Embed Size (px)
Citation preview

IL-13 signal transduction in human monocytes:phosphorylation of receptor components, associationwith Jaks, and phosphorylation/activation of Stats
Biswajit Roy,* Ashish Bhattacharjee,* Bo Xu,* Dwayne Ford,† Abby L. Maizel,†
and Martha K. Cathcart**Department of Cell Biology, Lerner Research Institute, Cleveland Clinic Foundation, Ohio; and †Roger WilliamsMedical Center, Boston University, School of Medicine, Massachusetts
Abstract: Interleukin (IL)-13 regulates monocytefunction and is a potent stimulator of 15-lipoxy-genase expression. In different cell types, the func-tional IL-13 receptor complex can be comprised ofvariable protein components and has not beenthoroughly examined in human monocytes. Here,we identify the receptor components and upstreamsignaling events initiated by IL-13 in primary hu-man blood monocytes. The expression, phosphor-ylation and associated Jak kinases of the known,variable receptor components, IL-4R�, IL-2R�c,IL-13R�1 and IL-13R�2, were examined. We de-termined that IL-4R� and IL13R�1 are phosphor-ylated upon exposure to IL-13. Although IL-2R�cis also expressed, it is not phosphorylated uponexposure to IL-13. Evaluation of the presence ofIL-13R�2 failed to reveal significant mRNA orprotein expression. Earlier, our laboratory showedthat IL-13 induced the phosphorylation of Jak2and Tyk2 in monocytes and that expression of bothJaks was essential for downstream signaling by IL-13. Here, we report that Jak2 is associated withIL-4R�, and Tyk2 is associated with the IL-13R�1component of the IL-13 receptor complex. Addi-tionally, Stat proteins 1�, 3, 5A, 5B, and 6 arephosphorylated in response to IL-13. Further, thenuclear translocation and DNA binding of each ofthese Stats were induced by IL-13. These datarepresent the first complete report of the func-tional IL-13 receptor complex and early signalingevents in human monocytes. This information iscritical for understanding the IL-13 response ofmonocytes in inflammation. J. Leukoc. Biol. 72:580–589; 2002.
Key Words: human macrophages � cytokine receptors � cytokines� inflammation � monocytes
INTRODUCTION
Interleukin (IL)-13 is a pleiotropic immune regulatory cytokinethat shares structural and biological properties with IL-4 [1].IL-13 is known to promote growth of preactivated B lymphocytes
[2], induce germ line ε transcripts, and direct naive B lymphocytesto switch to immunoglobulin IgE and IgG4 synthesis [3]. It alsoinduces expression of the low-affinity receptor for IgE/CD23 andup-regulates class II major histocompatibility complex expressionon B lymphocytes and monocytes [4, 5]. In monocytes, IL-13down-regulates surface expression of the Fc receptor for IgG [5]and inhibits synthesis of inflammatory cytokines including tumornecrosis factor-�, IL-1�, IL-6, and IL-8 [6, 7]. Moreover, itsuppresses synthesis of IL-12, a critical cytokine for differentia-tion of uncommitted T cells toward the T helper cell type 1phenotype [5, 8]. Recent reports highlight the central contributionof IL-13 to experimental allergic asthma [9, 10]. The role of thiscytokine in inducing monocyte 15-lipoxygenase (15-LO), a lipid-peroxidating enzyme of potential interest in atherosclerosis,asthma, and inflammation in general, is also well documented [11,12]. We recently reported that two Jak kinases involved in theinitial signaling response to IL-13 are required for induction of15-LO in human monocytes [11], yet the monocyte receptor com-plex components and association of the Jaks with these compo-nents remained to be defined.
Receptor complexes for many cytokines have been shown toshare components. The composition of the IL-13 receptor com-plex has been shown to vary between and among differing celltypes, but IL-4 and IL-13 receptor complexes can use theIL-4R� protein (140 kDa). The IL-13 receptor complex hasalso been reported to putatively share a component that is usedby IL-4 as well as several other cytokines, the IL-2R�c, afinding that remains controversial [13]. Two other human IL-13receptor components have recently been cloned. One of these,referred to here as IL-13R�1, was cloned by three separategroups [14–16]. The other, herein referred to as IL-13R�2,was cloned by Caput et al. [17]. Both components are 55–70kDa and bind IL-13 with different affinities. IL-13R�1 initiallybinds IL-13 with subsequent recruitment of the IL-4R� (140-kDa glycoprotein) to efficiently transduce a signal [14, 18],whereas the IL-13R�2 can bind IL-13 in the absence ofIL-4R�, but its role in IL-13 signaling is still unclear [17].Earlier, Doucet et al. [19] had shown that the pattern of
Correspondence: Dr. Martha K. Cathcart, Department of Cell Biology/NC10,Lerner Research Institute, Cleveland Clinic Foundation, 9500 Euclid Avenue,Cleveland, OH 44195. E-mail: [email protected]
Received March 6, 2002; revised April 8, 2002; accepted May 1, 2002.
580 Journal of Leukocyte Biology Volume 72, September 2002 http://www.jleukbio.org

expression of IL-13R�2 varied from cell line to cell line withinthe human lung fibroblast cell lineages. Although CCL202 andFPA cell lines expressed IL-13R�2, the other cell line, ICIG7,had no detectable expression of IL-13R�2 by reverse tran-scriptase-polymerase chain reaction (RT-PCR) or on immuno-blots. Studies such as these implicate cell lineage-specificvariability in the IL-13 receptor constituents.
A series of cytokines and growth factors are known to triggeractivation of members of the Jak family of kinases that asso-ciate with receptor components. In recently published studies,we found that among the Jaks, only Jak2 and Tyk2 werephosphorylated in response to IL-13 in human monocytes [11].Other studies revealed the inhibition of expression of either ofthese kinases blocked the IL-13 induction of the 15-LO andwere therefore critical components for transducing the cytokinesignal. The association of these kinases with IL-13 receptorcomponents was not investigated in these earlier studies; how-ever, Jak1 was shown to associate with IL-4R� in response toIL-4 in human monocytes [20], and another report showed thatJak2 associated with IL-4R� [21].
Members of the Jak family of kinases can mediate thephosphorylation of Stat proteins on a single tyrosine leading toStat translocation and DNA binding [22–28]. So far, six mem-bers of the Stat family have been identified and are referred toas Stats1–6. Several of the Stats can be expressed as alterna-tively spliced isoforms {e.g., 91-kDa form (Stat1�) and 84-kDaform (Stat1�); ref [29]}.
Stat proteins contain SH2 domains and dimerize after phos-phorylation [30]. This enables the proteins to be efficiently trans-ported to the nucleus and bind DNA [31]. DNA binding mayinvolve interactions with other proteins, such as p48, which formspart of the interferon (IFN)-stimulated gene factor complex bind-ing to the IFN-stimulated response element [32]. The ability ofindividual cytokine receptors to activate overlapping but distinctsets of homo- and heterodimerizing Stat proteins contributes tosignal specificity. For example, interferon-� activates Stats1, 2,and 3 and exerts antiviral- and growth-inhibitory effects in thetumorigenic cell line Daudi [33], and prolactin activates Stats1, 3,and 5 in mammary epithelial cells and induces milk protein geneexpression [34, 35].
In this study, we report that IL-13 induces the phosphory-lation of two receptor components, IL-13R�1 and IL-4R�. Incontrast, neither IL-13R�2 nor IL-2R�c appears to participatein the signaling process. Based on our earlier reports that Jak2and Tyk2 are phosphorylated in response to IL-13 in thesecells, we also investigated the association of Jak2 and Tyk2with the receptor constituents and found that Jak2 associateswith IL-4R�, and Tyk2 associates with IL-13R�1. Further, wefound that Stat1�, Stat3, Stat5A/Stat5B, and Stat6 are tyrosine-phosphorylated and activated in response to IL-13. Thesefindings delineate an IL-13-driven, selective signaling pathwayfrom the receptor to the nucleus in human monocytes.
MATERIALS AND METHODS
Materials
Recombinant human IL-13 was purchased from Upstate Biotechnology (LakePlacid, NY). Rabbit antisera against Jak1, Jak2, Tyk2, and Jak3 were pur-
chased from Upstate Biotechnology or Santa Cruz Biotechnology (Santa Cruz,CA). Each one of the Jak/Tyk antibodies was essentially noncrossreactive withthe other members of the Jak family of kinases and recognized correspondingantigens under native (in 1% Triton X-100 extracts) as well as denaturingconditions.
