Embed Size (px)
Citation preview

Eur. J. Biochem. 226, 23-30 (1994) 0 FEBS 1994
Identification of an endonuclease responsible for apoptosis in rat thymocytes Daisuke SHIOKAWA‘.’, Harumi OHYAMA3, Takeshi YAMADA4, Kenji TAKAHASHI‘ and Sei-ichi TANUMA*,’ ’ Department of Biophysics and Biochemistry, Faculty of Science, University of Tokyo, Japan
Research Institute for Bioscience, Science University of Tokyo, Japan National Institute of Radiological Sciences, Inage, Chiba, Japan Department of Biology, School of Medicine, Toho University, Tokyo, Japan Department of Biochemistry, Faculty of Pharmaceutical Sciences, Science University of Tokyo, Japan
(Received June l/August 22, 1994) - EJB 94 078914
Analyses of cleavage ends of DNA fragments in apoptotic rat thymocytes induced by y-ray irradiation or by treatment with dexamethasone revealed that in both cases the fragments produced had 3’-hydroxyl (OH) and 5’-phosphoryl ( P ) ends of DNA chains. Rat thymocyte nuclei contained at least three endonuclease activities (deoxyribonucleases a, p and y ) that were able to cleave chromatin to mononucleosomal and oligonucleosomal fragments. The nuclei of apoptotic rat thymo- cytes induced by y-ray irradiation or dexamethasone retained considerable deoxyribonuclease y activity, but not a or p deoxyribonuclease activity. During the induction of apoptosis, treatment with cycloheximide, which suppressed apoptosis, resulted in marked decreases of deoxyribonucleases a and p activities. After release of cycloheximide inhibition, DNA fragmentation associated with apoptosis occurred in the cycloheximide-treated thymocyte nuclei, in which deoxyribonuclease y activity was only observed. The purified deoxyribonucleases a and p were divalent cation-indepen- dent acidic endonucleases, which were separated on a CM5PW column by HPLC. The molecular masses of deoxyribonucleases a and p were 28 kDa and 30 kDa, respectively, as determined by TSK G-2000SW gel-filtration HPLC, and both were 32 kDa in molecular mass as determined by SDS/ PAGE. In contrast, deoxyribonuclease y , a neutral endonuclease, required both Ca2+ and Mg2+ for full activity and was inhibited by Zn2+. The molecular mass of deoxyribonuclease y was 31 kDa and 33 kDa when measured by gel filtration and SDS/PAGE, respectively. Under these optimal conditions, deoxyribonuclease y was shown to produce 3’-OH/S’-P ends of nucleosomal DNA frag- ments, while deoxyribonucleases a and p both formed DNA fragments with 3’-P/S’-OH ends. The ends formed by cleavage with deoxyribonuclease y were the same as those produced in apoptotic rat thymocytes. On the basis of these results, it seems likely that deoxyribonuclease y is responsible for internucleosomal cleavage of chromatin during thymic apoptosis.
Cell death by apoptosis occurs during embryogenesis, metamorphosis, normal tissue turnover, carcinogenesis, thy- mic selection, growth factor deprivation, cell-mediated cyto- toxicity, human immunodeficiency virus infection, tumor re- gression and ionizing radiation exposure [ l - 141. Elucidation of the molecular mechanism of apoptosis is critical for under- standing the physiological significance of cell death in the control of cell social community and developing chemothera- peutic agents for apoptosis-related diseases. At present, known processes associated with apoptosis are specific mor- phological changes and nuclear DNA fragmentation [ l , 2, 8, 10, 11, 131. Internucleosomal cleavage of chromosomal
Correspondence to S. Tanuma, Department of Biochemistry, Fa- culty of Pharmaceutical Sciences, Science University of Tokyo, Shinjuku-ku, Tokyo, Japan 162
Abbreviations. PhMeSO,F, phenylmethylsulfonyl fluoride ; IC,, concentration for half-maximal inhibition ; CHO, Chinese hamster ovary ; DNase, deoxyribonuclease.
Enzymes. Proteinase K (EC 3.4.21.64); RNase A (EC 3.1.27.5); terminal deoxynucleotidyl transferase (EC 2.7.7.31) ; T4 polynucleo- tide kinase (EC 2.7.1.78) ; calf intestinal alkaline phosphatase (EC 3.1.3.1); deoxyribonuclease I (DNase I) (EC 3.1.21.1); deoxyribo- nuclease I1 (DNase 11) (EC 3.1.22.1).
DNA, which is suggested to be catalyzed by a constitutive endonuclease, is the biochemical feature most commonly as- sociated with apoptotic processes leading to cell death [15- 181. The identification of this endonuclease is, therefore, im- portant for understanding the mechanism regulating apo- ptosis.
One of the best characterized systems for apoptosis is immature thymocytes that undergo apoptosis when exposed to irradiation, glucocorticoids or antibodies to the CD-3 T cell receptor complex [S, 15-20]. Studies using such sys- tems have suggested that a sustained increase in intracellular Ca2+ during apoptosis may activate a constitutive endonucle- ase that mediates DNA fragmentation [15 -201. A likely can- didate for such an enzyme is Ca2+/Mg2+-dependent endonu- clease(s) [21-291, such as Nuc 18 [26] and deoxyribonucle- ase I (DNase I) [27]. The involvement of this type of endonu- clease is supported by the observations that Ca2+ ionophores induce DNA fragmentation and cell death in thymocytes and that Ca2+ chelators can prevent both processes [16-201. Furthermore, apoptosis in thymocytes is known to be inhib- ited by Zn2+ [16]. Thus, an endogenous Ca2+-dependent en- donuclease that is inhibited by Zn2+ is suggested to function in DNA fragmentation during thymic apoptosis.

