(see Supporting information, Figure S2). These find-ings implied that aberrant activation of NF-κB is acritical event in NPC tumorigenesis. In several EBV-positive NPC tumours, NF-κB is activated by LMP1through the CTAR1 and CTAR2 domains that canengage TRAFs and induce both canonical and non-canonical NF-κB pathways. However, only 20–40%primary NPC shows LMP1 expression [2]. Aside fromepigenetic regulation, our recent study has illustratedthat several EBV microRNAs suppress LMP1 pro-tein expression in EBV-positive NPC cells [20]. Thus,
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

106 Y Y-Y Or et al
Figure 10. Effect of LTβR suppression in xeno-2117. Significant growth inhibition was observed in xeno-2117 treated with LTβRsiRNA in vivo. The size of dissected LTβR siRNA-treated tumours at day 8 was significantly smaller than that of controls. Undermicroscropic examination, massive cell death (∗) was found in the LTβR siRNA-treated xeno-2117
LTβR amplification may contribute as one of themechanisms activating NF-κB signalling in NPC. Thepresent study is the first to demonstrate that constitu-tive NF-κB signalling in NPC is activated by geneticalterations. Screening for other genetic and epigeneticchanges contributing to the constitutive activation ofNF-κB pathways in NPC is ongoing.
The oncogenic properties of LTβR have also beenshown in several tumour models. LTβR was foundto promote cell growth and induce angiogenesis infibrosarcoma by enhancing the production of MIP-2 and angiogenic CXC chemokine [21]. Fujiwaraet al [22] have also demonstrated the transformingactivity of LTβR in 3T3 cells by both in vitro and
in vivo tumorigenicity assay. Furthermore, constitu-tive activation of NF-κB by LTβR was reported to beassociated with the autonomous growth of melanomacells. Knockdown of LTβR expression in melanomacells by shRNA resulted in decreased NF-κB activity,cell growth and invasiveness [23]. The critical roleof LTβR amplification and over-expression in NPCtumorigenicity was also demonstrated in our study.After local administration of LTβR siRNA, in vivogrowth inhibition was observed in the tumour withLTβR amplification. Knockdown of LTβR may sup-press the expression of a number of NF-κB targets(eg EGFR, bcl2 and cIAP-2) that are important inthe survival and proliferation of NPC tumour cells[19,24]. Taken together, these findings are evidence
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.

12p13.3 amplicon in NPC 107
that LTβR amplification may play an important rolein the tumorigenesis of a subset of NPC tumours. Fur-ther investigation of LTβR function and activation ofNF-κB signals in NPC will allow us to target thecritical components and therefore provide new oppor-tunities for therapeutic intervention.
Acknowledgements
This work was supported by the Michael and Betty KadoorieCancer Genetics Research Program II (MBKCGRPII), Li KaShing Institute of Health Science and Hong Kong ResearchGrant Council (Grant No. 470708).
References
1. Lo KW, To KF, Huang DP. Focus on nasopharyngeal carcinoma.Cancer Cell 2004;5:423–428.
2. Raab-Traub N. Epstein–Barr virus in the pathogenesis of NPC.Semin Cancer Biol 2002;12:431–441.
3. Lo KW, Chung GTY, To KF. Acquired genetic and epigeneticalterations in nasopharyngeal carcinoma. In NasopharyngealCarcinoma, Busson P (ed.). Landes Bioscience: Austin, Texas,USA, 2009.
4. Hui ABY, Lo KW, Leung SF, Teo P, Fung MK, To KF, et al.Detection of recurrent chromosomal gains and losses in primarynasopharyngeal carcinoma by comparative genomic hybridization.Int J Cancer 1999;82:498–503.
5. Lo KW, Teo PM, Hui AB, Tsang YS, Chan SY, Mak KF,et al. High resolution allelotype of microdissected primarynasopharyngeal carcinoma. Cancer Res 2000;60:3348–3353.
6. Chan AS, To KF, Lo KW, Mak KF, Pak W, Chiu B, et al. Highfrequency of chromosome 3p deletion in histologically normalnasopharyngeal epithelia from southern Chinese. Cancer Res2000;60:5365–5370.
7. Chan AS, To KF, Lo KW, Ding M, Li X, Johnson P, et al. Highfrequency of chromosome 9p losses in histologically normalnasopharyngeal epithelia from southern Chinese. Int J Cancer2002;102:300–303.
8. Hui ABY, Or YY, Takano H, Tsang RK, To KF, Guan XY, et al.Array-based CGH analysis identified cyclin D1 as a targetoncogene at 11q13.3 in nasopharyngeal carcinoma. Cancer Res2005;65:8125–8133.