Antibodies to IL-4R� were purchased from Santa Cruz Biotechnology andR&D Systems (Minneapolis, MN). Affinity-purified antisera against deducedpeptides from the amino-terminal or carboxy-terminal regions of IL-13R�1were prepared as previously described [18] with the amino-terminal antibodydemonstrating higher specificity in Western blotting. Monoclonal rat anti-human IL-2R�c was purchased from BD Pharmingen (San Diego, CA) and wasused for immunoprecipitation. Rabbit anti-human IL-2R�c (C-20) used forWestern blots was obtained from Santa Cruz Biotechnology. Antisera againstthe IL-13R�2 were produced in collaboration with Zymed, Inc. (San Francisco,CA). This heterotypic antisera, against the carboxy-terminal amino acids364–375 of the deduced protein sequence [17], was purified by affinitychromatography using the peptide attached to a sulfolink column (Pierce,Rockford, IL) as described [18]. The purified antisera against the IL-13R�2were capable of recognizing that protein produced subsequent to Superfect™(Qiagen, Valencia, CA) mediated transfection of the receptor construct [17] inthe expression system pcDNA 3.1 (Invitrogen, Carlsbad, CA) into COS-7 cells.In addition, the antisera effectively immunoprecipitated radiolabeled 125I-IL-13 [36] subsequent to binding and cross-linking the ligand to cells bearingthe receptor.
Rabbit antisera against Stat proteins 1–5 were purchased from TransductionLaboratories (Lexington, KY; Stat1–4) as well as Santa Cruz Biotechnology(Stats2, 4, and 5). Antibodies to Stat5A and Stat5B were obtained from UpstateBiotechnology. Antibodies to Stat6 and phospho-Stat6 were purchased fromBD Pharmingen and Cell Signaling Technology Inc. (Beverly, MA), respec-tively. Antisera raised against two peptides or amino acids 1–178 or 592–731of the Stat1 protein were also used (Transduction Laboratories). Each of theStat antibodies used for these studies was essentially noncrossreactive withother Stat molecules and was good for detecting proteins on Western blots aswell as immunoprecipitating the respective antigens. Antiphosphotyrosine-Statantibodies raised against Stat1 (Y701), Stat3 (Y705), and Stat5A/B(Y694,Y699) were purchased from Upstate Biotechnology and were used todetect tyrosine-phosphorylated Stat proteins on Western blots.
For general detection of tyrosine phosphorylated proteins on immunoblots,PY-99 (Transduction Laboratories) or a mixture (1:1) of phosphotyrosineantibodies PY-20 (Santa Cruz Biotechnology) and 4G-10 (Upstate Biotechnol-ogy) was used at dilutions of 1:1000.
Isolation of human monocytes
Human peripheral blood monocytes were isolated from heparinized wholeblood by sequential centrifugation over a Ficoll-Paque solution and adherenceto serum-coated tissue culture flasks as described previously [37]. Nonadher-ent cells were removed from the flasks by subsequent washes using Dulbecco’smodified Eagle’s medium (DMEM; Life Technologies, Inc., Gaithersburg, MD)supplemented with 10% bovine calf serum (BCS; Hyclone, Logan, UT). Ad-herent cells were released from the flask using 5 mM ethylenediaminetetraac-etate and were plated after washing in tissue-culture plates (Costar, Cam-bridge, MA). The isolated cell preparations had typically more than 95%monocytes, were maintained in DMEM containing 10% BCS at 37°C in thepresence of 10% CO2, and were used immediately for experiments.
Immunoprecipitation and Western blotting
Freshly isolated monocytes were counted and plated in six-well or 10-cmplates and were allowed to adhere for 2 h. The cells were then pretreated for15 min with addition of sodium orthovanadate solution (100 �M) followed bytreatment with or without IL-13 (250–500 pM) or IL-4 (670 pM) for 10, 15, or30 min (as indicated). Orthovanadate does not enter the cells [38, 39].Extensive studies in our laboratory have shown that treatment of humanmonocytes with 100 �M sodium orthovanadate does not induce the Jak or Statpathway as is observed with cell-permeable pervanadate and instead preservesphosphorylation signals of the phosphoproteins upon Western analysis, likelyby reducing phosphatase activity immediately upon lysis. In these experi-ments, this treatment did not change the results qualitatively but merelyimproved the preservation of the phosphorylation. For receptor studies, post-nuclear lysates were prepared using a lysis buffer of 1% Triton X-100, 150 mM
Roy et al. IL-13 signal transduction in human monocytes 581

NaCl, 50 mM NaF, 30 mM � glycerophosphate, 0.5 mM phosphoserine, 0.5mM phosphotyrosine, 1.0 mM phosphothreonine, 1.5 mM p-nitrophenylphos-phate, 50 mM Tris, pH 7.4, 1 mM sodium orthovanadate, 500 �M phenyl-methylsulfonyl fluoride, and protease inhibitor cocktail (Sigma Chemical Co.,St. Louis, MO). The cells were kept on ice for 30 min and centrifuged at 9300g for 15 min at 4°C, the supernatant was collected, and protein concentrationwas determined. The lysates were run on sodium dodecyl sulfate-polyacryl-amide gel electrophoresis (SDS-PAGE) gels and transferred to polyvinylidenedifluoride (PVDF) membranes as previously described [11]. For Stat experi-ments, whole cell extracts and nuclear extracts were used as noted andprepared according to previously published protocols [40, 41] with the excep-tion of 0.5 M salt, which was used in the whole cell lysis buffer. The blockerused for direct Westerns with antiphospho Stat antibodies was 5% milk inphosphate-buffered saline (PBS) with 0.1% Tween 20; otherwise, it was 5%bovine serum albumin in PBS/Tween 20.
For immunoprecipitation experiments, the lysates (0.5–1.0 mg/ml, usually0.5–2.0 mg total lysates) were incubated with immunoprecipitating antibodies(4–6 �g/ml as noted) for 1–2 h at 4°C with constant rotation. Immunecomplexes were collected using prewashed Sepharose-protein A or Sepharose-protein G beads (20–50 �l packed bead vol per ml extract). The beads werewashed three times with lysis buffer. The immune complexes were released byboiling the beads in SDS sample buffer and then analyzed by SDS-PAGEfollowed by electrophoretic transfer to PVDF membranes (Bio-Rad, Hercules,CA). The membranes were blocked and subsequently probed with specificantibodies and developed using enhanced chemiluminescence (ECL; Pierce).For immunoprecipitation of IL-13R�1, the antisera against the aminoterminalpeptide were used. Sodium pyrophosphate, p-nitrophenyl phosphate, phospho-serine, phosphotyrosine, phosphothreonine, and �-glycerophosphate were notincluded in the lysis buffer for phosphoprotein immunoprecipitation experi-ments.
In numerous experiments, immunoblots were stripped and reprobed toassess equal loading and/or equal immunoprecipitation. In these instances, theblots were incubated at 50°C for 30 min in a stripping buffer containing 62.5mM Tris-HCl, pH 6.7, 2% SDS, and 100 mM �-mercaptoethanol. The blotswere then washed three to four times for 15 min each with PBS-Tween 20(0.1% v/v) and were then reprobed with a different primary antibody anddeveloped using ECL.
Electrophoretic mobility shift assay (EMSA)
To assess the DNA binding activity of Stat proteins upon IL-13 treatment,EMSA was performed using nuclear extracts from human monocytes andspecific Stat probes. Briefly, nuclear proteins from monocytes with or withoutIL-13 treatment were extracted by the method described above. The double-stranded Stat1-, Stat3-, and Stat5-specific probes were obtained from SantaCruz, and the SBE1 probe (5�-GCTCTTCTTCCCAGGAACTCAATG-3�) wasused for Stat6. The Stat probes were labeled with 32P using T4-polynucleotidekinase (Promega, Madison, WI) and were incubated with 3 �g nuclear proteinsat room temperature for 20 min. The protein-DNA complexes were resolved on5% denaturing polyacrylamide gels. After drying, the gels were exposed toX-ray films at �80°C.
Determination of mRNA for IL-13R�2
The presence of mRNA for the IL-13R�2 was performed as previously de-scribed using a nested protocol [18]. The first-stage reaction results in a1088-bp product, and the internal primers for the second reaction results inamplification of a 437-bp product.
RESULTS
Phosphorylation of receptor components in IL-13-treated human monocytes
To determine whether IL-4R� was phosphorylated on tyrosinein response to IL-13, its phosphorylation status was analyzedusing antiphosphotyrosine immunoprecipitation (using PY-99)followed by detection with an IL-4R� antibody on a Westernblot (Fig. 1A). PY-99-immunoprecipitated IL-4R� was de-
tected in the immunoprecipitate from untreated monocyte celllysates. This result was confirmed by performing experimentsusing the antibody to IL4-R� for immunoprecipitation and thenprobing with the antibody to phosphotyrosine. IL-13 caused asubstantial increase in tyrosine phosphorylation of IL4-R� inthese experiments (Fig. 1B, upper panel). After stripping, thisblot was reprobed with antibody to IL4-R�, and equal loadingwas evident in the two lanes (Fig. 1B, lower panel). A similarresult was obtained by probing a blot of the PY99 immunopre-cipitate with antibody to IL4-R� derived from another source(data not shown).