24
One promising approach for identifying this endonucle- ase is to study the nature of DNA fragmentation at the cellu- lar level and compare it with the fragmentation produced by an endonuclease(s) purified from apoptotic cell nuclei. In the present study, we use rat thymocytes to examine the molecu- lar mechanism by which chromosomal DNA is cleaved into nucleosomal oligomers. Our data show that DNA fragments cleaved at the 3’-OWS’-P are produced in apoptotic rat thy- mocytes induced by irradiation or glucocorticoids. During studies on a nuclear endonuclease(s) capable of internucleo- soma1 DNA cleavage in apoptotic rat thymocytes, we iso- lated a novel CaZ+/Mg2+-dependent endonuclease (DNase y ) from nuclei, which produces DNA fragments with 3’-OH/5’- P ends and is inhibited by Zn2+. The identity of the ends of these products of DNase y with the ends produced in apop- totic rat thymocytes and the similarity for the ion dependence suggest that this enzyme is involved in apoptotic DNA frag- mentation in rat thymocytes.
EXPERIMENTAL PROCEDURES
Materials [a-3ZP]dCTP (3000 Cilmmol) and‘ [y-”PIATP (3000 Cil
mmol) were purchased from DuPont/New England Nuclear. Terminal deoxynucleotidyl transferase, T4 polynucleotide ki- nase and calf intestinal alkaline phosphatase were from Ta- kara. S Sepharose was from Pharmacia. CMSPW, heparin 5PW and G-2000SW were from Tosoh. Dexamethasone and cycloheximide were from Sigma. All other reagents were the highest purity grade available.
Induction of apoptosis Thymocytes obtained from 10-week-old male albino
wistar strain rats were suspended in Krebs-Ringer phosphate solution containing 10 mM glucose [30]. Apoptosis was in- duced by incubation of the cells at 37°C after irradiation with y-rays of at a dose of I0 Gy or in the presence of 0.1 pM dexamethasone. The apoptotic thymocytes were collected for morphological examination by electron microscopy or analy- sis of DNA fragmentation by 2% agarose gel electrophoresis as described previously [14].
End labeling of DNA For analysis of cleavage ends of DNA fragments, DNA
was isolated by phenol/chloroform extraction. The 3’ ends of the DNA fragments (5 pg, equivalent to approximately 1 X lo6 cells) were labeled by incubation with 5 U terminal deoxynucleotidyl transferase and 0.83 mCi/ml [a-32P]dCTP in the presence of 25 mM Tris/HCI, pH 7.6, 10 mM dithio- threitol and 1 mM CaCl, (total volume 30 pl). The 5‘ ends of the DNA fragments (5 pg) were labeled by incubation with 5 U T4 polynucleotide kinase and 0.83 mCi/ml of [y-12P]ATP in the presence of 100 mM Tris/HCl, pH 7.6, 20 mM MgCI,, 10 mM dithiothreitol and 0.2 mM spermidine (total volume 30 pl). The phosphoryl groups in the ends of DNA chains were removed by pretreatment with 20 U calf intestinal alka- line phosphatase in the presence of 36 mM Tris/HCl, pH 8.0, and 1 mM MgC1,. Unincorporated labeled nucleotides were removed by precipitation with ammonium acetate/isopropa- no1 and the labeled DNA was subjected to 2% agarose gel electrophoresis. After transferring DNA in the gels to nylon membranes, the membranes were autoradiographed.
Endonuclease assay Endonuclease activity was measured as the amount of
fragmentation of chromosomal DNA of HeLa S3 cell nuclei (1.67 X 107/ml; assay 1) or of supercoiled pBSIISK(-) plas- mid DNA (6.67 mg/ml; assay 2) separated by 2% and 0.8% agarose gel electrophoresis, respectively [ 101. The reaction mixture (total volume 30 pl) for assays of DNases a and /3 contained 50 mM Mes/NaOH, pH 5.6, 3 mM MgCL 1 mM 2-mercaptoethanol and 0.1 mM phenylmethylsulfonyl fluo- ride (PhMeS0,F) and the mixture for the assay of DNase y contained 50 mM Mops/NaOH, pH 7.2, 3 mM CaCl,, 3 mM MgCl,, 1 mM 2-mercaptoethanol and 0.1 mM PhMeS0,F. The reactions were carried out at 37°C for 60 min and 10 min in assay 1 and assay 2, respectively, and were termi- nated by chilling on ice. For assay 1, the resulting reaction mixture was centrifuged at 20000Xg for 20 s and precipi- tated nuclei were lysed in S O mM Tris/HCl, pH 7.8, 10 mM EDTA, 0.5 % (mass/vol.) sodium N-lauroyl sarcosinate (buffer A). The DNA was prepared by successive treatments with 0.5 mg/ml ribonuclease A for 20 min and 0.5 nig/ml proteinase K for 30 min and was loaded onto a 2% agarose gel. DNA fragmentation patterns stained with ethidium bro- mide were examined in photographs taken under ultraviolet illumination [14]. The amount of fragmentation was deter- mined by densitometry of DNA fragments with molecular sizes of less than 5 kb. For assay 2, plasmid DNA was ex- tracted by phenol/chloroform, and the aqueous phase was subjected to 0.8 % agarose gel electrophoresis. After staining with ethidium bromide, the hydrolysis of plasmid DNA was detected under ultraviolet illumination.