9. Huang DP, Ho JH, Chan WK, Lau WH, Lui M. Cytogeneticsof undifferentiated nasopharyngeal carcinoma xenografts fromsouthern Chinese. Int J Cancer 1989;43:936–939.
10. Cheung ST, Huang DP, Hui ABY, Lo KW, Ko CW, Tsang YS,et al. Nasopharyngeal carcinoma cell line (C666–1) consistentlyharboring Epstein–Barr virus. Int J Cancer 1999;83:121–126.
11. Busson P, Ganem G, Flores P, Mugneret F, Clausse B, Caillou B,et al. Establishment and characterization of three transplantableEBV-containing nasopharyngeal carcinomas. Int J Cancer1988;42:599–606.
12. Lo KW, Kwong J, Hui AB, Chan SY, To KF, Chan AS, et al.High frequency of promoter hypermethylation of RASSF1A innasopharyngeal carcinoma. Cancer Res 2001;61:3877–3881.
13. Tsao SW, Wang X, Liu Y, Cheung YC, Feng H, Zheng Z, et al.Establishment of two immortalized nasopharyngeal epithelial celllines using SV40 large T and HPV16E6/E7 viral oncogenes.Biochim Biophys Acta 2002;1590:150–158.
14. Li HM, Man C, Jin Y, Deng W, Yip YL, Feng HC, et al.Molecular and cytogenetic changes involved in the immortalizationof nasopharyngeal epithelial cells by telomerase. Int J Cancer2006;119:1567–1576.
15. Rodriguez S, Khabir A, Keryer C, Perrot C, Drira M, Ghorbel A,et al. Conventional and array-based comparative genomichybridization analysis of nasopharyngeal carcinomas from theMediterranean area. Cancer Genet Cytogenet 2005;157:140–147.
16. Chen YJ, Ko JY, Chen PJ, Shu CH, Hsu MT, Tsai SF, et al.Chromosomal aberrations in nasopharyngeal carcinoma analyzedby comparative genomic hybridization. Genes ChromosomesCancer 1999;25:169–175.
17. Zarnegar B, Yamazaki S, He JQ, Cheng G. Control of canonicalNF-κB activation through the NIK–IKK complex pathway. ProcNatl Acad Sci USA 2008;105:3503–3508.
18. Keats JJ, Fonseca R, Chesi M, Schop R, Baker A, Chng WJ, et al.Promiscuous mutations activate the noncanonical NF-κB pathwayin multiple myeloma. Cancer Cell 2007;12:131–144.
19. Thornburg NJ, Pathmanathan R, Raab-Traub N. Activation ofnuclear factor-κB p50 homodimer/Bcl-3 complexes in nasopha-ryngeal carcinoma. Cancer Res 2003;63:8293–8301.
20. Lo AK, To KF, Lo KW, Lung RW, Hui JW, Liao G, et al.Modulation of LMP1 protein expression by EBV-encodedmicroRNAs. Proc Natl Acad Sci USA 2007;104:16164–16169.
21. Hehlgans T, Stoelcker B, Stopfer P, Muller P, Cernaianu G,Guba M, et al. Lymphotoxin-β receptor immune interactionpromotes tumor growth by inducing angiogenesis. Cancer Res2002;62:4034–4040.
22. Fujiwara S, Yamashita Y, Choi YL, Wada T, Kaneda R, Takada S,et al. Transforming activity of the lymphotoxin-β receptorrevealed by expression screening. Biochem Biophys Res Commun2005;338:1256–1262.
23. Dhawan P, Su Y, Thu YM, Yu Y, Baugher PJ, Ellis DL, et al.The lymphotoxin-β receptor is an upstream activator of NF-κB mediated transcription in melanoma cells. J Biol Chem283:15399–15408.
24. Friboulet L, Pioche-Durieu C, Rodriguez S, Valent A, Souquere S,Ripoche H, et al. Recurrent overexpression of c-IAP2 in EBV-associated nasopharyngeal carcinomas: critical role in resis-tance to Toll-like receptor 3-mediated apoptosis. Neoplasia2008;10:1183–1194.
SUPPORTING INFORMATION ON THE INTERNET
The following supporting information may be found in the online version of this article:
Supplementary Figure S1. Penetrancy summary of significant copy number changes in 18 NPC tumours
Supplementary Figure S2. NF-κB activation in NPC
Supplementary Table S1. Correlation of LTβR copy number changes, expression and clinical featuresin primary NPC tumours
Supplementary Table S2. Primer sequences for candidate genes on 12p13.3 amplicon
J Pathol 2010; 220: 97–107 DOI: 10.1002/pathCopyright 2009 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.





