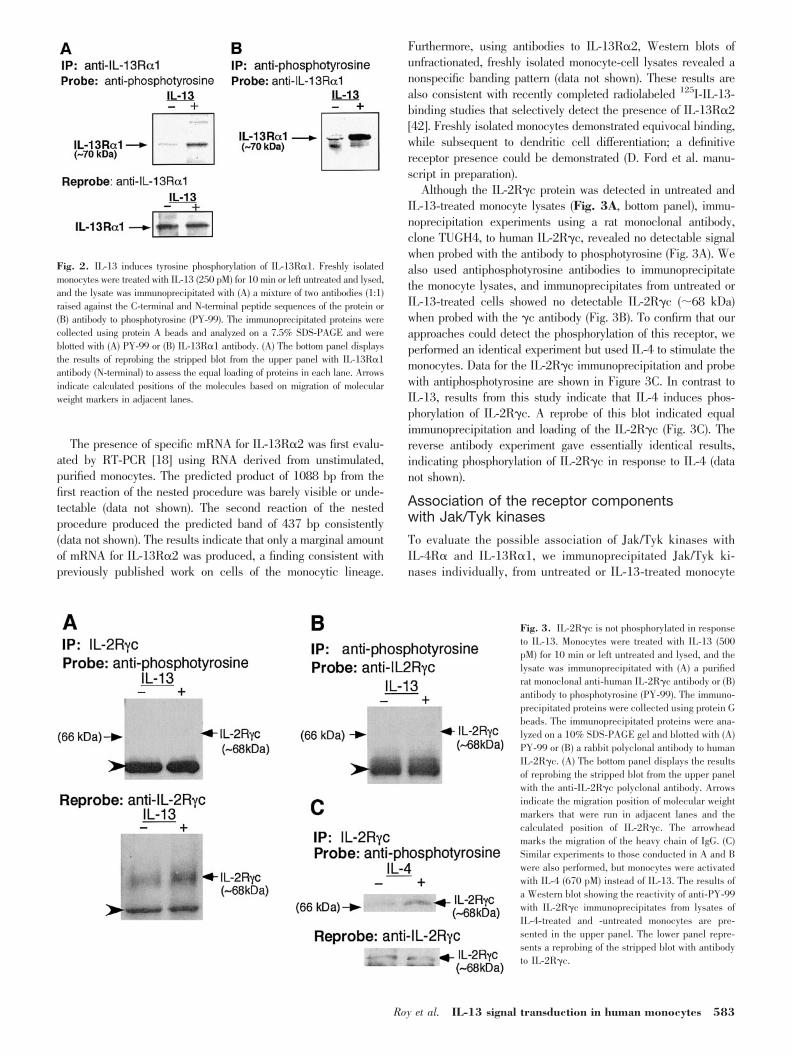
The phosphorylation status of IL-13R�1 and IL-13R�2receptor components was investigated next. Two different an-tibodies against N-terminal or C-terminal fragments of IL-13R�1 were used in our studies, as described in Materials andMethods [18]. Blots of SDS-PAGE gels of IL-13R�1 immuno-precipitates, derived from 1 mg each of untreated and IL-13-treated monocyte cell lysates, were probed with antiphospho-tyrosine (PY-99). The results indicate increased (�3.4-fold)phosphorylation of IL-13R�1 in IL-13-treated monocytes witha low but detectable basal level of phosphorylated protein inthe untreated cells (Fig. 2). The blot was stripped and re-probed with antibody to IL-13R�1 to evaluate the loading inthe two lanes, and equal loading was observed (Fig. 2A, bottompanel). This result was confirmed by detecting phosphorylatedIL-13R�1 by probing a blot of an SDS gel of the PY-99immunoprecipitate with the antibody raised against the N-terminal sequence of the receptor component (Fig. 2B). Theresults thus indicate that IL-13R�1 is an active constituent ofthe IL-13 signaling complex.
Fig. 1. IL-13 induces tyrosine phosphorylation of IL-4R�. Human bloodmonocytes were treated with IL-13 (250 pM) for 10 min or left untreated asindicated. (A) The cells were lysed; the cleared cell lysates were used forimmunoprecipitation with the phosphotyrosine antibody, PY-99. Immune com-plexes were collected using protein A beads and analyzed by immunoblottingwith antibody to IL-4R�. (B) Cell lysates were immunoprecipitated withantibody to IL-4R�, and immune complexes were analyzed by Western blot-ting with antibody to phosphotyrosine. Arrows indicate the position of IL-4R�as calculated from molecular weight markers in adjacent lanes. The lowerpanel displays the results of reprobing the stripped blot from the upper panelwith IL-4R� antibody to assess equal immunoprecipitation and loading ofprotein in each lane.
582 Journal of Leukocyte Biology Volume 72, September 2002 http://www.jleukbio.org
The presence of specific mRNA for IL-13R�2 was first evalu-ated by RT-PCR [18] using RNA derived from unstimulated,purified monocytes. The predicted product of 1088 bp from thefirst reaction of the nested procedure was barely visible or unde-tectable (data not shown). The second reaction of the nestedprocedure produced the predicted band of 437 bp consistently(data not shown). The results indicate that only a marginal amountof mRNA for IL-13R�2 was produced, a finding consistent withpreviously published work on cells of the monocytic lineage.
Furthermore, using antibodies to IL-13R�2, Western blots ofunfractionated, freshly isolated monocyte-cell lysates revealed anonspecific banding pattern (data not shown). These results arealso consistent with recently completed radiolabeled 125I-IL-13-binding studies that selectively detect the presence of IL-13R�2[42]. Freshly isolated monocytes demonstrated equivocal binding,while subsequent to dendritic cell differentiation; a definitivereceptor presence could be demonstrated (D. Ford et al. manu-script in preparation).
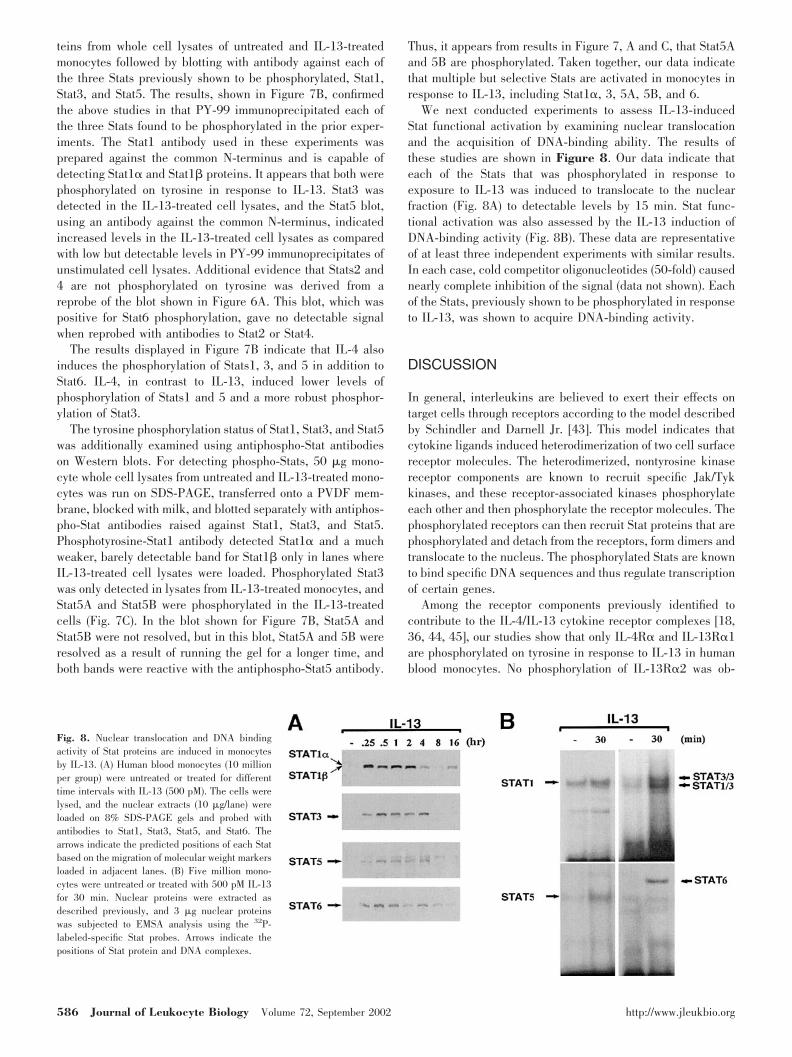
Although the IL-2R�c protein was detected in untreated andIL-13-treated monocyte lysates (Fig. 3A, bottom panel), immu-noprecipitation experiments using a rat monoclonal antibody,clone TUGH4, to human IL-2R�c, revealed no detectable signalwhen probed with the antibody to phosphotyrosine (Fig. 3A). Wealso used antiphosphotyrosine antibodies to immunoprecipitatethe monocyte lysates, and immunoprecipitates from untreated orIL-13-treated cells showed no detectable IL-2R�c (�68 kDa)when probed with the �c antibody (Fig. 3B). To confirm that ourapproaches could detect the phosphorylation of this receptor, weperformed an identical experiment but used IL-4 to stimulate themonocytes. Data for the IL-2R�c immunoprecipitation and probewith antiphosphotyrosine are shown in Figure 3C. In contrast toIL-13, results from this study indicate that IL-4 induces phos-phorylation of IL-2R�c. A reprobe of this blot indicated equalimmunoprecipitation and loading of the IL-2R�c (Fig. 3C). Thereverse antibody experiment gave essentially identical results,indicating phosphorylation of IL-2R�c in response to IL-4 (datanot shown).