Endonuclease purification For purification of nuclear DNases from rat thymocytes,
nuclei were isolated from 2X 10’” normal or apoptotic cells and were homogenized in 10 mM Tris/HCl, pH 7.8, 2 mM 2- mercaptoethanol, 0.3 mM PhMeSO,F, 3 mM MgC1, (buffer B) containing 0.1% Nonidet P-40. The DNase activities in the nuclei were solubilized by sonication in buffer B contain- ing 0.5 M (NH,),SO,, and the nuclear debris was removed by centrifugation at 150000Xg for 1 h. The supernatant frac- tion was applied onto S Sepharose equilibrated with 20 mM Tris/HCl, pH 7.8, 1 mM 2-mercaptoethanol, 0.1 mM PhMe- SO,F, 10% ethylene glycol (buffer N). The column was washed with the same buffer and developed with a linear gradient of 0-1 M KC1 in buffer N. DNase activity was eluted with 0.6 M KC1. This enzyme fraction was sub,jected to CMSPW HPLC in a column equilibrated with buffer N and proteins were eluted with a linear gradient of 0-1 M KC1 in buffer N. DNases a, /3 and y were eluted with 0.24, 0.34 and 0.55 M KCl, respectively. Each enzyme fraction was subjected to heparin 5PW HPLC in a column ttquili- brated with buffer N and proteins were eluted with a linear gradient of 0-1 M KC1 in the same buffer. The eluate\ were further purified by gel filtration HPLC on G-2000SW equili- brated with 0.3 M NaCl in 20 rnM Mops/NaOH, pH 7.0, 1 mM 2-mercaptoethanol, 0.1 mM PhMeSO,F, 5% ethylene glycol (buffer S). The molecular mass standards used for cal- ibration were IgG (158 kDa), BSA (66 kDa), ovalbumin (44 kDa) and myoglobin (17 kDa). The enzyme fractions were finally subjected to CMSPW HPLC in a column equili- brated with buffer S containing 0.3 M NaCl and proteins were eluted with a linear gradient of 0.3-1.5 M NaCl in buffer S. The enzyme fractions were desalted by dialysis against buffer S and stored at 0-4°C.

25
DNase activity gel system
For identification of properties having DNase activities and estimation of molecular masses, the activity gel system for DNase [31] was used with several modifications. Briefly, the purified DNases were separated by electrophoresis in a Laemmli SDS gel containing 200 pg/ml native calf thymus DNA. After electrophoresis, the gels were washed with 10 mM Tris/HCl, pH 7.8, and 5 mM 2-mercaptoethanol at 50°C for 1 h to remove SDS, and subsequently with 10 mM Tris/HCl, pH 7.8, at 4°C overnight to allow refolding of en- zymes. The gels were incubated in 10 mM TrisMCl, pH 7.8, 3 mM CaC1, and 3 mM MgCI, at 37°C for appropriate peri- ods of time. Apparent nuclease activities were detected as dark areas on a fluorescent background after transillumina- tion of gels with ultraviolet. A calibration curve determined by molecular mass standards was not affected by the pres- ence of 200 yglml double-stranded DNA in SDS gels.
RESULTS
Characterization of apoptosis in rat thymocytes
To characterize the apoptosis of rat thymocytes induced by ;/-ray irradiation or dexamethasone, we examined the mor- phological changes of the cells and the nature of the DNA fragmentation which are characteristic of apoptosis. Typical morphological changes in apoptotic thymocytes induced by irradiation were observed in transmission and scanning electron microscopies. Analysis revealed cell shrinkage, chromatin condensation, collapse along the nuclear envelope, and the disappearance of cell surface microvilli, as compared with normal thymocytes. Similar morphological changes characteristic of apoptosis were observed in dexamethasone- treated rat thymoiytes. In both cases, agarose gel electropho- resis of cellular DNA from the apoptotic thymocytes showed ‘ladder’ patterns, indicating the result of apoptotic inter- nucleosomal cleavage of chromosomal DNA.
The modes of DNA cleavage during apoptosis induced by irradiation or dexamethasone were first investigated by end-labeling methods (Fig. 1). If DNA fragments have free 3’-OH and 5’-P ends, the resulting nucleosome ladders should be detected simply (i.e. without alkaline phosphatase pretreatment) by 3‘ end labeling of the extracted DNA by terminal deoxynucleotidyl transferase and only one ”P-la- beled nucleotide triphosphate, [u-’*P]dCTP, and by 5’ end labeling by T4 polynucleotide kinase and [y-12P]ATP only after pretreatment of the DNA with alkaline phosphatase. In the opposite case, for the end labeling, 3‘ ends of DNA chains should not be labeled without alkaline phosphatase pretreatment. The resultant autoradiograms revealed that for apoptosis, the 5’ ends of the fragments (Fig. 1 A and B) could not be labeled without alkaline phosphatase pretreatment. Thus, the fragments produced at a cellular level had 3’-OH and 5‘-P ends. These results provide evidence that the apoptosis induced in these conditions is catalyzed by an en- donuclease that generates 3’-OH/5’-P cleavage ends of DNA chains. These findings are important for identifying the endo- nuclease responsible for apoptosis.
