Embed Size (px)
Citation preview

66 Journal of the Ncutvh)gwal 5cietJ~ e~, 122 t I ~)~4) Or) i~ 1994 Elsevier Science B.V. All rights reserved 0(122-5 IfiX/q4/S0v.(,~
JNS 4224
Immunohistochemical localization of glutathione peroxidase in infarcted human brain
S h u n y a T a k i z a w a a, K a z u s h i M a t s u s h i m a a, Y u k i t o S h i n o h a r a a , . , S a o r i O g a w a ~,
N o b u y u k i K o m a t s u b, H i r o t o s h i U t s u n o m i y a ~, K e i i c h i W a t a n a b e ~
Departments o f a Neurology, b Cell Biology and c Pathology, Tokai University School o f Medicine, Bohseidai, Isehara, Kanagawa, 259-11, Japan
(Received 18 May, 1993) (Revised, received 13 September, 1993)
(Accepted 26 September, 1993)
Key words: Immunohistochemical staining; Glutathione peroxidase; Cerebral infarction; Lipid peroxidation; Autopsied brain
Summary
This is the first report which demonstrates the presence of glutathione peroxidase in the autopsied brain of 5 patients without cerebral infarction and 21 patients with cerebral infarction by the indirect enzyme-labeled antibody technique with monoclonal antibody to human glutathione peroxidase. In 2 out of 5 patients without cerebral infarction, a weak reaction for glutathione peroxidase was demonstrated both in neurons and glia. In 6 patients who had died within 5 days after stroke, no staining was observed in infarcted brain tissue except in macrophages. In all 15 patients who had died more than 6 days after stroke, however, a reaction for glutathione peroxidase was demonstrated in the cytoplasm of glial cells in the marginal area around the infarction, and there was a patchy reaction in the cytoplasm of macrophages in the core lesion. These results suggest that glutathione peroxidase in glial cells of the marginal area around the infarction may play a protective role against lipid peroxidation after cerebral infarction, or alternatively, may be involved in the healing process after ischemia.
Introduction
It has been suggested that superoxide radicals and lipid peroxides are generated either during cerebral ischemia (Demopoulos 1973) or during reoxygenation after cerebral ischemia (Kogure et al. 1979, 1985; Yoshida et al. 1980). Furthermore, endogenous super- oxide dismutase (SOD) (Kontos and Wei 1986; Chan et al. 1987, 1988) and glutathione peroxidase (GSH-Px) (Prohaska and Ganther, 1976; Brannan et al. 1980; Carmagnol et al. 1983; Kaplan and Ansari 1984; Ansari et al. 1985) have been shown to play significant roles in reducing superoxides and various peroxides such as hydrogen peroxides and lipid peroxides, respectively, that occur in various brain lesions including ischemia. However, the anti-peroxidative function of GSH-Px in the brain is controversial. DeMarchena et al. (1974) reported that the GSH-Px activity in normal brains of various animals is insufficient to reduce lipid perox- ides. Chan et al. (1988) also reported that the activity of SOD was significantly decreased in a rat decapita-
* Corresponding author. Tel.: 0463-93-1121; Fax: 0463-94-8764.
tion model, but that of GSH-Px was not. It is, however. unclear whether biochemical measurement of the en- zyme activity using brain homogenate accurately repre- sents the activity in individual cells of the brain, espe- cially if the enzyme content is relatively small and its distribution is biased towards limited cell types. Fur- thermore, biochemical study may not enable us to distinguish the enzyme activity in cells from that in plasma. An immunohistochemical study is able to de- tect cellular GSH-Px selectively because of the im- munological difference between GSH-Px in cells and that in plasma (Maddipati and Marnett 1987), and should allow us to evaluate the regional distribution of GSH-Px in cerebral infarction.
Only one immunohistological study of GSH-Px in normal brain of experimental animals has been re- ported (Ushijima et al. 1986), and nothing is known about the distribution of GSH-Px in human brain with cerebral infarction. The purpose of this study was to investigate using immunohistochemical methods the specific localization of GSH-Px in autopsied human brain of patients with cerebral infarction, in order to cast light on the putative neuroprotective role of GSH- Px, in addition to SOD, against ischemic brain damage.