Association of the receptor componentswith Jak/Tyk kinases
To evaluate the possible association of Jak/Tyk kinases withIL-4R� and IL-13R�1, we immunoprecipitated Jak/Tyk ki-nases individually, from untreated or IL-13-treated monocyte
Fig. 2. IL-13 induces tyrosine phosphorylation of IL-13R�1. Freshly isolatedmonocytes were treated with IL-13 (250 pM) for 10 min or left untreated and lysed,and the lysate was immunoprecipitated with (A) a mixture of two antibodies (1:1)raised against the C-terminal and N-terminal peptide sequences of the protein or(B) antibody to phosphotyrosine (PY-99). The immunoprecipitated proteins werecollected using protein A beads and analyzed on a 7.5% SDS-PAGE and wereblotted with (A) PY-99 or (B) IL-13R�1 antibody. (A) The bottom panel displaysthe results of reprobing the stripped blot from the upper panel with IL-13R�1antibody (N-terminal) to assess the equal loading of proteins in each lane. Arrowsindicate calculated positions of the molecules based on migration of molecularweight markers in adjacent lanes.
Fig. 3. IL-2R�c is not phosphorylated in responseto IL-13. Monocytes were treated with IL-13 (500pM) for 10 min or left untreated and lysed, and thelysate was immunoprecipitated with (A) a purifiedrat monoclonal anti-human IL-2R�c antibody or (B)antibody to phosphotyrosine (PY-99). The immuno-precipitated proteins were collected using protein Gbeads. The immunoprecipitated proteins were ana-lyzed on a 10% SDS-PAGE gel and blotted with (A)PY-99 or (B) a rabbit polyclonal antibody to humanIL-2R�c. (A) The bottom panel displays the resultsof reprobing the stripped blot from the upper panelwith the anti-IL-2R�c polyclonal antibody. Arrowsindicate the migration position of molecular weightmarkers that were run in adjacent lanes and thecalculated position of IL-2R�c. The arrowheadmarks the migration of the heavy chain of IgG. (C)Similar experiments to those conducted in A and Bwere also performed, but monocytes were activatedwith IL-4 (670 pM) instead of IL-13. The results ofa Western blot showing the reactivity of anti-PY-99with IL-2R�c immunoprecipitates from lysates ofIL-4-treated and -untreated monocytes are pre-sented in the upper panel. The lower panel repre-sents a reprobing of the stripped blot with antibodyto IL-2R�c.
Roy et al. IL-13 signal transduction in human monocytes 583

cell lysates and analyzed the SDS-PAGE blots with antibodiesto the receptor components. Results of these experiments arepresented in Figures 4 and 5. Antibodies to Jak1 and Jak2immunoprecipitated IL-4R�, and Jak3 and Tyk2 did not (Fig.4A). The data indicate that the recruitment of Jak1 to IL-4R�is IL-13-dependent, Jak2 is endogenously associated with thereceptor, and association increases upon treatment with IL-13.Reversing the antibodies in immunoprecipitator and Westernconfirmed the coassociation of Jak2 with IL-4R� (Fig. 4B).Reprobing the blots with antibodies corresponding to thoseused for immunoprecipitation confirmed that equal amounts ofspecific proteins were immunoprecipitated from the lysates(Fig. 4, A and B, lower panels).
Similar experiments, where Jak/Tyk kinases were immuno-precipitated from untreated or IL-13-treated monocyte celllysates and were blotted with antibody to IL-13R�1, showedthat predominantly Tyk2 and not other Jaks distinctly coim-munoprecipitated IL-13R�1 (Fig. 5A). Similar findings weredemonstrated when the IP and Western antibodies were re-versed as shown in Figure 5B. Results indicated that theassociation between Tyk2 and IL-13R�1 existed in untreatedcells and was modestly enhanced (25–40%, n�3) by IL-13treatment. Reprobing the same blots with the antibodies usedfor immunoprecipitation confirmed equal loading of the immu-noprecipitated proteins (Fig. 5, A and B, lower panels).
Earlier reports from our lab [11] showed that both Jak2 andTyk2, but not Jak1 and Jak3, were phosphorylated on tyrosinein IL-13-treated human blood monocytes. The association ex-periments above thus show that although Jak2 associates withIL-4R�, Tyk2 associates with the other constituent of the IL-13receptor complex, IL-13R�1.
Tyrosine phosphorylation and activation of Stats
Because the Stat proteins are early and important substrates ofphosphorylated and activated Jak/Tyk kinases, we next studiedwhich of the six known Stats were involved in the IL-13signaling pathway. Previously, Stat6 has been reported asbeing phosphorylated on tyrosine in response to IL-13. Weconducted experiments to test this in our monocyte culturesystem and found that Stat6 does indeed become phosphory-lated. Data from a representative experiment are shown inFigure 6 where Stat6 was detected in PY-99 immunoprecipi-tates in IL-13-treated monocyte-nuclear lysates (Fig. 6A) or inwhole cell lysates when directly probed with antiphospho-Stat6antibody (Fig. 6B). Equal loading of protein is observed in thereprobe of the upper blot in Figure 6B with antibody to Stat6.
Phosphorylation of other Stat family members in monocyteshas remained unexplored. We next conducted experiments toevaluate whether any other Stat proteins were phosphorylatedin monocytes as a result of IL-13 stimulation. The remaining
Fig. 4. Jak1 and Jak2 associate with IL-4R�.Monocytes (20 million cells per group) were treatedwith IL-13 or left untreated. (A) Both groups werelysed and immunoprecipitated with antibodies spe-cific for Jak1, Jak2, Tyk2, or Jak3. The immuno-precipitated proteins were absorbed to protein Abeads, extracted, and analyzed on a 7.5% SDS-PAGE blot and probed with an antibody to IL-4R�.The arrow indicates the estimated position of theIL-4R� protein. The bottom panel displays the blotfrom subsequent reprobing, after stripping, of theindividual two-lane strips, using antibodies againstcorresponding Jak/Tyk kinases. Near-equal immu-noprecipitation of proteins was observed. (B) Mono-cytes were treated as above, but antibody to IL-4R�was used for immunoprecipitation, and the blot ofthe SDS-PAGE was then probed with antibody to Jak2. The bottom panel shows the results of reprobing this stripped immunoblot with antibody to IL-4R� toevaluate equal immunoprecipitation and loading.
Fig. 5. Tyk2 associates with IL-13R�1. (A)Monocytes were treated and immunoprecipitatedas described in the legend to Figure 4. The blotwas probed with an antibody against the N-ter-minal peptide sequence of the IL-13R�1 protein.The arrow indicates the calculated, expected po-sition of IL-13R�1. The bottom panel shows theresults of reprobing, after stripping, the individ-ual strips with corresponding Jak/Tyk antibodiesto assess equal immunoprecipitation and loadingof proteins in each lane. (B) IL-13R�1 was im-munoprecipitated from monocyte lysates (20 mil-lion per group), untreated or treated with IL-13,and analyzed on a 7.5% SDS-PAGE. The blot wasprobed with antibody to Tyk2, and the arrowindicates the calculated, predicted position ofTyk2. The bottom panel shows the same blot thatwas stripped and reprobed with IL-13R�1 antibody to assess equal immunoprecipitation and loading of proteins in each lane.
584 Journal of Leukocyte Biology Volume 72, September 2002 http://www.jleukbio.org

Stat proteins (Stats1–5) were individually immunoprecipitatedfrom untreated and IL-13-treated monocyte lysates. The im-munoprecipitates were then evaluated in Western blots. Theblot was probed with phosphotyrosine antibody (PY-99) todetect IL-13-induced changes in the phosphotyrosine contentof the immunoprecipitated Stat proteins. We observed thatalthough Stat1, Stat3, Stat5A, and Stat5B were phosphorylatedon tyrosine in response to IL-13, other Stats, Stat2 and Stat4,
were not detected by the PY-99 antibody (Fig. 7A). In eachcase, the blots were individually reprobed with correspondingStat antibodies to ensure adequate and equal immunoprecipi-tation and loading of each Stat protein. The antibody to Stat1,in Figure 7A, was against the C-terminus of the protein andtypically detects mostly Stat1� (91 kDa).