Purification of nuclear endonucleases
We next attempted to purify the endonuclease responsible for this internucleosomal cleavage from rat thymocytes. For detection of the endonuclease activity that cleaves linker re-
APase
A
DNA fragments
End-labeling
Results
APase labeling
B
(k) alkaline phosphatase (APase)
3’end-labeling Send-labeling terminal deoxy- polynucleotide nucleotidyl- kinase transferase [c~-~’P]~CTP [Y-~~P]ATP
3’end Send 3’end Send
+ - + - + - + - Yes Yes Yes No Yes No Yes Yes
Fig. 1. Analyses of cleavage ends of DNA fragments in apoptotic rat thymocytes. DNA was extracted from apoptotic rat thymocytes 4 h after irradiation (10 Gy) (A) or dexamethasone (0.1 pM) treat- ment (B). The DNA was incubated with (lanes 1 and 3) or without (lanes 2 and 4) alkaline phosphatase (APase) prior to 3’ end (lanes 1 and 2) or 5‘ end (lanes 3 and 4) labeling as described in the Experimental Procedures section. Aliquots of DNA were subjected to 2% agarose gel electrophoresis and autoradiography.
gions of chromatin, we used HeLa S3 cell nuclei as substrate, because these nuclei contain little endogenous endonuclease activity. An endonuclease activity specific for the inter- nucleosomal regions should produce nucleosomal ladders on agarose gel electrophoresis. Endonuclease activities present in isolated nuclei from rat thymocytes were solubilized with salts. Essentially complete solubilization of nuclear endonu- clease activities was obtained when a sonication step at 0.5 M (NH,),SO, was included. The soluble enzyme preparation was used for subsequent chromatographies.
Three endonuclease activities were resolved in the third step of CM5PW HPLC (Fig. 2A). These DNase activities, tentatively named DNase a, p and y in order of elution, cata- lyzed the cleavage of linker DNA of chromatin in HeLa S3 cell nuclei and also cleaved supercoiled plasmid DNA endo- nucleolytically. Interestingly, the induction of apoptosis by irradiation resulted in decreases in the activities of DNases a and p, without affecting DNase y activity appreciably (Fig. 2B). Similar results were obtained with apoptotic rat thymocytes induced by dexamethasone (data not shown). The active fractions of DNases u and p from normal thymo- cyes and DNase y from apoptotic rat thymocytes were further purified by sequential HFLC steps on heparin 5PW, G- 2000SW gel filtration and a second CM5PW chromatogra- phy. There was no evidence for complexes of a single DNase that can dissociate or for the ability of the various forms to interconvert in any of the chromatographies used. The physi-

26
I Fract. NO 20 30 40
100
P lool B 9
60
40
20
0 0 10 20 30 40 50 60
1 .oo
0.75
0.25 0 o y
.c co hl
0 - 0) 0 C
0 c 0 .- - 2
0 a - 1.00 C
0
0
L
0.75 :: C
0.25
0
Fraction Number
Fig.2. CMSPW HPLC of nuclear DNases from normal and apoptotic rat thymocytes. The active fractions of DNases obtained by S Sepharose chromatography from nuclei of normal (A) and apoptotic (B) rat thymocytes 4 h after irradiation were subjected to CMSPW HPLC. DNase activities of fractions were detected by a HeLa S3 nuclear assay after agarose gel electrophoresis (insert) and determination of the amount of fragmentation, as described in the Experimental Procedures section.
cal and catalytic properties of these enzyme preparations were studied.
Changes of nuclear endonucleases during apoptosis DNases a and j? appeared to turn over at high rates and
the 7 DNase was retained in apoptotic cell nuclei (Fig. 2). Previous work has shown that little internucleosomal cleav- age of DNA was observed following irradiation in rat thymo- cytes treated with cycloheximide [30]. These observations offered an experimental model to investigate the possible changes of these three nuclear DNases during apoptosis.
After a 3-h treatment with cycloheximide of irradiated rat thymocytes, nuclei were isolated and DNase activities were separated using CMSPW HPLC. As shown in Fig. 3A, cyclo- heximide treatment resulted in marked decreases in DNases a and j?. In contrast, a sample with considerable activity of DNase y was extracted from the cycloheximide-treated cell nuclei. In this time period, DNA fragmentation was sup- pressed (Fig. 3A). Incubation was subsequently carried out for another 3 h after washing in Krebs-Ringer phosphate so- lution of the thymocytes to remove cycloheximide. In the absence of cycloheximide, a DNA ladder characteristic of apoptosis was clearly observed (Fig. 3B). During this further incubation of thymocytes without cycloheximide, DNase y activity was only retained in the apoptotic thymocyte nuclei (Fig. 3B) comparable to that shown in Fig. 3A.