SSDI 0 0 2 2 - 5 1 0 X ( 9 3 ) E 0 2 3 5 - 2

Materials and methods
The subjects were 5 patients without cerebral infarc- tion and 21 autopsied patients with cerebral infarction, 13 male and 13 female, aged 39-92 years (mean + SD: 71 ___ 13 years). Table 1 shows the clinical characteris- tics of the patients we studied, including the location of the cerebral infarction, duration from onset to death, physical condition before death, and duration from death to autopsy (mean + SD: 7.35 + 5.30 h). Five pa- tients without cerebral infarction, patient Nos. 1-5 in Table 1, had suffered malignant thymoma (epithelial type), rectal carcinoma, lung carcinoma, acute myocar- dial infarction and malignant mesothelioma in pleura, respectively.
Preparation of anti-human glutathione peroxidase anti- body
The monoclonal anti-GSH-Px antibody was pro- duced in our laboratory and its specificity established (Suemizu et al. 1992). Briefly, human GSH-Px purified from human blood in Freund's incomplete adjuvant was injected subcutaneously into mice. Spleen ceils from the immunized mice were fused with NS-1 cells,
67
and finally the ELISA-positive hybridoma clones (GPX-347) were obtained. This monoclonal antibody reacted with a 21.5-22.5 kDa single polypeptide by immunoblot analysis, suggesting that this antibody was specific for human GSH-Px.
Immunohistochemical staining The indirect enzyme-labeled antibody technique with
anti-GSH-Px, prepared as described above, was ap- plied to determine the immunohistochemical localiza- tion of GSH-Px in infarcted human brain. The tissue blocks of autopsied brains were fixed in 10% formalde- hyde for 1 week and then embedded in paraffin. The blocks were sectioned at 4 mm thickness onto albumin-coated slides. After removal of the paraffin, the sections on the glass slides were incubated in a 5 mM solution of periodic acid for 10 min to oxidize pseudo-peroxidase compounds and exposed to 5% nor- mal goat serum for 10 min. The sections were first incubated with purified mouse anti-human GSH-Px antibody (GPX-347) at 50-fold dilution overnight in a humidified chamber at 4°C. After being washed thor- oughly with 0.01 M sodium phosphate-buffered saline (pH 7.2), the sections were incubated with anti-mouse
TABLE 1
CLINICAL CHARACTERISTICS OF PATIENTS
Patient Age Location of Days (from Physical condition before death Hours (from and sex cerebral infarction onset to death to
death) autopsy)
1 75M - 2 69M - 3 79M - 4 63F - 5 76F - 6 50M MCA 3 7 67M V-B 3 8 39F Bilateral ACA and MCA 4 9 50F ACA and MCA 4
10 90F MCA 4 11 70M MCA 5 12 66M MCA 6 13 67M V-B 9 14 66F MCA 9 15 77F ACA and MCA 9 16 56M MCA 10 17 84M ACA and MCA 13 18 81F ACA and MCA 16 19 70F V-B and MCA 26 20 72F MCA 31 21 77F V-B 33 22 92M MCA 36 23 74M V-B 73 24 84F Bilateral ACA and MCA 81 25 65M MCA 88 26 85F ACA and MCA 150
Cardiogenic shock 2 h before death 11 Septic shock 10 h before death 15 Hypoxia 1 day before death 15 Cardiogenic shock 5 h before death 6 Hypoxia 1 day before death 13 Hypoxia 1 day before death 2 Hypotension 1 day before death 6 Hypotension 10 h before death 18 Hypotension 2 h before death 3 Hypotension 2 h before death 5 Hypotension 15 h before death 2 Hypotension 9 h before death 5 Hypotension 6 h before death 6 Apnea at death 6 Apnea 3 h before death 3 Hypoxia 8 days before death 6 Cardiogenic shock 1 h before death 4 Hypoxia 1 h before death 12 Hypotension 9 h before death 3 exacerbation of paresis 2 days before death 1 Hypotension 14 h before death 1 Hypoxia 2 days before death 3 Hypotension 11 days before death 4 Hypotension 9 days before death 11 Decreased consciousness level 5 days before death 12 Hypoxia 10 days before death 18
ACA, anterior cerebral artery territory; MCA, middle cerebral artery territory; V-B, vertebro-basilar artery territory; - , a patient without cerebral infarction.