To confirm these data, we also used antiphosphotyrosine(PY-99) to immunoprecipitate tyrosine-phosphorylated pro-
Fig. 6. IL-13 induces tyrosine phosphorylation of Stat6protein in monocytes. (A) Monocytes (20 million per group)were untreated or treated with IL-13 (500 pM) or IL-4 (670pM) for 30 min and were lysed, and the nuclear extractswere immunoprecipitated with the phosphotyrosine antibodyPY-99. The immunoprecipitated proteins were collectedusing protein G beads and were run on 8% SDS-PAGE geland immunoblotted with Stat6 antibody. Jurkat cell extractwas run in an adjacent lane of the same gel as a positivecontrol, and the immunoreactive band migrated just above97 kDa, identically with the monocyte band (data notshown). (B) Human blood monocytes (5106) were un-treated or treated with IL-13 (500 pM) and IL-4 (670 pM) for30 min. The cells were lysed, and the whole cell extracts (75�g/lane) were loaded on an 8% SDS-PAGE gel and immu-noblotted with antiphospho Stat6 (Tyr 641) antibody. Thebottom panel shows the same blot that was reprobed with
Stat6 antibody to evaluate equal loading of proteins.
Fig. 7. IL-13 induces tyrosine phosphorylation of Stat pro-teins in monocytes. (A) Monocytes (30 million per group)were untreated or treated with IL-13 for 15 min, lysed, andimmunoprecipitated with antibodies specific for Stat1, Stat2,Stat3, Stat4, Stat5A, and Stat5B. The immunoprecipitatedproteins were collected on protein A beads and separated ona 7.5% SDS-PAGE, transferred onto a PVDF membrane, andprobed with PY-99. The bottom panel shows the stripped andreprobed individual sections of the membrane blot with cor-responding antibodies, demonstrating nearly equal immuno-precipitation and loading of proteins in each lane. (B) Mono-cytes (20 million per group) were untreated or treated withIL-13 (500 pM) or IL-4 (670 pM) for 30 min and were lysed,and the nuclear extracts were immunoprecipitated with thephosphotyrosine antibody PY-99. The immunoprecipitatedproteins were collected using protein G beads. Three differ-ent blots were prepared by running the samples on 8%SDS-PAGE gels, and the blots were probed individually withantibodies to Stat1, Stat3, and Stat5. Arrows indicate pre-dicted positions of Stat proteins, based on the migration ofmolecular weight markers. In the Stat3 blot, lysate from A431cells was run in an adjacent lane of the same gel as a positivecontrol and as an indicator of the migration of Stat3 (data notshown). (C) Monocyte lysates were prepared from untreated orIL-13-treated cells, and protein concentrations were deter-mined. Lysate protein (50 �g) from treated or untreatedmonocytes was loaded in adjacent lanes and probed sepa-rately using anti-phospho-Stat antibodies against Stat1, Stat3,and Stat5. The positions of the markers loaded in adjacentlanes are indicated.
Roy et al. IL-13 signal transduction in human monocytes 585
teins from whole cell lysates of untreated and IL-13-treatedmonocytes followed by blotting with antibody against each ofthe three Stats previously shown to be phosphorylated, Stat1,Stat3, and Stat5. The results, shown in Figure 7B, confirmedthe above studies in that PY-99 immunoprecipitated each ofthe three Stats found to be phosphorylated in the prior exper-iments. The Stat1 antibody used in these experiments wasprepared against the common N-terminus and is capable ofdetecting Stat1� and Stat1� proteins. It appears that both werephosphorylated on tyrosine in response to IL-13. Stat3 wasdetected in the IL-13-treated cell lysates, and the Stat5 blot,using an antibody against the common N-terminus, indicatedincreased levels in the IL-13-treated cell lysates as comparedwith low but detectable levels in PY-99 immunoprecipitates ofunstimulated cell lysates. Additional evidence that Stats2 and4 are not phosphorylated on tyrosine was derived from areprobe of the blot shown in Figure 6A. This blot, which waspositive for Stat6 phosphorylation, gave no detectable signalwhen reprobed with antibodies to Stat2 or Stat4.
The results displayed in Figure 7B indicate that IL-4 alsoinduces the phosphorylation of Stats1, 3, and 5 in addition toStat6. IL-4, in contrast to IL-13, induced lower levels ofphosphorylation of Stats1 and 5 and a more robust phosphor-ylation of Stat3.
The tyrosine phosphorylation status of Stat1, Stat3, and Stat5was additionally examined using antiphospho-Stat antibodieson Western blots. For detecting phospho-Stats, 50 �g mono-cyte whole cell lysates from untreated and IL-13-treated mono-cytes was run on SDS-PAGE, transferred onto a PVDF mem-brane, blocked with milk, and blotted separately with antiphos-pho-Stat antibodies raised against Stat1, Stat3, and Stat5.Phosphotyrosine-Stat1 antibody detected Stat1� and a muchweaker, barely detectable band for Stat1� only in lanes whereIL-13-treated cell lysates were loaded. Phosphorylated Stat3was only detected in lysates from IL-13-treated monocytes, andStat5A and Stat5B were phosphorylated in the IL-13-treatedcells (Fig. 7C). In the blot shown for Figure 7B, Stat5A andStat5B were not resolved, but in this blot, Stat5A and 5B wereresolved as a result of running the gel for a longer time, andboth bands were reactive with the antiphospho-Stat5 antibody.
Thus, it appears from results in Figure 7, A and C, that Stat5Aand 5B are phosphorylated. Taken together, our data indicatethat multiple but selective Stats are activated in monocytes inresponse to IL-13, including Stat1�, 3, 5A, 5B, and 6.
We next conducted experiments to assess IL-13-inducedStat functional activation by examining nuclear translocationand the acquisition of DNA-binding ability. The results ofthese studies are shown in Figure 8. Our data indicate thateach of the Stats that was phosphorylated in response toexposure to IL-13 was induced to translocate to the nuclearfraction (Fig. 8A) to detectable levels by 15 min. Stat func-tional activation was also assessed by the IL-13 induction ofDNA-binding activity (Fig. 8B). These data are representativeof at least three independent experiments with similar results.In each case, cold competitor oligonucleotides (50-fold) causednearly complete inhibition of the signal (data not shown). Eachof the Stats, previously shown to be phosphorylated in responseto IL-13, was shown to acquire DNA-binding activity.
DISCUSSION
In general, interleukins are believed to exert their effects ontarget cells through receptors according to the model describedby Schindler and Darnell Jr. [43]. This model indicates thatcytokine ligands induced heterodimerization of two cell surfacereceptor molecules. The heterodimerized, nontyrosine kinasereceptor components are known to recruit specific Jak/Tykkinases, and these receptor-associated kinases phosphorylateeach other and then phosphorylate the receptor molecules. Thephosphorylated receptors can then recruit Stat proteins that arephosphorylated and detach from the receptors, form dimers andtranslocate to the nucleus. The phosphorylated Stats are knownto bind specific DNA sequences and thus regulate transcriptionof certain genes.
Among the receptor components previously identified tocontribute to the IL-4/IL-13 cytokine receptor complexes [18,36, 44, 45], our studies show that only IL-4R� and IL-13R�1are phosphorylated on tyrosine in response to IL-13 in humanblood monocytes. No phosphorylation of IL-13R�2 was ob-
Fig. 8. Nuclear translocation and DNA bindingactivity of Stat proteins are induced in monocytesby IL-13. (A) Human blood monocytes (10 millionper group) were untreated or treated for differenttime intervals with IL-13 (500 pM). The cells werelysed, and the nuclear extracts (10 �g/lane) wereloaded on 8% SDS-PAGE gels and probed withantibodies to Stat1, Stat3, Stat5, and Stat6. Thearrows indicate the predicted positions of each Statbased on the migration of molecular weight markersloaded in adjacent lanes. (B) Five million mono-cytes were untreated or treated with 500 pM IL-13for 30 min. Nuclear proteins were extracted asdescribed previously, and 3 �g nuclear proteinswas subjected to EMSA analysis using the 32P-labeled-specific Stat probes. Arrows indicate thepositions of Stat protein and DNA complexes.
586 Journal of Leukocyte Biology Volume 72, September 2002 http://www.jleukbio.org

served. IL-13, in contrast to IL-4, did not induce the phos-phorylation of IL-2R�c. The lack of IL-2R�c involvement inthe IL-13 response is consistent with previous studies showinginvolvement of this receptor component in IL-4 responses butnot in IL-13 responses of a mast cell line [46], B lymphocytes[47], or lymphohematopoietic cells [48]. This finding might alsoexplain why Jak3 is involved in the IL-4 response in monocytesbut is not involved in the IL-13 signaling pathway, as Jak3 isknown to associate with IL-2R�c [49, 50]. IL-4 has also beenshown to be able to stimulate certain cells in the absence aswell as the presence of IL-2R�c. In contrast to signalingthrough Jak3/Stat6, in the absence of IL-2R�c, IL-4 activa-tion can still proceed but instead involves a Jak1/Stat6 path-way [51].