Properties of nuclear endonucleases Molecular muss
The molecular masses of these three DNases were deter- mined by the SDS/PAGE-renatured method (activity gel sys-
1 .oo
0.75
, 1.0
0.25
u- W N
0 -
0 C
0 a, c .- -
E - 1.00 c a,
0 0.75
0.50 I 0.5
0.25 ]
0
Fraction Number
Fig. 3. Changes in DNase a, p and y activities during apoptosis in cycloheximide-treated rat thymocytes. Irradiated rat thymo- cytes were incubated for 3 h in the presence of 20 pM cycloheximide (A), washed in Krebs-Ringer phosphate solution to remove cyclo- heximide and further incubated for 3 h (B). The chromosomal DNA was examined by agarose gel electrophoresis for internucleosomal cleavage (left insets), and DNase activities extracted from nuclei were subjected to CM5PW HPLC and measured by HeLa S3 nuclear assay (right insets). The amount of fragmentation was determined as described in the Experimental Procedures section.
kDa
f 97.4 f 66.2
f 45.0
f 31.0
f 21.5
1 2 3 Fig.4. Activity gel analysis of DNases a, p and y. The molecular masses of DNases a (lane 1) and 11 (lane 2) purified from normal rat thymocyte nuclei, and the masses of DNase y (lane 3) from irradi- ated apoptotic cells were analyzed by the activity gel system as described in the Experimental Procedures section. The molecular mass markers were phosphorylase b (97400), BSA (66200). oval- bumin (45 OOO), carbonic anhydrase (31 000), soybean trypsin inhibi- tor (21 500) and lysozyme (14400 Da).
tem). This activity gel system was based on the ability of DNase to be renatured after removal of SDS and to cleave DNA during incubation. As shown in Fig. 4, the localizations of DNases in gels could be detected by the disappearance of DNA fluorescence as dark bands on an ethidium bromide fluorescent background. On addition of Ca2+ and Mg" to the incubation medium with DNase y (Fig. 4), a non-fluores- cent band in a portion corresponding to a 33-kDa protein was observed. However, both DNases a and j? (Fig. 4) exhibited non-fluorescent bands corresponding to a protein of 32 kDa. On G-2000SW gel-filtration HPLC, DNases a, j? and y ap-
27
Acetate-KOH I Mes I MODS I Tris-HCI I Ches
A
Acetate-KOH I Mes I Mops I Tris-HCI I Ches
B
100
75
50
25
- 0
100 .- 3 5
m 0) v)
.- c 0 75
2 50
t o
n .- $ 25 m c -
Acetate-KOH I Mes I MODS I Tris-HCI I Ches
C
0 0 0.01 0.1 1 10
- Concentration of divalent cations (mM)
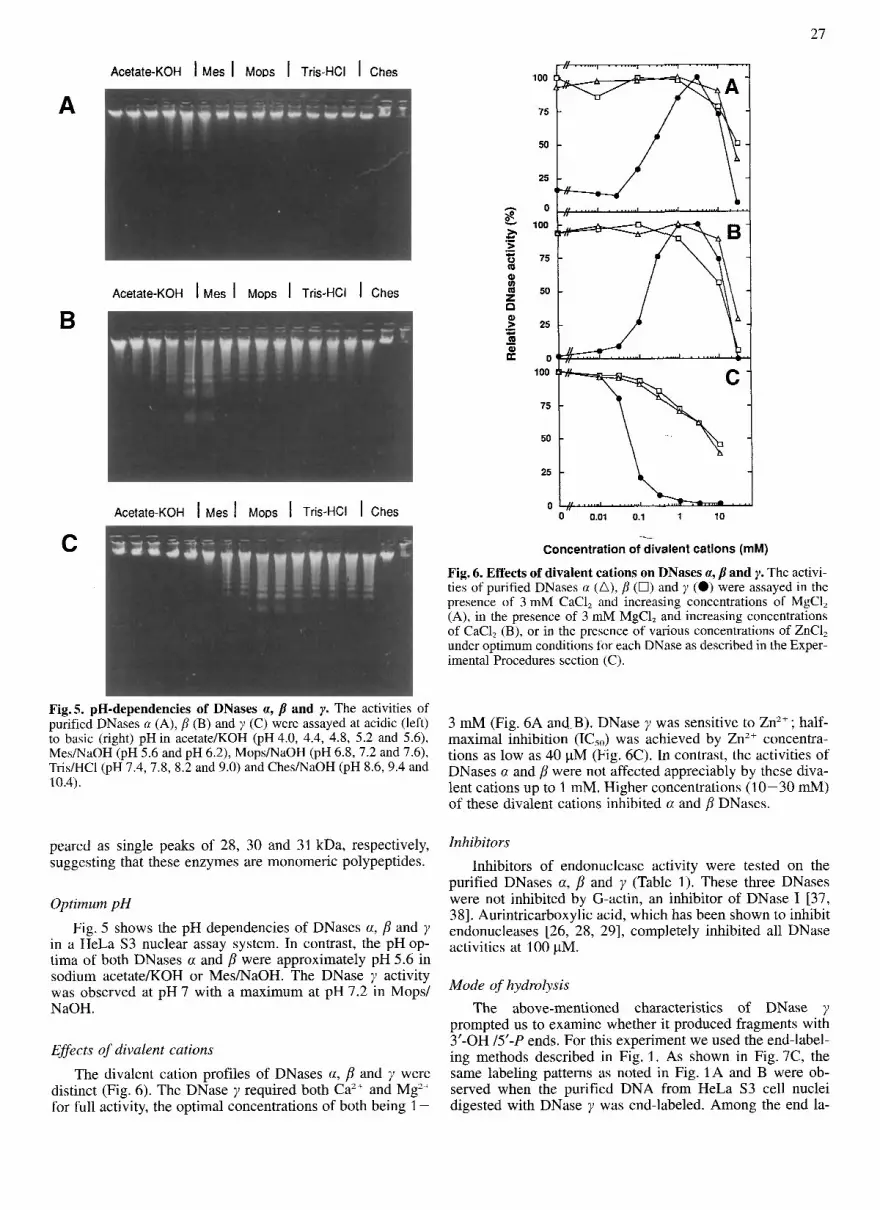
Fig. 6. Effects of divalent cations on DNases a, p and y. The activi- ties of purified DNases a (A), p (0) and y (0) were assayed in the presence of 3 mM CaC1, and increasing concentrations of MgCl, (A), in the presence of 3 mM MgC1, and increasing concentrations of CaC1, (B), or in the presence of various concentrations of ZnC1, under optimum conditions for each DNase as described in the Exper- imental Procedures section (C).