68
IgG-sheep IgG F(ab') 2 labeled with horseradish perox- idase (Amersham Japan, Tokyo) at 100-fold dilution for 3 h at room temperature in a humidified chamber. The sections were stained by the Graham-Karnovsky method with 3,3'-diaminobenzidine and hydrogen per- oxide (Graham and Karnovsky 1966). As an immuno- logic negative control, BrdU (Becton Dickinson, San Jose, CA, USA) at 50-fold dilution was employed in place of the anti-human GSH-Px antibody. The follow- ing grading scale for the evaluation of GSH-Px staining was used: 0 = no staining, 1 = weak staining (including patchy staining), 2 = strong staining. The pathological evaluation was carried out with the pathologist blinded as to patients' clinical status.
Neuron specific enolase (rabbit anti-human neuron specific enolase, Dako, Denmark), S-100 protein (rab- bit anti-cow S-100, Dako, Denmark) and lysozyme (rabbit anti-human lysozyme, Dako, Denmark), which are reliable histochemical markers of neurons, glial cells and macrophages, respectively, were stained for the purpose of identifying the cells stained with GSH- Px. For these immunohistochemical stainings, anti-rab- bit IgG F(ab') 2 (from donkey) labeled with horseradish peroxidase (Amersham Japan, Tokyo) was applied. Fi- nally, nuclear counter-staining with methyl green was performed on all immunohistochemically stained sec- tions mentioned above.
Results
Table 2 summarizes the immunohistochemical reac- tion for GSH-Px in the non-infarcted region, and marginal and core areas in the infarcted region in all patients examined. No reaction for BrdU as an im- munologic negative control was observed in any of the tissue sections studied.
In 2 out of 5 patients without cerebral infarction, a weak reaction for GSH-Px both in neuron and glia was demonstrated as shown in Fig. 1 (patient No. 5). A weak reaction for GSH-Px was observed in only glia in 1 patient without infarction, and no reaction for GSH- Px was detected in 2 patients without infarction. There was no relationship between GSH-Px staining and physical condition before death in 5 patients without cerebral infarction.
In the non-infarcted region in 21 patients with cere- bral infarction, a weak reaction for GSH-Px was simi- larly observed in neurons and glia in 1 patient, and in only neurons in 3 patients, whose tissues showed no significant pathologic changes in hematoxylin-eosin- stained sections.
In 6 patients who had died within 5 days after stroke, no staining of GSH-Px was clearly observed in neurons or glial cells in the infarcted region, except a patchy staining in macrophages in the core area of the infarction in 2 patients (Nos. 10 and 11).
' FABLE 2
I M M U N O H I S T O C H E M I C A L R E A C T I O N B R A I N
F O R GSH-Px 1N
No. Days Non- in fa rc t ed In fa rc ted region
(from onse t reg ion Marg ina l a rea Core a rea of infarc t N G M to dea th ) N G M N G M
1 - 1 1 0 . . . . .