Four models of IL-13 receptor complex composition havebeen proposed by Murata et al. [44] to represent the differingcomponents between and among cell types and cell lines.Although Model I for the IL-13R involves IL-13R�1 andIL-13R�2 subunits, Model II includes IL-13R�1 and IL-4R�.Both of the other two models, III and IV, contain the IL-2R�cchain of the IL-2 receptor along with IL-13R�1 and IL-4R�.Therefore, we believe that Model II best represents the IL-13receptor complex in monocytes.
So far, mostly nontyrosine kinase receptors have been re-ported to transduce signals involving the Jak/Tyk kinases; theepidermal growth factor, platelet-derived growth factor, andfibroblast growth factor tyrosine kinase receptors are excep-tions in that they can use Jak kinases for signal transduction[27]. Physical association of Jak/Tyk kinases with profoundlydifferent nontyrosine kinase receptors has been shown, andassociation and phosphorylation of Jak/Tyk kinases on a par-ticular receptor molecule vary between cells. Interactions be-tween Jak kinases and receptor components can be constitutiveas well as inducible, as reported in numerous studies includingthose presented here.
Our studies demonstrate that Jak1 and Jak2 associated withIL-4R�. Cross-immunoprecipitation studies confirmed thisfinding. Significantly, although the association of Jak1 to IL-4R� was induced in response to ligand binding, Jak2 wasbound to IL-4R�, irrespective of the cytokine treatment. Fur-ther, we showed that Tyk2, another kinase phosphorylated inresponse to IL-13, bound to IL-13R�1 prior to exposure toIL-13, and association was modestly induced in response to theligand. Reports by others have indicated that upon exposure ofT lymphocytes to IL-4, IL-4R� can bind and phosphorylateJak1 [52]. At present, we cannot explain the IL-13-responsivebinding of Jak1 to IL-4R� and noninvolvement of Jak1 phos-phorylation in the IL-13 response. This appears to be anotherimportant difference between the IL-4 and IL-13 activationpathways. One might speculate that if only one Jak associateswith each molecule of IL-4R�, then endogenous levels of Jak1,differing in various cell types, may alter the downstream sig-naling by IL-13. Further evidence that Jak1 is not involved inthe IL-13 response of human monocyte, at least as it relates to15-LO expression, is derived from our recently publishedstudies showing that inhibition of Jak1 expression did notinterfere with IL-13-mediated 15-LO expression [11]. Othershave reported that IL-4 and IL-13 induce Jak2 phosphorylation[21, 53] and that Jak2 is constitutively associated with IL-4R�
in human colon carcinoma cells [21]. This finding is consistentwith the IL-13 signaling in monocytes as reported here. Sim-ilarly, IL-13 induction of Tyk2 phosphorylation was also ob-served in human colon carcinoma cells, similar to our results inhuman monocytes [11, 21]. Thus, some aspects of Jak/Tykassociation and phosphorylation are similar, and some aredifferent than observations in other cell types. One commonfinding from numerous labs in a variety of cell types, andconsistent with our findings, is that Jak3 is not phosphorylatedin response to IL-13 [21, 48, 54–56].
Stat proteins are activated in response to the binding of anumber of cytokines and growth factors to their specific recep-tors. The Stats are activated by phosphorylation of one partic-ular tyrosine residue located in the C-terminus [27]. Tyrosinephosphorylation allows Stat proteins to dimerize via binding ofthe SH2 domain of one Stat molecule to the phosphotyrosine ofanother activated Stat molecule. The dimerized Stat complex istransported to the nucleus where it binds DNA and thusregulates transcription of target genes [57]. Our observationsindicate that Stat1�, Stat3, Stat5A, Stat5B, and Stat6 aretyrosine-phosphorylated in IL-13-treated primary humanmonocytes. A recent report has also observed the phosphory-lation of Stats1 and 3 in response to IL-13 in human normaland tumor lung fibroblasts [58]. In normal B cells, studies byIzuhara et al. [47] have shown that IL-13-induced phosphory-lation of Stat6, in contrast to that induced by IL-4, is indepen-dent of Jak3 and IL-2R�c. This finding is consistent with theresults presented here with primary human monocytes.
The serine phosphorylation of Stats is known to be essentialfor optimal promotion of transcription but not for dimer forma-tion or nuclear translocation of the Stat complex [59, 60].Recent reports suggest that there is serine phosphorylation ofStats, Stat1� (Ser-727), Stat3 (Ser-727), Stat5A (Ser-725), andStat5B (Ser-730), although predictably, this phosphorylation ismediated by different kinases [61, 62]. We have recentlyshown that Stat1 and Stat3 are phosphoryated on Serine 727 inresponse to IL-13 (B. Roy, A. Bhattacharjee, B. Xu, and M. K.Cathcart, manuscript in preparation). Further studies are re-quired to understand exactly how tyrosine and serine-phospho-rylated Stat1 and Stat3 participate in the IL-13-mediated reg-ulation of gene transcription.
The formation of homodimers and heterodimers betweentyrosine-phosphorylated Stats as well as serine phosphorylationappears to be an important mechanism for providing specificityin gene induction. These additional, regulatory events poten-tially enable a limited group of transcription factors to orches-trate cell type-specific or cell stage-specific effects observed inresponse to many cytokines and growth factors. Recently, inStat6-transfected, transformed epithelial cells, Stat-6 has beenreported to regulate the IL-4-mediated induction of 15-LO[63]. Prior studies also suggested Stat6 regulation of IL-4-induced 15-LO enzymatic activity in murine macrophages [64].Our studies indicate Stat6 is not the only Stat with potential tomediate the IL-13 induction of 15-LO expression in primaryhuman monocytes. Further, our results suggest quantitativelydifferential Stat activation induced by IL-4 and IL-13. Thismay explain the differing cellular responses to these cytokines.
In summary, we report here the immediate, early compo-nents of the IL-13 signaling pathway in human monocytes. Our
Roy et al. IL-13 signal transduction in human monocytes 587

results, schematically presented in Figure 9, indicate thatIL-13 signals through IL-13R�1 and IL-4R�, two receptorcomponents characterized earlier in different cells types. Bothof these receptor components were phosphorylated upon expo-sure of monocytes to IL-13. Each was associated with membersof the Jak/Tyk kinase family, which are known to be phosphor-ylated, activated, and linked to expression of 15-LO expressionin response to IL-13 in these cells; i.e., Jak2 and Tyk2 wereshown to be associated with IL-4R� and IL-13R�1, respec-tively. Jak1 also associated with IL-4R� in response to IL-13exposure, but Jak1 was not phosphorylated. We have thereforedepicted Jak1 or Jak2 interacting with this component of theIL-13 receptor. Tanner et al. have reported that the peptidesequence of PXXPXP is an absolutely required receptor se-quence for binding to Jaks [65]. It is interesting to note thatIL-4R� has two of these sequences, 42 amino acids apart. It istherefore theoretically possible that IL-4R� could bind morethan one Jak; however, there are no data to support this. Ourstudies have also revealed the selective tyrosine phosphoryla-tion and activation of Stat proteins 1�, 3, 5A, 5B, and 6 inresponse to IL-13 in this system. The identification of theIL-13-induced signal transduction cascade in monocytes sug-gests novel regulation of IL-13 responses in these cells. Dif-ferences between IL-4 and IL-13 signal transduction in mono-cytes were identified, as well as differences between IL-13signal transduction in monocytes as compared with other celltypes. Further studies are needed to evaluate the roles of thesepathway components in regulating gene expression in responseto IL-13. Identification of the IL-13-induced signaling path-ways may provide novel targets for therapeutic intervention inthe IL-13-driven, inflammatory processes that appear to con-tinue to be of critical importance in allergic asthma and athero-genesis.
ACKNOWLEDGMENTS
B. R. and A. B. contributed equally to this work and are bothconsidered first authors. This work was supported by funding toM. K. C. from NIH-HL51068.
REFERENCES
1. Chaouchi, N., Wallon, C., Goujard, C., Tertian, G., Rudent, A., Caput, D.,Ferrera, P., Minty, A., Vazquez, A., Delfraissy, J. F. (1996) Interleukin-13inhibits interleukin-2-induced proliferation and protects chronic lympho-cytic leukemia B cells from in vitro apoptosis. Blood 87, 1022–1029.
2. McKenzie, G. J., Bancroft, A., Grencis, R. K., McKenzie, A. N. (1998) Adistinct role for interleukin-13 in Th2-cell-mediated immune responses.Curr. Biol. 8, 339–342.
3. Punnonen, J., de Vries, J. E. (1994) IL-13 induces proliferation, Ig isotypeswitching, and Ig synthesis by immature human fetal B cells. J. Immunol.152, 1094–1102.
4. Defrance, T., Carayon, P., Billian, G., Guillemot, J. C., Minty, A., Caput,D., Ferrara, P. (1994) Interleukin 13 is a B cell stimulating factor. J. Exp.Med. 179, 135–143.