Fig.5. pH-dependencies of DNases a, and y. The activities of purified DNases a (A), p (B) and y (C) were assayed at acidic (left) to basic (right) pH in acetate/KOH (pH 4.0, 4.4, 4.8, 5.2 and 5.61, Mes/NaOH (PH 5.6 and PH 6.21, MoPsmaOH (PH 6.83 7.2 and 7.61, Tris/HCl (pH 7.4,7.8, 8.2 and 9.0) and Ches/NaOH (PH 8 6 9 . 4 and 10.4).
3 mM (Fig. 6A and.B). DNase y was sensitive to Zn2+ ; half- maximal inhibition (1C5") was achieved by Zn2+ concentra- tions as low as 40 pM (Fig. 6C). In contrast, the activities of DNases a and p were not affected appreciably by these diva- lent cations up to 1 mM. Higher concentrations (10-30 mM) of these divalent cations inhibited a and p DNases.
peared as single peaks of 28, 30 and 31 kDa, respectively, suggcsting that these enzymes are monomeric polypeptides.
Optimum pH
Fig. 5 shows the pH dependencies of DNases a, p and y in a HeLa S3 nuclear assay system. In contrast, the pH op- tima of both DNases a and p were approximately pH 5.6 in sodium acetate/KOH or Mes/NaOH. The DNase y activity was observed at pH 7 with a maximum at pH 7.2 in Mops/ NaOH.
Effects of divalent cations The divalent cation profiles of DNases a, p and y were
distinct (Fig. 6). The DNase y required both Caz+ and Mg2+ for full activity, the optimal concentrations of both being 1 -
Inhibitors Inhibitors of endonuclease activity were tested on the
purified DNases a, p and y (Table 1). These three DNases were not inhibited by G-actin, an inhibitor of DNase I 137, 381. Aurintricarboxylic acid, which has been shown to inhibit endonucleases [26, 28, 291, completely inhibited all DNase activities at 100 pM.
Mode of hydrolysis The above-mentioned characteristics of DNase y
prompted us to examine whether it produced fragments with 3'-OH /5'-P ends. For this experiment we used the end-label- ing methods described i n Fig. 1. As shown in Fig. 7C, the same labeling patterns as noted in Fig. 1A and B were ob- served when the purified DNA from HeLa S3 cell nuclei digested with DNase y was end-labeled. Among the end la-

x 9 C .- .- e P c k
S G
0 -0
5
+
I
02 N
0 3 8
2
3
Y I
t-
APase
A B c
1 2 3 4 1 2 3 4 1 2 3 4 Fig. 7. Modes of DNA cleavage by DNases a, fi and y. DN.\ was extracted from HeLa S3 cell nuclei digested by DNase a (A). fi (B) or y (C) and subjected to 3’ end (lanes 1 and 3) or 5’ end (lmes 3 and 4) labeling wlth (lanes 1 and 3, +) or without (lanes 2 and 4, -) Pretreatment with alkaline phosphatase (APase). Aliquots of DNA were analyzed by 2% agarose gel electrophoresis and autora- diography as described in the Experimental Procedures section.
belings, the 5’ ends of the fragments (Fig. 7C) could not be labeled without alkaline phosphatase pretreatment. Thus, the DNase y produced 3’-OH/S’-P ends of DNA chains. In con- trast, the DNA fragments formed by DNases 0: and p had 3’- P/S’-OH ends as evidenced by labeling of their 3‘ ends by terminal deoxynucleotidyl transferase only after alkaline phosphatase pretreatment and 5’ end fill-in reactions without alkaline phosphatase pretreatment (Fig. 7A and B).
DISCUSSION In a search for an endonuclease responsible for apoptosis
in rat thymocytcs, we found three activities of endonuclease, namely DNases a, p and y . These DNases are probably pre- sent in nuclei in monomeric polypeptide forms that cleave linker regions of chromatin to produce nucleosomal oligo- mers. The cognate forms of DNases a and p were easily solubilized from nuclei, suggesting a location in the nucleo- plasm. In contrast, DNase y was tightly bound to sgme un- identified nuclear structure. The DNases a and p differed from DNase y in physical and enzymic properties, especially in the modes of cleavage of chromosomal DNA and require- ments for divalent metal ions. These properties of DNases a, p and 7 allow comparison with other previously purified endonucleases. Endonucleases which are active in the pres- ence of only one divalent cation, such as Ca’+, Mg” and Mn’+ [32-35]. and are involved in DNA repair will not be discussed.