2 - 0 0 0 - -
3 - 0 0 0
4 - 0 1 0
5 - 1 1 0 - -
6 3 0 0 0 0 0 0 0 0 0
7 3 1 0 0 0 0 0 0 0 0 8 4 0 0 0 0 0 0 0 0 0
9 4 0 0 0 0 0 0 0 0 0
10 4 0 0 0 0 0 0 0 0 1
I I 5 0 0 0 0 0 0 0 0 1
12 6 0 0 0 0 2 1 0 1 1 13 9 0 0 0 0 2 1 0 1 1
14 9 0 0 0 0 2 1 0 1 1
15 9 0 0 0 0 1 1 0 0 1
16 10 0 0 0 0 2 0 0 0 1 17 13 1 1 0 1 1 0 0 0 1 18 16 1 0 0 1 2 0 0 1 1
19 26 0 0 0 0 1 0 0 1 l 20 31 1 0 0 1 2 l 0 0 1
21 33 0 0 0 0 2 1 0 l 1 22 36 0 0 0 0 2 1 0 1 1
23 73 0 0 0 0 1 0 0 0 1
24 81 0 0 0 0 1 0 0 0 1
25 88 0 0 0 0 1 1 0 0 1
26 150 0 0 0 0 1 0 0 0 1
N, neuron ; G, gl ial cell; M, mac rophage . The g rad ing scale for
GSH-Px s ta in ing was as follows: 0 = no s ta in ing , 1 = weak s ta in ing ( inc lud ing pa tchy staining) , 2 = s t rong staining.
In all 15 patients who had died more than 6 days after stroke an immunohistochemical reaction for GSH-Px was observed in cells scattered in the marginal area around the cerebral infarction, and an especially strong reaction was seen there in 8 out of the 15 patients. Patchy staining of GSH-Px was also observed in the core area of the infarction in all 15 patients. Fig. 2 shows representative GSH-Px- and hematoxylin- eosin-stained sections at low magnification in patient No. 18. Hematoxylin-eosin staining (Fig, 2A) shows pyknotic nuclei in neurons and glial proliferation in the marginal area of the infarct, and foamy cells in the core of the infarct. A strong reaction for GSH-Px is seen in the cells surrounding the cerebral infarction (Fig. 2B).
In order to identify the GSH-Px positive cells, fur- ther immunohistochemical staining of serial sections was performed. In the non-infarcted region (Fig. 3A, B), a weak reaction for GSH-Px was observed, involv- ing cells which were identified as neurons by the posi- tive reaction for neuron specific enolase. In the

Fig. 1. Immunohistochemical staining of GSH-Px (A, 480x) in a patient without cerebral infarction (patient No. 5), who had died 1 day after hypoxia. A weak reaction for glutathione peroxidase was observed, in cells which are identified as neuron (small arrows) and glia (large arrows) by the positive reaction for neuron specific eno- lase (B, a consecutive section to A, 480x) and S-100 protein (C, a
consecutive section to A, 480 x ), respectively.
69
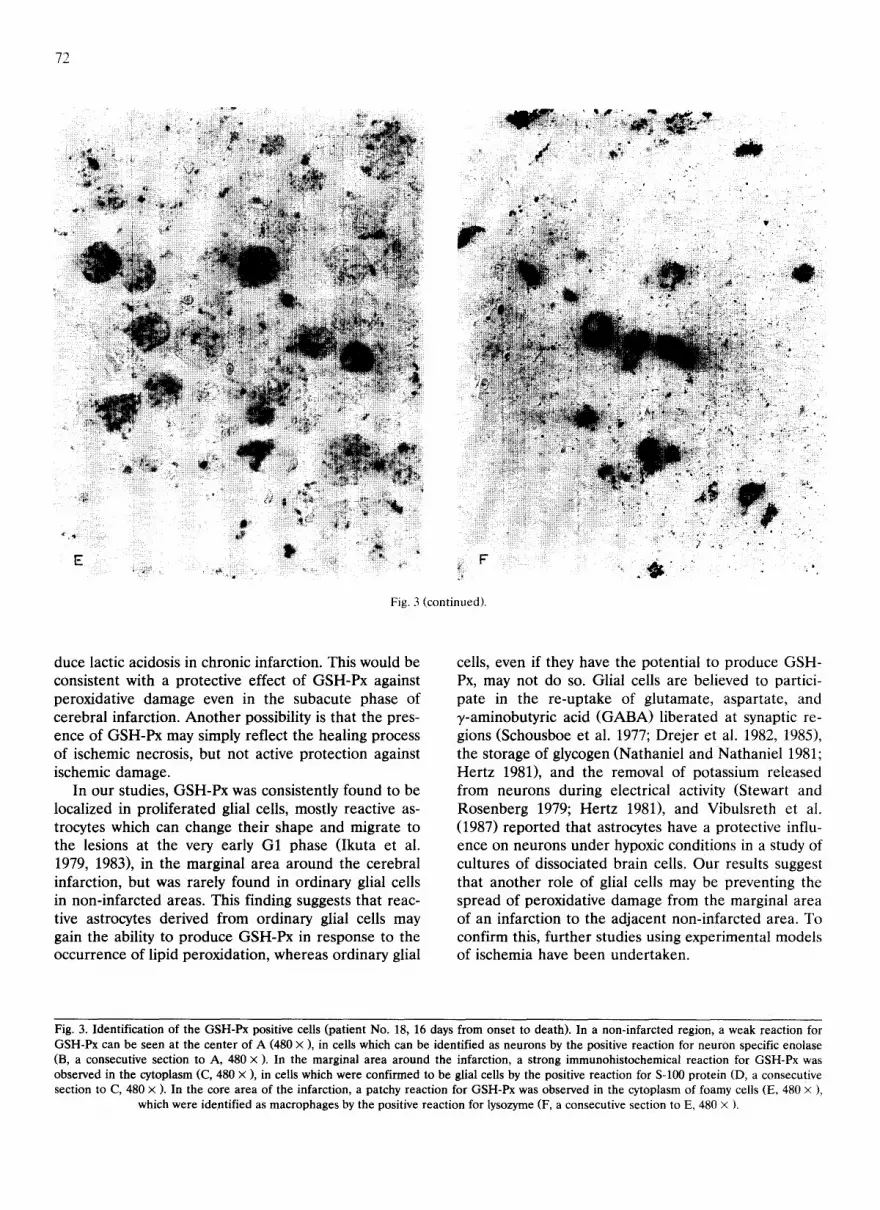