5. de Waal Malefyt, R., Figdor, C. G., Huijbens, R., Mohan-Peterson, S.,Bennett, B., Culpepper, J., Dang, W., Zurawski, G., de Vries, J. E. (1993)Effects of IL-13 on phenotype, cytokine production, and cytotoxic functionof human monocytes. Comparison with IL-4 and modulation by IFN-gamma or IL-10. J. Immunol. 151, 6370–6381.
6. de Waal Malefyt, R., Abrams, J. S., Zurawski, S. M., Lecron, J. C.,Mohan-Peterson, S., Sanjanwala, B., Bennett, B., Silver, J., de Vries, J. E.,Yssel, H. (1995) Differential regulation of IL-13 and IL-4 production byhuman CD8 and CD4 Th0, Th1 and Th2 T cell clones and EBV-transformed B cells. Int. Immunol. 7, 1405–1416.
7. Minty, A., Ferrara, P., Caput, D. (1997) Interleukin-13 effects on activatedmonocytes lead to novel cytokine secretion profiles intermediate betweenthose induced by interleukin-10 and by interferon-gamma. Eur. CytokineNetw. 8, 189–201.
8. Hsieh, C. S., Macatonia, S. E., Tripp, C. S., Wolf, S. F., O’Garra, A.,Murphy, K. M. (1993) Development of TH1 CD4 T cells through IL-12produced by Listeria-induced macrophages. Science 260, 547–549.
9. Wills-Karp, M., Luyimbazi, J., Xu, X., Schofield, B., Neben, T. Y., Karp,C. L., Donaldson, D. D. (1998) Interleukin-13: central mediator of allergicasthma. Science 282, 2258–2261.
10. Grunig, G., Warnock, M., Wakil, A. E., Venkayya, R., Brombacher, F.,Rennick, D. M., Sheppard, D., Mohrs, M., Donaldson, D. D., Locksley,R. M., Corry, D. B. (1998) Requirement for IL-13 independently of IL-4in experimental asthma. Science 282, 2261–2263.
11. Roy, B., Cathcart, M. K. (1998) Induction of 15-lipoxygenase expressionby IL-13 requires tyrosine phosphorylation of Jak2 and Tyk2 in humanmonocytes. J. Biol. Chem. 273, 32023–32029.
12. Nassar, G. M., Morrow, J. D., Roberts II, L. J., Lakkis, F. G., Badr, K. F.(1994) Induction of 15-lipoxygenase by interleukin-13 in human bloodmonocytes. J. Biol. Chem. 269, 27631–27634.
13. Obiri, N. I., Murata, T., Debinski, W., Puri, R. K. (1997) Modulation ofinterleukin (IL)-13 binding and signaling by the gamma c chain of the IL-2receptor. J. Biol. Chem. 272, 20251–20258.
14. Aman, M. J., Tayebi, N., Obiri, N. I., Puri, R. K., Modi, W. S., Leonard,W. J. (1996) cDNA cloning and characterization of the human interleukin13 receptor alpha chain. J. Biol. Chem. 271, 29265–29270.
15. Gauchat, J. F., Schlagenhauf, E., Feng, N. P., Moser, R., Yamage, M.,Jeannin, P., Alouani, S., Elson, G., Notarangelo, L. D., Wells, T., Eugster,H. P., Bonnefoy, J. Y. (1997) A novel 4-kb interleukin-13 receptor alphamRNA expressed in human B, T, and endothelial cells encoding analternate type-II interleukin-4/interleukin-13 receptor. Eur. J. Immunol.27, 971–978.
16. Miloux, B., Laurent, P., Bonnin, O., Lupker, J., Caput, D., Vita, N.,Ferrara, P. (1997) Cloning of the human IL-13R alpha1 chain and recon-stitution with the IL-4R alpha of a functional IL-4/IL-13 receptor complex.FEBS Lett. 401, 163–166.
17. Caput, D., Laurent, P., Kaghad, M., Lelias, J. M., Lefort, S., Vita, N.,Ferrara, P. (1996) Cloning and characterization of a specific interleukin(IL)-13 binding protein structurally related to the IL-5 receptor alphachain. J. Biol. Chem. 271, 16921–16926.
18. Ogata, H., Ford, D., Kouttab, N., King, T. C., Vita, N., Minty, A.,Stoeckler, J., Morgan, D., Girasole, C., Morgan, J. W., Maizel, A. L. (1998)Regulation of interleukin-13 receptor constituents on mature human Blymphocytes. J. Biol. Chem. 273, 9864–9871.
19. Doucet, C., Brouty-Boye, D., Pottin-Clemenceau, C., Canonica, G. W.,Jasmin, C., Azzarone, B. (1998) Interleukin (IL)-4 and IL-13 act on humanlung fibroblasts. Implication in asthma. J. Clin. Investig. 101, 2129–2139.
20. Musso, T., Johnston, J. A., Linnekin, D., Varesio, L., Rowe, T. K., O’Shea,J. J., McVicar, D. W. (1995) Regulation of JAK3 expression in humanmonocytes: phosphorylation in response to interleukins 2, 4, and 7. J. Exp.Med. 181, 1425–1431.
21. Murata, T., Noguchi, P. D., Puri, R. K. (1996) IL-13 induces phosphory-lation and activation of JAK2 Janus kinase in human colon carcinoma cell
Fig. 9. Model of the predicted IL-13 receptor complex and upstream signal-transducing components.
588 Journal of Leukocyte Biology Volume 72, September 2002 http://www.jleukbio.org

lines: similarities between IL-4 and IL-13 signaling. J. Immunol. 156,2972–2978.
22. Silvennoinen, O., Ihle, J. N., Schlessinger, J., Levy, D. E. (1993) Inter-feron-induced nuclear signalling by Jak protein tyrosine kinases. Nature366, 583–585.
23. Stahl, N., Boulton, T. G., Farruggella, T., Ip, N. Y., Davis, S., Witthuhn,B. A., Quelle, F. W., Silvennoinen, O., Barbieri, G., Pellegrini, S., et al.(1994) Association and activation of Jak-Tyk kinases by CNTF-LIF-OSM-IL-6 beta receptor components. Science 263, 92–95.
24. Lutticken, C., Wegenka, U. M., Yuan, J., Buschmann, J., Schindler, C.,Ziemiecki, A., Harpur, A. G., Wilks, A. F., Yasukawa, K., Taga, T., et al.(1994) Association of transcription factor APRF and protein kinase Jak1with the interleukin-6 signal transducer gp130. Science 263, 89–92.
25. Igarashi, K., Garotta, G., Ozmen, L., Ziemiecki, A., Wilks, A. F., Harpur,A. G., Larner, A. C., Finbloom, D. S. (1994) Interferon-gamma inducestyrosine phosphorylation of interferon-gamma receptor and regulated as-sociation of protein tyrosine kinases, Jak1 and Jak2, with its receptor.J. Biol. Chem. 269, 14333–14336.
26. Shuai, K., Schindler, C., Prezioso, V. R., Darnell Jr., J. E. (1992) Acti-vation of transcription by IFN-gamma: tyrosine phosphorylation of a 91-kDDNA binding protein. Science 258, 1808–1812.
27. Shuai, K., Stark, G. R., Kerr, I. M., Darnell Jr., J. E. (1993) A singlephosphotyrosine residue of Stat91 required for gene activation by inter-feron-gamma. Science 261, 1744–1746.
28. Witthuhn, B. A., Quelle, F. W., Silvennoinen, O., Yi, T., Tang, B., Miura,O., Ihle, J. N. (1993) JAK2 associates with the erythropoietin receptor andis tyrosine phosphorylated and activated following stimulation with eryth-ropoietin. Cell 74, 227–236.
29. Schindler, C., Fu, X. Y., Improta, T., Aebersold, R., Darnell Jr., J. E.(1992) Proteins of transcription factor ISGF-3: one gene encodes the 91-and 84-kDa ISGF-3 proteins that are activated by interferon alpha. Proc.Natl. Acad. Sci. USA 89, 7836–7839.
30. Shuai, K., Horvath, C. M., Huang, L. H., Qureshi, S. A., Cowburn, D.,Darnell Jr., J. E. (1994) Interferon activation of the transcription factorStat91 involves dimerization through SH2-phosphotyrosyl peptide inter-actions. Cell 76, 821–828.
31. Zhong, Z., Wen, Z., Darnell Jr., J. E. (1994) Stat3: a STAT family memberactivated by tyrosine phosphorylation in response to epidermal growthfactor and interleukin-6. Science 264, 95–98.
32. Fu, X. Y., Kessler, D. S., Veals, S. A., Levy, D. E., Darnell Jr., J. E. (1990)ISGF3, the transcriptional activator induced by interferon alpha, consistsof multiple interacting polypeptide chains. Proc. Natl. Acad. Sci. USA 87,8555–8559.