So far, several endonucleases that absolutely requirc both Caz+ and Mgz+ for full activity have been purified from some mammalian tissues and cell lines [21-291. However, these endonucleases differ from DNases y in their affinities for ion- exchange resins, molecular masses and optimum pH. The modes of hydrolysis of DNA by putative apoptotic endonu- cleases, such as Ca2+/Mg2+-dependent endonucleasek [24, 25, 291, Nuc 18 [26] [the amino acid sequence of Nuc 18 has recently been shown to have considerable similarity to the cyclophilin family of proteins which are ubiquitous intra- cellular binding proteins for the immunosuppressive drug cy- closporin A (Cidlowski, J. A,, Montagne, J. W., Hughes, F. M., Evans, R. B. and Bortner, C. D., unpublished results)] and CazT/Mn2’-depeudent endonuclease [28] are still un- known. DNase I, which is known to be active in the presence

29
of Ca” or Mg’+, cleaves DNA to produce 3’-OH/5’-P ends [36]. Unlike DNase I [37-391, G-actin did not inhibit DNase y . Thus, many of -the properties of these putative apoptotic enzymes appear to be distinct from those of purified DNases y (Table 1). The y type DNase is present in nuclei from calf thymus and rat spleen (unpublished results). It is conceivable that the nuclear y type DNase acts to produce DNA fragmen- tation during apoptosis in lymphocytes.
However, the properties of DNases a and p are similar to those of DNase I1 [40]. DNase I1 is known to be a lysosomal enzyme. Moreover, DNase I1 has a molecular mass of 36 kDa, clearly distinguishing it from DNases a and p. Re- cently, Barry and Eastman identified a DNase-11-like endonu- clease from Chinese hamster ovary (CHO) cell nuclei that could produce DNA fragmentation similar to that observed in apoptotic cells [41]. This acidic CHO endonuclease was not dependent on Ca2+ nor Mg“, and was insensitive to Zn’+ (Table 1). These properties are similar to the properties of DNases a and p reported in this study. The cognate form of DNases a and p are also present in calf thymocytes and rat splenocytes. Although the physiological significances of DNases a and p are still unknown, these enzymes are impor- tant for general roles in nuclear functions. The changes in the activities of these endonucleases on various stimuli (un- published results) may provide some clues to their functions.
A common event in thymic apoptosis is internucleosomal cleavage of nuclear DNA. This irreversible molecular pro- cess required for cell death is suggested to be catalyzed by a constitutive endogenous Ca2+/Mg2+-dependent endonuclease [16-191. As the ends of DNA fragments cleaved by DNase y are the same as those produced in apoptotic rat thymocytes, and micromolar amouts of Zn’+, known to prevent apoptosis, inhibit DNase y , this enzyme is considered to be the endonu- clease responsible for DNA fragmentation during thymic apoptosis. This conclusion for the role of DNase y in thymic apoptosis is supported by the observations that only DNase y activity was retained in apoptotic thymocyte nuclei. How- ever, in some cell lines, DNA fragmentation occurred with- out increase in the intracellular Ca2+ concentration [44, 451. These observations imply that an apoptotic endonuclease that causes DNA fragmentation may not always be a Ca2+/MgZ+- dependent endonuclease and there may be differences in dif- ferent cell types and states. Nevertheless, it will be essential to investigate whether intranuclear compartmentation of Ca’+ occurs and is sufficient for DNase y activation.
Confirmation of the essential role of DNase 7 in apoptosis in vivo and of the distinct DNA fragmentations by these three DNases must await further studies. Studies are also needed to establish whether these DNases are products of separate genes or whether alternative splicings are also necessary. These problems are best resolved by cloning the relevant DNases. Furthermore, the mechanisms of activation of these three nuclear DNases in thymocyte nuclei are un- clear. Possibly, the DNases interact with endogenous inhibi- tors or activators, and/or are modified post-translationally by poly(ADP-ribosy1)ation [44-461 or phosphorylation. Ca”/ Mg’+ -dependent or CaZ+/Mn2+-dependent endonuclease ac- tivities have been shown to be regulated by poly(ADP-ribo- sy1)ation [47,48]. Additional studies are required to elucidate these possibilities. Such information may provide important evidence for understanding the biological significance of these nuclear DNases in cell death or survival.
This study was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science and Culture of
Japan, and funds from the Naito Foundation, Cell Science Founda- tion, Terumo Life Science Foundation and the SUT Special Grant for Research Promotion 1994.
REFERENCES 1.
2.
3. 4.
5. 6.
7. 8.
9.
10.
11.
12. 13. 14.
15. 16. 17.
18. 19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
Kerr, J. F. R., Wyllie, A. H. & Currie, A. R. (1972) Bi: J. Cancer
Wyllie, A. H., Kerr, J. F. R. & Currie, A. R. (1980) Int. Rev.
Berger, N. A. (1985) Radiat. Res. 101, 4-15. Carson, D. A,, Seto, S., Wasson, D. B. & Camera, C. J. (1986)
Ucker, D. S. (1987) Nature 329, 62-64. Smith, C. A,, Williams, G. T., Kingston, R., Jenkinson, E. J. &
Dive, C. & Hickman, J. A. (1991) Bi: J. Cancer 64, 192-196. Arends, M. J. & Wyllie, A. H. (1991) Int. Rev. Exp. Pathol. 32,
Groux, H., Torpier, G., Monte, D., Mouton, Y., Capron, A. &
Williams, G. T., Smith, C. A,, McCarthy, N. J. & Grimes, E. A.