marginal area around the cerebral infarction (Fig. 3C, D), a strong immunohistochemical reaction for GSH-Px in the cytoplasm was observed in cells which were identified as glial cells by the positive reaction for S-100 protein. In the core area of the infarction (Fig. 3E, F), a patchy reaction for GSH-Px was observed in the cytoplasm of foamy cells, which were identified as macrophages by the positive reaction for lysozyme.
There was no relationship between GSH-Px staining and either age of the patients, location of the infarct, or hours from death to autopsy.
Discussion
The major finding of this study was the observation of immunohistochemical staining for GSH-Px in the cytoplasm of glial cells in the marginal area around the cerebral infarction, together with patchy staining in cytoplasm of macrophages in the core area of the infarction in patients who had died more than 6 days after the onset of occlusive cerebrovascular disease. In contrast, a few neurons and glia showed a weak reac- tion for GSH-Px in non-infarcted regions. This is the first report to demonstrate the presence of GSH-Px in human brain and has confirmed the cell types of the GSH-Px positive cells.
Our immunohistochemical examination suggests that the content of GSH-Px in neuron and glia in the non-infarcted region in patients with infarction and in brain tissue without cerebral infarction is rather small, although immunoreactivity of the enzyme protein would be partially lost during the period between death and autopsy, and during the tissue processing for immuno- histochemistry, including the fixation and paraffin em- bedding. On the other hand, the results of biochemical examination have been variable. Fukuzawa and Toku- mura (1976) could find no GSH-Px activity in the brain tissues of normal mice. DeMarchena et al. (1974) spec- ulated that GSH-Px activity of the cytosol fractions from the brains of various animals is insufficient to reduce lipid hydroperoxides. On the contrary, Pro- haska and Ganther (1976) found that GSH-Px activity was 3 times higher than that reported by DeMarchena et al. (1974), and several other investigators have found sufficient GSH-Px activity in the brain to reduce lipid hydroperoxides (Brannan et al. 1980; Carmagnol et al. 1983; Kaplan and Ansari 1984). The reason for the inconsistency seems to be that biochemical studies are not appropriate to evaluate the amount or activity of an enzyme whose content is rather small because the GSH-Px positive cells are sparsely and inhomogenously distributed in the brain as our immunohistochemical observations proved.