33. Davis, E., Krishnan, K., Yan, H., Newcomb, E. W., Krolewski, J. J. (1996)A mutant form of p135tyk2, an interferon-alpha inducible tyrosine kinase,suppresses the transformed phenotype of Daudi cells. Leukemia 10,543–551.
34. Groner, B., Altiok, S., Meier, V. (1994) Hormonal regulation of transcrip-tion factor activity in mammary epithelial cells. Mol. Cell. Endocrinol.100, 109–114.
35. O’Shea, J. J. (1997) Jaks, STATs, cytokine signal transduction, andimmunoregulation: are we there yet? Immunity 7, 1–11.
36. Obiri, N. I., Leland, P., Murata, T., Debinski, W., Puri, R. K. (1997) TheIL-13 receptor structure differs on various cell types and may share morethan one component with IL-4 receptor. J. Immunol. 158, 756–764.
37. Cathcart, M. K., McNally, A. K., Morel, D. W., Chisolm III, G. M. (1989)Superoxide anion participation in human monocyte-mediated oxidation oflow-density lipoprotein and conversion of low-density lipoprotein to acytotoxin. J. Immunol. 142, 1963–1969.
38. Heffetz, D., Bushkin, I., Dror, R., Zick, Y. (1990) The insulinomimeticagents H2O2 and vanadate stimulate protein tyrosine phosphorylation inintact cells. J. Biol. Chem. 265, 2896–2902.
39. Trudel, S., Paquet, M. R., Grinstein, S. (1991) Mechanism of vanadate-induced activation of tyrosine phosphorylation and of the respiratory burst inHL60 cells. Role of reduced oxygen metabolites. Biochem. J. 276, 611–619.
40. Schreiber, E., Matthias, P., Muller, M., Schaffner, W. (1989) Rapiddetection of octamer binding proteins with ‘mini-extracts’, prepared froma small number of cells. Nucleic Acids Res. 17, 6419.
41. Rosen, R. L., Winestock, K. D., Chen, G., Liu, X., Hennighausen, L.,Finbloom, D. S. (1996) Granulocyte-macrophage colony-stimulating factorpreferentially activates the 94-kD STAT5A and an 80-kD STAT5A iso-form in human peripheral blood monocytes. Blood 88, 1206–1214.
42. David, M., Ford, D., Bertoglio, J., Maizel, A. L., Pierre, J. (2001) Inductionof the IL-13 receptor alpha2-chain by IL-4 and IL-13 in human keratin-ocytes: involvement of STAT6, ERK and p38 MAPK pathways. Oncogene20, 6660–6668.
43. Schindler, C., Darnell Jr., J. E. (1995) Transcriptional responses to polypep-tide ligands: the JAK-STAT pathway. Annu. Rev. Biochem. 64, 621–651.
44. Murata, T., Obiri, N. I., Debinski, W., Puri, R. K. (1997) Structure ofIL-13 receptor: analysis of subunit composition in cancer and immunecells. Biochem. Biophys. Res. Commun. 238, 90–94.
45. Debinski, W., Miner, R., Leland, P., Obiri, N. I., Puri, R. K. (1996) Receptorfor interleukin (IL) 13 does not interact with IL4 but receptor for IL4 interactswith IL13 on human glioma cells. J. Biol. Chem. 271, 22428–22433.
46. He, Y. W., Malek, T. R. (1995) The IL-2 receptor gamma c chain does notfunction as a subunit shared by the IL-4 and IL-13 receptors. Implicationfor the structure of the IL-4 receptor. J. Immunol. 155, 9–12.
47. Izuhara, K., Heike, T., Otsuka, T., Yamaoka, K., Mayumi, M., Imamura,T., Niho, Y., Harada, N. (1996) Signal transduction pathway of interleu-kin-4 and interleukin-13 in human B cells derived from X-linked severecombined immunodeficiency patients. J. Biol. Chem. 271, 619–622.
48. Welham, M. J., Learmonth, L., Bone, H., Schrader, J. W. (1995) Inter-leukin-13 signal transduction in lymphohemopoietic cells. Similaritiesand differences in signal transduction with interleukin-4 and insulin.J. Biol. Chem. 270, 12286–12296.
49. Miyazaki, T., Kawahara, A., Fujii, H., Nakagawa, Y., Minami, Y., Liu,Z. J., Oishi, I., Silvennoinen, O., Witthuhn, B. A., Ihle, J. N., et al. (1994)Functional activation of Jak1 and Jak3 by selective association with IL- 2receptor subunits. Science 266, 1045–1047.
50. Dawson, C. H., Brown, B. L., Dobson, P. R. (1997) A 70-kDa proteinfacilitates interleukin-4 signal transduction in the absence of the commongamma receptor chain. Biochem. Biophys. Res. Commun. 233, 279–282.
51. Reichel, M., Nelson, B. H., Greenberg, P. D., Rothman, P. B. (1997) TheIL-4 receptor alpha-chain cytoplasmic domain is sufficient for activationof JAK1 and STAT6 and the induction of IL-4-specific gene expression.J. Immunol. 158, 5860–5867.
52. Yin, T., Tsang, M. L., Yang, Y. C. (1994) JAK1 kinase forms complexeswith interleukin-4 receptor and 4PS/insulin receptor substrate-1-like pro-tein and is activated by interleukin-4 and interleukin-9 in T lymphocytes.J. Biol. Chem. 269, 26614–26617.
53. Palmer-Crocker, R. L., Hughes, C. C., Pober, J. S. (1996) IL-4 and IL-13activate the JAK2 tyrosine kinase and Stat6 in cultured human vascularendothelial cells through a common pathway that does not involve thegamma c chain. J. Clin. Investig. 98, 604–609.
54. Izuhara, K., Feldman, R. A., Greer, P., Harada, N. (1994) Interaction ofthe c-fes proto-oncogene product with the interleukin-4 receptor. J. Biol.Chem. 269, 18623–18629.
55. Keegan, A. D., Johnston, J. A., Tortolani, P. J., McReynolds, L. J., Kinzer,C., O’Shea, J. J., Paul, W. E. (1995) Similarities and differences in signaltransduction by interleukin 4 and interleukin 13: analysis of Janus kinaseactivation. Proc. Natl. Acad. Sci. USA 92, 7681–7685.
56. Rolling, C., Treton, D., Pellegrini, S., Galanaud, P., Richard, Y. (1996) IL4and IL13 receptors share the gamma c chain and activate STAT6, STAT3 andSTAT5 proteins in normal human B cells. FEBS Lett. 393, 53–56.
57. Darnell Jr., J. E., Kerr, I. M., Stark, G. R. (1994) Jak-STAT pathways andtranscriptional activation in response to IFNs and other extracellularsignaling proteins. Science 264, 1415–1421.
58. Doucet, C., Jasmin, C., Azzarone, B. (2000) Unusual interleukin-4 and -13signaling in human normal and tumor lung fibroblasts. Oncogene 19,5898–5905.
59. Zhang, X., Blenis, J., Li, H. C., Schindler, C., Chen-Kiang, S. (1995)Requirement of serine phosphorylation for formation of STAT-promotercomplexes. Science 267, 1990–1994.
60. Wen, Z., Darnell Jr., J. E. (1997) Mapping of Stat3 serine phosphorylation to asingle residue (727) and evidence that serine phosphorylation has no influence onDNA binding of Stat1 and Stat3. Nucleic Acids Res. 25, 2062–2067.
61. Chung, J., Uchida, E., Grammer, T. C., Blenis, J. (1997) STAT3 serinephosphorylation by ERK-dependent and -independent pathways negativelymodulates its tyrosine phosphorylation. Mol. Cell. Biol. 17, 6508–6516.
62. Beadling, C., Ng, J., Babbage, J. W., Cantrell, D. A. (1996) Interleukin-2activation of STAT5 requires the convergent action of tyrosine kinases anda serine/threonine kinase pathway distinct from the Raf1/ERK2 MAPkinase pathway. EMBO J. 15, 1902–1913.
63. Conrad, D. J., Lu, M. (2000) Regulation of human 12/15-lipoxygenase byStat6-dependent transcription. Am. J. Respir. Cell Mol. Biol. 22, 226–234.
64. Heydeck, D., Thomas, L., Schnurr, K., Trebus, F., Thierfelder, W. E., Ihle,J. N., Kuhn, H. (1998) Interleukin-4 and -13 induce upregulation of themurine macrophage 12/15-lipoxygenase activity: evidence for the involve-ment of transcription factor STAT6. Blood 92, 2503–2510.
65. Tanner, J. W., Chen, W., Young, R. L., Longmore, G. D., Shaw, A. S.(1995) The conserved box 1 motif of cytokine receptors is required forassociation with JAK kinases. J. Biol. Chem. 270, 6523–6530.
Roy et al. IL-13 signal transduction in human monocytes 589





