Cohen, J. J., Duke, R. C., Fadok, U. A. & Sellins, K. S . (1992)
Raff, M. C. (1992) Nature 356, 397-400. Cohen, J. J. (1993) Imrnunol. Today 14, 126-130. Tanuma, S., Shiokawa, D., Tanimoto, Y., Ikekita, M., Sakagami,
H., Takeda, M., Fukuda, S. & Kochi, M. (1993) Biochem. Biophys. Res. Commun. 194, 29-35.
26, 239-257.
Cytol. 68, 251 - 306.
J. Exp. Med. 163, 746-751.
Owen, J. J. (1989) Nature 337, 181-184.
223-254.
Ameisen, J. C. (1992) J. Exp. Med. 175, 331-340.
(1992) Trends Cell Biol. 2, 263-267.
Annu. Res. Immunol. 10, 267-293.
Wyllie, A. H. (1980) Nature 284, 555-556. Cohen, J. J. & Duke, R. C. (1984) J. Immunol. 132, 38-42. Arends, M. J., Moms, R. G. & Wyllie, A. H. (1990) Am. J.
Compton, M. M. (1992) Cancer Metastasis Rev. 11, 105-119. Wyllie, A. H., Morris, R. G., Smith, A. L. & Dunlop, D. (1984)
J. Pathol. 142, 67-77. McConkey, D. J., Hartzell, P. & Orrenius, S. (1990) Arch. Bio-
chern. Biophys. 278, 284-287. Burgoyne, L. A., Wagar, M. A. & Atkinson, M. R. (1970) Bio-
chem. Biophys. Res. Commun. 39, 254-259. Hewish, D. R. & Burgoyne, L. A. (1973) Biochem. Biophys.
Res. Commun. 56, 703-710. Ishidd, R., Akiyoshi, H. & Takahashi, T. (1974) Biochem. Bio-
phys. Res. Commun. 59, 658-665. Nakamura, M., Sakaki, Y., Watanabe, N. & Takagi, Y. (1981) J.
Biochem. (Tokyo) 89, 143-152. Hashida, T., Tanaka, Y., Matsunami, N., Yoshihara, K., Kamiya,
T., Tanigawa, Y. & Koide, S. S. (1982) J. Biol. Chem. 257, 13 114-13 119.
Gaido, M. L. & Cidlowski, J. A. (1991) J. Biol. Chem. 266, 18580- 18 585.
Peitsch, M. C., Polzar, B., Stepham, H., Crompton, T., MacDon- ald, H. R., Mannherz, H. G. & Tshopp, J. (1993) EMBO J.
Nikonova, L. V., Belelsky, I. P. & Umansky, S. R. (1993) Eui:
Ribeiro, J. M. & Carson, D. A. (1993) Biochemistry 32, 9129-
Yamada, T. & Ohyama, H. (1988) Int. J. Radiat. Biol. 53, 65-
Rosenthal, A. L. & Lacks, S. A. (1977) Anal. Biochem. 80, 76-
Wang, E. C., Furth, J. J. & Rose, J. A. (1978) Biochemistry 17,
Nakayama, J., Fujiyoshi, T., Nakamura, M. & Anai, M. (1981)
Hibino, Y., Yamamura, T. & Sugano, N. (1989) Biochem. Bio-
Cottlieb, J. & Muzyczaka, N. (1990) J. Biol. Chem. 265,
Pathol. 136, 593 - 601.
12, 371-377.
J. Biochem. 215, 893-901.
9136.
75.
90.
544- 547.
J. Biol. Chem. 256, 1636-1642.
phys. Acta. 1008, 287-292.
10836- 10841.

30
36. Laskowski, M. (1971) in The enzymes (Boyer, P. D., Landy, H. & Myrback, K., eds) vol. 4, pp. 289-311, Academic Press, New York.
43. Bansal, N., Houle, A. G. & Melnykovych, G. (1990) J. Cell.
44. Sugimura, T. (1973) Prog. Nucleic Acid Res. Mol. Biol. 13,
45. Ueda, K. & Hayaishi, 0. (1985) Annu. Rev. Biochem. 54 73-
46, T ~ ~ ~ ~ ~ , s, (1989) Biochem, ~ i ~ ~ , $ ~ ~ . R ~ ~ , commun. 163,
47. Yoshihara, K., Tanigawa, Y., Burzio, L. & Koide, S. S. ( 1975)
48. Nelipovich, P. A,, Nikonova, L. V. & Umansky, S. R. (1988)
Physiol. 143, 105 - 109.
127- 151.
100.
1047 - 1055.
37. Lacks, S . A. (1981)J. Biol. Chem. 256, 2644-2648. 38. Anai, M., Sas&, M., Muta, A. 8-z MiYagawa, T. (1981) Bio-
39. Kreuder, v., Dieckhoff, J., Sittig, M. 82 Mannherz, H. G. (1984)
40. Murai, K., Yamanaka, M., Akagi, K. & Anai, M. (1980) J. Bio-
41. Barry, A. & Eastman, A. (1993) Arch. Biochem. Biophys. 300,
42. Alnemri, E. S . & Litwack, G. (1990) J. Biol. Chem. 265,
chirn. Biophys. Acta 656, 183-188.
Eur. J. Biochem. 139, 389-400.
chem. (Tokyo) 87, 1097-1103.
440 - 450.
17323 - 17 333.
Proc. Natl Acad. Sci. USA 72, 289-293.
Int. J. Radiat. Biol. 53, 749-765.





