As regards intracellular localization of GSH-Px, our data suggest that GSH-Px is located in the cytoplasm

70
of neurons and glia in non-infarcted brain. Ushijima et al. (1986), however, reported the immunohistochemical detection of GSH-Px in the nuclei of nerve cells in normal rat brain. These contradictory results may be due to the differences of species and /o r methods employed. In our preliminary experiments with is- chemic rat brain (reported at the 13th Congress of the Japanese Stroke Association, 1988, Tokyo), however, the positive reaction for GSH-Px was clearly and exten- sively localized in the cytoplasm of glial cells when the rapid freeze substitution technique was employed to ensure strict immobilization of the antigen, which is a very soluble cytosolic protein, and to minimize loss of antigenicity of the enzyme. When we applied the ordi- nary perfusion fixation technique with paraformalde- hyde, as used by Ushijima et al. (1986), immobilization of the antigen was poor, and artifactual entry of the cytosolic antigen into the nuclei was observed. Other investigators have reported that GSH-Px activity in the brain tissues was highest in the cytosol followed by mitochondrial and synaptosomal fractions (Prohaska and Ganther 1976; Victorica et al. 1984), in agreement with our results.
It is of interest that a strong staining ti)r GSH-Px was found in the marginal area around the infarction, which seems to be the critical zone of ischemia, in patients who had died more than 6 days after strokc. GSH-Px synthesis is probably induced in the marginal area around the infarction, where lipid peroxidation takes place actively owing to the restoration of the oxygen supply by collateral circulation (Kogure et al. 1979, 1985; Yoshida et al. 1980). Evidence that GSH-Px synthesis is enhanced by lipid peroxide in peritoneal macrophages (Watanabe and Murakoshi 1986) and liver cells (Watanabe 1986) may support the above specula- tion.
There are some possible explanations for the de- layed appearance of GSH-Px (more than 6 days after stroke) in the human brain. One possibility is the gradual synthesis of GSH-Px in response to the accu- mulation of lipid peroxides after ischemia. Hugg et al. (1992) reported that patients with chronic cerebral infarction had elevated lactate within the infarct, If lactate is produced by viable, yet persistently ischemic tissue, then ischemia, which stimulates the anaerobic production of lactic acid, would be expected to pro-
Fig. 2. Hematoxylin-eosin staining (A, 240 × ) and immunohistochemical staining for GSH-Px (B, 240 × ) in a patient with cerebral infarction (No. 18, 16 days from onset to death). (A) Neurons with pyknotic nuclei and proliferated glial cells with eosinophilic cytoplasm in the marginal area of the infarction, and foamy cells in the core of the infarct were stained with hematoxylin-eosin. (B) Strong immunohistochemical reaction
for GSH-Px was observed in the cells surrounding the infarction.

.-..I
72
F"
Fig. 3 (continued).
i ' :
L - d ~" i ii ~ ~ • i!iiiiiii!%i diiii!!~!ii; !iiiiiiiill ¸: ~ ! ....... ' ~ i'
duce lactic acidosis in chronic infarction. This would be consistent with a protect ive effect of GSH-Px against peroxidative damage even in the subacute phase of cerebral infarction. A n o t h e r possibility is that the pres- ence o f GSH-Px may simply reflect the healing process of ischemic necrosis, but not active protect ion against ischemic damage.
In our studies, GSH-Px was consistently found to be localized in prol i ferated glial cells, mostly reactive as- trocytes which can change their shape and migrate to the lesions at the very early G1 phase ( Ikuta et al. 1979, 1983), in the marginal area a round the cerebral infarction, but was rarely found in ordinary glial cells in non- infarc ted areas. This finding suggests that reac- tive astrocytes derived f rom ordinary glial cells may gain the ability to p roduce GSH-Px in response to the occurrence o f lipid peroxidation, whereas ordinary glial
cells, even if they have the potential to p roduce GSH- Px, may not do so. Glial cells are believed to partici- pate in the re-uptake of glutamate, aspartate, and ~,-aminobutyric acid ( G A B A ) l iberated at synaptic re- gions (Schousboe et al. 1977; Drejer et ali 1982, 1985), the storage of glycogen (Nathaniel and Nathaniel 1981; Her tz 1981), and the removal of potassium released f rom neurons during electrical activity (Stewart and Rosenberg 1979; Her tz 1981), and Vibulsreth et al. (1987) repor ted that astrocytes have a protective influ- ence on neurons under hypoxic conditions in a study of cultures of dissociated brain ceils. Our results suggest that another role of glial cells may be preventing the spread of peroxidative damage from the marginal area of an infarction to the adjacent non- infarc ted area. To confirm this, fur ther studies using experimental models of ischemia have been under taken.
Fig. 3. Identification of the GSH-Px positive cells (patient No. 18, 16 days from onset to death). In a non-infarcted region, a weak reaction for GSH-Px can be seen at the center of A (480 x ), in cells which can be identified as neurons by the positive reaction for neuron specific enolase (B, a consecutive section to A, 480 x ). In the marginal area around the infarction, a strong immunohistochemical reaction for GSH-Px was observed in the cytoplasm (C, 480 × ), in cells which were confirmed to be glial cells by the positive reaction for S-100 protein (D, a consecutive section to C, 480 x ). In the core area of the infarction, a patchy reaction for GSH-Px was observed in the cytoplasm of foamy cells (E, 480 x ),
which were identified as macrophages by the positive reaction for lysozyme (F, a consecutive section to E, 480 x ).

Acknowledgments This work was supported in part by grants from the Tokai University Research Foundation.
References
Ansari, K.A., D. Bigelow and E. Kaplan (1985) Glutathione peroxi- dase activity in surgical and autopsied human brains. Neurochem. Res., 10: 703-711.
Brannan, T.S., H.S. Maker, C. Weiss and G. Cohen (1980) Regional distribution of glutathione peroxidase in adult rat brain. J. Neu- rochem., 35: 1013-1014.
Carmagnol, F., P.M. Sinet and H. Jerome (1983) Selenium-depen- dent and non-selenium-dependent glutatbione peroxidases in hu- man tissue extracts. Biochim. Biophys. Acta, 759: 49-57.
Chan, P.H., S. Longar and R.A. Fishman (1987) Protective effects of liposome-entrapped superoxide dismutase on posttraumatic brain edema. Ann. Neurol., 21: 540-547.
Chan, P.H., R. Chu and R.A. Fishman (1988) Reduction of activities of superoxide dismutase but not of glutathione peroxidase in rat brain regions following decapitation ischemia. Brain Res., 439: 388-390.
DeMarchena, O., M. Guarnieri and G. McKhann (1974) Glutathione peroxidase levels in brain. J. Neurochem., 22: 773-776.
Demopoulos, H.B. (1973) The basis of free radical pathology. Fed. Proc., 32: 1859-1861.
Drejer, J., P.M. Larsson and A. Schousboe (1982) Characterization of L-glutamate uptake into and release from astrocytes and neu- rons cultured from different brain regions. Exp. Brain Res., 47: 259-269.
Drejer, J., H. Benveniste, N.H. Diemer and A. Schousboe (1985) Cellular origin of ischemia-induced glutamate release from brain tissue in vivo and in vitro. J. Neurochem., 45: 145-151.
Fukuzawa, K. and M. Tokumura (1976) Glutathione peroxidase activity in tissues of vitamin E-dependent mice. J. Nutr. Sci. Vitaminol., 22: 405-407.
Graham, R.C. and M.J. Karnovsky (1966) The early stages of absorp- tion of injected horseradish peroxidase in the proximal tubules of mouse kidney: Ultrastructual cytochemistry by a new technique. J. Histochem. Cytochem., 14: 291-302.
Hertz, L. (1981) Functional interactions between astrocytes and neurons. In: Federroff, S. (Ed.), Glial and Neuronal Cell Biology, A.R. Liss, New York, pp. 45-58.
Hugg, J.W., J.H. Duijin, G.B. Matson, A.M; Maudsley, J.S. Tsuruda, D.F. Gelinas and M.W. Weiner (1992) Elevated lactate and alkalosis in chronic human brain infarction observed by 1H and 31p MR spectroscopic imaging. J. Cereb. Blood Flow Metab., 12: 734-744.
Ikuta, F., E. Ohama, K. Yamazaki, S. Takeda, S. Egawa and T. Ichikawa (1979) Morphology of migrating glial cells in normal development, neoplasia and other disorders. In: Zimmerman,
73
H.M. (Ed.), Progress in Neuropathology, Vol. 4, Raven Press, New York, pp. 377-405.
Ikuta, F., Y. Yoshida, E. Ohama, K. Oyanagi, S. Takeda, K. Ya- mazaki and K. Watanabe (1983) Revised pathophysiology on BBB damage: The edema as an ingeniously provided condition for cell motility and lesion repair. Acta Neuropathol. (Berl.), Suppl. 8: 103-110.
Kaplan, E. and K. Ansari (1984) Reduction of polyunsaturated fatty acid hydroperoxides by human brain glutathione peroxidase. Lipids, 19: 784-789.
Kogure, K., H. Morooka, R. Busto and P. Scheinberg (1979) Involve- ment of lipid peroxidation in postischemic brain damage. Neurol- ogy, 29: 546.
Kogure K., H. Arai, K. Abe and M. Nakano (1985) Free radical damage of the brain following ischemia. Prog. Brain Res., 63: 237-259.
Kontos H.A. and E.P. Wei (1986) Superoxide production in experi- mental brain injury. J. Neurosurg., 64: 803-807.
Maddipati, K.R. and L.J. Marnett (1987) Characterization of the major hydroperoxide-reducing activity of human plasma. J. Biol. Chem., 262: 17398-17403.
Nathaniel, E.J.H. and D.R. Nathaniel (1981) The reactive astrocyte. Adv. Cell Neurobiol., 2: 249-301.
Prohaska, J.R. and H.E. Ganther (1976) Selenium and glutathione peroxidase in developing rat brain. J. Neurochem., 27: 1379-1387.
Schousboe, A., G. Svenneby and L. Hertz (1977) Uptake and metabolism of glutamate in astrocytes cultured from dissociated mouse brain hemispheres. J. Neurochem,, 29: 999-1005.
Stewart, R.M. and R,N. Rosenberg (1979) Physiology of glia: glial- neural interactions. Int. Rev. Neurobiol., 21: 275-309.
Suemizu, H., S. Yoshimura, N. Tada, K. Watanabe and T. Moriuchi (1992) Production and characterization of two monoclonal anti- bodies to human glutathione peroxidase. Hybridoma, 11: 795-801.
Ushijima, K., H. Miyazaki and T. Morioka (1986) Immunohistochem- ical localization of glutathione peroxidase in the brain of the rat. Resuscitation, 13: 97-105.
Vibulsreth, S., F. Hefti, M.D. Ginsberg, W.D. Dietrich and R. Busto (1987) Astrocytes protect cultured neurons from degeneration induced by anoxia. Brain Res., 422: 303-311.
Victorica, J., A. Machado and J. Satrustegui (1984) Age-dependent variations in peroxide-utilizing enzymes from rat brain mitochon- dria and cytoplasm. J. Neurochem., 42: 351-356.
Watanabe, K. (1986) Lipid peroxidation and cell injury; the role of glutathione peroxidase. Trans. Soc. Pathol. Jpn., 76: 39-74.
Watanabe, K. and M. Murakoshi (1986) Lipid peroxidation and arachidonate cascade in macrophages with a special reference to the change of glutathione peroxidase, lipid peroxidase scavenger. Tokai J. Exp. Clin. Med., 11: 105-109.
Yoshida, S., S. Inoh, T. Asano, K. Sano, M. Kubota, H. Shimazaki and N. Ueta (1980) Effect of transient ischemia on free fatty acids and phospholipids in the gerbil brain: Lipid peroxidation as a possible cause of postischemic injury. J. Neurosurg., 53: 323-331.





























